
Plant-Growth-Promoting Rhizobacteria Emerging as an Effective Bioinoculant to Improve the Growth, Production, and Stress Tolerance of Vegetable Crops

 ,
,  ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Effect of PGPR in Plant Growth Promotion
3. Role of PGPR in Vegetable Crop Production

{kind=link}
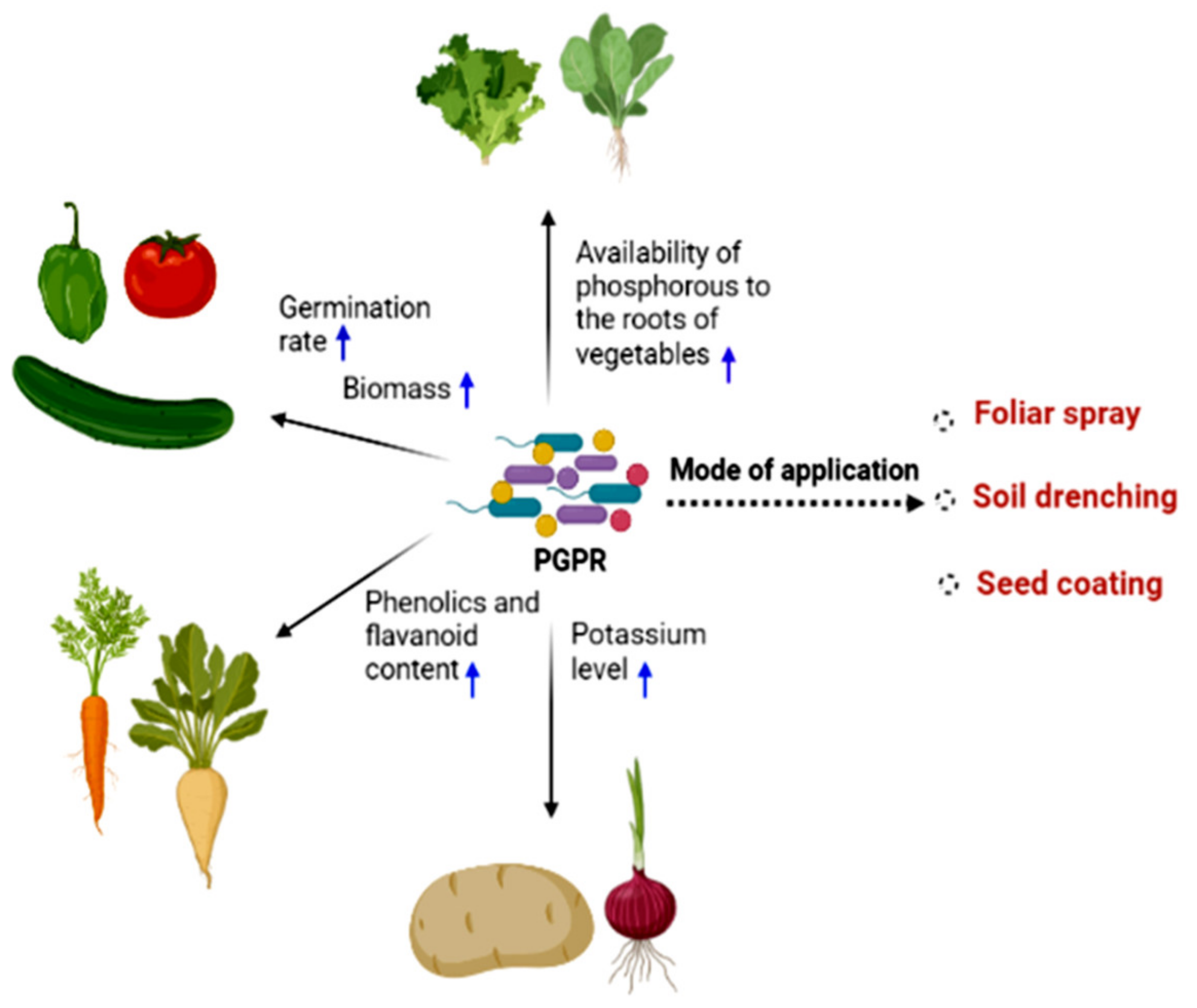{kind=link}
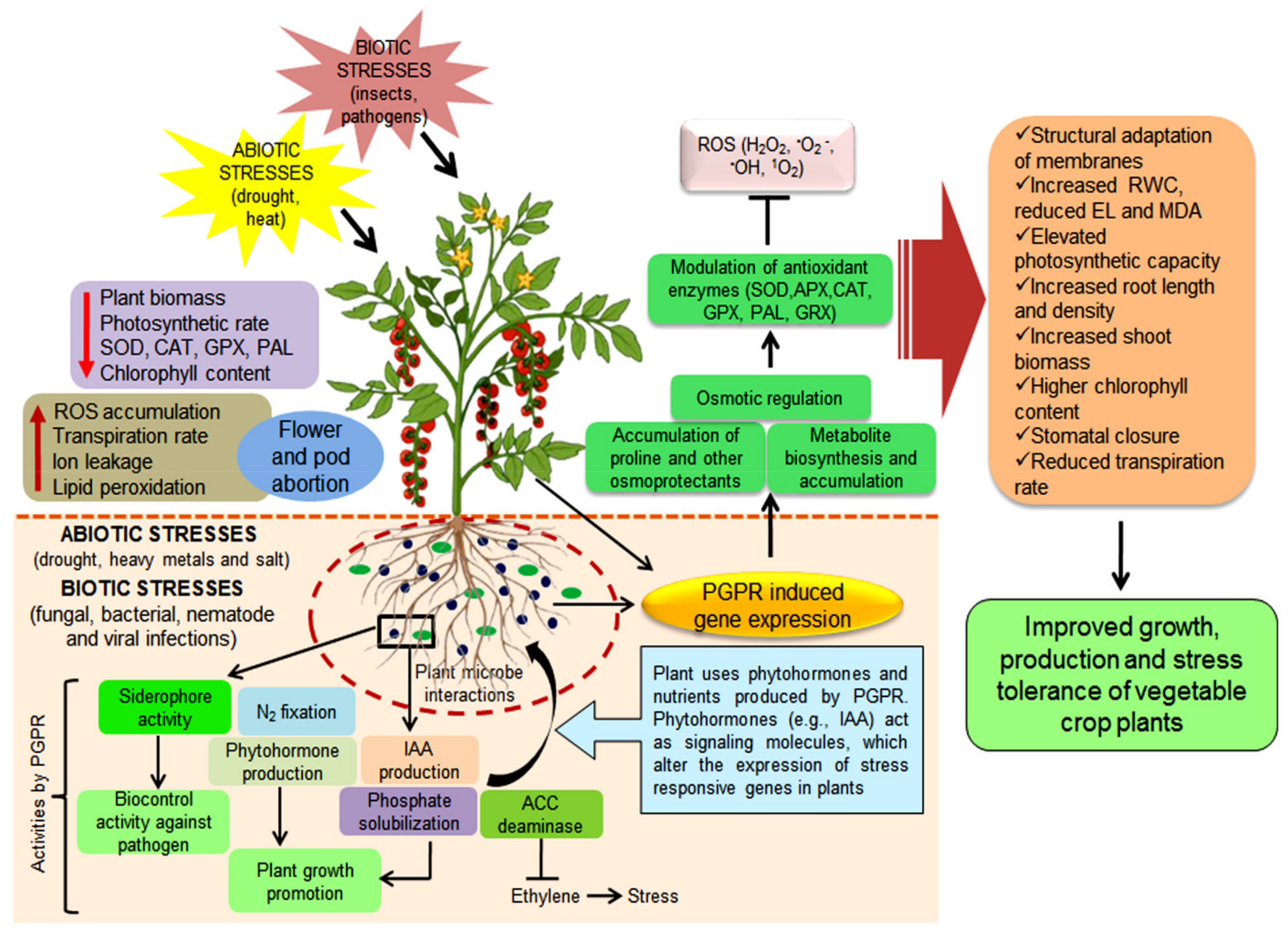{kind=link}
| PGPR | Vegetable Crop | Mode of Treatment | Effect on Crops | References |
|---|---|---|---|---|
| Alcaligenes faecalis and Bacillus amyloliquefaciens | Spinacia oleracea | Soil treatment | Mitigated lead toxicity | [95] |
| B. pumilus SE34 | Solanum lycopersicum | Seed treatment | Induced systemic response during infection | [96] |
| Jeotgalicoccus huakuii NBRI 13E | S. lycopersicum, Abelmoschus esculentus, Zea mays | Seed treatment and foliar spray | Increased yield and ameliorated salt stress | [97] |
| B. pumilus strain SE34 or B. amyloliquefaciens strain IN937a or B. subtilus strain IN937 | S. lycopersicum | Seed treatment and soil drenching | Induced resistance against CMV virus | [98] |
| Rhizobium spp. | S. lycopersicum, Capsicum annuum, Daucus carota, Lactuca sativa | Seed treatment | Increased biomass | [99,100] |
| Bacillus megaterium var. phosphaticum | S. oleracea | Soil and seed treatment | Ensured efficient absorption of P, water, and other microelements to alleviate water stress and resist fungal diseases | [101,102] |
| Bacillus amyloliquefaciens | L. esculentum | Spraying on leaves | Induced systemic resistance against tomato leaf curl virus disease | [103] |
| Bacillus cereus | S. lycopersicum | Soil drenching | Biotic stress resistance against bacterial speck disease caused by Pseudomonas syringae pv | [104] |
| Paenibacillus alvei and Bacillus velezensis | Sorghum bicolor | Seed treatment | Resistance to water stress and crown rot disease caused by Fusarium pseudograminearum | [105] |
| Pseudomonas fluorescens | Arachis hypogea | Seed treatment | Produced 1-aminocyclopropane-1-carboxylic acid (ACC) deaminase to confer resilience against salinity stress | [106] |
| PGPR Bacillus subtilis (RS2) and Bacillus spp. (RS7) | C. annuum | Seedling treatment | Increased productivity | [107] |
| Bacillus tequilensis | S. lycopersicum | Seedling and soil drenching | Produced ACC deaminase to confer resilience against salinity stress | [108] |
| Stenotrophomonas maltophilia, Achromobacter xylosoxidans, Achromobacter spp. | S. tuberosum | Potato tuber coating | Increased P solubilization, indole acetic acid, hydrogencyanide, and ammonia | [109] |
| Pseudomonas spp. PS1 | Vigna radiate | Seed treatment | Increased plant biomass, yield, and protein content | [110] |
| B. amyloliquefaciens | S. lycopersicum | Seed treatment | Resistance from bacterial wilt of tomato (Ralstonia solanacearum) | [111] |
| Bacillus cereus BC1AW and Pseudomonas putida PP3WT | S. lycopersicum | Seedling treatment | Ameliorated bacterial wilt disease | [112] |
| Pseudomonas fluorescens | Solanum tuberosum | Soil treatment | Protection from Ralstonia solanacearum pathogen. Reduced bacterial wilt incidence and improved growth | [113] |
| Trichoderma viride ES1 and Pseudomonas fluorescens Bak150 | S. tuberosum | Foliar spray | Suppressed early blight disease and increased yield | [114] |
| Trichoderma spp. | Brassica oleracea | - | [115] | |
| Trichoderma spp. | S. lycopersicum | Seed priming and soil treatment | Protection from F. oxysporum f. sp. lycopersici | [116] |
| T. harzianum+Pseudomonas spp. | S. lycopersicum | - | Protection from Sclerotium rolfsii | [117] |
| T. viride+T. harzianum+P. fluorescens+Azotobacter spp. + Azospirillum spp. + PSB | S. lycopersicum | Seed treatment and soil drenching | Disease management and protection from Pythium aphanidermatum, Ralstonia solanacearum, Fusarium oxysporum f. sp. lycopersici | [118] |
| Bacillus subtilis, Trichoderma spp. | S. lycopersicum, S. melongena | Seed treatment | Protection from Fusarium infection through secretion of extracellular cell-wall-degrading enzymes | [119,120] |
| Pseudomonas fluorescens | A. sesculentus | Seed and soil treatment | Protection from Rhizoctonia solani by the producing siderophores, HCN, and indole acetic acid | [121] |
| Lactic acid bacteria | C. annuum | Soil drenching and foliar spray | Protection from black rot by producing siderophores | [122] |
| Azospirillum brasilense, Pseudomonas fluorescens and Bacillus megaterium | Cucumis sativus | Seedling treatment and foliar spray | Improved fruit quality | [123] |
| Pseudomonas fluorescens, Pseudomonas spp., Bacillus subtilis | C. sativus | Seed treatment | Protection from damping off by producing antibiotics and metabolites and inducing systemic resistance | [124] |
| Chaetomium globosum, Burkholderia cepacia | S. tuberosum, C. annuum | Soil drenching and foliar spray | Protection from late blight disease by producing endo- and exo-glucanases; antimicrobial activity of organic acids | [125,126] |
| Trichoderma harzianum+Pseudomonas fluorescens | S. tuberosum | Seed treatment and foliar spray | Protection from early blight caused by Alternaria solani but active biomolecules not yet determined | [127] |
| Bacillus subtilis | C. sativus | Soilless potting mix drenching | Disease suppression against anthracnose disease | [128] |
| Stenotrophomonas maltophilia and Agrobacterium fabrum | Momordica charantia | Seed coating | Immobilized Cd in Cd-rich soil to improve growth | [95] |
| Bacillus velezensis isolates (Y6 and F7) | S. lycopersicum | Soil and seed treatment | Protection from fungal infections by producing antibiotic compounds | [129] |
4. Mechanistic Overview of PGPR-Mediated Plant Growth Promotion of Vegetable Crops under Stress Conditions
4.1. Role of PGPR against Biotic Stresses in Vegetable Crops
4.1.1. Role of PGPR in Fungal- and Bacterial-Induced Stress in Vegetable Crops
4.1.2. PGPR against Nematode and Insect Pests
4.2. Role of PGPR against Abiotic Stress in Vegetable Crops
| Stress | Crops | PGPR Isolates | PGP Activity | References |
|---|---|---|---|---|
| Abiotic stress | ||||
| Salinity | Abelmoschus esculentus | Enterobacter spp. | Increased ACC deaminase activity | [148] |
| Salinity | Lycopersicum esculentum | Streptomyces spp. strain PGPA39 | Increased ACC deaminase activity, phosphate solubilization, and IAA production | [149] |
| Drought | L. esculentum | Bacillus subtilis | Cytokinin signaling | [150] |
| Drought | Capsicum annuum | Bacillus licheniformis K11 | Reduced ethylene concentration | [151] |
| Salinity and drought | Cucumis sativus | Burkholderia cepacia, Promicromonospora spp. | Increased salicylic acid and gibberellic acid | [152] |
| Salinity | Solanum melongena | Pseudomonas spp. | Produced antioxidant enzymes | [153] |
| Salinity | Pisum sativum | Bacillus spp. | Increased IAA production, phosphate solubilization, ammonia production, ACC deaminase activity, siderophore production, and antioxidant enzyme production | [154] |
| Salinity | Mentha spp. | Halomonas desiderata STR8 and Exiguobacterium oxidotolerans STR36 | Reduced harmful effects of salinity | [155,156] |
| Salinity | M. polymorpha, Medicago lupulina, Medicago truncatula, Medicago sativa | Bacillus megaterium NMp082 | Induced tolerance to salt stress | [157] |
| Heat | Solanum lycopersicum | Bacillus cereus | Extended thermotolerance in tomato seedlings | [158] |
| Biotic stress | ||||
| Damping off | L. esculentum | Streptomyces isolate DBTB 13, Trichoderma viride, T. harzianum, and P. fluorescens + Azotobacter and Azospirillum | Reduced stunting and stem collapse in infected plants | [118,159,160] |
| Bottom rot | Lactuca sativa | Bacillus amyloliquefaciens strain FZB42 | Improved the quality of lettuce by preventing wilting and rotting | [161] |
| Powdery mildew | C. sativus | Ampelomyces quisqualis Ces., B. subtilis strain GB03 | Prevented crop from tiny white superficial spots, reduced severity of angular leaf spot disease (foliar disease) | [162] |
| White rust disease, Fusarium wilts | Spinacia oleracea | B. subtilis, Pseudomonas spp., Bacillus spp., Burkholderia spp., Penicillium oxalicum, Enterobacter cloacae, Trichoderma spp. | Controlled Fusarium wilt and white rust | [78,163] |
| Colletotrichum lindemuthianum | Phaseolus vulgaris | P. fluorescens | Disease management against biotic stress | [164] |
| Damping-off | Beta vulgaris | Pseudomonas fluorescens | Disease management by producing antifungal compounds | [165] |
| Plasmodiophora brassicae | Brassicae oleraceae | Trichoderma spp. | Prevented and managed club root disease in cabbage | [115] |
| Pythium aphanidermatum, Ralstonia solanacearum, Fusarium oxysporum f. sp. lycopersici | L. esculentum | T. viride+T. harzianum+P. fluorescens+Azotobacter+Azospirillum + PSB | Disease management from several biotic stress | [118] |
| Powdery mildew, Botrytis rot | Greenhouse crops | Ampelomyces quisqualis, Pseudomonas flocculosa, Ulocladium spp. | Disease control against Botrytis rot and powdery mildew | [166] |
| Fusarium wilt, bacterial wilt | S. melongena and L. esculentum | Trichoderma spp., Bacillus subtilis, Bacillus amyloliquefaciens, Pseudomonas fluorescens | Produced antibiotics and secondary metabolites to control bacterial wilt and fusarium diseases through the secretion of enzymes that degrade extracellular wall components | [119,120,167] |
| Root rot disease | Abelmoschus esculentus | Pseudomonas fluorescens | Disease management by producing siderophores, HCN, and indole acetic acid | [121] |
| Damping off, downy mildew | Cucumis sativus | Pseudomonas spp., Bacillus subtilis, consortium of Achromobacter spp., Streptomyces spp., Bacillus licheniformis | Disease management by producing numerous antibiotics, metabolites, and induced systemic resistance | [124] |
| Bacterial spot and blight disease | C. annuum | Lactic acid bacteria, P. fluorescens | Protection by producing siderophores, numerous chemicals, and microbial fungicides | [122,168] |
| Late blight | S. tuberosum | Burkholderia cepacia; Chaetomium globosum | Protection by generating antimicrobial activity through organic acids and enzymes, such as exo- and endo-glucanases | [125,126] |
| Pythium aphanidermatum | L. esculentum Mill. | Streptomyces isolate H2 | Prevented damping off, thus acting as a biocontrol agent | [160] |
| Squash mosaic virus | C. sativus | P.fluorescens, B. polymyxa | Protection from pathogenic viruses | [169] |
| Watermelon mosaic potyvirus | C. maxima | B. subtilis, B. pumilus | Biocontrol mechanism for pathogenic viruses | [170] |
| Bacterial wilt, Fusarium wilt, leaf spot, anthracnose, Alternaria leaf blight, downy and powdery mildew | Citrullus lanatus (Thunb.) | P. polymyxa (SN-22), Sinomonas atrocyanea (NSB27) | Reduced angular leaf spot lesions and gummy stem blight lesions and inhibited bacterial fruit blotch | [156] |
| Fusarium wilt | Raphanus sativus | Pseudomonas putida strains WCS358 and RE8 | Provided biocontrol mechanism against biotic agent | [156] |
4.2.1. PGPR-Mediated Drought Tolerance in Vegetable Crops
4.2.2. PGPR-Mediated Salinity Tolerance in Vegetable Crops
4.2.3. PGPR-Mediated Tolerance to Heat, Metal Toxicity, and Other Stresses in Vegetable Crops
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AcdS | 1-Aminocyclopropane-1-carboxylate deaminase |
| CAT | Catalase |
| GPX | Guaiacol peroxidase |
| IAA | Indole-3-acetic acid |
| IST | Induced systemic tolerance |
| PAL | Phenylalanine ammonia-lyase |
| PGPR | Plant-growth-promoting rhizobacteria |
| P | Phosphorus |
| K | Potassium |
| ROS | Reactive oxygen species |
| SOD | Superoxide dismutase |
| WHO | World Health Organization |
References
- Schreinemachers, P.; Simmons, E.B.; Wopereis, M.C.S. Tapping the economic and nutritional power of vegetables. Glob. Food Secur. 2018, 16, 36–45. [Google Scholar] [CrossRef]
- Padulosi, S.; Sthapit, B.; Lamers, H.; Kennedy, G.; Hunter, D. Horticultural biodiversity to attain sustainable food and nutrition security. In Proceedings of the International Symposia on Tropical and Temperate Horticulture, Cairns, QLD, Australia, 20 November 2016; ISHS Acta Horticulturae: Leuven, Belgium, 2018; pp. 21–34. [Google Scholar] [CrossRef]
- Ebert, A.W. The Role of Vegetable Genetic Resources in Nutrition Security and Vegetable Breeding. Plants 2020, 9, 736. [Google Scholar] [CrossRef]
- Global Genetically Modified Seeds Market to Witness Growth Through 2020 Due to Rise in Adoption of Bio-Fuels: Reports Technavio; BusinessWire: London, UK, 2016.
- FAO Crop Prospects and Food Situation. Global Information and Early Warning System on Food and Agriculture (GIEWS) Trade and Markets Division (EST); FAO: Rome, Italy, 2018; pp. 1–40. [Google Scholar]
- Rasool, M.; Akhter, A.; Soja, G.; Haider, M.S. Role of biochar, compost and plant growth promoting rhizobacteria in the management of tomato early blight disease. Sci. Rep. 2021, 11, 6092. [Google Scholar] [CrossRef] [PubMed]
- Drozdowska, M.; Leszczyńska, T.; Koronowicz, A.; Piasna-Słupecka, E.; Domagała, D.; Kusznierewicz, B. Young shoots of red cabbage are a better source of selected nutrients and glucosinolates in comparison to the vegetable at full maturity. Eur. Food Res. Technol. 2020, 246, 2505–2515. [Google Scholar] [CrossRef]
- Van Treuren, R.; Van Eekelen, H.D.L.M.; Wehrens, R.; De Vos, R.C.H. Metabolite variation in the lettuce gene pool: Towards healthier crop varieties and food. Metabolomics 2018, 14, 146. [Google Scholar] [CrossRef] [Green Version]
- Park, J.-A.; Cho, S.K.; Kim, J.E.; Chung, H.S.; Hong, J.-P.; Hwang, B.; Hong, C.B.; Kim, W.T. Isolation of cDNAs differentially expressed in response to drought stress and characterization of the Ca-LEAL1 gene encoding a new family of atypical LEA-like protein homologue in hot pepper (Capsicum annuum L. cv. Pukang). Plant Sci. 2003, 165, 471–481. [Google Scholar] [CrossRef]
- Vimala, K.; Mohan, Y.M.; Sivudu, K.S.; Varaprasad, K.; Ravindra, S.; Reddy, N.N.; Padma, Y.; Sreedhar, B.; MohanaRaju, K. Fabrication of porous chitosan films impregnated with silver nanoparticles: A facile approach for superior antibacterial application. Colloids Surf. B Biointerfaces 2010, 76, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Gill, N.S.; Sharma, G.; Arora, R. Cucumis Trigonus Roxb: A Review. Int. J. Adv. Pharm. Sci. 2015, 5, 45–50. [Google Scholar]
- Khan, M.A.M.; Ulrichs, C.; Mewis, I. Effect of water stress and aphid herbivory on flavonoids in broccoli (Brassica oleracea var. italica Plenck). J. Appl. Bot. Food Qual. 2011, 84, 178–182. [Google Scholar]
- Owis, A.I. Broccoli; the green beauty: A review. J. Pharm. Sci. Res. 2015, 7, 696–703. [Google Scholar]
- Talalay, P.; Fahey, J.W. Phytochemicals from Cruciferous Plants Protect against Cancer by Modulating Carcinogen Metabolism. J. Nutr. 2001, 131, 3027S–3033S. [Google Scholar] [CrossRef]
- Emuh, F.N.; Ofuoku, A.E.; Oyefia, E. Effect of intercropping okra (Hibiscus esculentus) with pumpkin (Cucurbita maxima Duch Ex. Lam.) on some growth parameters and economic yield of maize (Zea mays) and maximization of land use in a Fadama soil. Res. J. Biol. Sci. 2006, 1, 50–54. [Google Scholar]
- Storey, M.; Anderson, P. Total fruit and vegetable consumption increases among consumers of frozen fruit and vegetables. Nutrition 2018, 46, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Schwingshackl, L.; Schwedhelm, C.; Hoffmann, G.; Lampousi, A.-M.; Knüppel, S.; Iqbal, K.; Bechthold, A.; Schlesinger, S.; Boeing, H. Food groups and risk of all-cause mortality: A systematic review and meta-analysis of prospective studies. Am. J. Clin. Nutr. 2017, 105, 1462–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Zeng, X.-T.; Liu, T.-Z.; Zhang, C.; Yang, Z.-H.; Li, S.; Chen, X.-Y. Fruits and Vegetables Intake and Risk of Bladder Cancer: A PRISMA-compliant systematic review and dose-response meta-analysis of prospective cohort studies. Medicine 2015, 94, e759. [Google Scholar] [CrossRef]
- Dosil-Díaz, O.; Ruano-Ravina, A.; Gestal-Otero, J.J.; Barros-Dios, J.M. Consumption of fruit and vegetables and risk of lung cancer: A case-control study in Galicia, Spain. Nutrition 2008, 24, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G. Protective effects of fruits and vegetables in the diet. Nutr. Food Sci. 1996, 96, 6–10. [Google Scholar] [CrossRef]
- Adebawo, O.; Salau, B.; Ezima, E.; Oyefuga, O.; Ajani, E.; Idowu, G.; Famodu, A.; Osilesi, O. Fruits and vegetables moderate lipid cardiovascular risk factor in hypertensive patients. Lipids Health Dis. 2006, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Celik, F.; Topcu, F. Nutritional risk factors for the development of chronic obstructive pulmonary disease (COPD) in male smokers. Clin. Nutr. 2006, 25, 955–961. [Google Scholar] [CrossRef]
- Payne, M.E.; Steck, S.E.; George, R.R.; Steffens, D.C. Fruit, Vegetable, and Antioxidant Intakes Are Lower in Older Adults with Depression. J. Acad. Nutr. Diet. 2012, 112, 2022–2027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMartin, S.E.; Jacka, F.N.; Colman, I. The association between fruit and vegetable consumption and mental health disorders: Evidence from five waves of a national survey of Canadians. Prev. Med. 2013, 56, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.J.; Edwards, I.; Hamernig, I.; Jian, L.; James, A.; Johnson, S.; Tapsell, L.C. Vegetables containing phytochemicals with potential anti-obesity properties: A review. Food Res. Int. 2013, 52, 323–333. [Google Scholar] [CrossRef] [Green Version]
- Kubec, R.; Svobodová, M.; Velíšek, J. Distribution of S-Alk(en)ylcysteine Sulfoxides in Some Allium Species. Identification of a New Flavor Precursor: S-Ethylcysteine Sulfoxide (Ethiin). J. Agric. Food Chem. 2000, 48, 428–433. [Google Scholar] [CrossRef]
- Clinton, S.K. Lycopene: Chemistry, Biology, and Implications for Human Health and Disease. Nutr. Rev. 1998, 56, 35–51. [Google Scholar] [CrossRef]
- Nielsen, S.E.; Young, J.F.; Daneshvar, B.; Lauridsen, S.T.; Knuthsen, P.; Sandström, B.; Dragsted, L.O. Effect of parsley (Petroselinum crispum) intake on urinary apigenin excretion, blood antioxidant enzymes and biomarkers for oxidative stress in human subjects. Br. J. Nutr. 1999, 81, 447–455. [Google Scholar] [CrossRef] [Green Version]
- Ching, L.S.; Mohamed, S. Alpha-Tocopherol Content in 62 Edible Tropical Plants. J. Agric. Food Chem. 2001, 49, 3101–3105. [Google Scholar] [CrossRef] [PubMed]
- Lila, M.A. Anthocyanins and Human Health: An In Vitro Investigative Approach. J. Biomed. Biotechnol. 2004, 5, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Horbowicz, M.; Kosson, R.; Grzesiuk, A.; Dębski, H. Anthocyanins of Fruits and Vegetables—Their Occurrence, Analysis and Role in Human Nutrition. J. Fruit Ornam. Plant Res. 2008, 68, 5–22. [Google Scholar] [CrossRef]
- Adesemoye, A.O.; Kloepper, J.W. Plant–microbes interactions in enhanced fertilizer-use efficiency. Appl. Microbiol. Biotechnol. 2009, 85, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Costa, P.B.; Beneduzi, A.; De Souza, R.; Schoenfeld, R.; Vargas, L.; Passaglia, L.M.P. The effects of different fertilization conditions on bacterial plant growth promoting traits: Guidelines for directed bacterial prospection and testing. Plant Soil 2012, 368, 267–280. [Google Scholar] [CrossRef]
- Pahalvi, H.N.; Rafiya, L.; Rashid, S.; Nisar, B.; Kamili, A.N. Chemical Fertilizers and Their Impact on Soil Health. Microbiota Biofertil. 2021, 2, 1–20. [Google Scholar] [CrossRef]
- Ye, H.; Cheng, J.; Yu, K. In situ reduction of silver nanoparticles by gelatin to obtain porous silver nanoparticle/chitosan composites with enhanced antimicrobial and wound-healing activity. Int. J. Biol. Macromol. 2018, 121, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Caris-Veyrat, C.; Amiot, M.-J.; Tyssandier, V.; Grasselly, D.; Buret, M.; Mikolajczak, M.; Guilland, J.-C.; Bouteloup-Demange, C.; Borel, P. Influence of Organic versus Conventional Agricultural Practice on the Antioxidant Microconstituent Content of Tomatoes and Derived Purees; Consequences on Antioxidant Plasma Status in Humans. J. Agric. Food Chem. 2004, 52, 6503–6509. [Google Scholar] [CrossRef] [PubMed]
- Luthria, D.; Singh, A.P.; Wilson, T.; Vorsa, N.; Banuelos, G.S.; Vinyard, B.T. Influence of conventional and organic agricultural practices on the phenolic content in eggplant pulp: Plant-to-plant variation. Food Chem. 2010, 121, 406–411. [Google Scholar] [CrossRef]
- Vallverdu-Queralt, A.; Medina-Remón, A.; Casals-Ribes, I.; Lamuela-Raventos, R.M. Is there any difference between the phenolic content of organic and conventional tomato juices? Food Chem. 2012, 130, 222–227. [Google Scholar] [CrossRef]
- Oliveira, A.B.; Moura, C.F.H.; Gomes-Filho, E.; Marco, C.A.; Urban, L.; Miranda, M.R.A. The Impact of Organic Farming on Quality of Tomatoes Is Associated to Increased Oxidative Stress during Fruit Development. PLoS ONE 2013, 8, e56354. [Google Scholar] [CrossRef] [Green Version]
- Ruano-Rosa, D.; Mercado-Blanco, J. Combining Biocontrol Agents and Organics Amendments to Manage Soil-Borne Phytopathogens. In Organic Amendments and Soil Suppressiveness in Plant Disease Management; Springer: Cham, Switzerland, 2015; pp. 457–478. [Google Scholar] [CrossRef]
- Beckles, D. Factors affecting the postharvest soluble solids and sugar content of tomato (Solanum lycopersicum L.) fruit. Postharvest Biol. Technol. 2012, 63, 129–140. [Google Scholar] [CrossRef]
- Ezura, H. Tomato is a Next-generation Model Plant for Research and Development. J. Jpn. Soc. Hortic. Sci. 2009, 78, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Ye, L.; Zhao, X.; Bao, E.; Li, J.; Zou, Z.; Cao, K. Bio-organic fertilizer with reduced rates of chemical fertilization improves soil fertility and enhances tomato yield and quality. Sci. Rep. 2020, 10, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weller, D.M. Biological Control of Soilborne Plant Pathogens in the Rhizosphere with Bacteria. Annu. Rev. Phytopathol. 1988, 26, 379–407. [Google Scholar] [CrossRef]
- Ferchichi, N.; Toukabri, W.; Boularess, M.; Smaoui, A.; Mhamdi, R.; Trabelsi, D. Isolation, identification and plant growth promotion ability of endophytic bacteria associated with lupine root nodule grown in Tunisian soil. Arch. Microbiol. 2019, 201, 1333–1349. [Google Scholar] [CrossRef] [PubMed]
- Artyszak, A.; Gozdowski, D. The Effect of Growth Activators and Plant Growth-Promoting Rhizobacteria (PGPR) on the Soil Properties, Root Yield, and Technological Quality of Sugar Beet. Agronomy 2020, 10, 1262. [Google Scholar] [CrossRef]
- Kour, D.; Rana, K.L.; Yadav, N.; Yadav, A.N.; Kumar, A.; Meena, V.S.; Singh, B.; Chauhan, V.S.; Dhaliwal, H.S.; Saxena, A.K. Rhizospheric Microbiomes: Biodiversity, Mechanisms of Plant Growth Promotion, and Biotechnological Applications for Sustainable Agriculture. In Plant Growth Promoting Rhizobacteria for Agricultural Sustainability; Springer: Singapore, 2019; pp. 19–65. [Google Scholar] [CrossRef]
- Gupta, S.; Kaushal, R.; Gupta, S. Plant Growth Promoting Rhizobacteria: Bioresouce for Enhanced Productivity of Solanaceous Vegetable Crops. Acta Sci. Agric. 2017, 1, 10–15. [Google Scholar]
- Sudewi, S.; Ala, A.; Patandjengi, B.; BDR, M.F.; Rahim, A. Scereening of Plant Growth Promotion Rhizobacteria (PGPR) to increase local aromatic rice plant growth. Int. J. Pharm. Res. 2020, 13. [Google Scholar] [CrossRef]
- Kumar, M.; Yusuf, M.A.; Chauhan, P.S.; Nigam, M. Pseudomonas putida and Bacillus amyloliquefaciens alleviates the adverse effect of pesticides and poise soil enzymes activities in chickpea (Cicer arietinum L.) rhizosphere. Trop. Plant Res. 2017, 4, 405–418. [Google Scholar] [CrossRef]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Patle, B.J.; Wagh, A.P.; Umbarkar, P.S.; Bondre, S. V Integrated nutrient management studies in bottle gourd. J. Pharmacogn. Phytochem. 2018, 7, 1383–1385. [Google Scholar]
- Seymen, M.; Türkmen, Ö.; Dursun, A.; Paksoy, M.; Dönmez, M.F. Effects of Bacteria Inoculation on Yield, Yield Components and Mineral Composition in Eggplant (Solanum melongena L.). In Proceedings of the ICOEST Conference, Urgüp, Turkey, 18–21 June 2013; pp. 403–413. [Google Scholar]
- Yildirim, E.; Karlidag, H.; Turan, M.; Dursun, A.; Goktepe, F. Promoition of Broccoli by Plant Growth Promoting Rhizobacteria. Hort. Sci. 2011, 46, 932–936. [Google Scholar]
- Gajbhiye, V.T.; Gupta, S.; Gupta, R.K. Persistence of Imidacloprid in/on Cabbage and Cauliflower. Bull. Environ. Contam. Toxicol. 2004, 72, 283–288. [Google Scholar] [CrossRef]
- Physiology, C.; Mog, B.; Ad, D. Effect of Organics and Biofertilizers on Productivity Potential in Carrot (Daucus carota L.). Ph.D. Thesis, UAS Dharwad, Dharwad, India, 2007. [Google Scholar]
- Silva, L.R.; Azevedo, J.; Pereira, M.J.; Valentão, P.; Andrade, P.B. Chemical assessment and antioxidant capacity of pepper (Capsicum annuum L.) seeds. Food Chem. Toxicol. 2012, 53, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Gül, A.; Özaktan, H.; Kıdoğlu, F.; Tüzel, Y. Rhizobacteria promoted yield of cucumber plants grown in perlite under Fusarium wilt stress. Sci. Hortic. 2013, 153, 22–25. [Google Scholar] [CrossRef]
- Chamangasht, S.; Ardakani, M.R.; Khavazi, K. Improving Lettuce (Lactuca sativa L.) Growth and Yield by the Application of Biofertilizers. Ann. Biol. Res. 2012, 3, 1876–1879. [Google Scholar]
- Kumar, M.; Baishya, L.K.; Ghosh, D.C.; Ghosh, M.; Gupta, V.K.; Verma, M.R. Effects of organic manures, chemical fertilizers and biofertilizers on growth and productivity of rainfed potato in the eastern himalayas. J. Plant Nutr. 2013, 36, 1065–1082. [Google Scholar] [CrossRef]
- Lee, J. Effect of application methods of organic fertilizer on growth, soil chemical properties and microbial densities in organic bulb onion production. Sci. Hortic. 2010, 124, 299–305. [Google Scholar] [CrossRef]
- Habibi, A.; Heidari, G.R.; Sohrabi, Y.; Mohamadi, K. Effect of biofertilizers and chemical fertilizers on yield and yield components of pumpkin (Cucurbita pepo L. Convar. pepo Var. styriaca). Iran. J. Med. Aromat. Plants 2013, 28, 604–615. [Google Scholar]
- Kumar, S.; Kumar, S.; Maji, S.; Pandey, V.K. Effect of inorganic fertilizers and bio-fertilizers on growth, yield and quality of radish (Raphanus sativus L.). Int. J. Plant Sci. 2016, 11, 71–74. [Google Scholar] [CrossRef]
- Bernabeu, P.R.; Pistorio, M.; Tejerizo, G.T.; Santos, P.E.D.L.; Galar, M.L.; Boiardi, J.L.; Luna, M.F. Colonization and plant growth-promotion of tomato by Burkholderia tropica. Sci. Hortic. 2015, 191, 113–120. [Google Scholar] [CrossRef]
- Ruzzi, M.; Aroca, R. Plant growth-promoting rhizobacteria act as biostimulants in horticulture. Sci. Hortic. 2015, 196, 124–134. [Google Scholar] [CrossRef]
- Vetrano, F.; Miceli, C.; Angileri, V.; Frangipane, B.; Moncada, A.; Miceli, A. Effect of Bacterial Inoculum and Fertigation Management on Nursery and Field Production of Lettuce Plants. Agronomy 2020, 10, 1477. [Google Scholar] [CrossRef]
- Moncada, A.; Vetrano, F.; Esposito, A.; Miceli, A. Fertigation Management and Growth-Promoting Treatments Affect Tomato Transplant Production and Plant Growth after Transplant. Agronomy 2020, 10, 1504. [Google Scholar] [CrossRef]
- Dey, R.; Pal, K.K.; Bhatt, D.M.; Chauhan, S.M. Growth promotion and yield enhancement of peanut (Arachis hypogaea L.) by application of plant growth-promoting rhizobacteria. Microbiol. Res. 2004, 159, 371–394. [Google Scholar] [CrossRef]
- Choudhary, D.K.; Sharma, K.P.; Gaur, R.K. Biotechnological perspectives of microbes in agro-ecosystems. Biotechnol. Lett. 2011, 33, 1905–1910. [Google Scholar] [CrossRef]
- Voisard, C.; Keel, C.; Haas, D.; Dèfago, G. Cyanide production by Pseudomonas fluorescens helps suppress black root rot of tobacco under gnotobiotic conditions. EMBO J. 1989, 8, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Rijavec, T.; Lapanje, A. Hydrogen Cyanide in the Rhizosphere: Not Suppressing Plant Pathogens, but Rather Regulating Availability of Phosphate. Front. Microbiol. 2016, 7, 1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agbodjato, N.A.; Noumavo, P.A.; Baba-Moussa, F.; Salami, H.A.; Sina, H.; Sèzan, A.; Bankolé, H.; Adjanohoun, A.; Baba-Moussa, L. Characterization of Potential Plant Growth Promoting Rhizobacteria Isolated from Maize (Zea mays L.) in Central and Northern Benin (West Africa). Appl. Environ. Soil Sci. 2015, 2015, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Honma, M.; Shimomura, T. Metabolism of 1-Aminocyclopropane-1-carboxylic Acid. Agric. Biol. Chem. 1978, 42, 1825–1831. [Google Scholar] [CrossRef]
- Glick, B.R.; Cheng, Z.; Czarny, J.; Duan, J. Promotion of plant growth by ACC deaminase-producing soil bacteria. Eur. J. Plant Pathol. 2007, 119, 329–339. [Google Scholar] [CrossRef]
- Singh, R.P.; Jha, P.N. The PGPR Stenotrophomonas maltophilia SBP-9 Augments Resistance against Biotic and Abiotic Stress in Wheat Plants. Front. Microbiol. 2017, 8, 1945. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Kloepper, J.W.; Ryu, C.-M. Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 2009, 14, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Igual, J.M.; Valverde, A.; Cervantes, E.; Velázquez, E. Phosphate-solubilizing bacteria as inoculants for agriculture: Use of updated molecular techniques in their study. Agronomie 2001, 21, 561–568. [Google Scholar] [CrossRef]
- Shahid, M.; Zaidi, A.; Khan, M.S.; Rizvi, A.; Saif, S.; Ahmed, B. Recent Advances in Management Strategies of Vegetable Diseases. Microbial Strategies for Vegetable Production; Springer: Cham, Switzerland, 2017; pp. 197–226. [Google Scholar] [CrossRef]
- Malboobi, M.A.; Owlia, P.; Behbahani, M.; Sarokhani, E.; Moradi, S.; Yakhchali, B.; Deljou, A.; Heravi, K.M. Solubilization of organic and inorganic phosphates by three highly efficient soil bacterial isolates. World J. Microbiol. Biotechnol. 2009, 25, 1471–1477. [Google Scholar] [CrossRef]
- Ali, A.M.; Awad, M.Y.M.; Hegab, S.A.; El Gawad, A.M.A.; Eissa, M.A. Effect of potassium solubilizing bacteria (Bacillus cereus) on growth and yield of potato. J. Plant Nutr. 2020, 44, 411–420. [Google Scholar] [CrossRef]
- Kushwaha, A.; Baily, S.B.; Maxton, A.; Ram, G.D. Isolation and characterization of PGPR associated with cauliflower roots and its effect on plant growth. Bioscan 2013, 8, 95–99. [Google Scholar]
- Altuntaş, A. Comparative study on the effects of different conventional, organic and bio-fertilizers on broccoli yield and quality. Appl. Ecol. Environ. Res. 2018, 16, 1595–1608. [Google Scholar] [CrossRef]
- Tanwar, A.; Aggarwal, A.; Parkash, V. Effect of bioinoculants and superphosphate fertilizer on the growth and yield of broccoli (Brassica oleracea L. var. italica Plenck). N. Zealand J. Crop. Hortic. Sci. 2014, 42, 288–302. [Google Scholar] [CrossRef] [Green Version]
- Nabi, R.B.S.; Shahzad, R.; Tayade, R.; Shahid, M.; Hussain, A.; Ali, M.W.; Yun, B.-W. Evaluation potential of PGPR to protect tomato against Fusarium wilt and promote plant growth. PeerJ 2021, 9, e11194. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, P.; Oh, Y.; Panthee, D.R. Current Status of Early Blight Resistance in Tomato: An Update. Int. J. Mol. Sci. 2017, 18, 2019. [Google Scholar] [CrossRef] [Green Version]
- Rasool, M.; Akhter, A.; Haider, M.S. Molecular and biochemical insight into biochar and Bacillus subtilis induced defense in tomatoes against Alternaria solani. Sci. Hortic. 2021, 285, 110203. [Google Scholar] [CrossRef]
- Tariq, M.; Ali, Q.; Khan, A.; Khan, G.A.; Rashid, B. “Advancements in Life Sciences” Yield potential study of Capsicum annuum L. under the application of PGPR. Adv. Life Sci. 2014, 1, 202–207. [Google Scholar]
- El-Azeem, S.A.M.A.; Elwan, M.W.M.; Sung, J.-K.; Ok, Y.S. Alleviation of Salt Stress in Eggplant (Solanum melongena L.) by Plant-Growth-Promoting Rhizobacteria. Commun. Soil Sci. Plant Anal. 2012, 43, 1303–1315. [Google Scholar] [CrossRef]
- Azarmi-Atajan, F.; Sayyari-Zohan, M.H. Alleviation of salt stress in lettuce (Lactuca sativa L.) by plant growth-promoting rhizobacteria. J. Hortic. Postharvest. Res. 2020, 3, 67–78. [Google Scholar] [CrossRef]
- Ünlükara, A.; Cemek, B.; Karaman, S.; Ersahin, S. Response of lettuce (Lactuca sativa var. crispa) to salinity of irrigation water. N. Zealand J. Crop. Hortic. Sci. 2008, 36, 265–273. [Google Scholar] [CrossRef]
- Julia, I.; Oscar, M.; Analía, L.; Guilherme, J.Z.; Virginia, L. Biofertilization with Macrocystis pyrifera algae extracts combined with PGPR-enhanced growth in Lactuca sativa seedlings. Environ. Boil. Fishes 2020, 32, 4361–4371. [Google Scholar] [CrossRef]
- Ayuso-Calles, M.; García-Estévez, I.; Jiménez-Gómez, A.; Flores-Félix, J.D.; Escribano-Bailón, M.T.; Rivas, R. Rhizobium laguerreae Improves Productivity and Phenolic Compound Content of Lettuce (Lactuca sativa L.) under Saline Stress Conditions. Foods 2020, 9, 1166. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, S.; Naveen, K.; Panneerselvam, K.; Sabitha, V. Investigation of in vivo antioxidant property of Abelmoschus esculentus (L) moench. fruit seed and peel powders in streptozotocin-induced diabetic rats. J. Ayurveda Integr. Med. 2012, 3, 188–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adesemoye, A.O.; Ugoji, E.O. Evaluating Pseudomonas aeruginosaas plant growth-promoting rhizobacteria in West Africa. Arch. Phytopathol. Plant Prot. 2009, 42, 188–200. [Google Scholar] [CrossRef] [Green Version]
- Zafar-Ul-Hye, M.; Tahzeeb-Ul-Hassan, M.; Abid, M.; Fahad, S.; Brtnicky, M.; Dokulilova, T.; Datta, R.; Danish, S. Potential role of compost mixed biochar with rhizobacteria in mitigating lead toxicity in spinach. Sci. Rep. 2020, 10, 12159. [Google Scholar] [CrossRef]
- Yan, Z.; Reddy, M.S.; Ryu, C.-M.; McInroy, J.A.; Wilson, M.; Kloepper, J.W. Induced Systemic Protection Against Tomato Late Blight Elicited by Plant Growth-Promoting Rhizobacteria. Phytopathology 2002, 92, 1329–1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misra, S.; Dixit, V.K.; Mishra, S.K.; Chauhan, P.S. Demonstrating the potential of abiotic stress-tolerant Jeotgalicoccus huakuii NBRI 13E for plant growth promotion and salt stress amelioration. Ann. Microbiol. 2019, 69, 419–434. [Google Scholar] [CrossRef]
- Zehnder, G.W.; Yao, C.; Murphy, J.F.; Sikora, E.D.R.; Kloepper, J.W. Induction of resistance in tomato against cucumber mosaic cucumovirus by plant growth-promoting rhizobacteria. Entomophaga 2000, 45, 127–137. [Google Scholar] [CrossRef]
- García-Pinilla, S.; Villalobos-Espinosa, J.C.; Cornejo-Mazón, M.; Gutiérrez-López, G.F. Nanotechnology in food processing. In Advances in Processing Technologies Bio-Based Nanosystem in Food; CRC Press: Boca Raton, FL, USA, 2019; pp. 259–276. [Google Scholar] [CrossRef]
- Felix, J.D.F.; Menendez, E.; Rivera, L.P.; Marcos-García, M.; Martínez-Hidalgo, P.; Mateos, P.; Martínez-Molina, E.; Velázquez, M.D.L.E.; García-Fraile, P.; Rivas, R. Use of Rhizobium leguminosarum as a potential biofertilizer for Lactuca sativaand Daucus carota crops. J. Plant Nutr. Soil Sci. 2013, 176, 876–882. [Google Scholar] [CrossRef]
- Khalid, M.; Hassani, D.; Bilal, M.; Asad, F.; Huang, D. Influence of bio-fertilizer containing beneficial fungi and rhizospheric bacteria on health promoting compounds and antioxidant activity of Spinacia oleracea L. Bot. Stud. 2017, 58, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Mishra, B.K.; Lal, G.; Sharma, Y.K.; Kant, K.; Saxena, S.N.; Dubey, P.N. Effect of microbial inoculants on cumin (Cuminum cyminum Linn.) growth and yield. Int. J. Seed Spices 2019, 53, 53–56. [Google Scholar]
- Guo, Q.; Li, Y.; Lou, Y.; Shi, M.; Jiang, Y.; Zhou, J.; Sun, Y.; Xue, Q.; Lai, H. Bacillus amyloliquefaciens Ba13 induces plant systemic resistance and improves rhizosphere microecology against tomato yellow leaf curl virus disease. Appl. Soil Ecol. 2019, 137, 154–166. [Google Scholar] [CrossRef]
- Niu, D.-D.; Wang, C.-J.; Guo, Y.-H.; Jiang, C.-H.; Zhang, W.-Z.; Wang, Y.-P.; Guo, J.-H. The plant growth-promoting rhizobacteriumBacillus cereusAR156 induces resistance in tomato with induction and priming of defence response. Biocontrol Sci. Technol. 2012, 22, 991–1004. [Google Scholar] [CrossRef]
- Carlson, R.; Tugizimana, F.; Steenkamp, P.A.; Dubery, I.A.; Hassen, A.I.; Labuschagne, N. Rhizobacteria-induced systemic resilience in Sorghum bicolor (L.) moench against Fusarium pseudograminearum crown rot under drought stress conditions. Biol. Control. 2020, 151, 104395. [Google Scholar] [CrossRef]
- Saravanakumar, D.; Samiyappan, R. ACC deaminase from Pseudomonas fluorescens mediated saline resistance in groundnut (Arachis hypogea) plants. J. Appl. Microbiol. 2006, 102, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Kaushal, R.; Spehia, R.S.; Pathania, S.S.; Sharma, V. Productivity of capsicum influenced by conjoint application of isolated indigenous PGPR and chemical fertilizers. J. Plant Nutr. 2017, 40, 921–927. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Giri, V.P.; Singh, S.P.; Pandey, S.; Chauhan, P.; Soni, S.K.; Srivastava, S.; Singh, P.C.; Mishra, A. Intervention of bio-protective endophyte Bacillus tequilensis enhance physiological strength of tomato during Fusarium wilt infection. Biol. Control. 2019, 139, 104074. [Google Scholar] [CrossRef]
- Dawwam, G.E.; Elbeltagy, A.; Emara, H.M.; Abbas, I.H.; Hassan, M.M. Beneficial effect of plant growth promoting bacteria isolated from the roots of potato plant. Ann. Agric. Sci. 2013, 58, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Ahemad, M.; Khan, M.S. Productivity of greengram in tebuconazole-stressed soil, by using a tolerant and plant growth-promoting Bradyrhizobium sp. MRM6 strain. Acta Physiol. Plant. 2011, 34, 245–254. [Google Scholar] [CrossRef]
- Tan, S.; Jiang, Y.; Song, S.; Huang, J.; Ling, N.; Xu, Y.; Shen, Q. Two Bacillus amyloliquefaciens strains isolated using the competitive tomato root enrichment method and their effects on suppressing Ralstonia solanacearum and promoting tomato plant growth. Crop. Prot. 2013, 43, 134–140. [Google Scholar] [CrossRef]
- Kurabachew, H.; Wydra, K. Characterization of plant growth promoting rhizobacteria and their potential as bioprotectant against tomato bacterial wilt caused by Ralstonia solanacearum. Biol. Control. 2013, 67, 75–83. [Google Scholar] [CrossRef]
- Kuarabachew, H.; Assefa, F.; Hiskias, Y. Evaluation of ethiopian isolates of Pseudomonas fluorescens as biocontrol agent against potato bacterial wilt caused by Ralstonia (Pseudomonas) solanacearum. Acta Agric. Slov. 2007, 2, 125–135. [Google Scholar]
- Zegeye, E.D.; Santhanam, A.; Gorfu, D.; Kassa, B. Biocontrol activity of Trichoderma viride and Pseudomonas fluorescens against Phytophthora infestans under greenhouse conditions. J. Agric. Technol. 2011, 7, 1589–1602. [Google Scholar]
- Cuevas, V.C.; Kebasen, S.B. Ecological approach in the control of club root disease of cabbage. In Proceedings of the 7th Annual Scientific Meeting and Symposium, Mycological Society of the Philippines, Laguna, Philippines, 8 April 2005. [Google Scholar]
- Bhagat, S.; Bambawale, O.M.; Tripathi, A.K.; Ahmad, I.; Srivastava, R.C. Biological management of fusarial wilt of tomato by Trichoderma spp. in Andamans. Indian J. Hortic. 2013, 70, 397–403. [Google Scholar]
- Singh, S.P.; Singh, H.B.; Singh, D.K. Biocontrol potential of mixture of trichoderma isolates on damping-off and collar rot of tomato. Bioscan 2014, 9, 1301–1304. [Google Scholar]
- Thakur, N.; Tripathi, A. Biological Management of Damping-Off, Buckeye Rot and Fusarial Wilt of Tomato (cv. Solan Lalima) under Mid-Hill Conditions of Himachal Pradesh. Agric. Sci. 2015, 6, 535–544. [Google Scholar] [CrossRef] [Green Version]
- Loganathan, M.; Garg, R.; Venkataravanappa, V.; Saha, S.; Rai, A.B. Plant growth promoting rhizobacteria (PGPR) induces resistance against Fusarium wilt and improves lycopene content and texture in tomato. Afr. J. Microbiol. Res. 2014, 8, 1105–1111. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Monaim, M.F.; Abdel-Gaid, M.A.; Zayan, S.A.; Nassef, D.M.T. Enhancement of Growth Parameters and Yield Components in Eggplant using Antagonism of Trichoderma spp. Against Fusarium Wilt Disease. Int. J. Phytopathol. 2014, 3, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, A.; Dutta, S.; Nandi, S.; Bhattacharya, I.; De Roy, M.; Sarkar, G.; Mandal, T. Antagonistic potentiality of native rhizobacterial isolates against root rot disease of okra, incited by Rhizoctonia solani. Afr. J. Agric. Res. 2013, 8, 405–412. [Google Scholar] [CrossRef]
- Shrestha, A.; Kim, B.S.; Park, D.H. Biological control of bacterial spot disease and plant growth-promoting effects of lactic acid bacteria on pepper. Biocontrol Sci. Technol. 2014, 24, 763–779. [Google Scholar] [CrossRef]
- Salim, H.A.; Kadhum, A.A.; Ali, A.F.; Saleh, U.N.; Jassim, N.H.; Hamad, A.R.; Attia, J.A.; Darwish, J.J.; Hassan, A.F. Response of cucumber plants to PGPR bacteria (Azospirillum brasilense, Pseudomonas fluorescens and Bacillus megaterium) and bread yeast (Saccharomyces cerevisiae). Syst. Rev. Pharm. 2021, 12, 969–975. [Google Scholar] [CrossRef]
- Khabbaz, S.E.; Abbasi, P.A. Isolation, characterization, and formulation of antagonistic bacteria for the management of seedlings damping-off and root rot disease of cucumber. Can. J. Microbiol. 2014, 60, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Shanthiyaa, V.; Saravanakumar, D.; Rajendran, L.; Karthikeyan, G.; Prabakar, K.; Raguchander, T. Use of Chaetomium globosum for biocontrol of potato late blight disease. Crop. Prot. 2013, 52, 33–38. [Google Scholar] [CrossRef]
- Sopheareth, M.; Chan, S.; Naing, K.W.; Lee, Y.S.; Hyun, H.N.; Kim, Y.C.; Kim, K.Y. Biocontrol of Late Blight (Phytophthora capsici) Disease and Growth Promotion of Pepper by Burkholderia cepacia MPC-7. Plant Pathol. J. 2013, 29, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Mane, M.M.; Lal, A.; Zghair, Q.N.; Simon, S. Efficacy of certain bio agents and fungicides against early blight of potato (Solanum tuberosum L.). Int. J. Plant Prot. 2014, 7, 433–436. [Google Scholar] [CrossRef]
- Park, K.; Park, J.-W.; Lee, S.-W.; Balaraju, K. Disease suppression and growth promotion in cucumbers induced by integrating PGPR agent Bacillus subtilis strain B4 and chemical elicitor ASM. Crop. Prot. 2013, 54, 199–205. [Google Scholar] [CrossRef]
- Cao, Y.; Pi, H.; Chandrangsu, P.; Li, Y.; Wang, Y.; Zhou, H.; Xiong, H.; Helmann, J.D.; Cai, Y. Antagonism of Two Plant-Growth Promoting Bacillus velezensis Isolates against Ralstonia solanacearum and Fusarium oxysporum. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Boyce, A.N. Role of Plant Growth Promoting Rhizobacteria in Agricultural Sustainability—A Review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef]
- Mhatre, P.H.; Karthik, C.; Kadirvelu, K.; Divya, K.L.; Venkatasalam, E.P.; Srinivasan, S.; Ramkumar, G.; Saranya, C.; Shanmuganathan, R. Plant growth promoting rhizobacteria (PGPR): A potential alternative tool for nematodes bio-control. Biocatal. Agric. Biotechnol. 2018, 17, 119–128. [Google Scholar] [CrossRef]
- Etesami, H.; Adl, S.M. Can interaction between silicon and non–rhizobial bacteria help in improving nodulation and nitrogen fixation in salinity–stressed legumes? A review. Rhizosphere 2020, 15, 100229. [Google Scholar] [CrossRef]
- Porter, S.S.; Bantay, R.; Friel, C.A.; Garoutte, A.; Gdanetz, K.; Ibarreta, K.; Moore, B.M.; Shetty, P.; Siler, E.; Friesen, M.L. Beneficial microbes ameliorate abiotic and biotic sources of stress on plants. Funct. Ecol. 2019, 34, 2075–2086. [Google Scholar] [CrossRef] [Green Version]
- Kalozoumis, P.; Savvas, D.; Aliferis, K.; Ntatsi, G.; Marakis, G.; Simou, E.; Tampakaki, A.; Karapanos, I. Impact of Plant Growth-Promoting Rhizobacteria Inoculation and Grafting on Tolerance of Tomato to Combined Water and Nutrient Stress Assessed via Metabolomics Analysis. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Subiramani, S.; Ramalingam, S.; Muthu, T.; Nile, S.H.; Venkidasamy, B. Development of Abiotic Stress Tolerance in Crops by Plant Growth-Promoting Rhizobacteria (PGPR). In Phyto-Microbiome Stress Regulation; Springer: Singapore, 2020; pp. 125–145. [Google Scholar] [CrossRef]
- Bhattacharyya, C.; Banerjee, S.; Acharya, U.; Mitra, A.; Mallick, I.; Haldar, A.; Haldar, S.; Ghosh, A.; Ghosh, A. Evaluation of plant growth promotion properties and induction of antioxidative defense mechanism by tea rhizobacteria of Darjeeling, India. Sci. Rep. 2020, 10, 1–19. [Google Scholar] [CrossRef]
- Paul, D.; Lade, H. Plant-growth-promoting rhizobacteria to improve crop growth in saline soils: A review. Agron. Sustain. Dev. 2014, 34, 737–752. [Google Scholar] [CrossRef]
- Jyothsna, P.; Murthy, S.D.S. A review on effect of senescence in plants and the role of phytohormones in delaying senescence. Int. J. Plant Anim. Environ. Sci. 2016, 6, 152–162. [Google Scholar]
- Kaushal, M.; Wani, S.P. Plant-growth-promoting rhizobacteria: Drought stress alleviators to ameliorate crop production in drylands. Ann. Microbiol. 2015, 66, 35–42. [Google Scholar] [CrossRef]
- Slettengren, M.; Mohanty, S.; Kamolvit, W.; van der Linden, J.; Brauner, A. Making medical devices safer: Impact of plastic and silicone oil on microbial biofilm formation. J. Hosp. Infect. 2020, 106, 155–162. [Google Scholar] [CrossRef]
- Xia, Y.; Farooq, A.; Javed, M.T.; Kamran, M.A.; Mukhtar, T.; Ali, J.; Tabassum, T.; Rehman, S.U.; Munis, M.F.H.; Sultan, T.; et al. Multi-stress tolerant PGPR Bacillus xiamenensis PM14 activating sugarcane (Saccharum officinarum L.) red rot disease resistance. Plant Physiol. Biochem. 2020, 151, 640–649. [Google Scholar] [CrossRef]
- Ali, S.; Hameed, S.; Shahid, M.; Iqbal, M.; Lazarovits, G.; Imran, A. Functional characterization of potential PGPR exhibiting broad-spectrum antifungal activity. Microbiol. Res. 2019, 232, 126389. [Google Scholar] [CrossRef] [PubMed]
- Kousar, B.; Bano, A.; Khan, N. PGPR Modulation of Secondary Metabolites in Tomato Infested with Spodoptera litura. Agronomy 2020, 10, 778. [Google Scholar] [CrossRef]
- Choudhary, D.K.; Varma, A. Microbial-mediated induced systemic resistance in plants. In Microbial-Mediated Induced Systemic Resistance in Plants; Springer: Singapore, 2016; pp. 147–162. [Google Scholar] [CrossRef]
- Calvo-Polanco, M.; Romera, B.S.; Aroca, R.; Asins, M.J.; Declerck, S.; Dodd, I.C.; Martínez-Andújar, C.; Albacete, A.; Ruiz-Lozano, J.M. Exploring the use of recombinant inbred lines in combination with beneficial microbial inoculants (AM fungus and PGPR) to improve drought stress tolerance in tomato. Environ. Exp. Bot. 2016, 131, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Lastochkina, O.; Pusenkova, L.; Yuldashev, R.; Babaev, M.; Garipova, S.; Blagova, D.; Khairullin, R.; Aliniaeifard, S. Effects of Bacillus subtilis on some physiological and biochemical parameters of Triticum aestivum L. (wheat) under salinity. Plant Physiol. Biochem. 2017, 121, 80–88. [Google Scholar] [CrossRef]
- Sarkar, A.; Ghosh, P.K.; Pramanik, K.; Mitra, S.; Soren, T.; Pandey, S.; Mondal, M.H.; Maiti, T.K. A halotolerant Enterobacter sp. displaying ACC deaminase activity promotes rice seedling growth under salt stress. Res. Microbiol. 2018, 169, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Habib, S.H.; Kausar, H.; Saud, H.M. Plant Growth-Promoting Rhizobacteria Enhance Salinity Stress Tolerance in Okra through ROS-Scavenging Enzymes. BioMed Res. Int. 2016, 2016, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palaniyandi, S.A.; Damodharan, K.; Yang, S.H.; Suh, J.W. Streptomyces sp. strain PGPA39 alleviates salt stress and promotes growth of ‘Micro Tom’ tomato plants. J. Appl. Microbiol. 2014, 117, 766–773. [Google Scholar] [CrossRef]
- Arkhipova, T.N.; Prinsen, E.; Veselov, S.U.; Martinenko, E.V.; Melentiev, A.I.; Kudoyarova, G.R. Cytokinin producing bacteria enhance plant growth in drying soil. Plant Soil 2007, 292, 305–315. [Google Scholar] [CrossRef]
- Lim, J.-H.; Kim, S.-D. Induction of Drought Stress Resistance by Multi-Functional PGPR Bacillus licheniformis K11 in Pepper. Plant Pathol. J. 2013, 29, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.-M.; Radhakrishnan, R.; Khan, A.L.; Kim, M.-J.; Park, J.-M.; Kim, B.-R.; Shin, D.-H.; Lee, I.-J. Gibberellin secreting rhizobacterium, Pseudomonas putida H-2-3 modulates the hormonal and stress physiology of soybean to improve the plant growth under saline and drought conditions. Plant Physiol. Biochem. 2014, 84, 115–124. [Google Scholar] [CrossRef]
- Fu, Q.; Liu, C.; Ding, N.; Lin, Y.; Guo, B. Ameliorative effects of inoculation with the plant growth-promoting rhizobacterium Pseudomonas sp. DW1 on growth of eggplant (Solanum melongena L.) seedlings under salt stress. Agric. Water Manag. 2010, 97, 1994–2000. [Google Scholar] [CrossRef]
- Gupta, A.; Rai, S.; Bano, A.; Khanam, A.; Sharma, S.; Pathak, N. Comparative Evaluation of Different Salt-Tolerant Plant Growth-Promoting Bacterial Isolates in Mitigating the Induced Adverse Effect of Salinity in Pisum sativum. Biointerface Res. Appl. Chem. 2021, 11, 13141–13154. [Google Scholar] [CrossRef]
- Bharti, N.; Barnawal, D.; Awasthi, A.; Yadav, A.; Kalra, A. Plant growth promoting rhizobacteria alleviate salinity induced negative effects on growth, oil content and physiological status in Mentha arvensis. Acta Physiol. Plant. 2013, 36, 45–60. [Google Scholar] [CrossRef]
- Kaymak, H.C. Potential of PGPR in Improvement of Environmental-Friendly Vegetable Production. In Field Crops: Sustainable Management by PGPR; Springer: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]
- Chinnaswamy, A.; de la Peña, T.C.; Stoll, A.; de la Peña Rojo, D.; Bravo, J.; Rincón, A.; Lucas, M.M.; Pueyo, J.J. A nodule endophytic Bacillus megaterium strain isolated from Medicago polymorpha enhances growth, promotes nodulation byEnsifer medicaeand alleviates salt stress in alfalfa plants. Ann. Appl. Biol. 2018, 172, 295–308. [Google Scholar] [CrossRef]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Jan, R.; Kang, S.-M.; Kim, K.-M.; Lee, I.-J. Extending thermotolerance to tomato seedlings by inoculation with SA1 isolate of Bacillus cereus and comparison with exogenous humic acid application. PLoS ONE 2020, 15, e0232228. [Google Scholar] [CrossRef]
- Dhanasekar, D.; Sivamani, P.; Panneersel, A.; Thajuddin, N.; Rajakumar, G.; Selvamani, S. Biological Control of Tomato Seedling Damping off with Streptomyces sp. Plant Pathol. J. 2005, 4, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Hassanisaadi, M.; Bonjar, G.S.; Hosseinipour, A.; Abdolshahi, R.; Barka, E.A.; Saadoun, I. Biological Control of Pythium aphanidermatum, the Causal Agent of Tomato Root Rot by two Streptomyces Root Symbionts. Agronomy 2021, 11, 846. [Google Scholar] [CrossRef]
- Chowdhury, S.P.; Dietel, K.; Rändler, M.; Schmid, M.; Junge, H.; Borriss, R.; Hartmann, A.; Grosch, R. Effects of Bacillus amyloliquefaciens FZB42 on Lettuce Growth and Health under Pathogen Pressure and Its Impact on the Rhizosphere Bacterial Community. PLoS ONE 2013, 8, e68818. [Google Scholar] [CrossRef] [Green Version]
- William, Q. Least-Toxic Controls of Plant Diseases. Nat. Dis. Control. 2000, 11, 225. [Google Scholar]
- Rose, S.; Parker, M.; Punja, Z. Efficacy of Biological and Chemical Treatments for Control of Fusarium Root and Stem Rot on Greenhouse Cucumber. Plant Dis. 2003, 87, 1462–1470. [Google Scholar] [CrossRef] [Green Version]
- Ravi, S.; Sabitha, D.; Valluvaparidasan, V.; Jeyalakshmi, C.; Doraiswamy, S. Effect of biocontrol agents on seed-borne Colletotrichum in French bean. Plant Dis. Res. 1999, 14, 146–151. [Google Scholar]
- Ramesh Kumar, N.; Thirumalai Arasu, V.; Gunasekaran, P. Genotyping of antifungal compounds producing plant growth-promoting rhizobacteria, Pseudomonas fluorescens. Curr. Sci. 2002, 82, 1463–1466. [Google Scholar]
- Paulitz, T.C.; Bélanger, R.R. Biological control in greenhouse systems. Annu. Rev. Phytopathol. 2001, 39, 103–133. [Google Scholar] [CrossRef]
- Singh, D.; Yadav, D.K.; Chaudhary, G.; Rana, V.S.; Sharma, R.K. Potential of Bacillus amyloliquefaciens for Biocontrol of Bacterial Wilt of Tomato Incited by Ralstonia solanacearum. J. Plant Pathol. Microbiol. 2016, 7, 1–6. [Google Scholar] [CrossRef]
- Hausbeck, M.K.; Lamour, K.H. Phytophthora capsici on Vegetable Crops: Research Progress and Management Challenges. Plant Dis. 2004, 88, 1292–1303. [Google Scholar] [CrossRef] [Green Version]
- Firmansyah, D.; Hidayat, S.H. Chitosan and Plant Growth Promoting Rhizobacteria Application to Control Squash mosaic virus on Cucumber Plants. Asian J. Plant Pathol. 2017, 11, 148–155. [Google Scholar] [CrossRef]
- Elbeshehy, E.K.F.; Youssef, S.A.; Elazzazy, A.M. Resistance induction in pumpkin Cucurbita maxima L. against Watermelon mosaic potyvirus by plant growth-promoting rhizobacteria. Biocontrol Sci. Technol. 2015, 25, 525–542. [Google Scholar] [CrossRef]
- Belimov, A.A.; Dodd, I.C.; Safronova, V.I.; Shaposhnikov, A.I.; Azarova, T.S.; Makarova, N.M.; Davies, W.J.; Tikhonovich, I. Rhizobacteria that produce auxins and contain 1-amino-cyclopropane-1-carboxylic acid deaminase decrease amino acid concentrations in the rhizosphere and improve growth and yield of well-watered and water-limited potato (Solanum tuberosum). Ann. Appl. Biol. 2015, 167, 11–25. [Google Scholar] [CrossRef]
- Ullah, U.; Ashraf, M.; Shahzad, S.M.; Siddiqui, A.R.; Piracha, M.A.; Suleman, M. Growth behavior of tomato (Solanum lycopersicum L.) under drought stress in the presence of silicon and plant growth promoting rhizobacteria. Soil Environ. 2016, 35, 65–75. [Google Scholar]
- Kuşçu, H.; Turhan, A.; Demir, A.O. The response of processing tomato to deficit irrigation at various phenological stages in a sub-humid environment. Agric. Water Manag. 2014, 133, 92–103. [Google Scholar] [CrossRef]
- Lulsdorf, M.M.; Yuan, H.Y.; Slater, S.M.H.; Vandenberg, A.; Han, X.; Zaharia, L.I.; Abrams, S.R. Endogenous hormone profiles during early seed development of C. arietinum and C. anatolicum. Plant Growth Regul. 2013, 71, 191–198. [Google Scholar] [CrossRef]
- Lidon, Z.Z.; Cebola, F. An overview on drought induced changes in plant growth, water relations and photosynthesis. Emirates J. Food Agric. 2012, 24, 57–72. [Google Scholar] [CrossRef] [Green Version]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Mishra, S.; Dixit, V.; Agarwal, L.; Chauhan, P.S.; Nautiyal, C.S. Synergistic effect of Pseudomonas putida and Bacillus amyloliquefaciens ameliorates drought stress in chickpea (Cicer arietinum L.). Plant Signal. Behav. 2016, 11, e1071004. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Xing, S.; Ma, H.; Du, Z.; Ma, B. Cytokinin-producing, plant growth-promoting rhizobacteria that confer resistance to drought stress in Platycladus orientalis container seedlings. Appl. Microbiol. Biotechnol. 2013, 97, 9155–9164. [Google Scholar] [CrossRef]
- Creus, C.M.; Graziano, M.; Casanovas, E.M.; Pereyra, M.A.; Simontacchi, M.; Puntarulo, S.; Barassi, C.A.; LaMattina, L. Nitric Oxide is Involved in the Azospirillum brasilense-induced Lateral Root Formation in Tomato. Planta 2005, 221, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Batool, T.; Ali, S.; Seleiman, M.F.; Naveed, N.H.; Ali, A.; Ahmed, K.; Abid, M.; Rizwan, M.; Shahid, M.R.; Alotaibi, M.; et al. Plant growth promoting rhizobacteria alleviates drought stress in potato in response to suppressive oxidative stress and antioxidant enzymes activities. Sci. Rep. 2020, 10, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Guo, Y.; Wang, C.; Liu, H.; Niu, D.; Wang, Y.; Guo, J. Enhancement of tomato (Lycopersicon esculentum) tolerance to drought stress by plant-growth-promoting rhizobacterium (PGPR) Bacillus cereus AR156. J. Agric. Biotechnol. 2012, 20, 1097–1105. [Google Scholar]
- Shannon, M.C.; Grieve, C.M. Tolerance of vegetable crops to salinity. Sci. Hortic. 1998, 78, 5–38. [Google Scholar] [CrossRef]
- Shahbaz, M.; Ashraf, M.; Al-Qurainy, F.; Harris, P.J.C. Salt Tolerance in Selected Vegetable Crops. Crit. Rev. Plant Sci. 2012, 31, 303–320. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Lauchli, A.; Epstein, E. Plant Responses to Saline and Sodic Conditions and Agricultural Salinity Assessment and Management; ASCE: New York, NY, USA, 1990; pp. 113–137. [Google Scholar]
- Ekinci, M.; Turan, M.; Yildirim, E.; Güneş, A.; Kotan, R.; Dursun, A. Effect of plant growth promoting rhizobacteria on growth, nutrient, organic acid, amino acid and hormone content of cauliflower (Brassica oleracea L. var. botrytis) transplants. Acta Sci. Pol. Hortorum Cultus 2014, 13, 71–85. [Google Scholar]
- Xu, C.; Mou, B. Evaluation of Lettuce Genotypes for Salinity Tolerance. HortScience 2015, 50, 1441–1446. [Google Scholar] [CrossRef] [Green Version]
- Kidoglu, F.; Gül, A.; Özaktan, H.; Tüzel, Y. Effect of rhizobacteria on plant growth of different vegetables. Acta Hortic. 2008, 801, 1471–1478. [Google Scholar] [CrossRef]
- Bochow, H.; El-Sayed, S.F.; Junge, H.; Stavropoulou, A.; Schmiedeknecht, G. Use of Bacillus subtilis as biocontrol agent. IV. Salt-stress tolerance induction by Bacillus subtilis FZB24 seed treatment in tropical vegetable field crops, and its mode of action. Z. Pflanzenkrankh. Pflanzenschutz 2001, 108, 21–30. [Google Scholar]
- Bensalim, S.; Nowak, J.; Asiedu, S.K. A plant growth promoting rhizobacterium and temperature effects on performance of 18 clones of potato. Am. J. Potato Res. 1998, 75, 145–152. [Google Scholar] [CrossRef]
- Martin, C.A.; Stutz, J.C. Interactive effects of temperature and arbuscular mycorrhizal fungi on growth, P uptake and root respiration of Capsicum annuum L. Mycorrhiza 2003, 14, 241–244. [Google Scholar] [CrossRef]
- Mukhtar, T.; Rehman, S.U.; Smith, D.; Sultan, T.; Seleiman, M.F.; Alsadon, A.A.; Ali, S.; Chaudhary, H.J.; Solieman, T.H.I.; Saad, M.A.; et al. Mitigation of Heat Stress in Solanum lycopersicum L. by ACC-deaminase and Exopolysaccharide Producing Bacillus cereus: Effects on Biochemical Profiling. Sustainability 2020, 12, 2159. [Google Scholar] [CrossRef] [Green Version]
- Rubio, M.I.; Escrig, I.; Martínez-Cortina, C.; López-Benet, F.J.; Sanz, A. Cadmium and nickel accumulation in rice plants. Effects on mineral nutrition and possible interactions of abscisic and gibberellic acids. Plant Growth Regul. 1994, 14, 151–157. [Google Scholar] [CrossRef]
- Schuurmann, G.; Markert, B. Effects of heavy metals in plants at the cellular and organismic level. In Ecotoxicology; John Wiley and Sons: Heidelberg, Germany, 1998; pp. 587–620. [Google Scholar]
- Fatnassi, I.C.; Chiboub, M.; Saadani, O.; Jebara, M.; Jebara, S.H. Impact of dual inoculation with Rhizobium and PGPR on growth and antioxidant status of Vicia faba L. under copper stress. Comptes Rendus Biol. 2015, 338, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Gururani, M.A.; Upadhyaya, C.P.; Baskar, V.; Venkatesh, J.; Nookaraju, A.; Park, S.W. Plant Growth-Promoting Rhizobacteria Enhance Abiotic Stress Tolerance in Solanum tuberosum Through Inducing Changes in the Expression of ROS-Scavenging Enzymes and Improved Photosynthetic Performance. J. Plant Growth Regul. 2012, 32, 245–258. [Google Scholar] [CrossRef]
- Safronova, V.I.; Stepanok, V.V.; Engqvist, G.L.; Alekseyev, Y.V.; Belimov, A.A. Root-associated bacteria containing 1-aminocyclopropane-1-carboxylate deaminase improve growth and nutrient uptake by pea genotypes cultivated in cadmium supplemented soil. Biol. Fertil. Soils 2005, 42, 267–272. [Google Scholar] [CrossRef]
- I Burd, G.; Dixon, D.G.; Glick, B.R. Plant growth-promoting bacteria that decrease heavy metal toxicity in plants. Can. J. Microbiol. 2000, 46, 237–245. [Google Scholar] [CrossRef]
- Singh, A.D.; Sharma, P.; Kohli, S.K.; Kumar, P.; Singh, R.; Arora, P.; Sharma, P.; Kaur, R.; Sharma, A.; Bhardwaj, R. Role of Plant Growth Regulators (PGRs) in Mitigation of Heavy Metal Phytotoxicity in Plants. In Cellular and Molecular Phytotoxicity of Heavy Metals; Springer: Cham, Switzerland, 2020; pp. 263–304. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, M.; Giri, V.P.; Pandey, S.; Gupta, A.; Patel, M.K.; Bajpai, A.B.; Jenkins, S.; Siddique, K.H.M. Plant-Growth-Promoting Rhizobacteria Emerging as an Effective Bioinoculant to Improve the Growth, Production, and Stress Tolerance of Vegetable Crops. Int. J. Mol. Sci. 2021, 22, 12245. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212245
Kumar M, Giri VP, Pandey S, Gupta A, Patel MK, Bajpai AB, Jenkins S, Siddique KHM. Plant-Growth-Promoting Rhizobacteria Emerging as an Effective Bioinoculant to Improve the Growth, Production, and Stress Tolerance of Vegetable Crops. International Journal of Molecular Sciences. 2021; 22(22):12245. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212245
Chicago/Turabian StyleKumar, Manoj, Ved Prakash Giri, Shipra Pandey, Anmol Gupta, Manish Kumar Patel, Atal Bihari Bajpai, Sasha Jenkins, and Kadambot H. M. Siddique. 2021. "Plant-Growth-Promoting Rhizobacteria Emerging as an Effective Bioinoculant to Improve the Growth, Production, and Stress Tolerance of Vegetable Crops" International Journal of Molecular Sciences 22, no. 22: 12245. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212245