
Comparing the Osteogenic Potential and Bone Regeneration Capacities of Dedifferentiated Fat Cells and Adipose-Derived Stem Cells In Vitro and In Vivo: Application of DFAT Cells Isolated by a Mesh Method

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
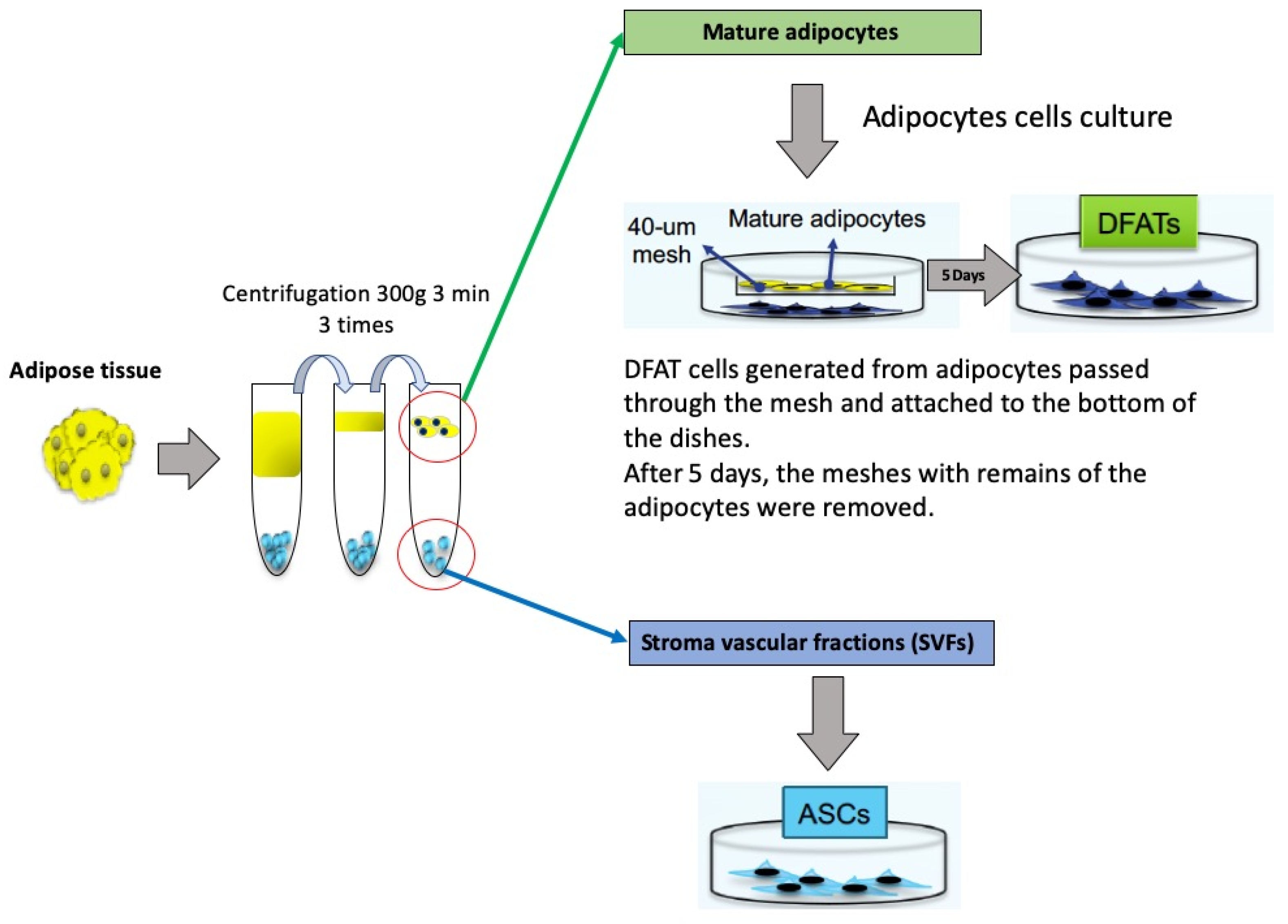
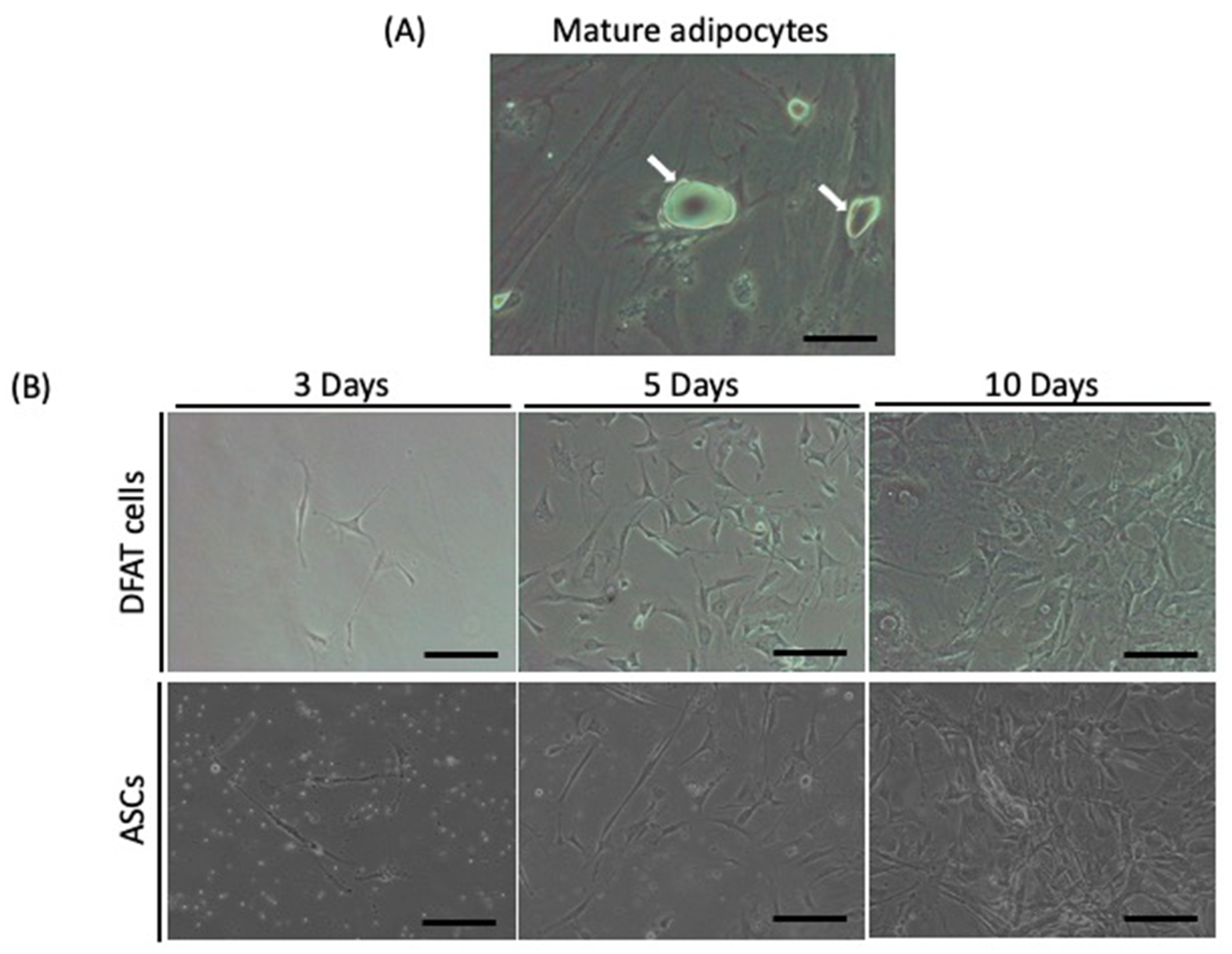
2.1. Isolation of DFAT Cells and ASCs from Adipose Tissue
2.2. Osteoblastic Differentiation of DFAT Cells and ASCs In Vitro
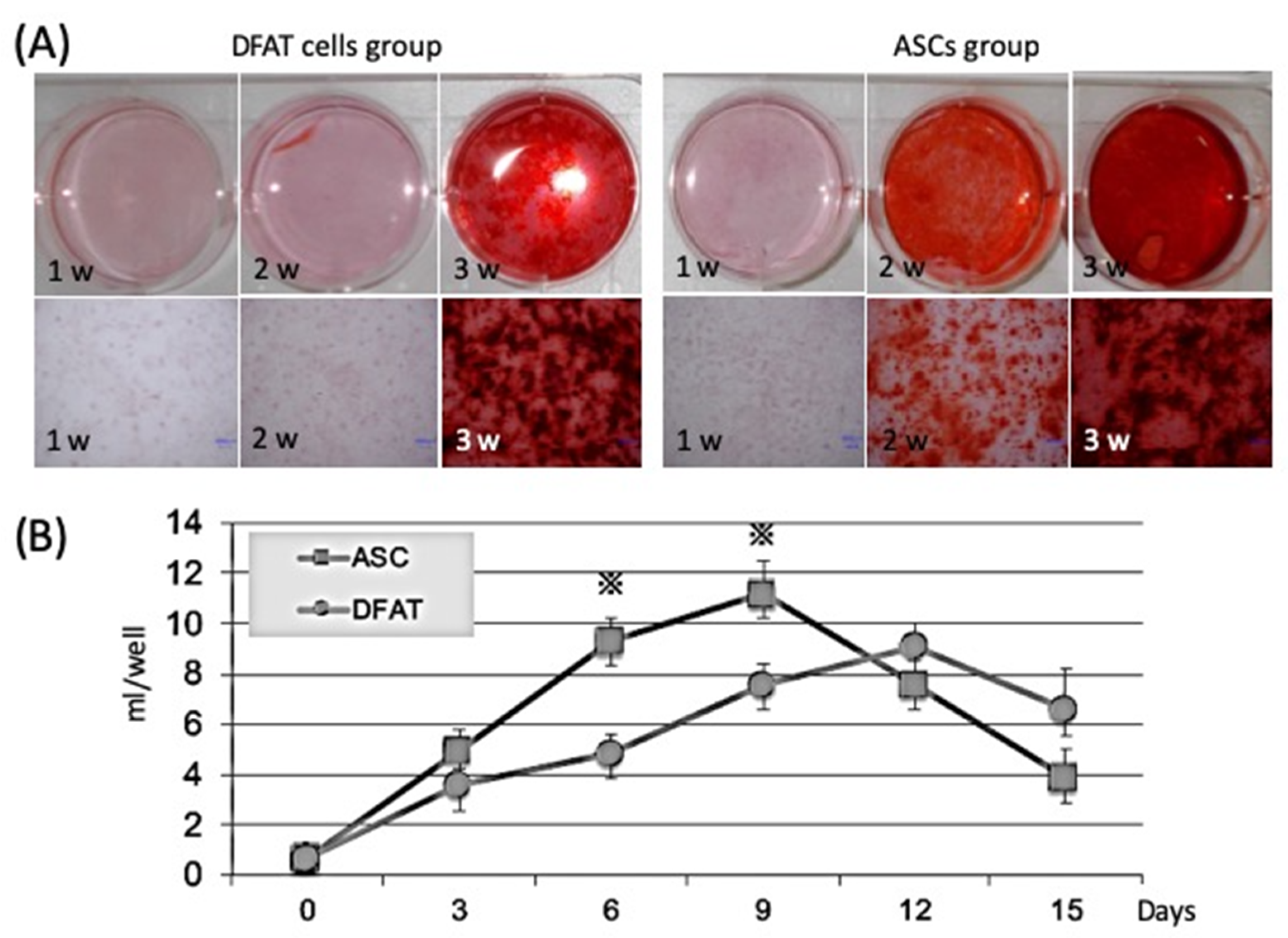
2.2.1. Comparison of Calcification Ability by Alizarin Red Staining
2.2.2. Comparison of Alkaline Phosphatase (ALP) Activity
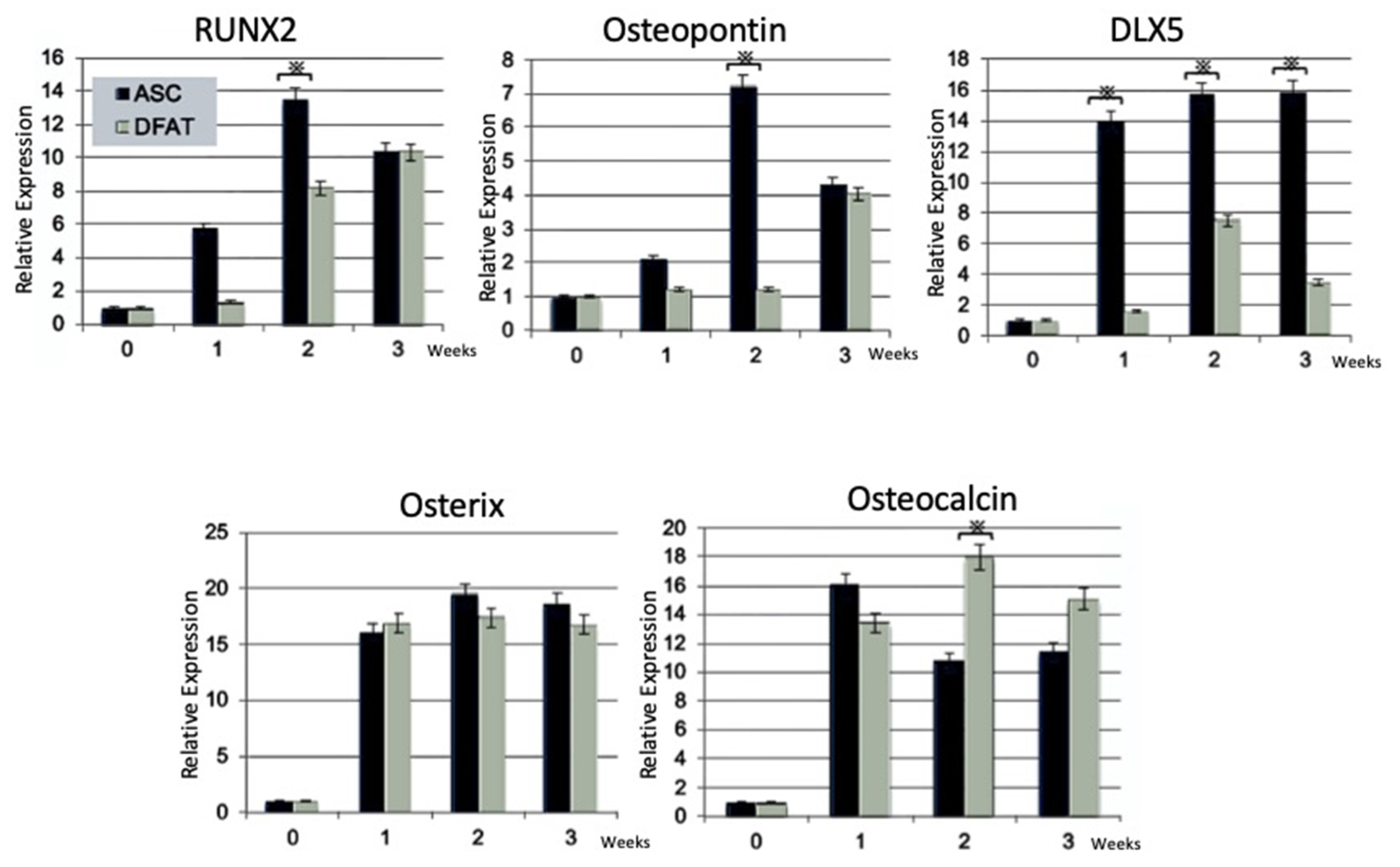
2.2.3. Reverse Transcription-Polymerase Chain Reaction (RT-PCR) Analysis
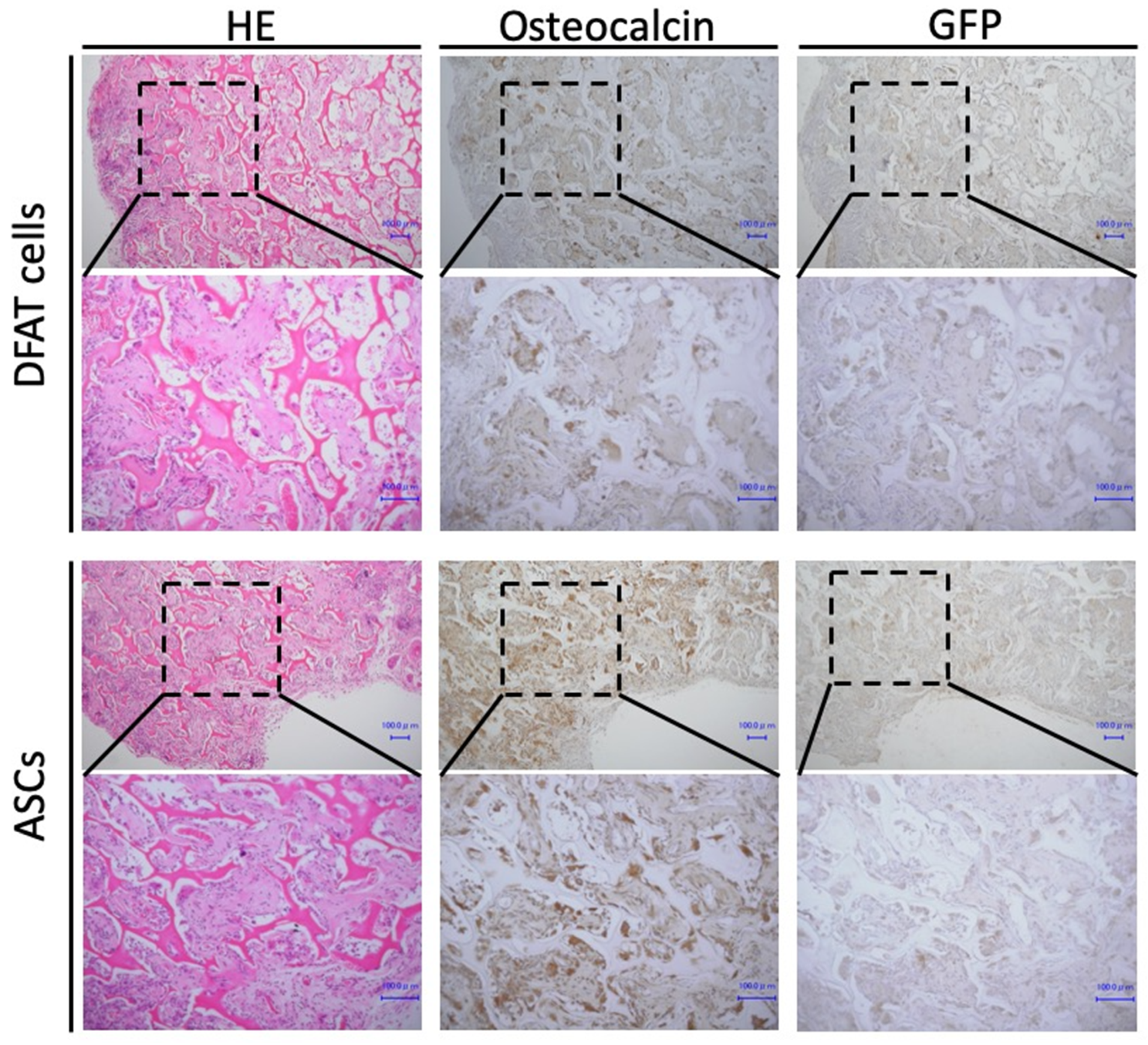
2.3. Formation of Osteoid Tissue by Transplanted DFAT Cells and ASCs
3. Discussion
4. Materials and Methods
4.1. Isolation and Culture of DFAT Cells and ASCs
4.2. Mineralization Assay by Alizarin Red Staining
4.3. ELISA for Quantification of Alkaline Phosphatase
4.4. Immunohistochemical Staining of GFP and Osteocalcin
4.5. Transplantation and Histological Evaluation
4.6. Reverse Transcription-Polymerase Chain Reaction (PCR) Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Liu, J.; Yu, F.; Sun, Y.; Jiang, B.; Zhang, W.; Yang, J.; Xu, G.T.; Liang, A.; Liu, S. Concise reviews: Characteristics and potential applications of human dental tissue-derived mesenchymal stem cells. Stem Cells 2015, 33, 627–638. [Google Scholar] [CrossRef]
- Xu, L.; Liu, Y.; Sun, Y.; Wang, B.; Xiong, Y.; Lin, W.; Wei, Q.; Wang, H.; He, W.; Wang, B.; et al. Tissue source determines the differentiation potentials of mesenchymal stem cells: A comparative study of human mesenchymal stem cells from bone marrow and adipose tissue. Stem Cell Res. Ther. 2017, 8, 275. [Google Scholar] [CrossRef] [Green Version]
- Abdel Meguid, E.; Ke, Y.; Ji, J.; El-Hashash, A.H.K. Stem cells applications in bone and tooth repair and regeneration: New insights, tools, and hopes. J. Cell Physiol. 2018, 233, 1825–1835. [Google Scholar] [CrossRef] [PubMed]
- Marco, T.; Bruna, C.; Francesco, P.; Manuel, N.; Irina, M. Strategic tools in regenerative and translational dentistry. Int. J. Mol. Sci. 2019, 20, 1879. [Google Scholar]
- Hajime, H.; Kimito, S.; Hitoshi, I.; Shogo, O. Differentiation of dental pulp-derived MSCs into Hepatocyte-like cells and their therapeutic use for chemical liver injuries of rat. J. Hard Tissue Biol. 2020, 29, 215–222. [Google Scholar]
- Shintaro, H.; Kaname, S.; Akira, T. A study on the effect of human dental pulp stem cell conditioned medium on human oral squamous cell carcinoma cell lines. J. Hard Tissue Biol. 2019, 28, 281–288. [Google Scholar]
- Lee, Y.C.; Chan, Y.H.; Hsieh, S.C.; Lew, W.Z.; Feng, S.W. Comparing the Osteogenic potentials and bone regeneration capacities of bone marrow and dental pulp mesenchymal stem cells in a rabbit calvarial bone defect model. Int. J. Mol. Sci. 2019, 20, 5015. [Google Scholar] [CrossRef] [Green Version]
- Park, J.C.; Oh, S.Y.; Lee, J.S.; Park, S.Y.; Choi, E.Y.; Cho, K.S.; Kim, C.S. In vivo bone formation by human alveolar-bone-derived mesenchymal stem cells obtained during implant osteotomy using biphasic calcium phosphate ceramics or Bio-Oss as carriers. J. Biomed. Mater. Res. B Appl. Biomater. 2016, 104, 515–524. [Google Scholar] [CrossRef]
- Kunkel, N.; Wagner, A.; Gehwolf, R.; Heimel, P.; Tempfer, H.; Korntner, S.; Augat, P.; Resch, H.; Redl, H.; Betz, O.; et al. Comparing the osteogenic potential of bone marrow and tendon-derived stromal cells to repair a critical-sized defect in the rat femur. J. Tissue Eng. Regen. Med. 2017, 11, 2014–2023. [Google Scholar] [CrossRef] [PubMed]
- Janko, M.; Sahm, J.; Schaible, A.; Brune, J.C.; Bellen, M.; Schroder, K.; Seebach, C.; Marzi, I.; Henrich, D. Comparison of three different types of scaffolds preseeded with human bone marrow mononuclear cells on the bone healing in a femoral critical size defect model of the athymic rat. J. Tissue Eng. Regen. Med. 2018, 12, 653–666. [Google Scholar] [CrossRef]
- Zhang, J.; Xu, D.-p.; Shang, J.; Liang, X.-j.; Zhang, X.-b. Effects of Naringin on the proliferation and osteogenic differentiation of canine bone marrow stromal cell in vitro. J. Hard Tissue Biol. 2021, 30, 73–78. [Google Scholar]
- Hua, S.; Shiqi, S.; Chao, N.; Yang, Z.; Weidong, M.; Song, C. Bone marrow mesenchymal stem cells transplantation on acute spinal cord injury. J. Hard Tissue Biol. 2020, 29, 91–98. [Google Scholar]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage cells from human adipose tissue: Implications for cell-based therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizuno, H.; Tobita, M.; Uysal, A.C. Concise review: Adipose-derived stem cells as a novel tool for future regenerative medicine. Stem Cells 2012, 30, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, R.; Mizuno, H.; Watanabe, A.; Migita, M.; Shimada, T.; Hyakusoku, H. Osteogenic and chondrogenic differentiation by adipose-derived stem cells harvested from GFP transgenic mice. Biochem. Biophys. Res. Commun. 2004, 313, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Mikako, I.; Masanori, T.; Kumiko, O.; Naohiro, S. Effects of adipose tissue-derived stem cells transplanted to a bone defect after irradiation. J. Hard Tissue Biol. 2020, 29, 111–122. [Google Scholar]
- Chengdong, P.; Zhengwei, L.; Jie, D.; Daliang, K. Comparative analysis of bone mechanical properties of adipose-derived mesenchymal stem cells and raloxifene in treatment of osteoporosis. J. Hard Tissue Biol. 2020, 29, 71–76. [Google Scholar]
- Farré-Guasch, E.; Martí-Pagè, C.; Hernádez-Alfaro, F.; Klein-Nulend, J.; Casals, N. Buccal fat pad, an oral access source of human adipose stem cells with potential for osteochondral tissue engineering: An in vitro study. Tissue Eng. Part C Methods 2010, 16, 1083–1094. [Google Scholar] [CrossRef] [Green Version]
- Yoon, E.; Dhar, S.; Chun, D.E.; Gharibjanian, N.A.; Evans, G.R. In vivo osteogenic potential of human adipose-derived stem cells/poly lactide-co-glycolic acid constructs for bone regeneration in a rat critical-sized calvarial defect model. Tissue Eng. 2007, 13, 619–627. [Google Scholar] [CrossRef]
- Lin, Y.; Tang, W.; Wu, L.; Jing, W.; Li, X.; Wu, Y.; Liu, L.; Long, J.; Tian, W. Bone regeneration by BMP-2 enhanced adipose stem cells loading on alginate gel. Histochem. Cell Biol. 2008, 129, 203–210. [Google Scholar] [CrossRef]
- Li, H.; Dai, K.; Tang, T.; Zhang, X.; Yan, M.; Lou, J. Bone regeneration by implantation of adipose-derived stromal cells expressing BMP-2. Biochem. Biophys. Res. Commun. 2007, 356, 836–842. [Google Scholar] [CrossRef]
- Matsumoto, T.; Kano, K.; Kondo, D.; Fukuda, N.; Iribe, Y.; Tanaka, N.; Matsubara, Y.; Sakuma, T.; Satomi, A.; Otaki, M.; et al. Mature adipocyte-derived dedifferentiated fat cells exhibit multilineage potential. J. Cell Physiol. 2008, 215, 210–222. [Google Scholar] [CrossRef]
- Yagi, K.; Kondo, D.; Okazaki, Y.; Kano, K. A novel preadipocyte cell line established from mouse adult mature adipocytes. Biochem. Biophys. Res. Commun. 2004, 321, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, T.; Matsumoto, T.; Kano, K.; Fukuda, N.; Obinata, D.; Yamaguchi, K.; Yoshida, T.; Takahashi, S.; Mugishima, H. Mature, adipocyte derived, dedifferentiated fat cells can differentiate into smooth muscle-like cells and contribute to bladder tissue regeneration. J. Urol. 2009, 182, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Kazama, T.; Fujie, M.; Endo, T.; Kano, K. Mature adipocyte-derived dedifferentiated fat cells can transdifferentiate into skeletal myocytes in vitro. Biochem. Biophys. Res. Commun. 2008, 377, 780–785. [Google Scholar] [CrossRef]
- Umabay, M.; Matsumoto, T.; Yokoyama, S.; Kano, K.; Kusumi, Y.; Masuko, T.; Mitsumata, M.; Saito, S.; Hirayama, A.; Mugishima, H.; et al. Dedifferentiated fat cells convert to cardiomyocyte phenotype and repair infarcted cardiac tissue in rats. J. Mol. Cell Cardiol. 2009, 47, 565–575. [Google Scholar]
- Obinata, D.; Matsumoto, T.; Ikado, Y.; Sakuma, T.; Kano, K.; Fukuda, N.; Yamaguchi, K.; Mugishima, H.; Takahashi, S. Transplantation of mature adipocyte-derived dedifferentiated fat (DFAT) cells improves urethral sphincter contractility in a rat model. Int. J. Urol. 2011, 18, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Nobusue, H.; Endo, T.; Kano, K. Establishment of a preadipocyte cell line derived from mature adipocytes of GFP transgenic mice and formation of adipose tissue. Cell Tissue Res. 2008, 332, 435–446. [Google Scholar] [CrossRef]
- Jumabay, M.; Zhang, R.; Yao, Y.; Goldhaber, J.I.; Boström, K.I. Spontaneously beating cardiomyocytes derived from white mature adipocytes. Cardiovasc. Res. 2010, 85, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Sugihara, H.; Funatsumaru, S.; Yonemitsu, N.; Miyabara, S.; Toda, S.; Hikichi, Y. A simple culture method of fat cells from mature fat tissue fragments. J. Lipid Res. 1989, 30, 1987–1995. [Google Scholar] [CrossRef]
- Zhang, H.H.; Kumar, S.; Barnett, A.H.; Eggo, M.C. Ceiling culture of mature human adipocytes: Use in studies of adipocyte functions. J. Endocrinol. 2000, 164, 119–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akita, D.; Kano, K.; Saito-Tamura, Y.; Mashimo, T.; Sato-Shionome, M.; Tsurumachi, N.; Yamanaka, K.; Kaneko, T.; Toriumi, T.; Arai, Y.; et al. Use of Rat Mature Adipocyte-Derived Dedifferentiated Fat Cells as a Cell Source for Periodontal Tissue Regeneration. Front. Physiol. 2016, 7, 50. [Google Scholar] [CrossRef] [Green Version]
- Kono, S.; Kazama, T.; Kano, K.; Harada, K.; Uechi, M.; Matsumoto, T. Phenotypic and functional properties of feline dedifferentiated fat cells and adipose-derived stem cells. Vet. J. 2014, 199, 88–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishimoto, N.; Momota, Y.; Hashimoto, Y.; Tatsumi, S.; Ando, K.; Omasa, T.; Kotani, J. The osteoblastic differentiation ability of human dedifferentiated fat cells is higher than that of adipose stem cells from the buccal fat pad. Clin. Oral. Investig. 2014, 18, 1893–1901. [Google Scholar] [CrossRef] [Green Version]
- Kishimoto, N.; Momota, Y.; Hashimoto, Y.; Ando, K.; Omasa, T.; Kotani, J. Dedifferentiated fat cells differentiate into osteoblasts in titanium fiber mesh. Cytotechnology 2013, 65, 15–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Q.; Zhao, L.; Song, Z.; Yang, G. Expression pattern of embryonic stem cell markers in DFAT cells and ADSCs. Mol. Biol. Rep. 2012, 39, 5791–5804. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takabatake, K.; Matsubara, M.; Yamachika, E.; Fujita, Y.; Arimura, Y.; Nakatsuji, K.; Nakano, K.; Nagatsuka, H.; Iida, S. Comparing the Osteogenic Potential and Bone Regeneration Capacities of Dedifferentiated Fat Cells and Adipose-Derived Stem Cells In Vitro and In Vivo: Application of DFAT Cells Isolated by a Mesh Method. Int. J. Mol. Sci. 2021, 22, 12392. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212392
Takabatake K, Matsubara M, Yamachika E, Fujita Y, Arimura Y, Nakatsuji K, Nakano K, Nagatsuka H, Iida S. Comparing the Osteogenic Potential and Bone Regeneration Capacities of Dedifferentiated Fat Cells and Adipose-Derived Stem Cells In Vitro and In Vivo: Application of DFAT Cells Isolated by a Mesh Method. International Journal of Molecular Sciences. 2021; 22(22):12392. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212392
Chicago/Turabian StyleTakabatake, Kiyofumi, Masakazu Matsubara, Eiki Yamachika, Yuki Fujita, Yuki Arimura, Kazuki Nakatsuji, Keisuke Nakano, Histoshi Nagatsuka, and Seiji Iida. 2021. "Comparing the Osteogenic Potential and Bone Regeneration Capacities of Dedifferentiated Fat Cells and Adipose-Derived Stem Cells In Vitro and In Vivo: Application of DFAT Cells Isolated by a Mesh Method" International Journal of Molecular Sciences 22, no. 22: 12392. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212392