
Biochemical Characterisation of Human Transglutaminase 4

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Analysis of Proteomics Datasets for the Expression of hTG4 Protein in the Human Body
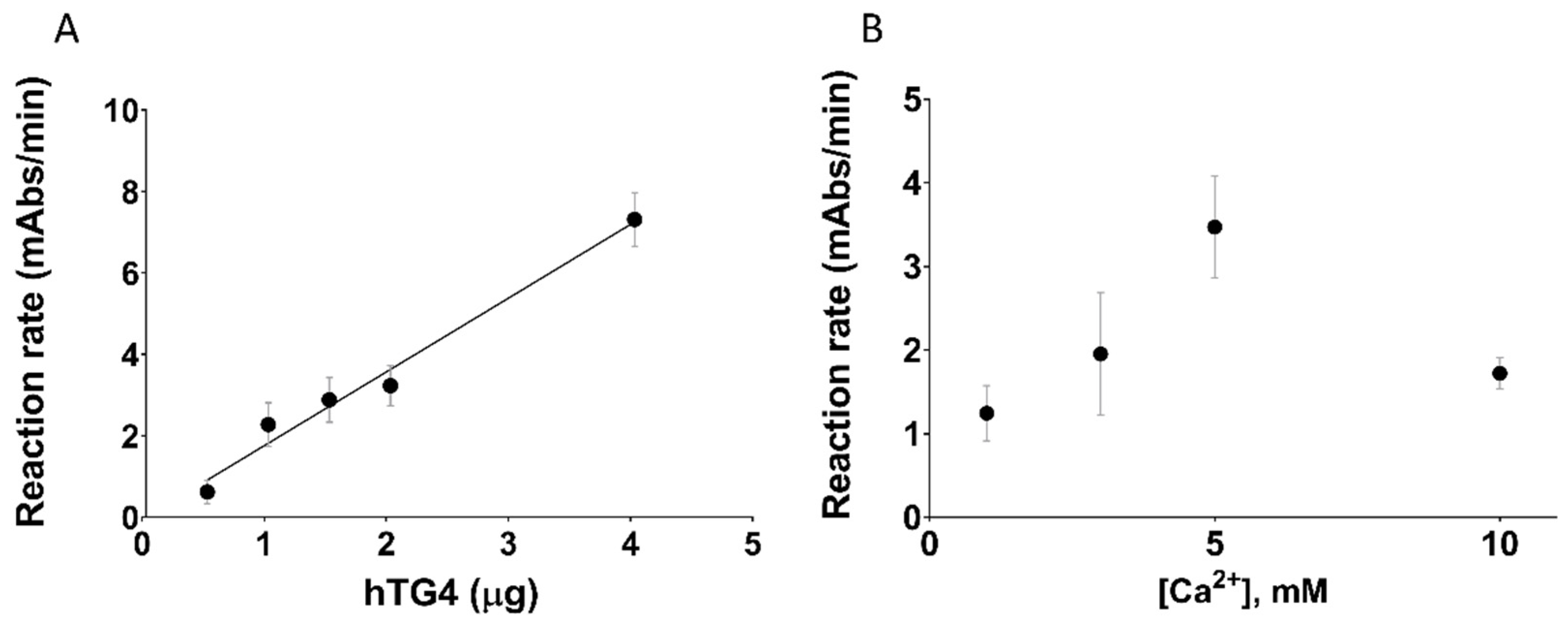
2.2. hTG4 Has Curiously Low Transamidase Activity in the Transglutaminase Family
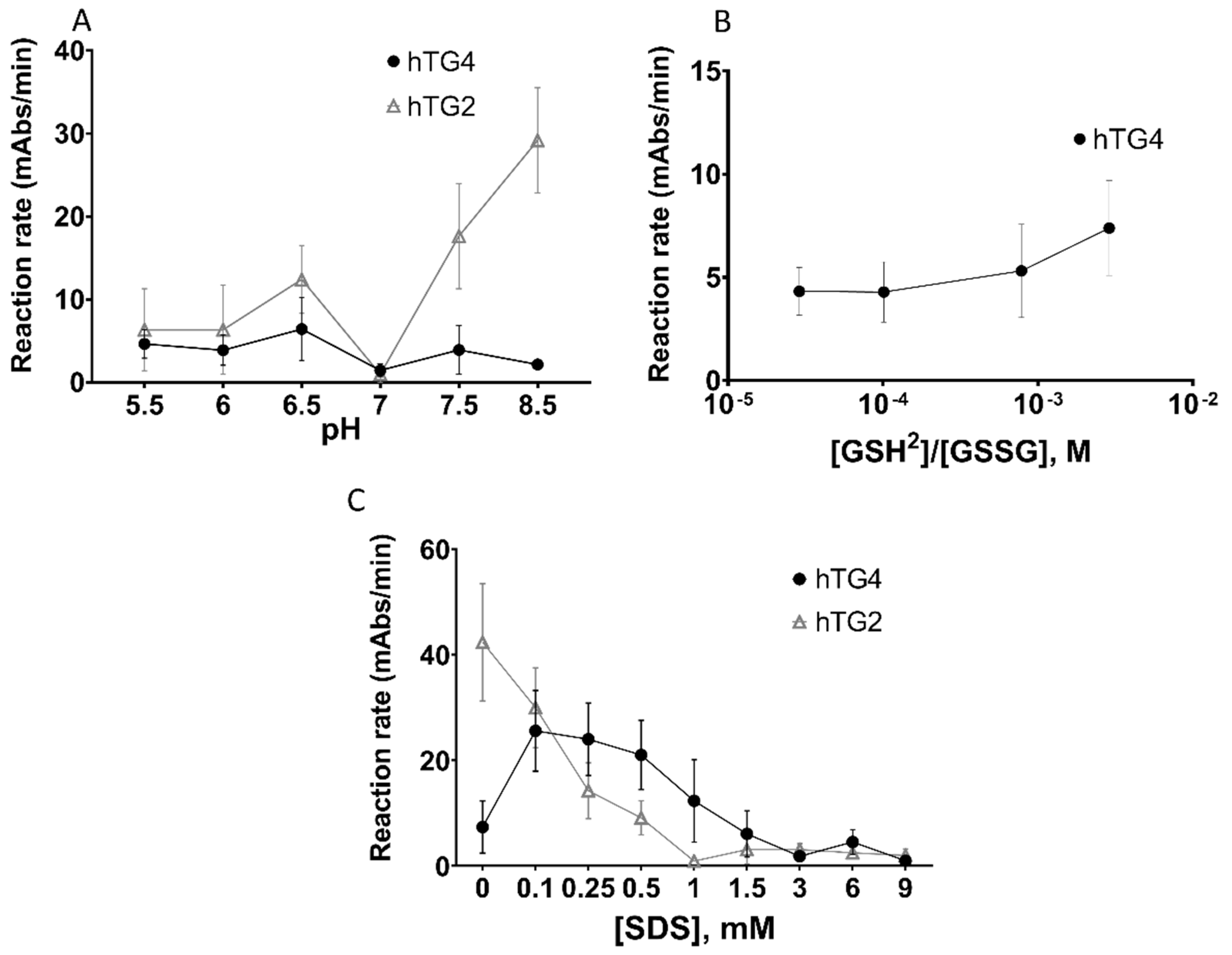
2.3. Effect of pH, Reducing/Oxidising Environment, and Sodium-Dodecyl Sulphate on hTG4 Transamidase Activity
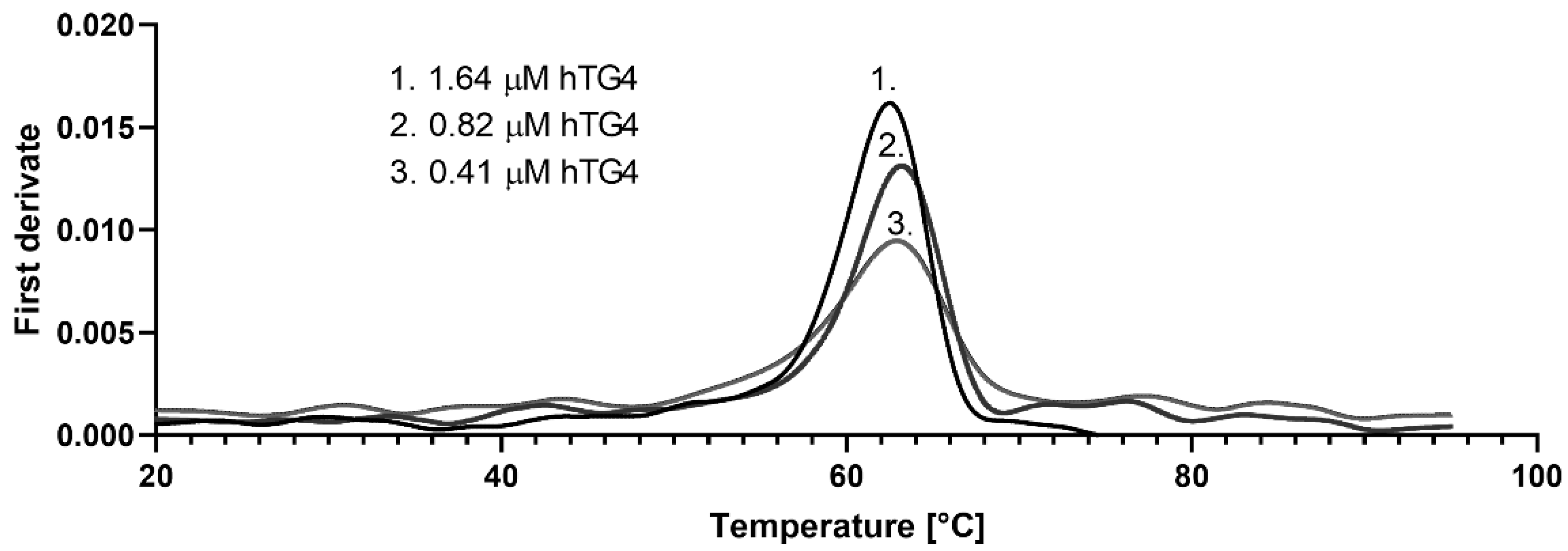
2.4. Thermal Stability of Recombinant hTG4 Protein
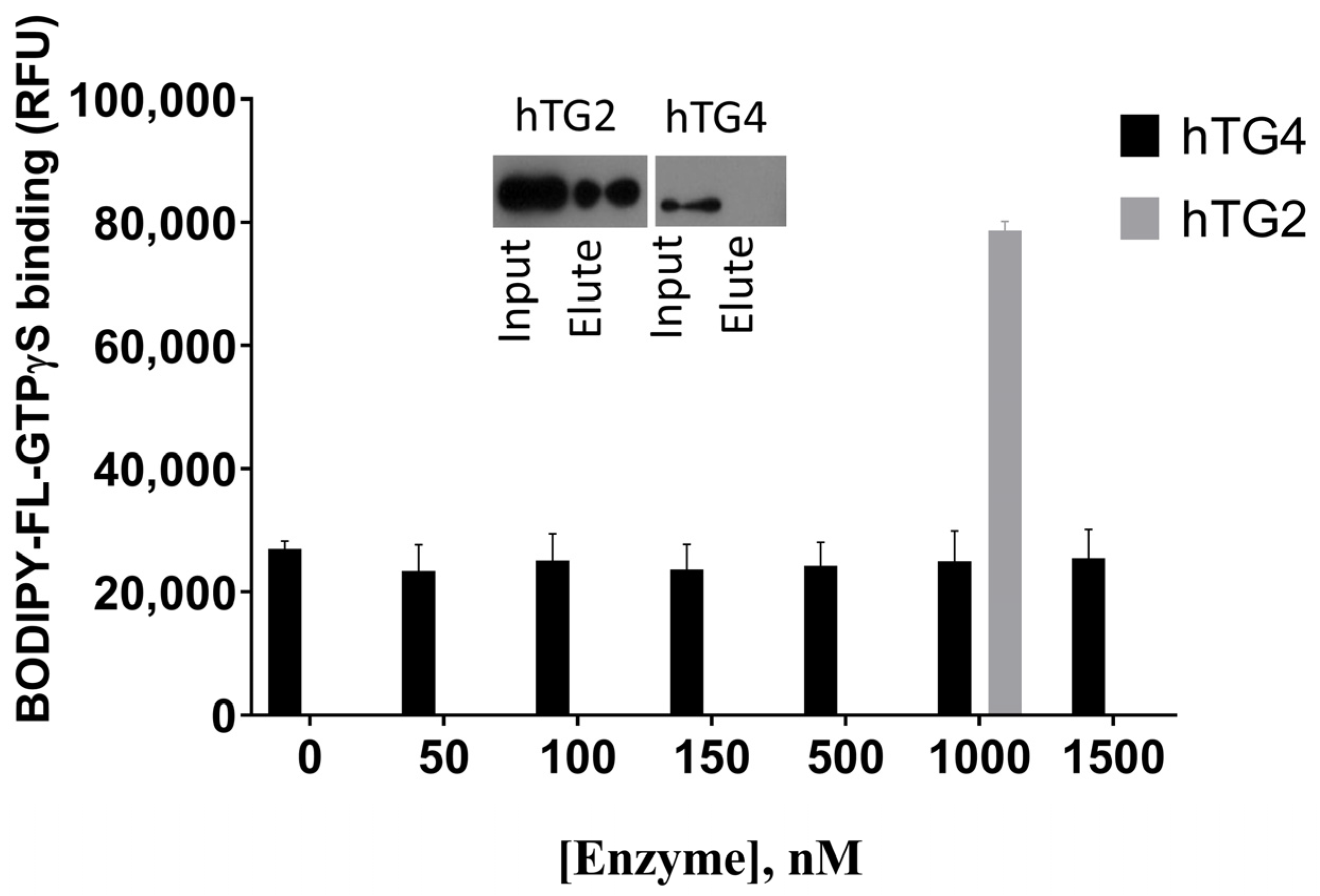
2.5. Human TG4 Does Not Bind GTP
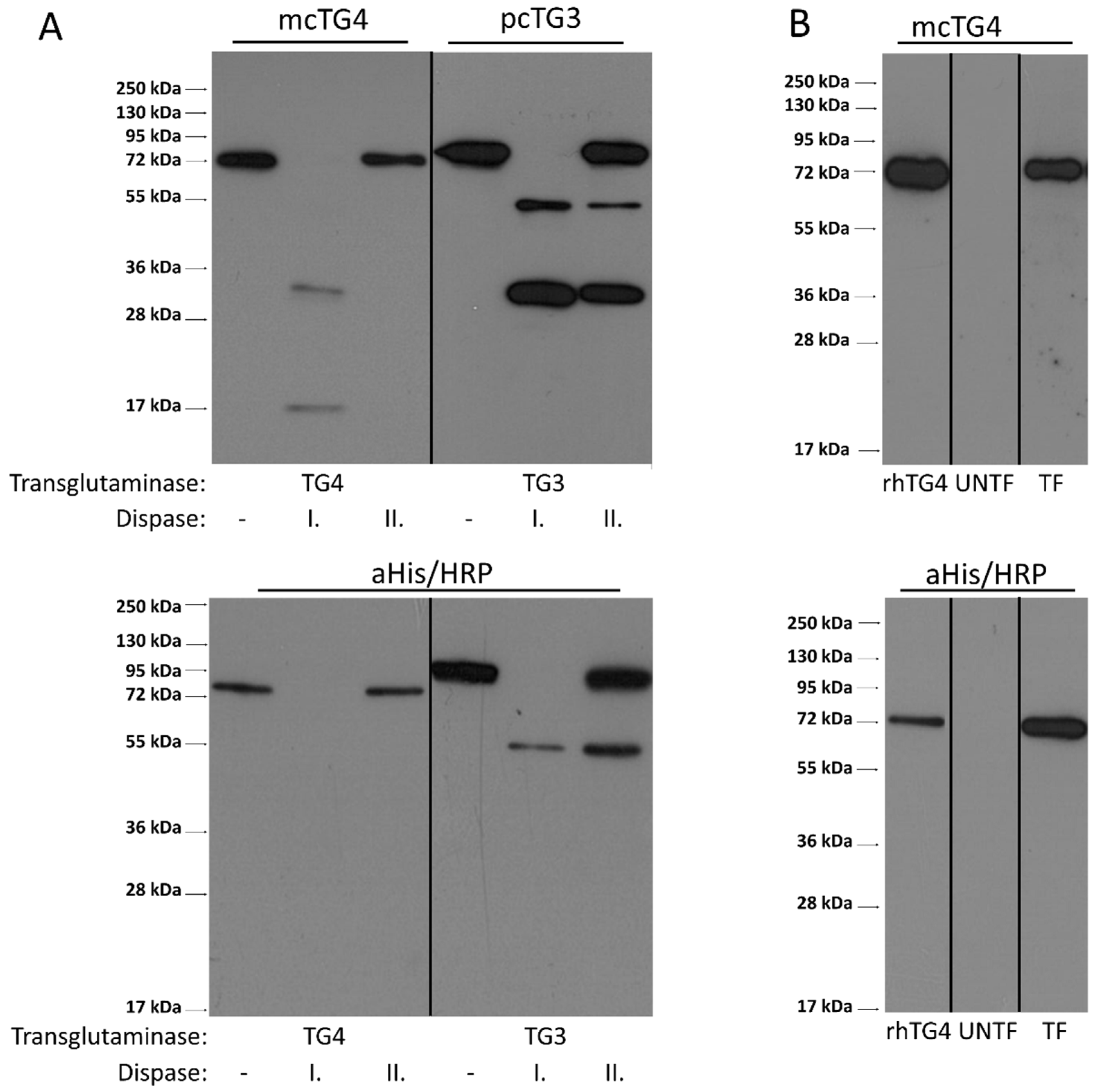
2.6. Limited Proteolysis of hTG4 by Thrombin or Dispase Is Not a Potential Mechanism for Activation of hTG4
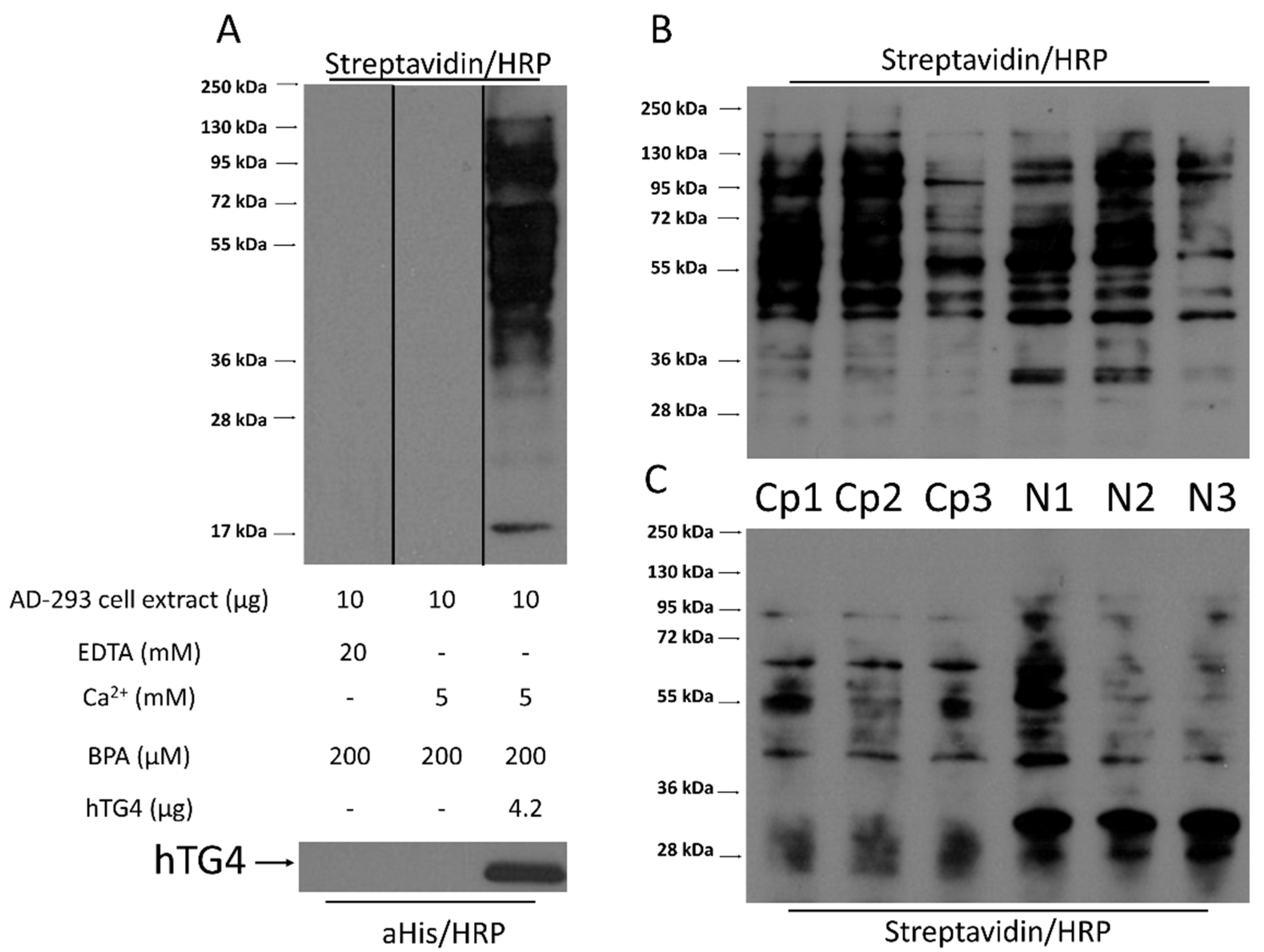
2.7. Human TG4 Incorporates Biotin-Pentylamine into Several Proteins in AD-293 Cell Extract
3. Discussion
4. Materials and Methods
4.1. Proteomics Database Analysis
4.2. Production of Recombinant Human TG4
4.3. Transamidase Activity Assay
4.4. Nucleotide-Binding Assays
4.5. Differential Scanning Fluorimetry
4.6. Dispase Digestion
4.7. Exogenous Expression of hTG4 in AD-293 Cells
4.8. Human TG4 Substrate Search
4.9. Western Blot
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| TG2, 3, 4, 7 | transglutaminase 2, 3, 4, 7, respectively |
| TGp | prostate transglutaminase (TG4) |
| FXIII-A | human blood coagulation factor XIII-A |
| SDS | sodium dodecyl sulphate |
| BPA | biotin-pentylamine |
| GSH/GSSG | reduced/oxidised glutathione |
| nanoDSF | nano differential scanning fluorimetry |
References
- Lorand, L.; Graham, R.M. Transglutaminases: Crosslinking enzymes with pleiotropic functions. Nat. Rev. Mol. Cell Biol. 2003, 4, 140–156. [Google Scholar] [CrossRef]
- Eckert, R.L.; Kaartinen, M.T.; Nurminskaya, M.; Belkin, A.M.; Colak, G.; Johnson, G.V.; Mehta, K. Transglutaminase regulation of cell function. Physiol. Rev. 2014, 94, 383–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muszbek, L.; Bereczky, Z.; Bagoly, Z.; Komáromi, I.; Katona, É. Factor XIII: A coagulation factor with multiple plasmatic and cellular functions. Physiol. Rev. 2011, 91, 931–972. [Google Scholar] [CrossRef] [Green Version]
- Williams-Ashman, H.G. Transglutaminases and the clotting of mammalian seminal fluids. Mol. Cell. Biochem. 1984, 58, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Dean, M.D. Genetic disruption of the copulatory plug in mice leads to severely reduced fertility. PLoS Genet. 2013, 9, e1003185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, D.C.; Agrawal, A.K.; Manjunath, R.; Mukherjee, A.B. Suppression of epididymal sperm antigenicity in the rabbit by uteroglobin and transglutaminase in vitro. Science 1983, 219, 989–991. [Google Scholar] [CrossRef] [PubMed]
- Peluso, G.; Porta, R.; Esposito, C.; Tufano, M.A.; Toraldo, R.; Vuotto, M.L.; Ravagnan, G.; Metafora, S. Suppression of rat epididymal sperm immunogenicity by a seminal vesicle secretory protein and transglutaminase both in vivo and in vitro. Biol. Reprod. 1994, 50, 593–602. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.B.; Petersen-Jones, H.; Thompson, J.M.; Hitomi, K.; Itoh, M.; Bakker, E.N.; Johnson, G.V.; Colak, G.; Watts, S.W. Vena cava and aortic smooth muscle cells express transglutaminases 1 and 4 in addition to transglutaminase 2. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H1355–H1366. [Google Scholar] [CrossRef] [Green Version]
- Seitz, J.; Keppler, C.; Hüntemann, S.; Rausch, U.; Aumüller, G. Purification and molecular characterisation of a secretory transglutaminase from coagulating gland of the rat. Biochim. Biophys. Acta 1991, 1078, 139–146. [Google Scholar] [CrossRef]
- Esposito, C.; Pucci, P.; Amoresano, A.; Marino, G.; Cozzolino, A.; Porta, R. Transglutaminase from rat coagulating gland secretion. Post-translational modifications and activation by phosphatidic acids. J. Biol. Chem. 1996, 271, 27416–27423. [Google Scholar] [CrossRef] [Green Version]
- Spina, A.M.; Esposito, C.; Pagano, M.; Chiosi, E.; Mariniello, L.; Cozzolino, A.; Porta, R.; Illiano, G. GTPase and transglutaminase are associated in the secretion of the rat anterior prostate. Biochem. Biophys. Res. Commun. 1999, 260, 351–356. [Google Scholar] [CrossRef]
- Mariniello, L.; Esposito, C.; Caputo, I.; Sorrentino, A.; Porta, R. N-terminus end of rat prostate transglutaminase is responsible for its catalytic activity and GTP binding. Int. J. Biochem. Cell Biol. 2003, 35, 1098–1108. [Google Scholar] [CrossRef]
- Tseng, H.C.; Lin, H.J.; Gandhi, P.S.S.; Wang, C.Y.; Chen, Y.H. Purification and identification of transglutaminase from mouse coagulating gland and its crosslinking activity among seminal vesicle secretion proteins. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2008, 876, 198–202. [Google Scholar] [CrossRef]
- Tseng, H.C.; Tang, J.B.; Gandhi, P.S.; Luo, C.W.; Ou, C.M.; Tseng, C.J.; Lin, H.J.; Chen, Y.H. Mutual adaptation between mouse transglutaminase 4 and its native substrates in the formation of copulatory plug. Amino Acids 2012, 42, 951–960. [Google Scholar] [CrossRef]
- Peter, A.; Lilja, H.; Lundwall, A.; Malm, J. Semenogelin I and semenogelin II, the major gel-forming proteins in human semen, are substrates for transglutaminase. Eur. J. Biochem. 1998, 252, 216–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porta, R.; Esposito, C.; De Santis, A.; Fusco, A.; Iannone, M.; Metafora, S. Sperm maturation in human semen: Role of transglutaminase-mediated reactions. Biol. Reprod. 1986, 35, 965–970. [Google Scholar] [CrossRef] [Green Version]
- Perez, A.M.; Thomas, V.; Martin, G.; El Alaoui, S. Identification of human salivary transglutaminases. Amino Acids 2013, 44, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Hogan, M.C.; Johnson, K.L.; Zenka, R.M.; Charlesworth, M.C.; Madden, B.J.; Mahoney, D.W.; Oberg, A.L.; Huang, B.Q.; Leontovich, A.A.; Nesbitt, L.L.; et al. Subfractionation, characterisation, and in-depth proteomic analysis of glomerular membrane vesicles in human urine. Kidney Int. 2014, 85, 1225–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.G.; Ablin, R.J.; Kynaston, H.G.; Mason, M.D. The prostate transglutaminase (TGase-4, TGaseP) regulates the interaction of prostate cancer and vascular endothelial cells, a potential role for the ROCK pathway. Microvasc. Res. 2009, 77, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Davies, G.; Ablin, R.J.; Mason, M.D.; Jiang, W.G. Expression of the prostate transglutaminase (TGase-4) in prostate cancer cells and its impact on the invasiveness of prostate cancer. J. Exp. Ther. Oncol. 2007, 6, 257–264. [Google Scholar]
- Landegren, N.; Sharon, D.; Shum, A.K.; Khan, I.S.; Fasano, K.J.; Hallgren, Å.; Kampf, C.; Freyhult, E.; Ardesjö-Lundgren, B.; Alimohammadi, M.; et al. Transglutaminase 4 as a prostate autoantigen in male subfertility. Sci. Transl. Med. 2015, 7, 292ra101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, X.; Pascal, G.; Fouchécourt, S.; Pontarotti, P.; Monget, P. Gene birth, death, and divergence: The different scenarios of reproduction-related gene evolution. Biol. Reprod. 2009, 80, 616–621. [Google Scholar] [CrossRef] [Green Version]
- Thangaraju, K.; Király, R.; Demény, M.A.; András, M.J.; Fuxreiter, M.; Fésüs, L. Genomic variants reveal differential evolutionary constraints on human transglutaminases and point towards unrecognised significance of transglutaminase 2. PLoS ONE 2017, 12, e0172189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, E.W.; Csordas, A.; Sun, Z.; Jarnuczak, A.; Perez-Riverol, Y.; Ternent, T.; Campbell, D.S.; Bernal-Llinares, M.; Okuda, S.; Kawano, S.; et al. The ProteomeXchange consortium in 2017: Supporting the cultural change in proteomics public data deposition. Nucleic Acids Res. 2017, 45, D1100–D1106. [Google Scholar] [CrossRef]
- Desiere, F.; Deutsch, E.W.; King, N.L.; Nesvizhskii, A.I.; Mallick, P.; Eng, J.; Chen, S.; Eddes, J.; Loevenich, S.N.; Aebersold, R. The PeptideAtlas project. Nucleic Acids Res. 2006, 34, D655–D658. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, M.; Schlegl, J.; Hahne, H.; Gholami, A.M.; Lieberenz, M.; Savitski, M.M.; Ziegler, E.; Butzmann, L.; Gessulat, S.; Marx, H.; et al. Mass-spectrometry-based draft of the human proteome. Nature 2014, 509, 582–587. [Google Scholar] [CrossRef]
- Stamnaes, J.; Pinkas, D.M.; Fleckenstein, B.; Khosla, C.; Sollid, L.M. Redox regulation of transglutaminase 2 activity. J. Biol. Chem. 2010, 285, 25402–25409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietroni, V.; Di Giorgi, S.; Paradisi, A.; Ahvazi, B.; Candi, E.; Melino, G. Inactive and highly active, proteolytically processed transglutaminase-5 in epithelial cells. J. Investig. Dermatol. 2008, 128, 2760–2766. [Google Scholar] [CrossRef]
- Rivera-Gonzalez, G.C.; Droop, A.P.; Rippon, H.J.; Tiemann, K.; Pellacani, D.; Georgopoulos, L.J.; Maitland, N.J. Retinoic acid and androgen receptors combine to achieve tissue specific control of human prostatic transglutaminase expression: A novel regulatory network with broader significance. Nucleic Acids Res. 2012, 40, 4825–4840. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Bujanda, Z.A.; Obradovic, A.; Nirschl, T.R.; Crowley, L.; Macedo, R.; Papachristodoulou, A.; O’Donnell, T.; Laserson, U.; Zarif, J.C.; Reshef, R.; et al. TGM4: An immunogenic prostate-restricted antigen. J. Immunother. Cancer 2021, 9, e001649. [Google Scholar] [CrossRef]
- Drabovich, A.P.; Saraon, P.; Drabovich, M.; Karakosta, T.D.; Dimitromanolakis, A.; Hyndman, M.E.; Jarvi, K.; Diamandis, E.P. Multi-omics Biomarker Pipeline Reveals Elevated Levels of Protein-glutamine Gamma-glutamyltransferase 4 in Seminal Plasma of Prostate Cancer Patients. Mol. Cell Proteom. 2019, 18, 1807–1823. [Google Scholar] [CrossRef] [PubMed]
- Moore, B.M.; Flurkey, W.H. Sodium dodecyl sulfate activation of a plant polyphenoloxidase. Effect of sodium dodecyl sulfate on enzymatic and physical characteristics of purified broad bean polyphenoloxidase. J. Biol. Chem. 1990, 265, 4982–4988. [Google Scholar] [CrossRef]
- Bhuyan, A.K. On the mechanism of SDS-induced protein denaturation. Biopolymers 2010, 93, 186–199. [Google Scholar] [CrossRef]
- Carnahan, S.J.; Jensen-Seaman, M.I. Hominoid seminal protein evolution and ancestral mating behavior. Am. J. Primatol. 2008, 70, 939–948. [Google Scholar] [CrossRef]
- Liu, S.; Cerione, R.A.; Clardy, J. Structural basis for the guanine nucleotide-binding activity of tissue transglutaminase and its regulation of transamidation activity. Proc. Natl. Acad. Sci. USA 2002, 99, 2743–2747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.G.; Ablin, R.J. Prostate transglutaminase: A unique transglutaminase and its role in prostate cancer. Biomark. Med. 2011, 5, 285–291. [Google Scholar] [CrossRef]
- Kim, S.Y.; Chung, S.I.; Steinert, P.M. Highly active soluble processed forms of the transglutaminase 1 enzyme in epidermal keratinocytes. J. Biol. Chem. 1995, 270, 18026–18035. [Google Scholar] [CrossRef] [Green Version]
- Hitomi, K.; Yamagiwa, Y.; Ikura, K.; Yamanishi, K.; Maki, M. Characterization of human recombinant transglutaminase 1 purified from baculovirus-infected insect cells. Biosci. Biotechnol. Biochem. 2000, 64, 2128–2137. [Google Scholar] [CrossRef]
- Cheng, T.; Hitomi, K.; van Vlijmen-Willems, I.M.; de Jongh, G.J.; Yamamoto, K.; Nishi, K.; Watts, C.; Reinheckel, T.; Schalkwijk, J.; Zeeuwen, P.L. Cystatin M/E is a high affinity inhibitor of cathepsin V and cathepsin L by a reactive site that is distinct from the legumain-binding site. A novel clue for the role of cystatin M/E in epidermal cornification. J. Biol. Chem. 2006, 281, 15893–15899. [Google Scholar] [CrossRef] [Green Version]
- Hitomi, K.; Kanehiro, S.; Ikura, K.; Maki, M. Characterization of recombinant mouse epidermal-type transglutaminase (TGase 3): Regulation of its activity by proteolysis and guanine nucleotides. J. Biochem. 1999, 125, 1048–1054. [Google Scholar] [CrossRef] [PubMed]
- Kanchan, K.; Ergülen, E.; Király, R.; Simon-Vecsei, Z.; Fuxreiter, M.; Fésüs, L. Identification of a specific one amino acid change in recombinant human transglutaminase 2 that regulates its activity and calcium sensitivity. Biochem. J. 2013, 455, 261–272. [Google Scholar] [CrossRef] [Green Version]
- Csosz, E.; Meskó, B.; Fésüs, L. Transdab wiki: The interactive transglutaminase substrate database on web 2.0 surface. Amino Acids 2009, 36, 615–617. [Google Scholar] [CrossRef]
- Tseng, H.C.; Lin, H.J.; Tang, J.B.; Gandhi, P.S.; Chang, W.C.; Chen, Y.H. Identification of the major TG4 crosslinking sites in the androgen-dependent SVS I exclusively expressed in mouse seminal vesicle. J. Cell. Biochem. 2009, 107, 899–907. [Google Scholar] [CrossRef]
- Yu, W.; Yang, L.; Li, T.; Zhang, Y. Cadherin Signaling in Cancer: Its Functions and Role as a Therapeutic Target. Front. Oncol. 2019, 9, 989. [Google Scholar] [CrossRef] [PubMed]
- Biri, B.; Kiss, B.; Király, R.; Schlosser, G.; Láng, O.; Kőhidai, L.; Fésüs, L.; Nyitray, L. Metastasis-associated S100A4 is a specific amine donor and an activity-independent binding partner of transglutaminase-2. Biochem. J. 2016, 473, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Slaughter, T.F.; Achyuthan, K.E.; Lai, T.S.; Greenberg, C.S. A microtiter plate transglutaminase assay utilising 5-(biotinamido)pentylamine as substrate. Anal. Biochem. 1992, 205, 166–171. [Google Scholar] [CrossRef]
- Ruan, Q.; Tucholski, J.; Gundemir, S.; Johnson, V.G.V. The Differential Effects of R580A Mutation on Transamidation and GTP Binding Activity of Rat and Human Type 2 Transglutaminase. Int. J. Clin. Exp. Med. 2008, 1, 248–259. [Google Scholar]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualised p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
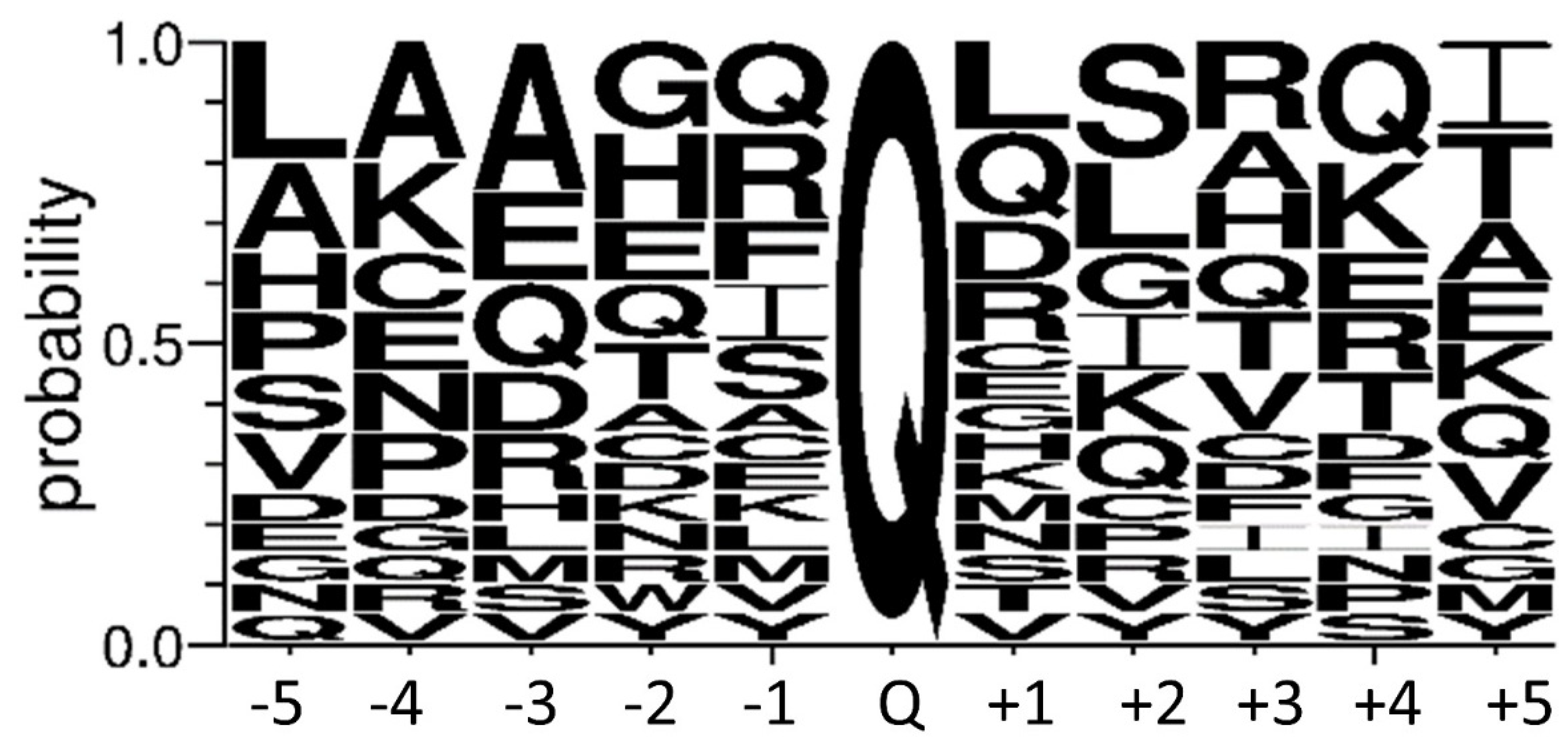{kind=link}
| Tissue | MassIVE | PeptideAtlas | ProteomicsDB |
|---|---|---|---|
| Colon | ++ | ++ | + |
| Heart | + | ++ | ++ |
| Prostate | ++ | ++ | ++ |
| Salivary gland | − | − | ++ |
| Seminal plasma | ++ | ++ | ++ |
| Seminal vesicle | ++ | ++ | ++ |
| Spermatozoon | ++ | ++ | − |
| Spleen | − | + | ++ |
| Urinary bladder | ++ | ++ | ++ |
| Human Transglutaminase | Article-No. | Transamidase Activity (U/mg) | Expression Host |
|---|---|---|---|
| TG1 | T009 | ~2500 | E. coli |
| TG2 | T001 | ~750 | E. coli |
| TG3 | T024 | ~1000 | Insect cells |
| TG4 | T042 | ~30 | E. coli |
| TG7 | T011 | ~1000 | E. coli |
| Accession | Identified Protein | Cellular Localisation | Total Spectrum Count | Reactive Q ± 5 AA | |
|---|---|---|---|---|---|
| 1. | B4GT5 | Beta-1,4-galactosyltranseferase 5 | Golgi apparatus | 13 | AQVYEQVLRSA |
| 2. | AGRL3 | Adhesion G protein-coupled receptor L3 | Cell membrane | 2 | HGSTIQLSANT |
| 3. | ENO4 | Enolase 4 | Cytosol | 4 | SKRGQQQITGK |
| 4. | CMYA5 | Cardiomyopathy-associated protein 5 | Cytoplasm, Nucleus | 4 | SDLGRQSGSIG |
| 5. | CHD1 | Chromodomain-helicase-DNA-binding protein 1 | Cytoplasm, Nucleus | 1 | LEHTRQCLIKI |
| 6. | SAM15 | Sterile alpha motif domain-containing protein 15 | n.a. | 1 | VPEEMQRKATE |
| 7. | GRIN3 | G protein-regulated inducer of neurite outgrowth 3 | Plasma membrane | 2 | LPAQRQMSRFK |
| 8. | K2C79 | Keratin, type II cytoskeletal 79 | Cytoskeleton, Cytosole, Extracellular exosome | 1 | AEAWYQTKYEE |
| 9. | PCDH17 | Protocadherin-17 | Cell membrane | 1 | HNAKCQLSLEV |
| 10. | F10A5 | Putative protein FAM 10A5 | Cytoplasm | 1 | PNAAIQDCDRA |
| 11. | PCDHB3 | Protocadherin beta-3 | Cell membrane | 1 | NKQHFQLSHQT |
| 12. | STAB2 | Stabilin-2 | Cytoplasm, Cell membrane | 1 | VARCSQKGTKV |
| 13. | PLXA2 | Plexin-A2 | Cell membrane | 1 | AVDGKQDYFPT |
| 14. | F155A | Transmembrane protein FAM155A | Membrane | 1 | DKEHQQQQRQQ |
| QRQQQQQQQQQ | |||||
| 15. | ALKB8 | Alkylated DNA repair protein alkB homolog 8 | Nucleus, Cytoplasm | 1 | GCDRSQNLVDI |
| 16. | IL6RA | Interleukin-6 receptor subunit alpha | Cell membrane, Secreted | 1 | PAEDFQEPCQY |
| 17. | ZN363 | RING finger and CHY zinc finger domain-containing protein 1 | Cytoplasm, Nucleus | 1 | LAMNLQGRHKC |
| 18. | FAM184A | Protein FAM184A | ECM | 1 | LCAEAQHVQRI |
| EAQHVQRIVTM |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Csobán-Szabó, Z.; Bécsi, B.; El Alaoui, S.; Fésüs, L.; Korponay-Szabó, I.R.; Király, R. Biochemical Characterisation of Human Transglutaminase 4. Int. J. Mol. Sci. 2021, 22, 12448. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212448
Csobán-Szabó Z, Bécsi B, El Alaoui S, Fésüs L, Korponay-Szabó IR, Király R. Biochemical Characterisation of Human Transglutaminase 4. International Journal of Molecular Sciences. 2021; 22(22):12448. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212448
Chicago/Turabian StyleCsobán-Szabó, Zsuzsa, Bálint Bécsi, Saïd El Alaoui, László Fésüs, Ilma Rita Korponay-Szabó, and Róbert Király. 2021. "Biochemical Characterisation of Human Transglutaminase 4" International Journal of Molecular Sciences 22, no. 22: 12448. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212448