
Genome Features of a New Double-Stranded RNA Helper Virus (LBCbarr) from Wine Torulaspora delbrueckii Killer Strains



Abstract
:1. Introduction
2. Results
2.1. Analysis of the dsRNA Genomes from TdV-LBCbarr1 and TdV-LBCbarr2
2.2. Analysis of the Gag-Pol Sequences from TdV-LBCbarr1 and TdV-LBCbarr2
2.3. Comparison of TdV-LBCbarr with LBC Viruses from Saccharomyces Yeasts
3. Discussion
3.1. Analysis of TdV-LBCbarr Genomes
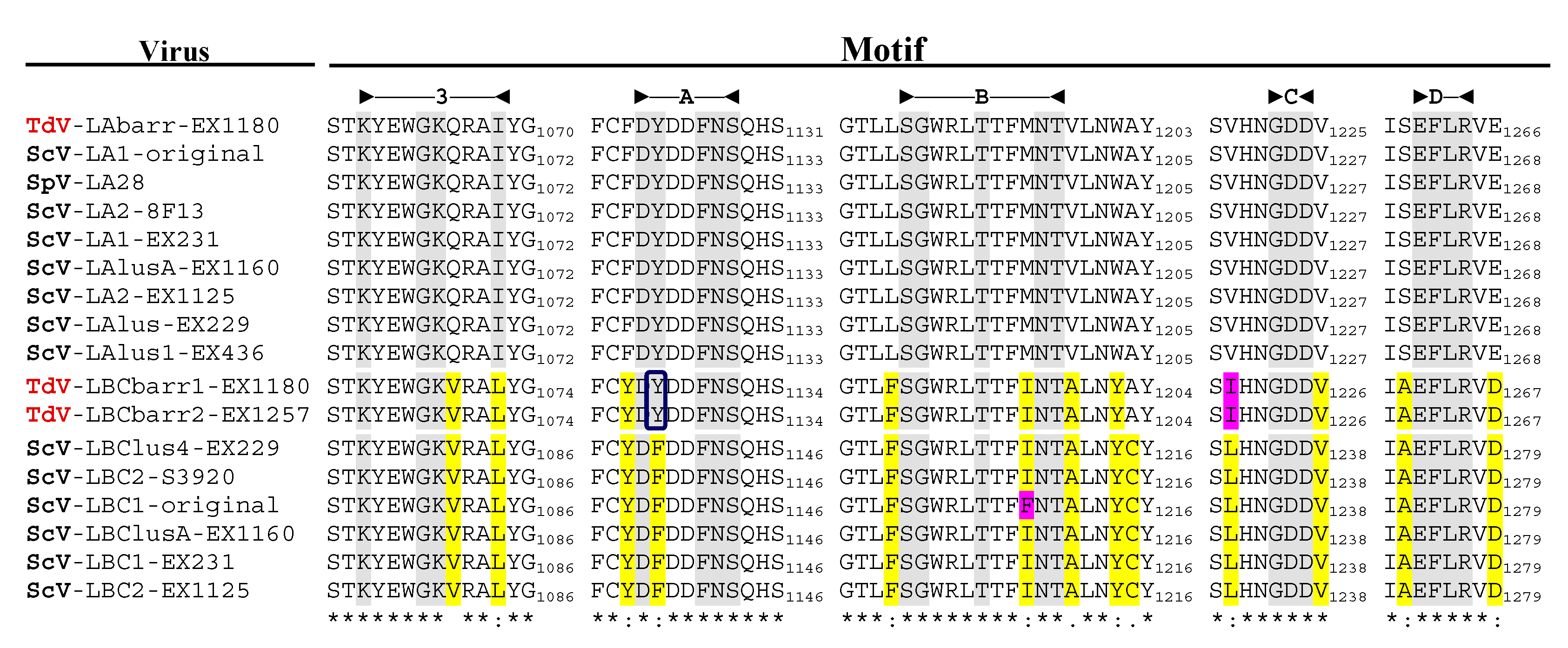
3.2. Analysis of TdV-LBCbarr Gag-Pol Sequences
3.3. Phylogenetic Relationship and Transmission of LBC Viruses
4. Materials and Methods
4.1. Yeast Strains and Culture Media
4.2. Purification of V-LBC dsRNA from Killer Yeasts
4.3. Preparation of cDNA Libraries from Purified V-LBC dsRNA and DNA Sequencing
4.4. Sequence Assembly of Virus Genomes
4.5. Sequence Analysis Tools
4.6. Nucleotide Sequence Accession Numbers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schmitt, M.J.; Breinig, F. Yeast viral killer toxins: Lethality and self-protection. Nat. Rev. Microbiol. 2006, 4, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Wickner, R.B.; Fujimura, T.; Esteban, R. Viruses and prions of Saccharomyces cerevisiae. Adv. Virus Res. 2013, 86, 1–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez, M.; Velázquez, R.; López-Pineiro, A.; Naranjo, B.; Roig, F.; Llorens, C. New Insights into the genome organization of yeast killer viruses based on “atypical” killer strains characterized by high-throughput sequencing. Toxins 2017, 9, 292. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Cousiño, N.; Esteban, R. Relationships and evolution of double-stranded RNA Totiviruses of yeasts inferred from analysis of L-A-2 and L-BC variants in wine yeast strain populations. Appl. Environ. Microbiol. 2017, 83, e02991-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Cousiño, N.; Maqueda, M.; Ambrona, J.; Zamora, E.; Esteban, E.; Ramírez, M. A new wine Saccharomyces cerevisiae double-stranded RNA virus encoded killer toxin (Klus) with broad antifungal activity is evolutionarily related to a chromosomal host gene. Appl. Environ. Microbiol. 2011, 77, 1822–1832. [Google Scholar] [CrossRef] [Green Version]
- Vepštaitė-Monstavičė, I.; Lukša, J.; Konovalovas, A.; Ežerskytė, D.; Stanevičienė, R.; Strazdaitė-Žielienė, Ž.; Serva, S.; Servienė, E. Saccharomyces paradoxus K66 killer system evidences expanded assortment of helper and satellite viruses. Viruses 2018, 10, 564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.M.; Lopinski, J.D.; Masuda, J.; Tzeng, T.H.; Bruen, J.A. A second double-stranded RNA virus from yeast. Virology 1996, 216, 451–454. [Google Scholar] [CrossRef]
- Wickner, R.B. Double-stranded RNA viruses of Saccharomyces cerevisiae. Microbiol. Rev. 1996, 60, 250–265. [Google Scholar] [CrossRef]
- Dinman, J.D.; Wickner, R.B. Ribosomal frameshifting efficiency and gag/gag-pol ratio are critical for yeast M1 double-stranded RNA virus propagation. J. Virol. 1992, 66, 3669–3676. [Google Scholar] [CrossRef] [Green Version]
- Fujimura, T.; Ribas, J.C.; Makhov, A.M.; Wickner, R.B. Pol of gag-pol fusion protein required for encapsidation of viral RNA of yeast L-A virus. Nature 1992, 359, 746–749. [Google Scholar] [CrossRef]
- Icho, T.; Wickner, R.B. The double-stranded RNA genome of yeast virus L-A encodes its own putative RNA polymerase by fusing two open reading frames. J. Biol. Chem. 1989, 264, 6716–6723. [Google Scholar] [CrossRef]
- Ramírez, M.; Velázquez, R.; Maqueda, M.; Martínez, A. Genome organization of a new double-stranded RNA LA helper virus from wine Torulaspora delbrueckii killer yeast as compared with its Saccharomyces counterparts. Front. Microbiol. 2020, 11, 1–17. [Google Scholar] [CrossRef]
- Rodríguez-Cousiño, N.; Gómez, P.; Esteban, R. L-A-lus, a new variant of the L-A Totivirus found in wine yeasts with Klus killer toxin-encoding Mlus double-stranded RNA: Possible role of killer toxin-encoding satellite RNAs in the evolution of their helper viruses. Appl. Environ. Microbiol. 2013, 79, 4661–4674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimura, T.; Esteban, R.; Esteban, L.M.; Wickner, R.B. Portable encapsidation signal of the L-A double-stranded-RNA virus of Saccharomyces cerevisiae. Cell 1990, 62, 819–828. [Google Scholar] [CrossRef]
- Ramírez, M.; Velázquez, R.; Maqueda, M.; López-Piñeiro, A.; Ribas, J.C. A new wine Torulaspora delbrueckii killer strain with broad antifungal activity and its toxin-encoding double-stranded RNA virus. Front. Microbiol. 2015, 6, 983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Cousiño, N.; Gómez, P.; Esteban, R. Variation and distribution of L-A helper Totiviruses in Saccharomyces sensu stricto yeasts producing different killer toxins. Toxins 2017, 9, 313. [Google Scholar] [CrossRef]
- Thiele, D.J.; Leibowitz, M.J. Structural and functional analysis of separated strands of killer double-stranded RNA of yeast. Nucleic Acids Res. 1982, 10, 6903–6918. [Google Scholar] [CrossRef] [Green Version]
- Field, L.J.; Bobek, L.A.; Brennan, V.E.; Reilly, J.D.; Bruenn, J.A. There are at least two yeast viral double-stranded RNAs of the same size: An explanation for viral exclusion. Cell 1982, 31, 193–200. [Google Scholar] [CrossRef]
- Sommer, S.S.; Wickner, R.B. Yeast L dsRNA consists of at least three distinct RNA’s; evidence that the non-Mendelian genes [HOK], [NEX] and [EXL] are on one of these dsRNA’s. Cell 1982, 31, 429–441. [Google Scholar] [CrossRef]
- Esteban, R.; Fujimura, T.; Wickner, R.B. Internal and terminal cis-acting sites are necessary for in vitro replication of the L-A double-stranded RNA virus of yeast. EMBO J. 1989, 8, 947–954. [Google Scholar] [CrossRef]
- Ribas, J.C.; Wickner, R.B. Saccharomyces cerevisiae L-BC double-stranded RNA virus replicase recognizes the L-A positive-strand RNA end. J. Virol. 1996, 70, 292–297. [Google Scholar] [CrossRef] [Green Version]
- Thiele, D.J.; Wang, R.W.; Leibowitz, M.J. Separation and sequence of the 3′ termini of M double-stranded RNA from killer yeast. Nucleic Acids Res. 1982, 10, 1661–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribas, J.C.; Wickner, R.B. RNA-dependent RNA polymerase consensus sequence of the L-A double-stranded RNA virus: Definition of essential domains. Proc. Natl. Acad. Sci. USA 1992, 89, 2185–2189. [Google Scholar] [CrossRef] [Green Version]
- Routhier, E.; Bruenn, J.A. Functions of conserved motifs in the RNA-dependent RNA polymerase of a yeast double-stranded RNA virus. J. Virol. 1998, 72, 4427–4429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimura, T.; Esteban, R. Cap-snatching mechanism in yeast L-A double-stranded RNA virus. Proc. Natl. Acad. Sci. USA 2011, 108, 17667–17671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimura, T.; Esteban, R. Cap snatching in yeast L-BC double-stranded RNA totivirus. J. Biol. Chem. 2013, 288, 23716–23724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konovalovas, A.; Serviené, E.; Serva, S. Genome sequence of Saccharomyces cerevisiae double-stranded RNA Virus L-A-28. Genome Announc. 2016, 4, e00549-16. [Google Scholar] [CrossRef] [Green Version]
- Kurtzman, C.P. Torulaspora Lindner (1904). In The Yeasts: A Taxonomic Study, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: London, UK, 2011; Volume 2, pp. 867–874. [Google Scholar]
- Kurtzman, C.P. Saccharomyces Meyen ex Reess (1870). In The Yeasts: A Taxonomic Study, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: London, UK, 2011; Volume 2, pp. 733–763. [Google Scholar]
- Dinman, J.D.; Icho, T.; Wickner, R.B. A -1 ribosomal frameshift in a double-stranded RNA virus of yeast forms a gag-pol fusion protein. Proc. Natl. Acad. Sci. USA 1991, 88, 174–178. [Google Scholar] [CrossRef] [Green Version]
- Tzeng, T.H.; Tu, C.L.; Bruenn, J.A. Ribosomal frameshifting requires a pseudoknot in the Saccharomyces cerevisiae double-stranded RNA virus. J. Virol. 1992, 66, 999–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maqueda, M.; Zamora, E.; Álvarez, M.L.; Ramírez, M. Characterization, ecological distribution, and population dynamics of Saccharomyces “sensu stricto” killer yeasts in the spontaneous grape-must fermentations of south-western Spain. Appl. Environ. Microbiol. 2012, 78, 735–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guthrie, C.; Fink, G.R. Guide to yeast genetics and molecular biology. Methods Enzymol. 1991, 194, 3–57. [Google Scholar]
- Maqueda, M.; Zamora, E.; Rodríguez-Cousiño, N.; Ramírez, M. Wine yeast molecular typing using a simplified method for simultaneously extracting mtDNA, nuclear DNA and virus dsRNA. Food Microbiol. 2010, 27, 205–209. [Google Scholar] [CrossRef]
- Toh-e, A.; Guerry, P.; Wickner, R.B. Chromosomal superkiller mutants of Saccharomyces cerevisiae. J. Bacteriol. 1978, 136, 1002–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y.; et al. SOAPdenovo2: An empirically improved memory-efficient short-read de novo assembler. Gigascience 2012, 1, 18–24. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Futami, R.; Muñoz-Pomer, L.; Dominguez-Escriba, L.; Covelli, L.; Bernet, G.P.; Sempere, J.M. GPRO The professional tool for annotation, management and functional analysis of omic databases. Biotechvana Bioinform. 2011, 2011, SOFT3. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Zuker, M.; Mathews, D.H.; Turner, D.H.; Barciszewski, J.; Clark, B.F.C. Algorithms and thermodynamics for RNA secondary structure prediction: A practical guide. In RNA Biochemistry and Biotechnology; Kluwer Academic Publishers: Amsterdam, The Netherlands, 1999. [Google Scholar]
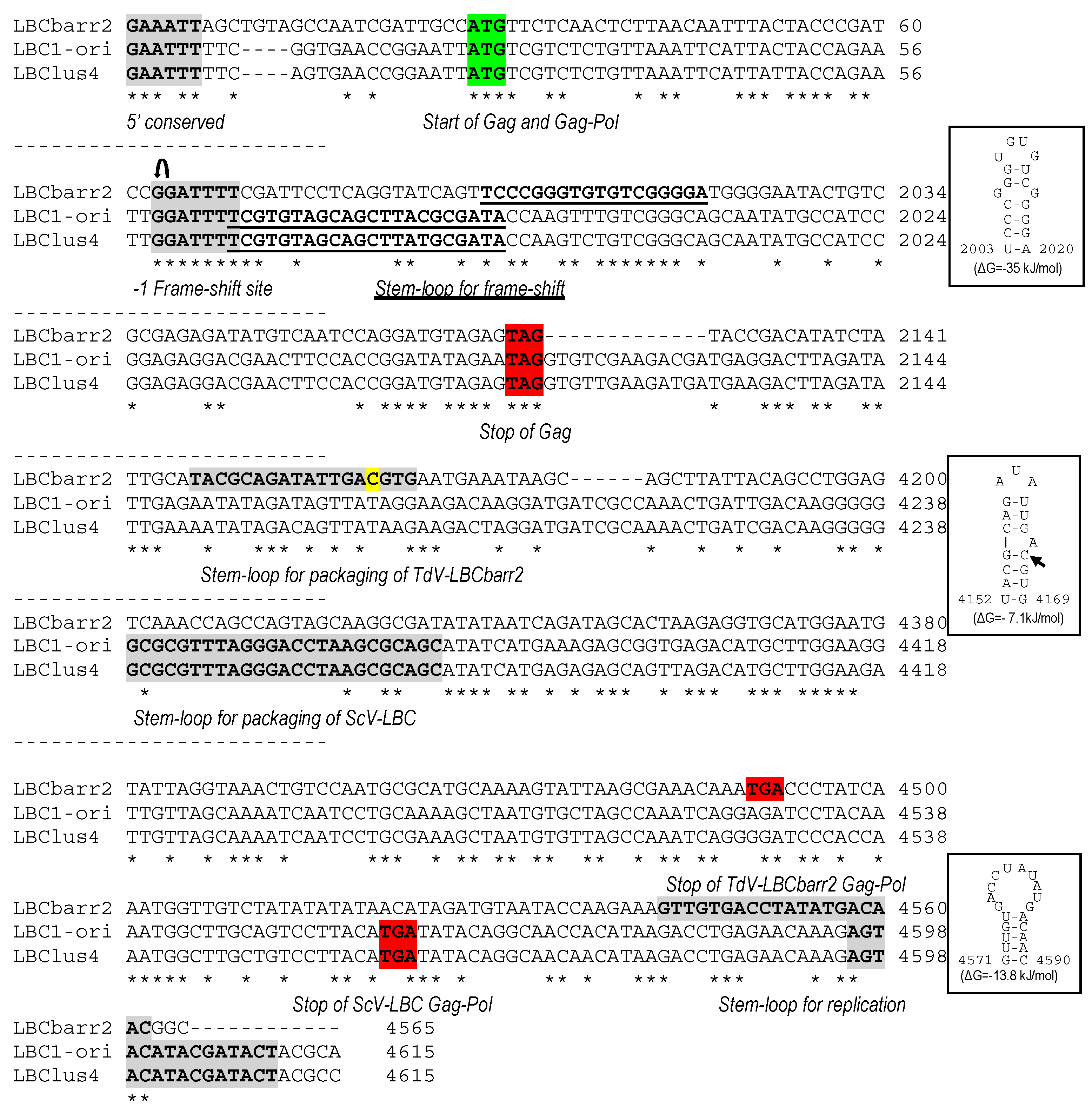
 , ribosomal frameshift. Asterisks (*) indicate identical nucleotide positions. The secondary structures of the putative cis signals for frameshifting, packaging, and replication of TdV-LBCbarr2 are displayed at the right of the sequence panel.
, ribosomal frameshift. Asterisks (*) indicate identical nucleotide positions. The secondary structures of the putative cis signals for frameshifting, packaging, and replication of TdV-LBCbarr2 are displayed at the right of the sequence panel.
, ribosomal frameshift. Asterisks (*) indicate identical nucleotide positions. The secondary structures of the putative cis signals for frameshifting, packaging, and replication of TdV-LBCbarr2 are displayed at the right of the sequence panel.
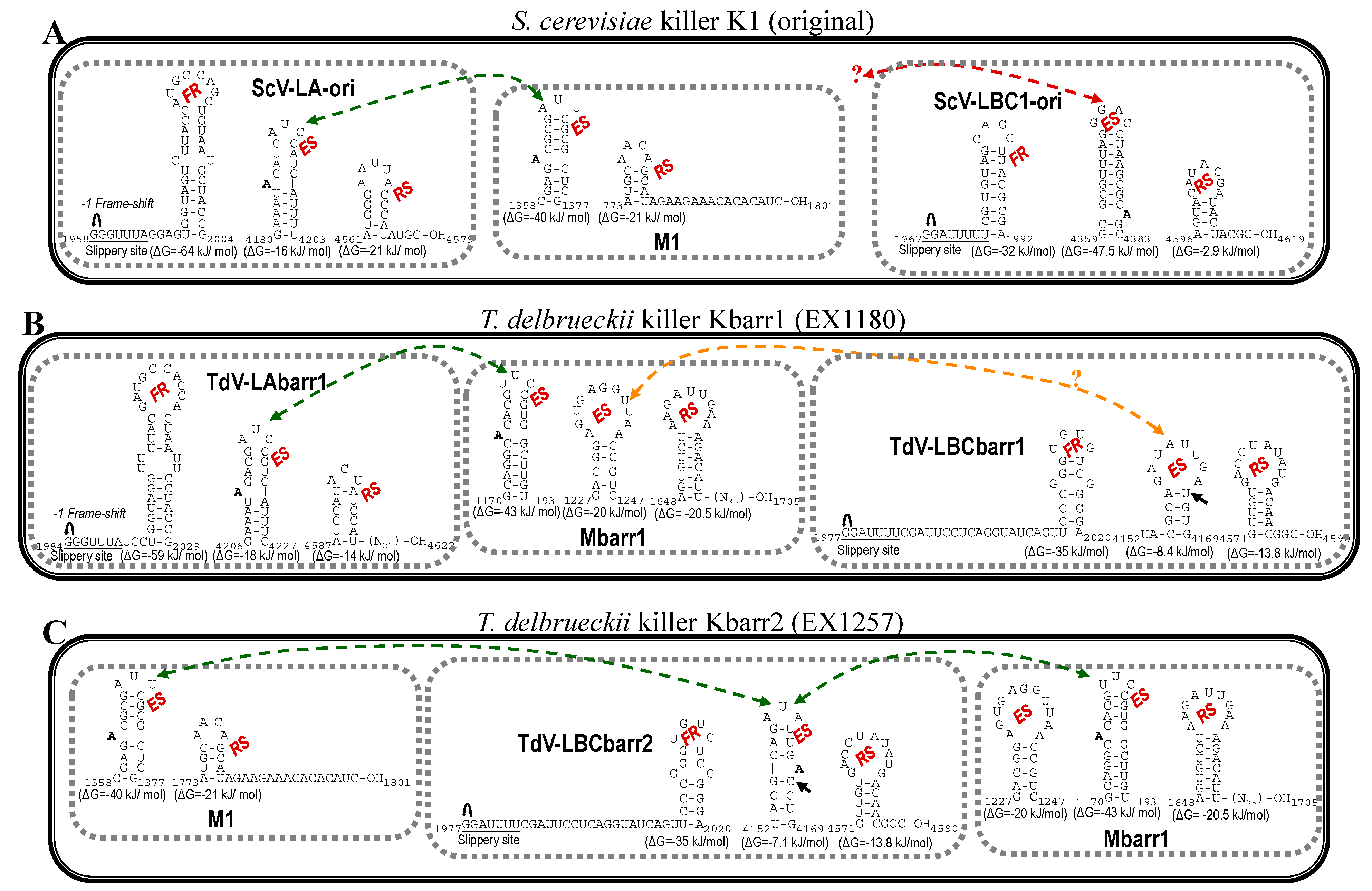
, ribosomal frameshift. Asterisks (*) indicate identical nucleotide positions. The secondary structures of the putative cis signals for frameshifting, packaging, and replication of TdV-LBCbarr2 are displayed at the right of the sequence panel. , ribosomal frameshift. Black arrow indicates the single nucleotide changes between ES of LBCbarr1 and LBCbarr2. Green dot-line arrow indicates equivalent ES for L and M viruses in the same yeast. Orange dot-line arrow indicates possible equivalent ES for LBCbarr1 and Mbarr1 in Kbarr1 EX1180 yeast. Red dot-line arrow indicates unknown equivalent ES in M viruses for the ES of ScV-LBC1-original.
, ribosomal frameshift. Black arrow indicates the single nucleotide changes between ES of LBCbarr1 and LBCbarr2. Green dot-line arrow indicates equivalent ES for L and M viruses in the same yeast. Orange dot-line arrow indicates possible equivalent ES for LBCbarr1 and Mbarr1 in Kbarr1 EX1180 yeast. Red dot-line arrow indicates unknown equivalent ES in M viruses for the ES of ScV-LBC1-original.
, ribosomal frameshift. Black arrow indicates the single nucleotide changes between ES of LBCbarr1 and LBCbarr2. Green dot-line arrow indicates equivalent ES for L and M viruses in the same yeast. Orange dot-line arrow indicates possible equivalent ES for LBCbarr1 and Mbarr1 in Kbarr1 EX1180 yeast. Red dot-line arrow indicates unknown equivalent ES in M viruses for the ES of ScV-LBC1-original.
, ribosomal frameshift. Black arrow indicates the single nucleotide changes between ES of LBCbarr1 and LBCbarr2. Green dot-line arrow indicates equivalent ES for L and M viruses in the same yeast. Orange dot-line arrow indicates possible equivalent ES for LBCbarr1 and Mbarr1 in Kbarr1 EX1180 yeast. Red dot-line arrow indicates unknown equivalent ES in M viruses for the ES of ScV-LBC1-original.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Previous Sequenced Length (bp)/Yeast Strain | Newly Analysed Yeast Strain | Killer Phenotype/dsRNA Isotype | Sequenced Length (bp) (Canonical) |
|---|---|---|---|---|
| LBC1 | 4615/Sc 299 (LBC1-original) | Sc EX231 | K1/M1-1 | 4971 (4615) |
| LBC2 | 4614/Sc S3920 | Sc EX1125 | K2/M2-4 | 4722 (4615) |
| LBClus | 4614/Sc EX229 | Sc EX229 | Klus/Mlus4 | 4839 (4615) |
| Sc EX1160 | Klus/MlusA | 4661 (4615) | ||
| LBCbarr-1 | Unknown/Td EX1180 | Td EX1180 | Kbarr1/LBCbarr1 | 4763 (4565) |
| LBCbarr-2 | Unknown/Td EX1257 | Td EX1257 | Kbarr2/LBCbarr2 | 5115 (4565) |
| Virus | Accession Number | Reference/Comment |
|---|---|---|
| ScV-LBC1-original | U01060.1 | Previously [7] known as ScV-La or L-B-C. Renamed in this study to distinguish it from other LBC viruses from different K1 killer yeasts |
| ScV-LBClus4-EX229 | KT784813.1 | [4] |
| ScV-LBC2-S3920 | KX906605.1 | [4] |
| TdV-LAbarr1-EX1180 | MW174763 | [12] |
| ScV-LA1-original | J04692.1 | Previously [11] known as ScV-LA. Renamed in this study to distinguish it from other LA viruses from different K1 killer yeast strains |
| ScV-LA1-EX231 | MW174760 | [12] |
| ScV-LAlus-EX229 | JN819511.1 | [13] |
| ScV-LAlus4-EX229 | MW174758 | [12] |
| ScV-LAlus1-EX436 | MW174761 | [12] |
| ScV-LAlusA-EX1160 | MW174762 | [12] |
| ScV-LA2-S3920 | KC677754.1 | [4] |
| ScV-LA2-EX1125 | MW174759 | [12] |
| SpV-LA28 | KU845301.2 | Formerly [27] assigned to S. cerevisiae but recently re-assigned to S. paradoxus [6] |
| Strain | Genotype [Relevant Phenotype] | Origin, Date, Grape Variety, Geographical Location in Extremadura (Spain) |
|---|---|---|
| Td EX1180 | wt LAbarr1 LBCbarr1 Mbarr1 [Kbarr1+] | Wine, 2006, Cayetana, Mérida |
| Td EX1257 | wt LAbarr2 LBCbarr2 Mbarr1 M1 [Kbarr2+] | Wine, 2006, Tempranillo, Badajoz |
| Sc EX231 | wt MAT a/α HO/HO LA1 LBC1 M1-1 [K1+] | Wine, 2003, Macabeo, Guadajira |
| Sc EX1125 | wt MAT a/α HO/HO LA2 LBC1 M2-4 [K2+] | Wine, 2005, Moscatel, La Albuera |
| Sc EX229 | wt MAT a/α HO/HO LAlus4 LBClus4 Mlus4 [Klus+] | Wine, 2003, Macabeo, Guadajira |
| Sc EX1160 | wt MAT a/α HO/HO LAlusA LBClusA MlusA MlusB MlusC [Klus+] | Wine, 2005, Moscatel, La Albuera |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramírez, M.; Velázquez, R.; López-Piñeiro, A.; Martínez, A. Genome Features of a New Double-Stranded RNA Helper Virus (LBCbarr) from Wine Torulaspora delbrueckii Killer Strains. Int. J. Mol. Sci. 2021, 22, 13492. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413492
Ramírez M, Velázquez R, López-Piñeiro A, Martínez A. Genome Features of a New Double-Stranded RNA Helper Virus (LBCbarr) from Wine Torulaspora delbrueckii Killer Strains. International Journal of Molecular Sciences. 2021; 22(24):13492. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413492
Chicago/Turabian StyleRamírez, Manuel, Rocío Velázquez, Antonio López-Piñeiro, and Alberto Martínez. 2021. "Genome Features of a New Double-Stranded RNA Helper Virus (LBCbarr) from Wine Torulaspora delbrueckii Killer Strains" International Journal of Molecular Sciences 22, no. 24: 13492. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413492