
Characterization of a Novel Thermostable Dye-Linked l-Lactate Dehydrogenase Complex and Its Application in Electrochemical Detection

 , ,
, ,
Abstract
:1. Introduction
2. Results
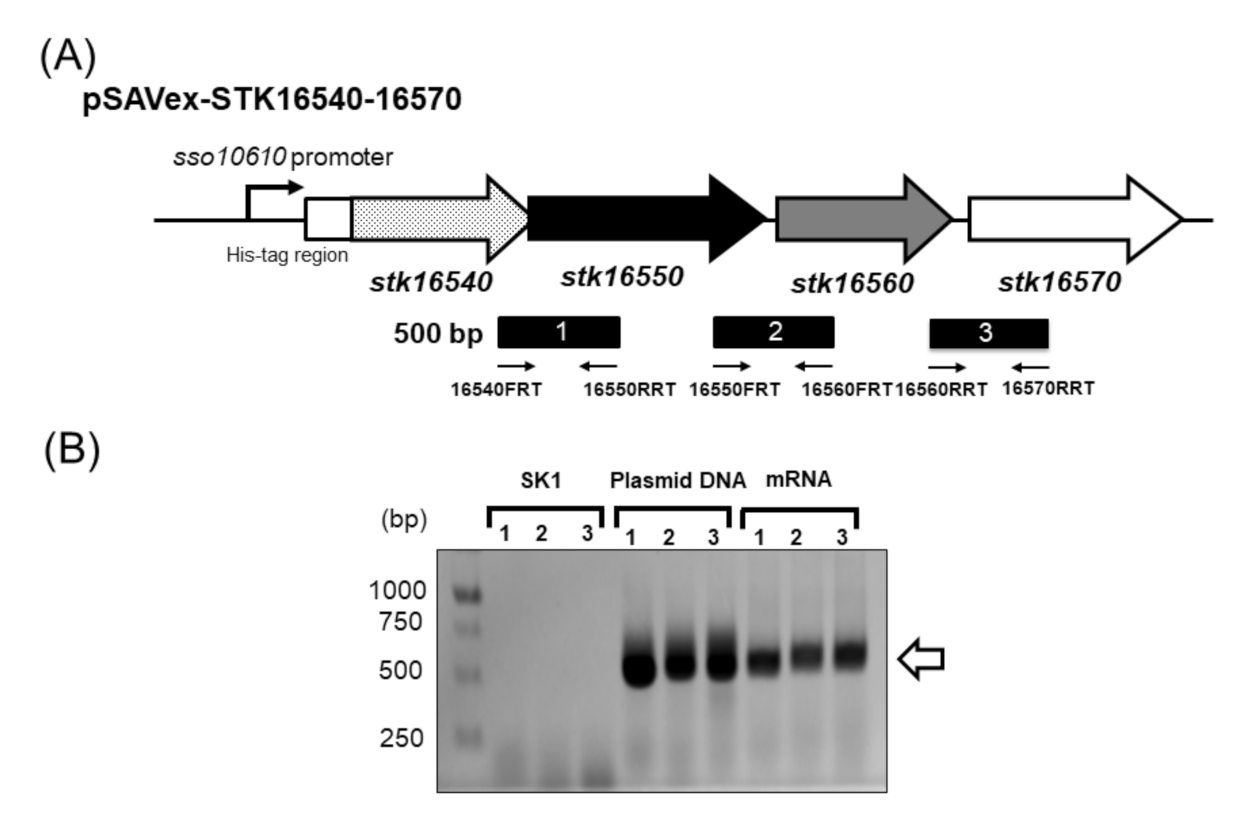
2.1. Expression of Dye-LDH in S. acidocaldarius SK1
2.2. Purification and Molecular Composition of Recombinant Dye-LDH from S. acidocaldarius SK1 Carrying pSAVex-STK16540-16570
2.3. Detection of Prosthetic Group
2.4. Effect of pH and Temperature on Dye-LDH Activity and Stability
2.5. Electron Donor and Acceptor Specificities
2.6. Characterization of PES-Modified Dye-LDH Electrode
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Strains and Cultivation Media
4.3. Plasmid Construction for Gene Expression in S. acidocaldarius SK1
4.4. Total RNA Preparation and Reverse Transcription PCR
4.5. Determination of Dye-LDH Activity and Protein Concentrations
4.6. Expression and Purification of Recombinant Dye-LDH from S. solfataricus
4.7. Molecular Mass Determination of Protein
4.8. Western Blot and N-Terminal Amino Acid Analysis
4.9. Determination of Prosthetic Group
4.10. Effect of Temperature and pH on Enzyme Activity and Stability
4.11. Kinetic Parameters
4.12. Preparation of a Dye-LDH Immobilized Electrode
4.13. Electrochemical Measurements
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| Dye-LDH | dye-linked l-lactate dehydrogenase |
| DCIP | 2,6-dichlorophenolindophenol |
| FAD | flavin adenine dinucleotide |
| FMN | flavin mononucleotide |
| INT | p-iodonitrotetrazolium violet |
| MTT | 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide |
| MWCNT | multi-walled carbon nanotube |
| PES | phenazine ethosulfate |
References
- Garvie, E.I. Bacterial lactate dehydrogenases. Microbiol. Rev. 1980, 44, 106–139. [Google Scholar] [CrossRef] [PubMed]
- Tabacchi, G.; Zucchini, D.; Caprini, G.; Gamba, A.; Lederer, F.; Vanoni, M.A.; Fois, E. L-lactate dehydrogenation in flavocytochrome b2: A first principles molecular dynamics study. FEBS J. 2009, 276, 2368–2680. [Google Scholar] [CrossRef] [PubMed]
- Shkil, H.; Stoica, L.; Dmytruk, K.; Smutok, O.; Gonchar, M.; Sibirny, A.; Schuhmann, W. Bioelectrochemical detection of L-lactate respiration using genetically modified Hansenula polymorpha yeast cells overexpressing flavocytochrome b2. Bioelectrochemistry 2009, 76, 175–179. [Google Scholar] [CrossRef]
- Smutok, O.; Gayda, G.; Gonchar, M.; Schuhmann, W. A novel L-lactate-selective biosensor based on flavocytochrome b2 from methylotrophic yeast Hansenula polymorpha. Biosens. Bioelectron. 2005, 20, 1285–1290. [Google Scholar] [CrossRef] [PubMed]
- Smutok, O.; Dmytruk, K.; Gonchar, M.; Sibirny, A.; Schuhmann, W. Permeabilized cells of flavocytochrome b2 over-producing recombinant yeast Hansenula polymorpha as biological recognition element in amperometric lactate biosensors. Biosens. Bioelectron. 2007, 23, 599–605. [Google Scholar] [CrossRef]
- Shkil, H.; Schulte, A.; Guschin, D.A.; Schuhmann, W. Electron transfer between genetically modified Hansenula polymorpha yeast cells and electrode surfaces via Os-complex modified redox polymers. ChemPhysChem 2011, 12, 806–813. [Google Scholar] [CrossRef]
- Garjonyte, R.; Melvydas, V.; Malinauskas, A. Mediated amperometric biosensors for lactic acid based on carbon paste electrodes modified with baker’s yeast Saccharomyces cerevisiae. Bioelectrochemistry 2006, 68, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Garjonyte, R.; Melvydas, V.; Malinauskas, A. Effect of yeast pretreatment on the characteristics of yeast-modified electrodes as mediated amperometric biosensors for lactic acid. Bioelectrochemistry 2008, 74, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Karkovska, M.; Smutok, O.; Stasyuk, N.; Gonchar, M. L-Lactate-selective microbial sensor based on flavocytochrome b2-enriched yeast cells using recombinant and nanotechnology approaches. Talanta 2015, 44, 1195–1200. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.W. Enzymes and proteins from organisms that grow near and above 100 degrees C. Annu. Rev. Microbiol. 1993, 47, 627–658. [Google Scholar] [CrossRef]
- Li, W.F.; Zhou, X.X.; Lu, P. Structural features of thermozymes. Biotechnol. Adv. 2005, 23, 271–281. [Google Scholar] [CrossRef]
- Ohshima, T.; Soda, K. Thermostable amino acid dehydrogenases: Application and gene cloning. Trends Biotechnol. 1989, 7, 210–214. [Google Scholar] [CrossRef]
- Berkner, S.; Wlodkowski, A.; Albers, S.V.; Lipps, G. Inducible and constitutive promoters for genetic systems in Sulfolobus acidocaldarius. Extremophiles 2010, 14, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Schägger, H.; von Jagow, G. Blue native electrophoresis for isolation of membrane protein complexes in enzymatically active form. Anal. Biochem. 1991, 199, 223–231. [Google Scholar] [CrossRef]
- Schägger, H.; Cramer, W.A.; von Jagow, G. Analysis of molecular masses and oligomeric states of protein complexes by blue native electrophoresis and isolation of membrane protein complexes by two-dimensional native electrophoresis. Anal. Biochem. 1994, 217, 220–230. [Google Scholar] [CrossRef]
- Schägger, H. Blue native gels to isolate protein complexes from mitochondria. Methods Cell Biol. 2001, 65, 231–244. [Google Scholar] [PubMed]
- Tsuge, H.; Kawakami, R.; Sakuraba, H.; Ago, H.; Miyano, M.; Aki, K.; Katunuma, N.; Ohshima, T. Crystal structure of a novel FAD-, FMN-, and ATP-containing L-proline dehydrogenase complex from Pyrococcus horikoshii. J. Biol. Chem. 2005, 280, 31045–31049. [Google Scholar] [CrossRef] [Green Version]
- Sakuraba, H.; Takamatsu, Y.; Satomura, T.; Kawakami, R.; Ohshima, T. Purification, characterization, and application of a novel dye-linked L-proline dehydrogenase from a hyperthermophilic archaeon, Thermococcus profundus. Appl. Environ. Microbiol. 2001, 67, 1470–1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawakami, R.; Sakuraba, H.; Ohshima, T. Gene and primary structures of dye-linked L-proline dehydrogenase from the hyperthermophilic archaeon Thermococcus profundus show the presence of a novel heterotetrameric amino acid dehydrogenase complex. Extremophiles 2004, 8, 99–108. [Google Scholar] [CrossRef]
- Kawakami, R.; Sakuraba, H.; Tsuge, H.; Goda, S.; Katunuma, N.; Ohshima, T. A second novel dye-linked L-proline dehydrogenase complex is present in the hyperthermophilic archaeon Pyrococcus horikoshii OT-3. FEBS J. 2005, 272, 4044–4054. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Lee, J.; Loew, N.; Takahashi-Inose, Y.; Okuda-Shimazaki, J.; Kojima, K.; Mori, K.; Tsugawa, W.; Sode, K. Engineered glucose oxidase capable of quasi-direct electron transfer after a quick-and-easy modification with a mediator. Int. J. Mol. Sci. 2020, 21, 1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiraka, K.; Kojima, K.; Tsugawa, W.; Asano, R.; Ikebukuro, K.; Sode, K. Rational engineering of Aerococcus viridans L-lactate oxidase for the mediator modification to achieve quasi-direct electron transfer type lactate sensor. Biosens. Bioelectron. 2020, 151, 111974. [Google Scholar] [CrossRef] [PubMed]
- Hatada, M.; Saito, S.; Yonehara, S.; Tsugawa, W.; Asano, R.; Ikebukuro, K.; Sode, K. Development of glycated peptide enzyme sensor based flow injection analysis system for haemoglobin A1c monitoring using quasi-direct electron transfer type engineered fructosyl peptide oxidase. Biosens. Bioelectron. 2012, 177, 112984. [Google Scholar] [CrossRef] [PubMed]
- Hatada, M.; Loew, N.; Inose-Takahashi, Y.; Okuda-Shimazaki, J.; Tsugawa, W.; Mulchandani, A.; Sode, K. Development of a glucose sensor employing quick and easy modification method with mediator for altering electron acceptor preference. Bioelectrochemistry 2018, 121, 185–190. [Google Scholar] [CrossRef]
- Suzuki, S.; Kurosawa, N. Disruption of the gene encoding restriction endonuclease Sua I and development of a host-vector system for the thermoacidophilic archaeon Sulfolobus acidocaldarius. Extremophiles 2016, 20, 139–148. [Google Scholar] [CrossRef]
- Satomura, T.; Emoto, S.; Kurosawa, N.; Ohshima, T.; Sakuraba, H.; Suye, S. Characterization of dye-linked D-amino acid dehydrogenase from Sulfurisphaera tokodaii expressed using an archaeal recombinant protein expression system. J. Biosci. Bioeng. 2020, 130, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, R.; Krummel, B.; Saiki, R.K. A general method of in vitro preparation and specific mutagenesis of DNA fragments: Study of protein and DNA interactions. Nucleic Acids Res. 1998, 16, 7351–7367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Satomura, T.; Kawakami, R.; Sakuraba, H.; Ohshima, T. Dye-linked D-proline dehydrogenase from hyperthermophilic archaeon, Pyrobaculum islandicum is a novel FAD-dependent amino acid dehydrogenase. J. Biol. Chem. 2002, 277, 12861–12867. [Google Scholar] [CrossRef] [Green Version]
- Davis, B.J. Disc electrophoresis. 2. Method and application to human serum proteins. Ann. N. Y. Acad. Sci. 1975, 121, 404–427. [Google Scholar] [CrossRef] [PubMed]
- Brill, W.J.; Westphal, J.; Stieghorst, M.; Davis, L.C.; Shah, V.K. Detection of nitrogenase components and other nonheme iron proteins in polyacrylamide gels. Anal. Biochem. 1974, 60, 237–241. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Protein (mg) | Total Activity (units) | Specific Activity (units/mg) | Yield (%) | Fold Purification | |
|---|---|---|---|---|---|
| Crude extract | 477 | 6.99 | 0.0156 | 100 | 1.00 |
| HisTrap FF Crude | 1.51 | 0.684 | 0.453 | 9.79 | 28.9 |
| HiTrap Q HP | 0.441 | 0.676 | 1.53 | 9.67 | 98.1 |
| Primer | Sequence |
|---|---|
| pET16540F | CCGCGCGGCAGCCATATGCTGGGAAAATTAATTTATGATA |
| pET16570R | TTAGCAGCCGGATCCTTAAGTAGTTTCGTATTTTATTTTT |
| pET15bF | ATGGCTGCCGCGCGGCACCAGGCCGCTGCT |
| pET15bR | GGATCCGGCTGCTAACAAAGCCCGAAAGGA |
| SSO16540F | GTGAAACCTTTAAGAATAGCATGGGCAGCAGCCATCATCA |
| SSO16570R | GGCCTCTTGCGGGATATCCGGATATAGTTC |
| pSSOF | AGCCATTGTCATAGTTTTCACTAGCTAAAA |
| pSSOR | GCTATTCTTAAAGGTTTCACATAAATAAAC |
| Fusion-pSSO | TGATGATGGCTGCTGCCCATGCTATTCTTAAAGGTTTCAC |
| pSAVF | AGCGCAACGCAATTAATGTGAGTTAGCTCA |
| pSAVR | CACTGCCCGCTTTCCAGTCGGGAAACCTGT |
| Infusion pSAV2F | GGAAAGCGGGCAGTGAGCCATTGTCATAGTTTTCA |
| Infusion pSAV2R | TAATTGCGTTGCGCTGGCCTCTTGCGGGATATCCG |
| 16540FRT | GGGTAGAACAGTTGTCTAAGCTTAGTAAGA |
| 16550RRT | AATATGTGAAGGTGGTTCATCAGCTAACTG |
| 16550FRT | CTGTATTAGATGCGGAAGATGTCATCTTCA |
| 16560FRT | GTAGTATATTAGCTCCGGTTATACCATTCC |
| 16560RRT | TGAAAAGACTTTCACCATCTCCAATAGGAC |
| 16570RRT | AGAAATCCATTAATTGTCACGTTCATGGGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Satomura, T.; Uno, K.; Kurosawa, N.; Sakuraba, H.; Ohshima, T.; Suye, S.-i. Characterization of a Novel Thermostable Dye-Linked l-Lactate Dehydrogenase Complex and Its Application in Electrochemical Detection. Int. J. Mol. Sci. 2021, 22, 13570. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413570
Satomura T, Uno K, Kurosawa N, Sakuraba H, Ohshima T, Suye S-i. Characterization of a Novel Thermostable Dye-Linked l-Lactate Dehydrogenase Complex and Its Application in Electrochemical Detection. International Journal of Molecular Sciences. 2021; 22(24):13570. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413570
Chicago/Turabian StyleSatomura, Takenori, Kohei Uno, Norio Kurosawa, Haruhiko Sakuraba, Toshihisa Ohshima, and Shin-ichiro Suye. 2021. "Characterization of a Novel Thermostable Dye-Linked l-Lactate Dehydrogenase Complex and Its Application in Electrochemical Detection" International Journal of Molecular Sciences 22, no. 24: 13570. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413570