
Characterizing the Interaction between the HTLV-1 Transactivator Tax-1 with Transcription Elongation Factor ELL2 and Its Impact on Viral Transactivation

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
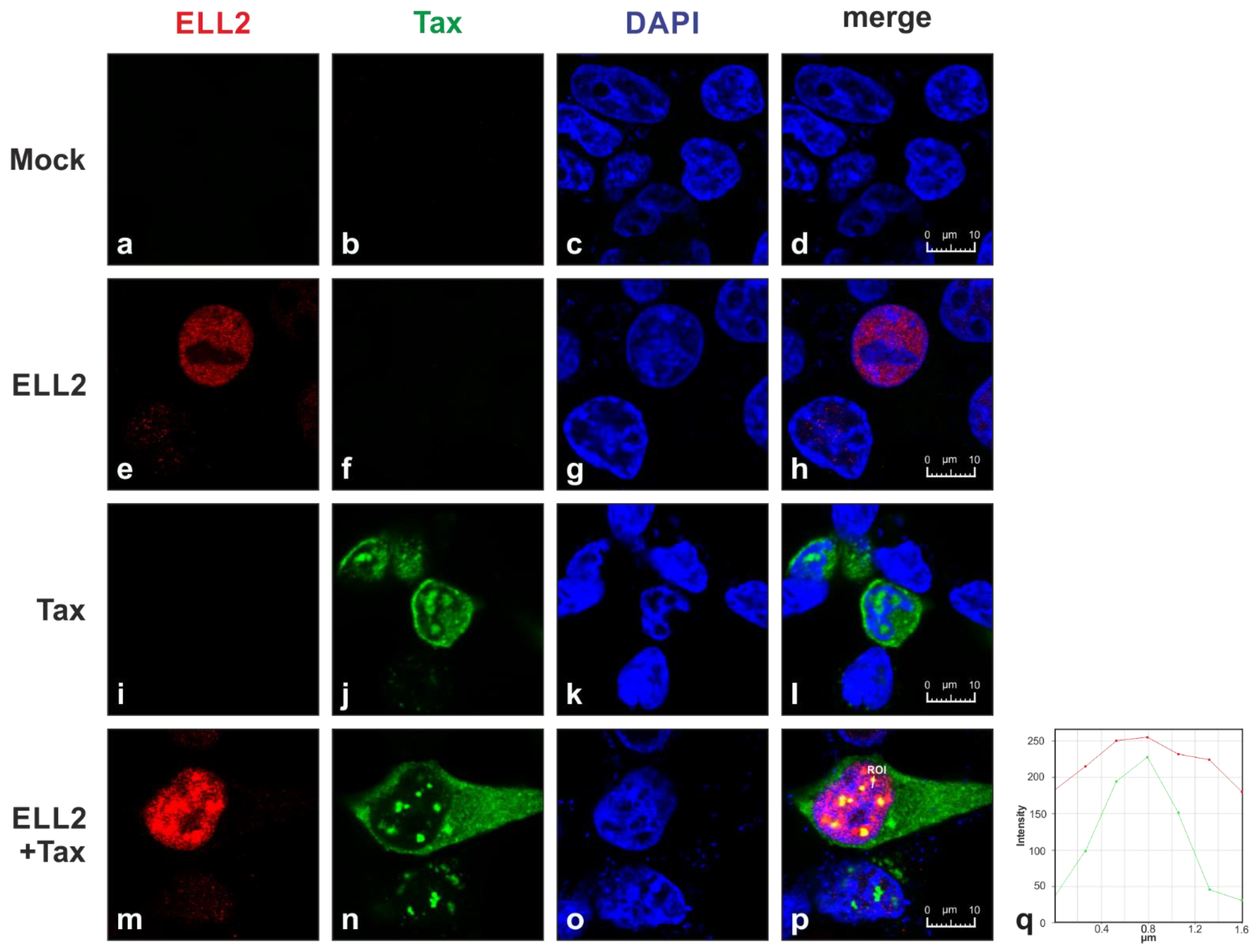
2.1. Tax-1 and ELL2 Co-Localize in Dot-like Structures in the Nucleus
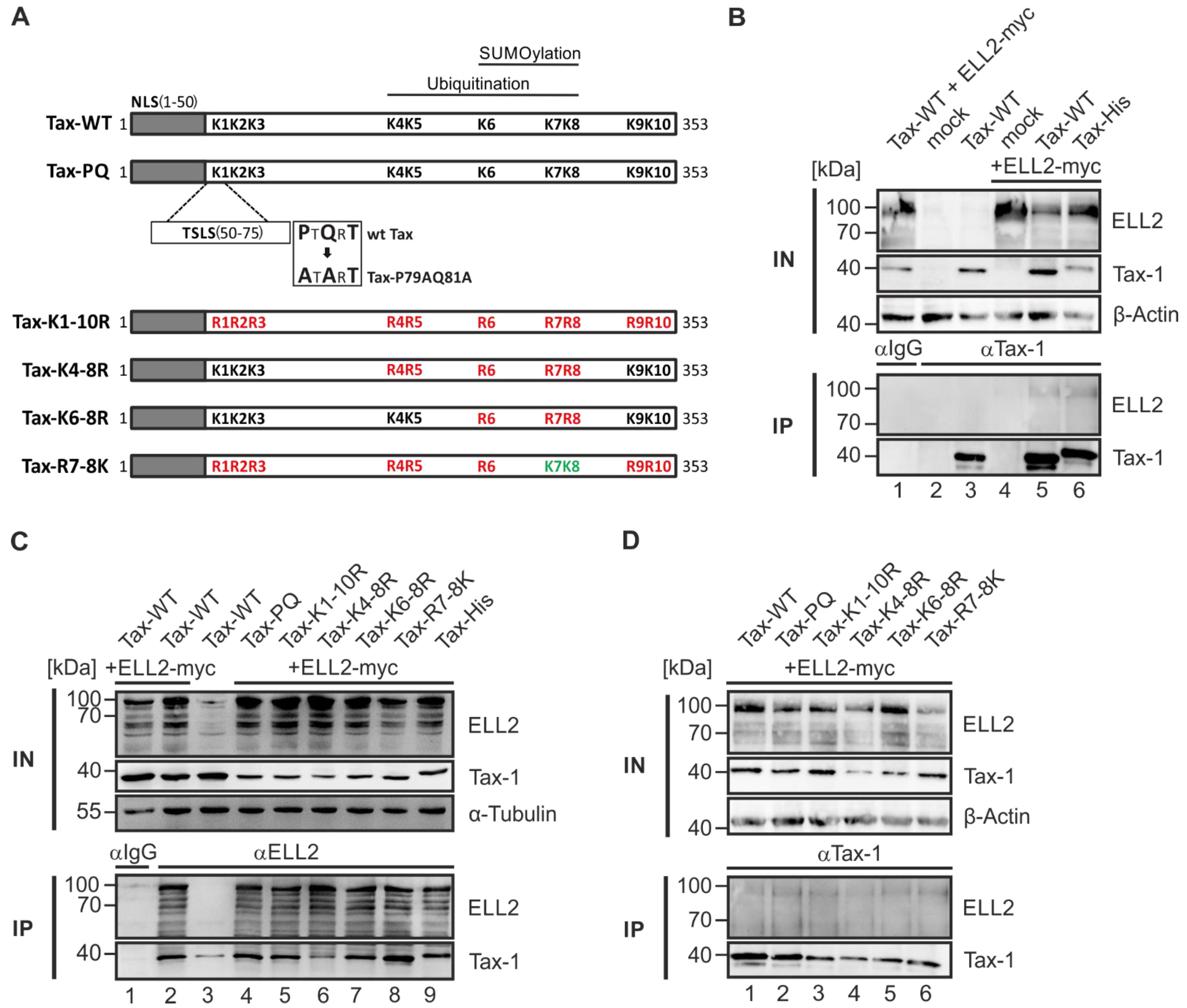
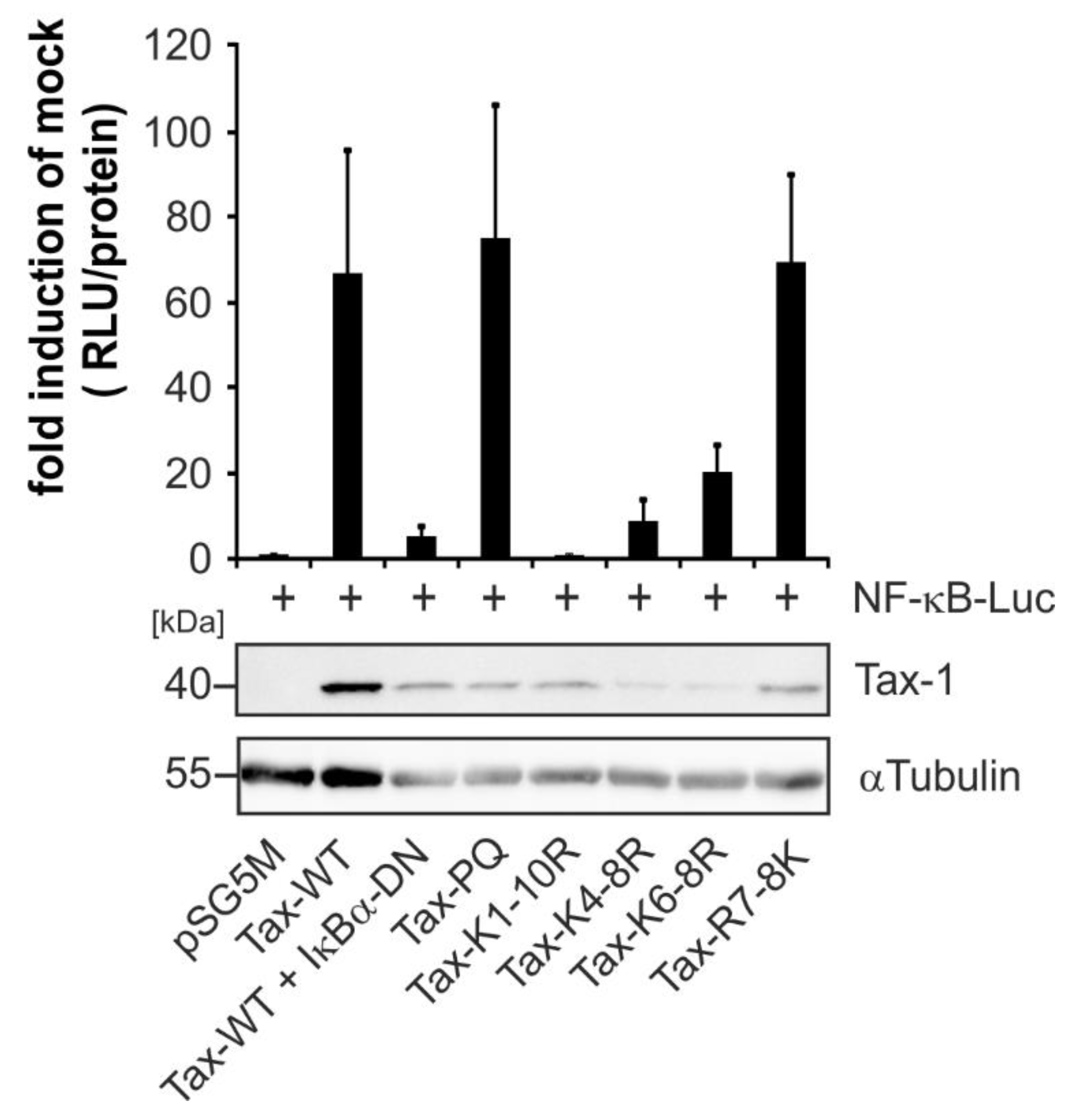
2.2. Point Mutants Impairing Post Translational Modifications of Tax-1 Do Not Affect Tax-1:ELL2 Complex Formation
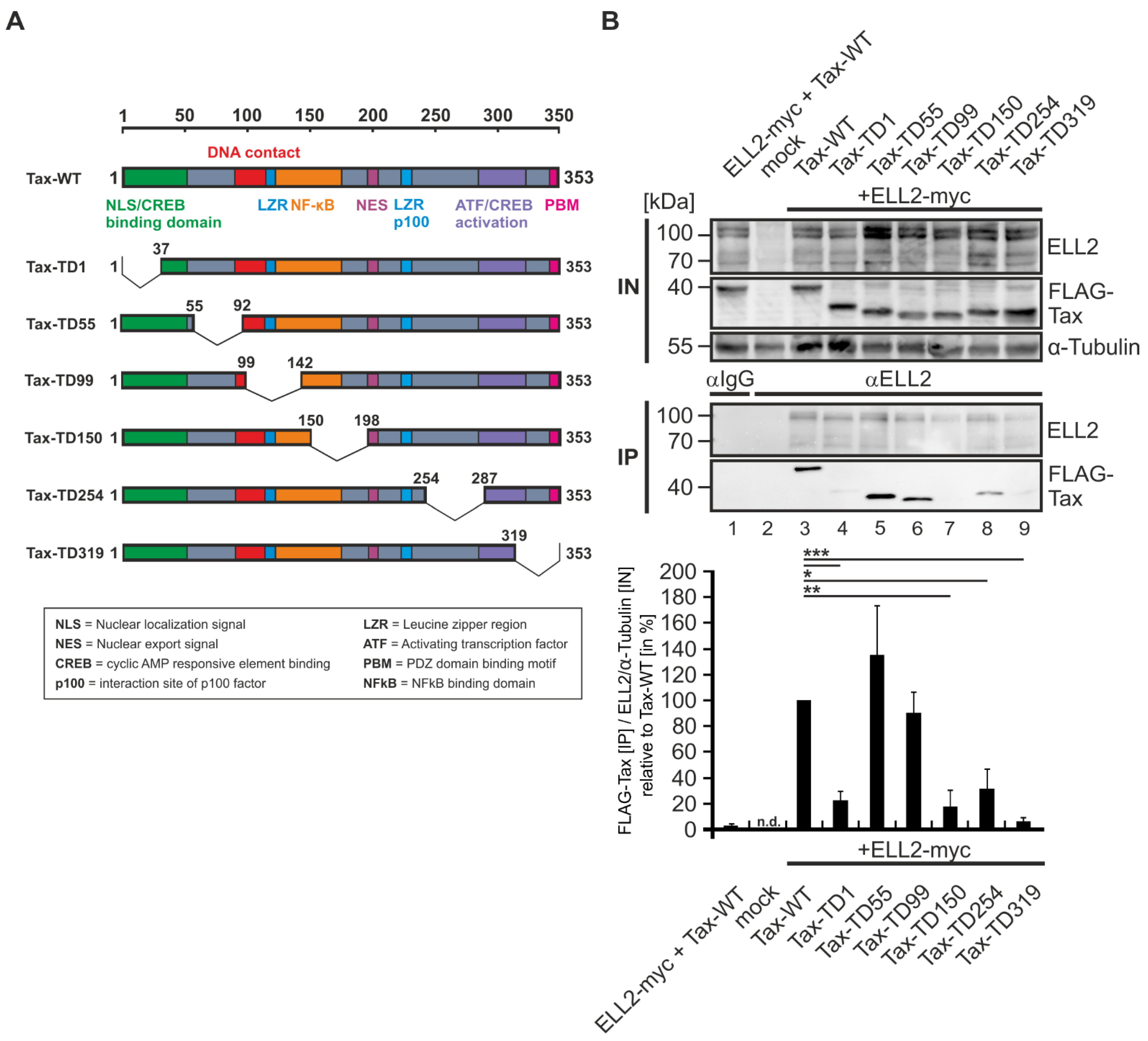
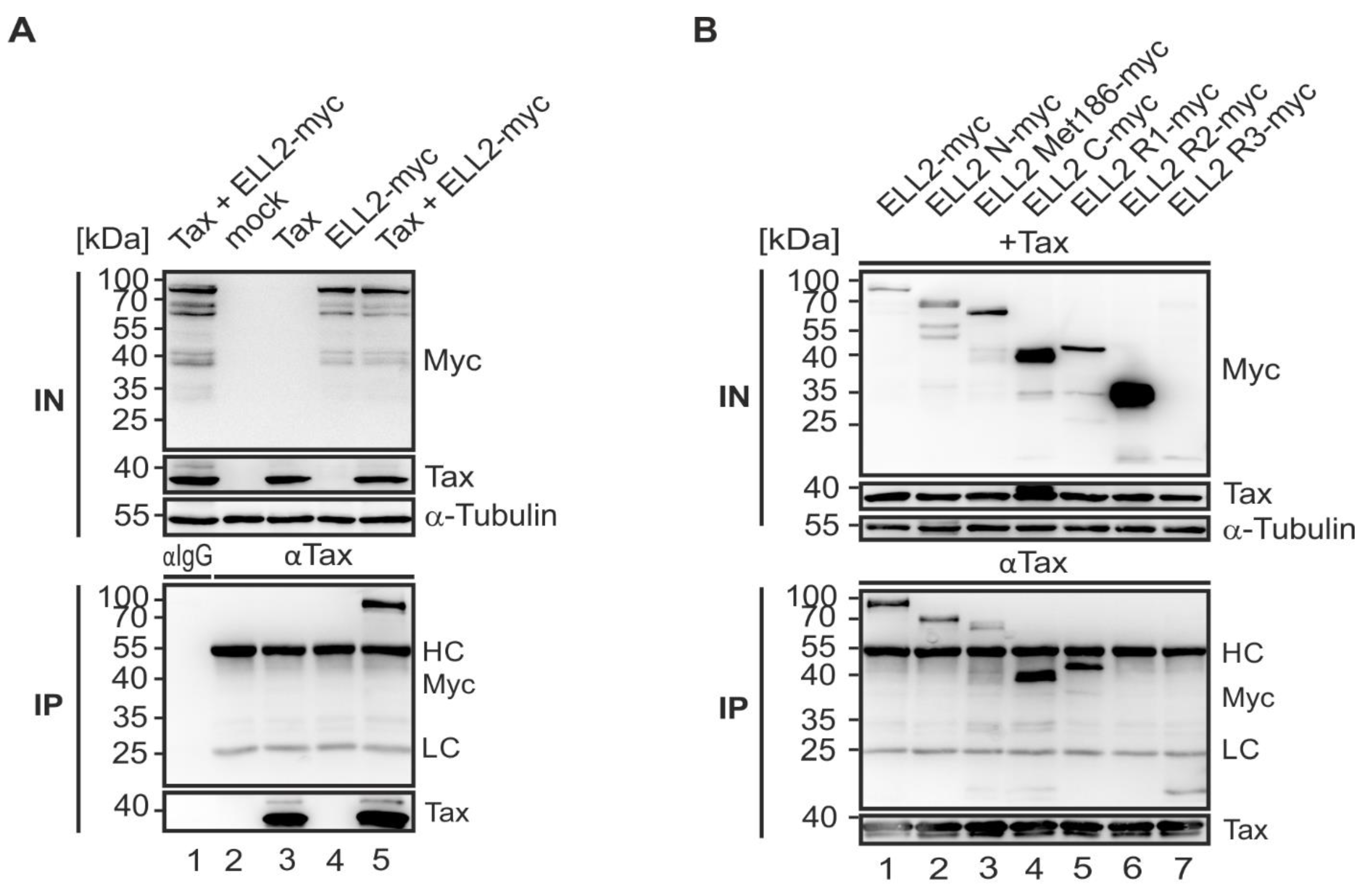
2.3. N- and C-Terminal Domains of Tax-1 Are Critical for Tax-1:ELL2 Complex Formation
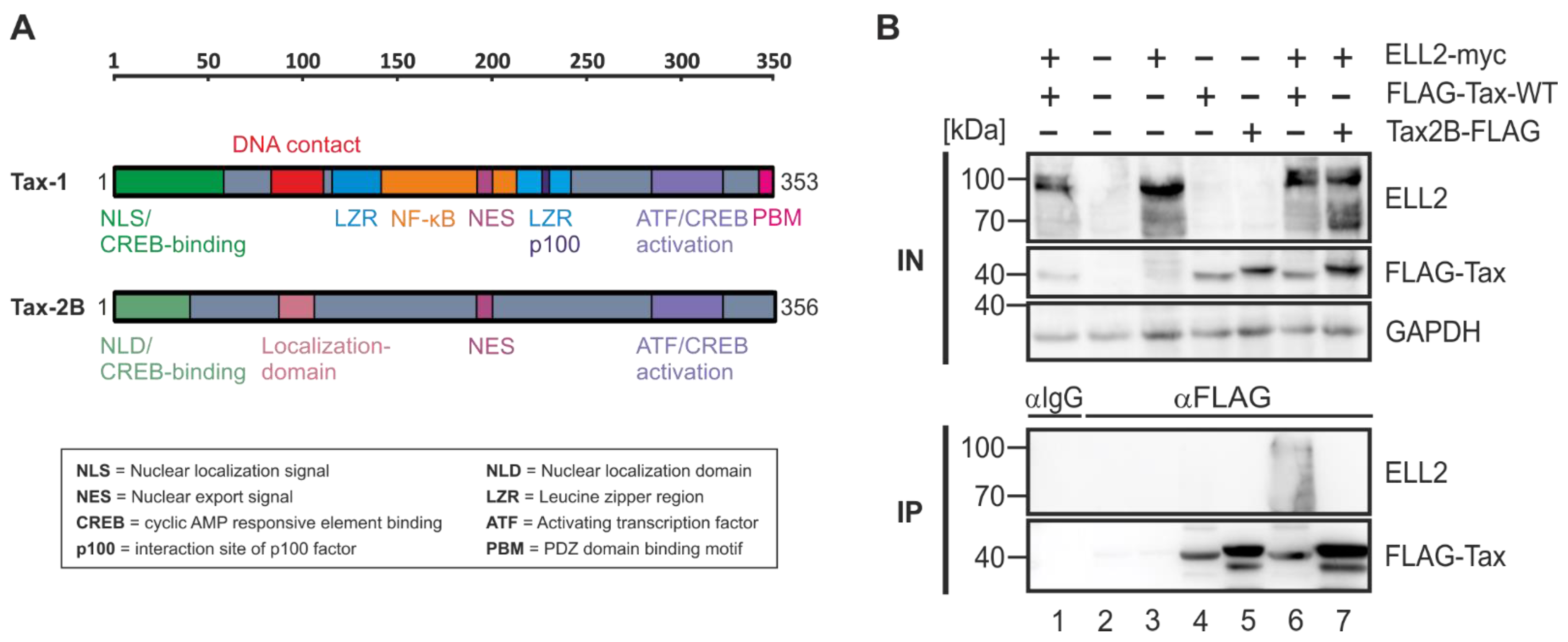
2.4. Tax-2B from HTLV-2B Does Not Interact with ELL2
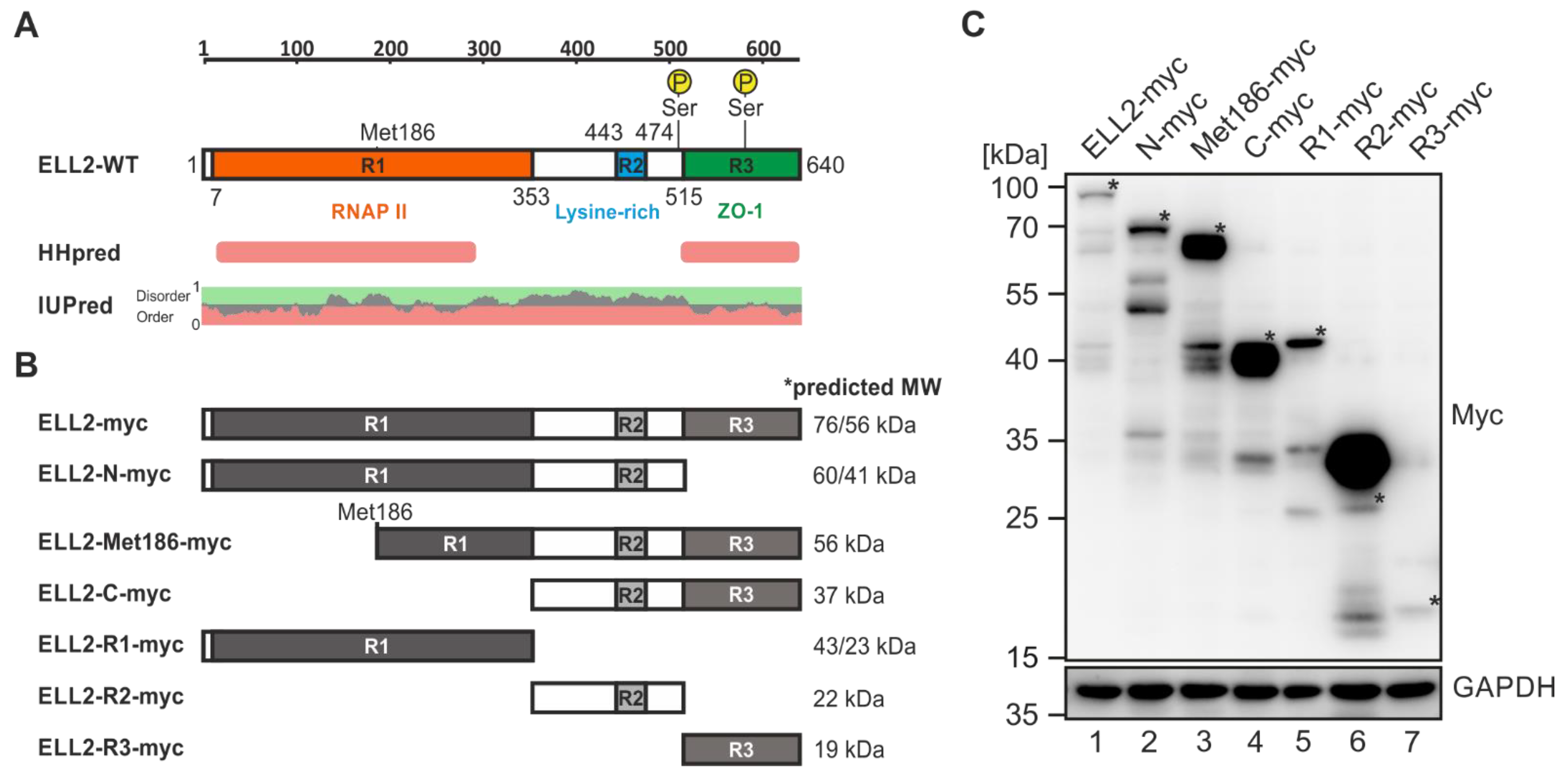
2.5. ELL2 Is a Highly Post Translationally Modified Protein That Contains Two Globular Domains
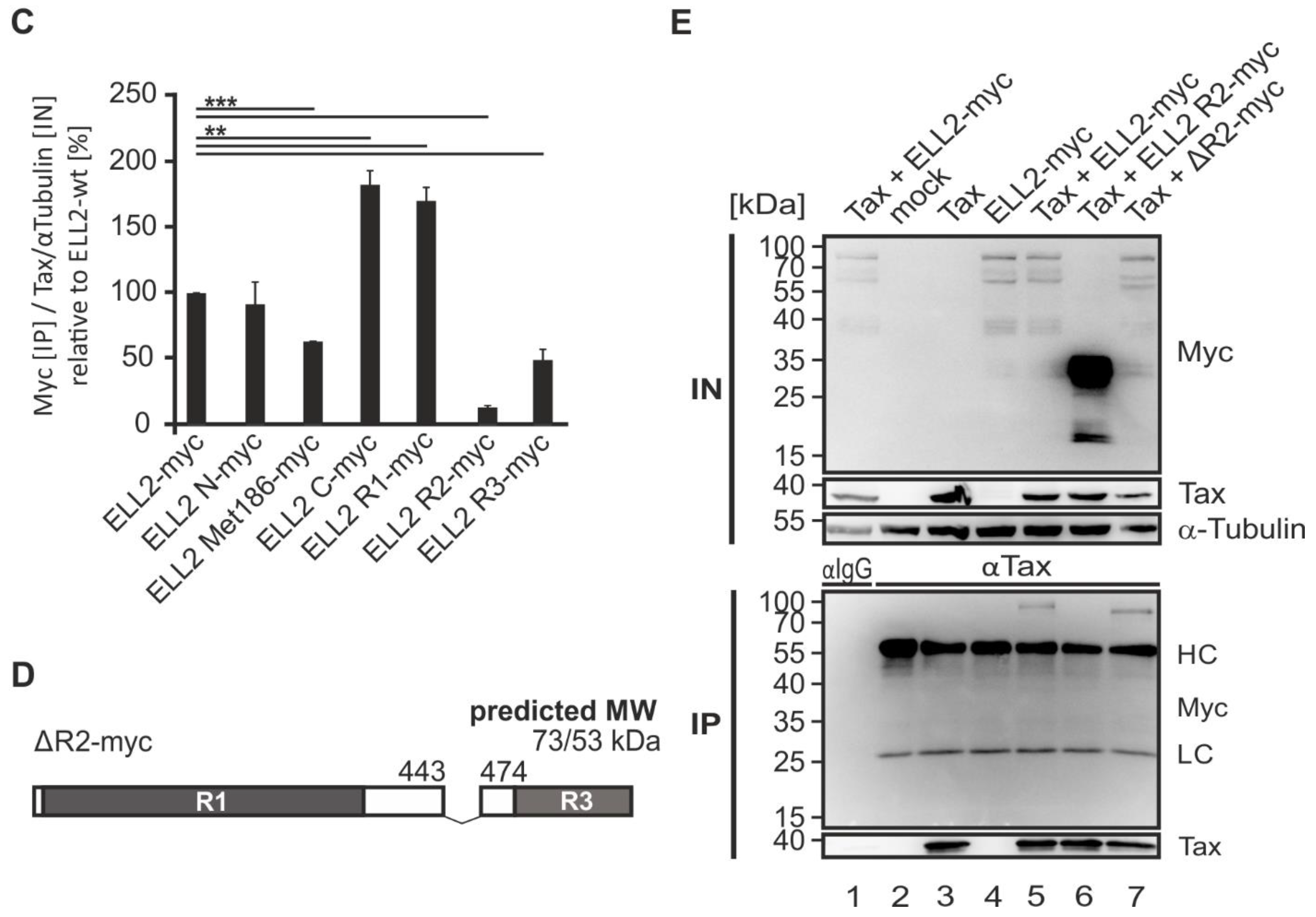
2.6. N- and C-Terminal Domains of ELL2 Are Critical for Tax-1:ELL2 Complex Formation
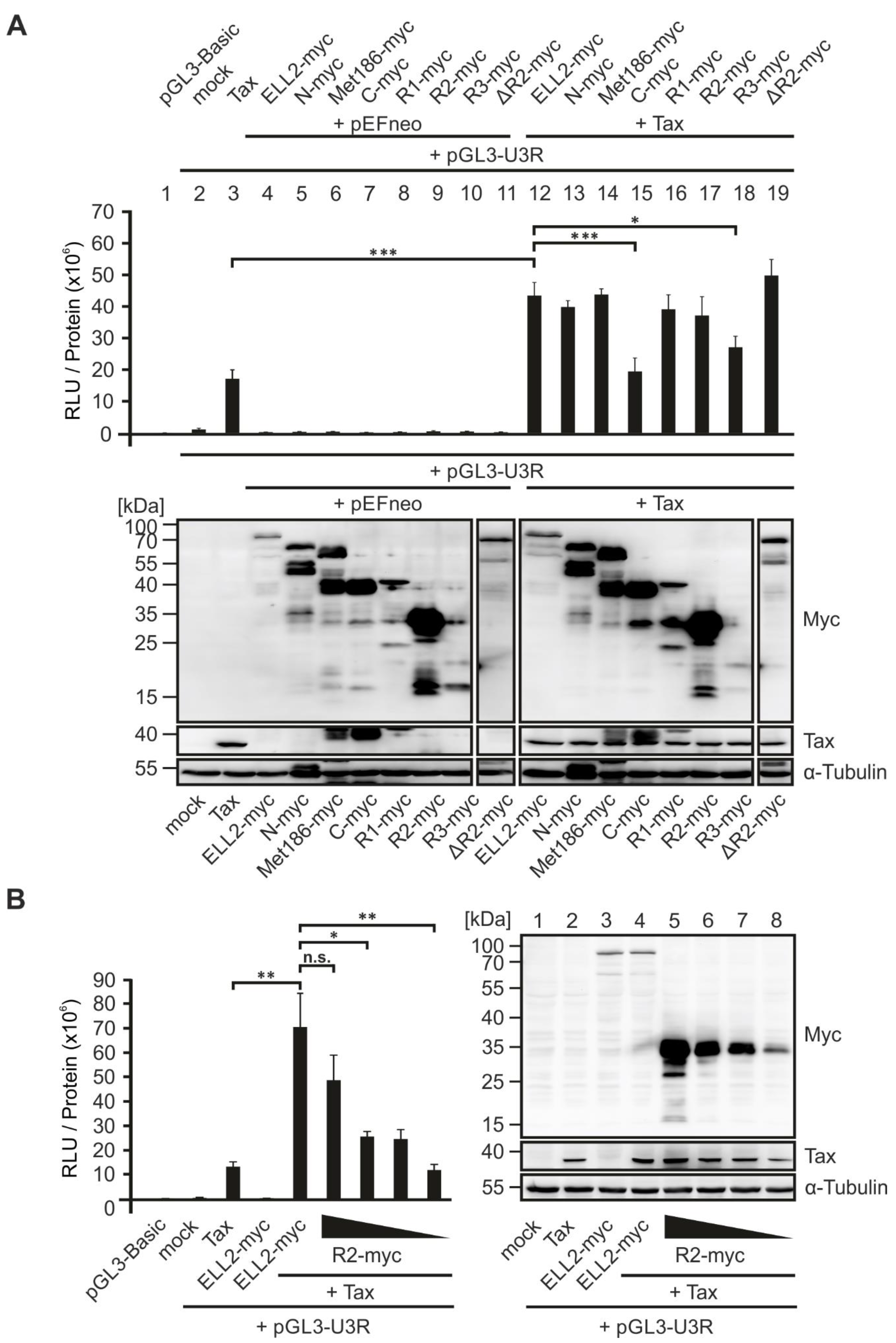
2.7. An N-Terminal Conserved Region of ELL2, ELL2-R1, Is Crucial for Enhancing Tax-1-Mediated Transactivation of the HTLV-1 Promoter
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Plasmids and Cloning
4.3. Transient Transfections
4.4. Western Blot and Antibodies
4.5. Luciferase Reporter Assays
4.6. Co-Immunoprecipitation
4.7. Immunofluorescence and Confocal Laser Scanning Microscopy
4.8. Statistics
4.9. Bioinformatics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Gessain, A.; Vernant, J.; Maurs, L.; Barin, F.; Gout, O.; Calender, A.D.; De Thé, G. Antibodies to human T-lymphotropic virus type-I in patients with tropical spastic paraparesis. Lancet 1985, 326, 407–410. [Google Scholar] [CrossRef]
- Poiesz, B.J.; Ruscetti, F.W.; Gazdar, A.F.; Bunn, P.A.; Minna, J.D.; Gallo, R.C. Detection and isolation of type C retrovirus particles from fresh and cultured lymphocytes of a patient with cutaneous T-cell lymphoma. Proc. Natl. Acad. Sci. USA 1980, 77, 7415–7419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, M.; Miyoshi, I.; Hinuma, Y. Isolation and characterization of retrovirus from cell lines of human adult T-cell leukemia and its implication in the disease. Proc. Natl. Acad. Sci. USA 1982, 79, 2031–2035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osame, M.; Usuku, K.; Izumo, S.; Ijichi, N.; Amitani, H.; Igata, A.; Matsumoto, M.; Tara, M. HTLV-I associated myelopathy, a new clinical entity. Lancet 1986, 1, 1031–1032. [Google Scholar] [CrossRef]
- Gessain, A.; Cassar, O. Epidemiological aspects and world distribution of HTLV-1 infection. Front. Microbiol. 2012, 3, 388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igakura, T.; Stinchcombe, J.C.; Goon, P.K.C.; Taylor, G.P.; Weber, J.N.; Griffiths, G.M.; Tanaka, Y.; Osame, M.; Bangham, C.R.M. Spread of HTLV-I Between Lymphocytes by Virus-Induced Polarization of the Cytoskeleton. Science 2003, 299, 1713–1716. [Google Scholar] [CrossRef] [Green Version]
- Pais-Correia, A.-M.; Sachse, M.; Guadagnini, S.; Robbiati, V.; Lasserre, R.; Gessain, A.; Gout, O.; Alcover, A.; Thoulouze, M.-I. Biofilm-like extracellular viral assemblies mediate HTLV-1 cell-to-cell transmission at virological synapses. Nat. Med. 2010, 16, 83–89. [Google Scholar] [CrossRef]
- Van Prooyen, N.; Gold, H.; Andresen, V.; Schwartz, O.; Jones, K.; Ruscetti, F.; Lockett, S.; Gudla, P.; Venzon, D.; Franchini, G. Human T-cell leukemia virus type 1 p8 protein increases cellular conduits and virus transmission. Proc. Natl. Acad. Sci. USA 2010, 107, 20738–20743. [Google Scholar] [CrossRef] [Green Version]
- Gross, C.; Thoma-Kress, A.K. Molecular Mechanisms of HTLV-1 Cell-to-Cell Transmission. Viruses 2016, 8, 74. [Google Scholar] [CrossRef] [PubMed]
- Pique, C.; Jones, K.S. Pathways of cell-cell transmission of HTLV-1. Front. Microbiol. 2012, 3, 378. [Google Scholar] [CrossRef] [Green Version]
- Sodroski, J.G.; Rosen, C.A.; Haseltine, W.A. Trans -Acting Transcriptional Activation of the Long Terminal Repeat of Human T Lymphotropic Viruses in Infected Cells. Science 1984, 225, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Caswell, R.J.; Nall, P.; Boothby, M.; Taylor, G.P. Rapid onset and progression of myelopathy following an STI: A case for screening? Sex. Transm. Infect. 2019, 95, 244–245. [Google Scholar] [CrossRef] [PubMed]
- Bangham, C.R.M. CTL quality and the control of human retroviral infections. Eur. J. Immunol. 2009, 39, 1700–1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnell, A.P.; Kohrt, S.; Thoma-Kress, A.K. Latency Reversing Agents: Kick and Kill of HTLV-1? Int. J. Mol. Sci. 2021, 22, 5545. [Google Scholar] [CrossRef] [PubMed]
- Wodarz, D.; Nowak, M.A.; Bangham, C.R. The dynamics of HTLV-I and the CTL response. Immunol. Today 1999, 20, 220–227. [Google Scholar] [CrossRef]
- Lin, H.-C.; Dezzutti, C.S.; Lal, R.B.; Rabson, A.B. Activation of human T-cell leukemia virus type 1 tax gene expression in chronically infected T cells. J. Virol. 1998, 72, 6264–6270. [Google Scholar] [CrossRef] [Green Version]
- Giam, C.-Z.; Jeang, K.-T. HTLV-1 Tax and adult T-cell leukemia. Front. Biosci. 2007, 1, 1. [Google Scholar] [CrossRef] [Green Version]
- Boxus, M.; Willems, L. How the DNA damage response determines the fate of HTLV-1 Tax-expressing cells. Retrovirology 2012, 9, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Currer, R.; Van Duyne, R.; Jaworski, E.; Guendel, I.; Sampey, G.; Das, R.; Narayanan, A.; Kashanchi, F. HTLV Tax: A Fascinating Multifunctional Co-Regulator of Viral and Cellular Pathways. Front. Microbiol. 2012, 3, 406. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, M.; Jeang, K.-T. Human T-cell leukaemia virus type 1 (HTLV-1) infectivity and cellular transformation. Nat. Rev. Cancer 2007, 7, 270–280. [Google Scholar] [CrossRef]
- Zhou, M.; Lu, H.; Park, H.; Wilson-Chiru, J.; Linton, R.; Brady, J.N. Tax Interacts with P-TEFb in a Novel Manner To Stimulate Human T-Lymphotropic Virus Type 1 Transcription. J. Virol. 2006, 80, 4781–4791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ott, M.; Geyer, M.; Zhou, Q. The Control of HIV Transcription: Keeping RNA Polymerase II on Track. Cell Host Microbe 2011, 10, 426–435. [Google Scholar] [CrossRef] [Green Version]
- Peterlin, B.M.; Price, D.H. Controlling the Elongation Phase of Transcription with P-TEFb. Mol. Cell 2006, 23, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Price, D.H. P-TEFb, a Cyclin-Dependent Kinase Controlling Elongation by RNA Polymerase II. Mol. Cell. Biol. 2000, 20, 2629–2634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.; Lin, C.; Shilatifard, A. The super elongation complex (SEC) family in transcriptional control. Nat. Rev. Mol. Cell Biol. 2012, 13, 543–547. [Google Scholar] [CrossRef]
- He, N.; Liu, M.; Hsu, J.; Xue, Y.; Chou, S.; Burlingame, A.; Krogan, N.J.; Alber, T.; Zhou, Q. HIV-1 Tat and Host AFF4 Recruit Two Transcription Elongation Factors into a Bifunctional Complex for Coordinated Activation of HIV-1 Transcription. Mol. Cell 2010, 38, 428–438. [Google Scholar] [CrossRef] [Green Version]
- Shilatifard, A.; Duan, D.R.; Haque, D.; Florence, C.; Schubach, W.H.; Conaway, J.; Conaway, R.C. ELL2, a new member of an ELL family of RNA polymerase II elongation factors. Proc. Natl. Acad. Sci. USA 1997, 94, 3639–3643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sims, R.J.; Belotserkovskaya, R.; Reinberg, D. Elongation by RNA polymerase II: The short and long of it. Genes Dev. 2004, 18, 2437–2468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, N.; Zhou, Q. New Insights into the Control of HIV-1 Transcription: When Tat Meets the 7SK snRNP and Super Elongation Complex (SEC). J. Neuroimmune Pharmacol. 2011, 6, 260–268. [Google Scholar] [CrossRef] [Green Version]
- Martincic, K.; Alkan, S.A.; Cheatle, A.; Borghesi, L.; Milcarek, C. Transcription elongation factor ELL2 directs immunoglobulin secretion in plasma cells by stimulating altered RNA processing. Nat. Immunol. 2009, 10, 1102–1109. [Google Scholar] [CrossRef]
- Nelson, A.M.; Carew, N.T.; Smith, M.S.; Milcarek, C. RNA Splicing in the Transition from B Cells to Antibody-Secreting Cells: The Influences of ELL2, Small Nuclear RNA, and Endoplasmic Reticulum Stress. J. Immunol. 2018, 201, 3073–3083. [Google Scholar] [CrossRef] [Green Version]
- Ghobrial, A.; Flick, N.; Daly, R.; Hoffman, M.; Milcarek, C. ELL2 Influences Transcription Elongation, Splicing, Ig Secretion and Growth. J. Mucosal Immunol. Res. 2019, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Mann, M.C.; Strobel, S.; Fleckenstein, B.; Kress, A.K. The transcription elongation factor ELL2 is specifically upregulated in HTLV-1-infected T-cells and is dependent on the viral oncoprotein Tax. Virology 2014, 464, 98–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kfoury, Y.; Setterblad, N.; El-Sabban, M.; Zamborlini, A.; Dassouki, Z.; El Hajj, H.; Hermine, O.; Pique, C.; de Thé, H.; Saïb, A. Tax ubiquitylation and SUMOylation control the dynamic shuttling of Tax and NEMO between Ubc9 nuclear bodies and the centrosome. Blood J. Am. Soc. Hematol. 2011, 117, 190–199. [Google Scholar]
- Semmes, O.J.; Jeang, K.-T. Localization of human T-cell leukemia virus type 1 tax to subnuclear compartments that overlap with interchromatin speckles. J. Virol. 1996, 70, 6347–6357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bex, F.; McDowall, A.; Burny, A.; Gaynor, R. The human T-cell leukemia virus type 1 transactivator protein Tax colocalizes in unique nuclear structures with NF-kappaB proteins. J. Virol. 1997, 71, 3484–3497. [Google Scholar] [CrossRef] [Green Version]
- Bonnet, A.; Randrianarison-Huetz, V.; Nzounza, P.; Nedelec, M.; Chazal, M.; Waast, L.; Pene, S.; Bazarbachi, A.; Mahieux, R.; Bénit, L. Low nuclear body formation and tax SUMOylation do not prevent NF-kappaB promoter activation. Retrovirology 2012, 9, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasr, R.; Chiari, E.; El-Sabban, M.; Mahieux, R.; Kfoury, Y.; Abdulhay, M.; Yazbeck, V.; Hermine, O.; de Thé, H.; Pique, C. Tax ubiquitylation and sumoylation control critical cytoplasmic and nuclear steps of NF-κB activation. Blood 2006, 107, 4021–4029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voll, R.E.; Jimi, E.; Phillips, R.J.; Barber, D.F.; Rincon, M.; Hayday, A.C.; Flavell, R.A.; Ghosh, S. NF-κB activation by the pre-T cell receptor serves as a selective survival signal in T lymphocyte development. Immunity 2000, 13, 677–689. [Google Scholar] [CrossRef] [Green Version]
- Millen, S.; Meretuk, L.; Göttlicher, T.; Schmitt, S.; Fleckenstein, B.; Thoma-Kress, A.K. A novel positive feedback-loop between the HTLV-1 oncoprotein Tax and NF-κB activity in T-cells. Retrovirology 2020, 17, 1–19. [Google Scholar] [CrossRef]
- Peloponese Jr, J.-M.; Yasunaga, J.; Kinjo, T.; Watashi, K.; Jeang, K.-T. Peptidylproline cis-trans-isomerase Pin1 interacts with human T-cell leukemia virus type 1 tax and modulates its activation of NF-κB. J. Virol. 2009, 83, 3238–3248. [Google Scholar] [CrossRef] [Green Version]
- Haller, K.; Wu, Y.; Derow, E.; Schmitt, I.; Jeang, K.-T.; Grassmann, R. Physical Interaction of Human T-Cell Leukemia Virus Type 1 Tax with Cyclin-Dependent Kinase 4 Stimulates the Phosphorylation of Retinoblastoma Protein. Mol. Cell. Biol. 2002, 22, 3327–3338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araujo, A.; Hall, W.W. Human T-lymphotropic virus type II and neurological disease. Ann. Neurol. 2004, 56, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Turci, M.; Lodewick, J.; Righi, P.; Polania, A.; Romanelli, M.G.; Bex, F.; Bertazzoni, U. HTLV-2B Tax oncoprotein is modified by ubiquitination and sumoylation and displays intracellular localization similar to its homologue HTLV-1 Tax. Virology 2009, 386, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Dooneief, G.; Marlink, R.; Bell, K.; Marder, K.; Renjifo, B.; Stern, Y.; Mayeux, R. Neurologic consequences of HTLV-II infection in injection-drug users. Neurology 1996, 46, 1556–1560. [Google Scholar] [CrossRef]
- Hjelle, B.; Torrez-Martinez, N.; Mills, R.; Appenzeller, O.; Jahnke, R.; Alexander, S.; Ross, G. Chronic neurodegenerative disease associated with HTLV-II infection. Lancet 1992, 339, 645–646. [Google Scholar] [CrossRef]
- Murphy, E.L.; Fridey, J.; Smith, J.W.; Engstrom, J.; Sacher, R.A.; Miller, K.; Gibble, J.; Stevens, J.; Thomson, R.; Hansma, D.; et al. HTLV-associated myelopathy in a cohort of HTLV-I and HTLV-II-infected blood donors. Neurology 1997, 48, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Romanelli, M.G.; Diani, E.; Bergamo, E.; Casoli, C.; Ciminale, V.; Bex, F.; Bertazzoni, U. Highlights on distinctive structural and functional properties of HTLV Tax proteins. Front. Microbiol. 2013, 4, 271. [Google Scholar] [CrossRef] [Green Version]
- Eiraku, N.; Novoa, P.; da Costa Ferreira, M.; Monken, C.; Ishak, R.; da Costa Ferreira, O.; Zhu, S.W.; Lorenco, R.; Ishak, M.; Azvedo, V. Identification and characterization of a new and distinct molecular subtype of human T-cell lymphotropic virus type 2. J. Virol. 1996, 70, 1481–1492. [Google Scholar] [CrossRef] [Green Version]
- Endo, K.; Hirata, A.; Iwai, K.; Sakurai, M.; Fukushi, M.; Oie, M.; Higuchi, M.; Hall, W.W.; Gejyo, F.; Fujii, M. Human T-Cell Leukemia Virus Type 2 (HTLV-2) Tax Protein Transforms a Rat Fibroblast Cell Line but Less Efficiently than HTLV-1 Tax. J. Virol. 2002, 76, 2648–2653. [Google Scholar] [CrossRef] [Green Version]
- Lewis, M.J.; Sheehy, N.; Salemi, M.; VanDamme, A.-M.; Hall, W.W. Comparison of CREB-and NF-κB-mediated transactivation by human T lymphotropic virus type II (HTLV-II) and type I (HTLV-I) tax proteins. Virology 2002, 295, 182–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feuer, G.; Green, P.L. Comparative biology of human T-cell lymphotropic virus type 1 (HTLV-1) and HTLV-2. Oncogene 2005, 24, 5996–6004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoji, T.; Higuchi, M.; Kondo, R.; Takahashi, M.; Oie, M.; Tanaka, Y.; Aoyagi, Y.; Fujii, M. Identification of a novel motif responsible for the distinctive transforming activity of human T-cell leukemia virus (HTLV) type 1 Tax1 protein from HTLV-2 Tax2. Retrovirology 2009, 6, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertazzoni, U.; Turci, M.; Avesani, F.; Di Gennaro, G.; Bidoia, C.; Romanelli, M.G. Intracellular Localization and Cellular Factors Interaction of HTLV-1 and HTLV-2 Tax Proteins: Similarities and Functional Differences. Viruses 2011, 3, 541–560. [Google Scholar] [CrossRef] [Green Version]
- Pérès, E.; Blin, J.; Ricci, E.P.; Artesi, M.; Hahaut, V.; van den Broeke, A.; Corbin, A.; Gazzolo, L.; Ratner, L.; Jalinot, P.; et al. PDZ domain-binding motif of Tax sustains T-cell proliferation in HTLV-1-infected humanized mice. PLoS Pathog. 2018, 14, e1006933. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Yamamoto, B.; Haoudi, A.; Semmes, O.J.; Green, P.L. PDZ binding motif of HTLV-1 Tax promotes virus-mediated T-cell proliferation in vitro and persistence in vivo. Blood 2006, 107, 1980–1988. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.; Li, Z.; Schulze-Gahmen, U.; Stjepanovic, G.; Zhou, Q.; Hurley, J.H. Structural basis for ELL2 and AFF4 activation of HIV-1 proviral transcription. Nat. Commun. 2017, 8, 14076. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, L.; Stephens, A.; Nam, S.-Z.; Rau, D.; Kübler, J.; Lozajic, M.; Gabler, F.; Söding, J.; Lupas, A.N.; Alva, V. A Completely Reimplemented MPI Bioinformatics Toolkit with a New HHpred Server at its Core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef]
- Erdős, G.; Dosztányi, Z. Analyzing Protein Disorder with IUPred2A. Curr. Protoc. Bioinform. 2020, 70, e99. [Google Scholar] [CrossRef] [Green Version]
- Mészáros, B.; Erdős, G.; Dosztányi, Z. IUPred2A: Context-dependent prediction of protein disorder as a function of redox state and protein binding. Nucleic Acids Res. 2018, 46, W329–W337. [Google Scholar] [CrossRef]
- Kumar, M.; Gouw, M.; Michael, S.; Sámano-Sánchez, H.; Pancsa, R.; Glavina, J.; Diakogianni, A.; Valverde, J.A.; Bukirova, D.; Čalyševa, J.; et al. ELM—the eukaryotic linear motif resource in 2020. Nucleic Acids Res. 2019, 48, D296–D306. [Google Scholar] [CrossRef] [Green Version]
- Sobhian, B.; Laguette, N.; Yatim, A.; Nakamura, M.; Levy, Y.; Kiernan, R.; Benkirane, M. HIV-1 Tat Assembles a Multifunctional Transcription Elongation Complex and Stably Associates with the 7SK snRNP. Mol. Cell 2010, 38, 439–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.-L.; Adya, N.; Siores, E.; Gao, Q.; Giam, C.-Z. Cellular factors involved in transcription and Tax-mediated trans-activation directed by the TGACGT motifs in human T-cell leukemia virus type I promoter. J. Biol. Chem. 1990, 265, 20285–20292. [Google Scholar] [CrossRef]
- He, N.; Chan, C.K.; Sobhian, B.; Chou, S.; Xue, Y.; Liu, M.; Alber, T.; Benkirane, M.; Zhou, Q. Human Polymerase-Associated Factor complex (PAFc) connects the Super Elongation Complex (SEC) to RNA polymerase II on chromatin. Proc. Natl. Acad. Sci. USA 2011, 108, E636–E645. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.; Garrett, A.S.; De Kumar, B.; Smith, E.R.; Gogol, M.; Seidel, C.; Krumlauf, R.; Shilatifard, A. Dynamic transcriptional events in embryonic stem cells mediated by the super elongation complex (SEC). Genes Dev. 2011, 25, 1486–1498. [Google Scholar] [CrossRef] [Green Version]
- Thénoz, M.; Vernin, C.; Mortada, H.; Karam, M.; Pinatel, C.; Gessain, A.; Webb, T.R.; Auboeuf, D.; Wattel, E.; Mortreux, F. HTLV-1-infected CD4+ T-cells display alternative exon usages that culminate in adult T-cell leukemia. Retrovirology 2014, 11, 119. [Google Scholar] [CrossRef] [Green Version]
- Semmes, O.J.; Jeang, K.-T. HTLV-I tax is a zinc-binding protein: Role of zinc in tax structure and function. Virology 1992, 188, 754–764. [Google Scholar] [CrossRef]
- Smith, M.R.; Greene, W.C. Characterization of a novel nuclear localization signal in the HTLV-I tax transactivator protein. Virology 1992, 187, 316–320. [Google Scholar] [CrossRef]
- Goren, I.; Semmes, O.J.; Jeang, K.-T.; Moelling, K. The amino terminus of Tax is required for interaction with the cyclic AMP response element binding protein. J. Virol. 1995, 69, 5806–5811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boxus, M.; Twizere, J.-C.; Legros, S.; Dewulf, J.-F.; Kettmann, R.; Willems, L. The HTLV-1 Tax interactome. Retrovirology 2008, 5, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alefantis, T.; Barmak, K.; Harhaj, E.W.; Grant, C.; Wigdahl, B. Characterization of a Nuclear Export Signal within the Human T Cell Leukemia Virus Type I Transactivator Protein Tax. J. Biol. Chem. 2003, 278, 21814–21822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, M.R.; Greene, W.C. Identification of HTLV-I tax trans-activator mutants exhibiting novel transcriptional phenotypes. Genes Dev. 1990, 4, 1875–1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basbous, J.; Bazarbachi, A.; Granier, C.; Devaux, C.; Mesnard, J.-M. TheCentral Region of Human T-Cell Leukemia Virus Type 1 TaxProtein Contains Distinct Domains Involved inSubunitDimerization. J. Virol. 2003, 77, 13028–13035. [Google Scholar] [CrossRef] [Green Version]
- Chevalier, S.; Meertens, L.; Pise-Masison, C.; Calattini, S.; Park, H.; Alhaj, A.; Zhou, M.; Gessain, A.; Kashanchi, F.; Brady, J.N.; et al. The tax protein from the primate T-cell lymphotropic virus type 3 is expressed in vivo and is functionally related to HTLV-1 Tax rather than HTLV-2 Tax. Oncogene 2006, 25, 4470–4482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higuchi, M.; Fujii, M. Distinct functions of HTLV-1 Tax1 from HTLV-2 Tax2 contribute key roles to viral pathogenesis. Retrovirology 2009, 6, 117. [Google Scholar] [CrossRef] [Green Version]
- Lodewick, J.; Lamsoul, I.; Bex, F. Move or Die: The Fate of the Tax Oncoprotein of HTLV-1. Viruses 2011, 3, 829–857. [Google Scholar] [CrossRef] [Green Version]
- Lodewick, J.; Lamsoul, I.; Polania, A.; Lebrun, S.; Burny, A.; Ratner, L.; Bex, F. Acetylation of the human T-cell leukemia virus type 1 Tax oncoprotein by p300 promotes activation of the NF-κB pathway. Virology 2009, 386, 68–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, W.-K.; Zhou, M.; Jang, M.K.; Huang, K.; Jeong, S.-J.; Ozato, K.; Brady, J.N. Modulation of the Brd4/P-TEFb interaction by the human T-lymphotropic virus type 1 tax protein. J. Virol. 2007, 81, 11179–11186. [Google Scholar] [CrossRef] [Green Version]
- Hirata, A.; Higuchi, M.; Niinuma, A.; Ohashi, M.; Fukushi, M.; Oie, M.; Akiyama, T.; Tanaka, Y.; Gejyo, F.; Fujii, M. PDZ domain-binding motif of human T-cell leukemia virus type 1 Tax oncoprotein augments the transforming activity in a rat fibroblast cell line. Virology 2004, 318, 327–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousset, R.; Fabre, S.; Desbois, C.; Bantignies, F.; Jalinot, P. The C-terminus of the HTLV-1 Tax oncoprotein mediates interaction with the PDZ domain of cellular proteins. Oncogene 1998, 16, 643–654. [Google Scholar] [CrossRef] [Green Version]
- Ota, T.; Suzuki, Y.; Nishikawa, T.; Otsuki, T.; Sugiyama, T.; Irie, R.; Wakamatsu, A.; Hayashi, K.; Sato, H.; Nagai, K.; et al. Complete sequencing and characterization of 21,243 full-length human cDNAs. Nat. Genet. 2004, 36, 40–45. [Google Scholar] [CrossRef]
- Liu, M.; Hsu, J.; Chan, C.; Li, Z.; Zhou, Q. The Ubiquitin Ligase Siah1 Controls ELL2 Stability and Formation of Super Elongation Complexes to Modulate Gene Transcription. Mol. Cell 2012, 46, 325–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daub, H.; Olsen, J.; Bairlein, M.; Gnad, F.; Oppermann, F.S.; Körner, R.; Greff, Z.; Kéri, G.; Stemmann, O.; Mann, M. Kinase-Selective Enrichment Enables Quantitative Phosphoproteomics of the Kinome across the Cell Cycle. Mol. Cell 2008, 31, 438–448. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.V.; Vermeulen, M.; Santamaria, A.; Kumar, C.; Miller, M.L.; Jensen, L.J.; Gnad, F.; Cox, J.; Jensen, T.S.; Nigg, E.A.; et al. Quantitative Phosphoproteomics Reveals Widespread Full Phosphorylation Site Occupancy During Mitosis. Sci. Signal. 2010, 3, ra3. [Google Scholar] [CrossRef] [PubMed]
- Rigbolt, K.T.G.; Prokhorova, T.A.; Akimov, V.; Henningsen, J.; Johansen, P.T.; Kratchmarova, I.; Kassem, M.; Mann, M.; Olsen, J.V.; Blagoev, B. System-Wide Temporal Characterization of the Proteome and Phosphoproteome of Human Embryonic Stem Cell Differentiation. Sci. Signal. 2011, 4, rs3. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Liu, R.; Yang, G.; Zhou, Q. The PARP1-Siah1 Axis Controls HIV-1 Transcription and Expression of Siah1 Substrates. Cell Rep. 2018, 23, 3741–3749. [Google Scholar] [CrossRef] [PubMed]
- Langton, B.; Sliwkowski, M.; Tran, K.; Knapp, S.; Keitelmann, E.; Smith, C.; Wallingford, S.; Liu, H.; Ralston, J.; Brandis, J. Development and characterization of monoclonal antibodies to the HTLV-I Tax (P40X) protein. Med. Virol. 1988, 8, 295. [Google Scholar]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kohrt, S.; Strobel, S.; Mann, M.C.; Sticht, H.; Fleckenstein, B.; Thoma-Kress, A.K. Characterizing the Interaction between the HTLV-1 Transactivator Tax-1 with Transcription Elongation Factor ELL2 and Its Impact on Viral Transactivation. Int. J. Mol. Sci. 2021, 22, 13597. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413597
Kohrt S, Strobel S, Mann MC, Sticht H, Fleckenstein B, Thoma-Kress AK. Characterizing the Interaction between the HTLV-1 Transactivator Tax-1 with Transcription Elongation Factor ELL2 and Its Impact on Viral Transactivation. International Journal of Molecular Sciences. 2021; 22(24):13597. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413597
Chicago/Turabian StyleKohrt, Stephan, Sarah Strobel, Melanie C. Mann, Heinrich Sticht, Bernhard Fleckenstein, and Andrea K. Thoma-Kress. 2021. "Characterizing the Interaction between the HTLV-1 Transactivator Tax-1 with Transcription Elongation Factor ELL2 and Its Impact on Viral Transactivation" International Journal of Molecular Sciences 22, no. 24: 13597. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413597