
Effects of Microvesicles Derived from NK Cells Stimulated with IL-1β on the Phenotype and Functional Activity of Endothelial Cells

 , , , ,
, , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
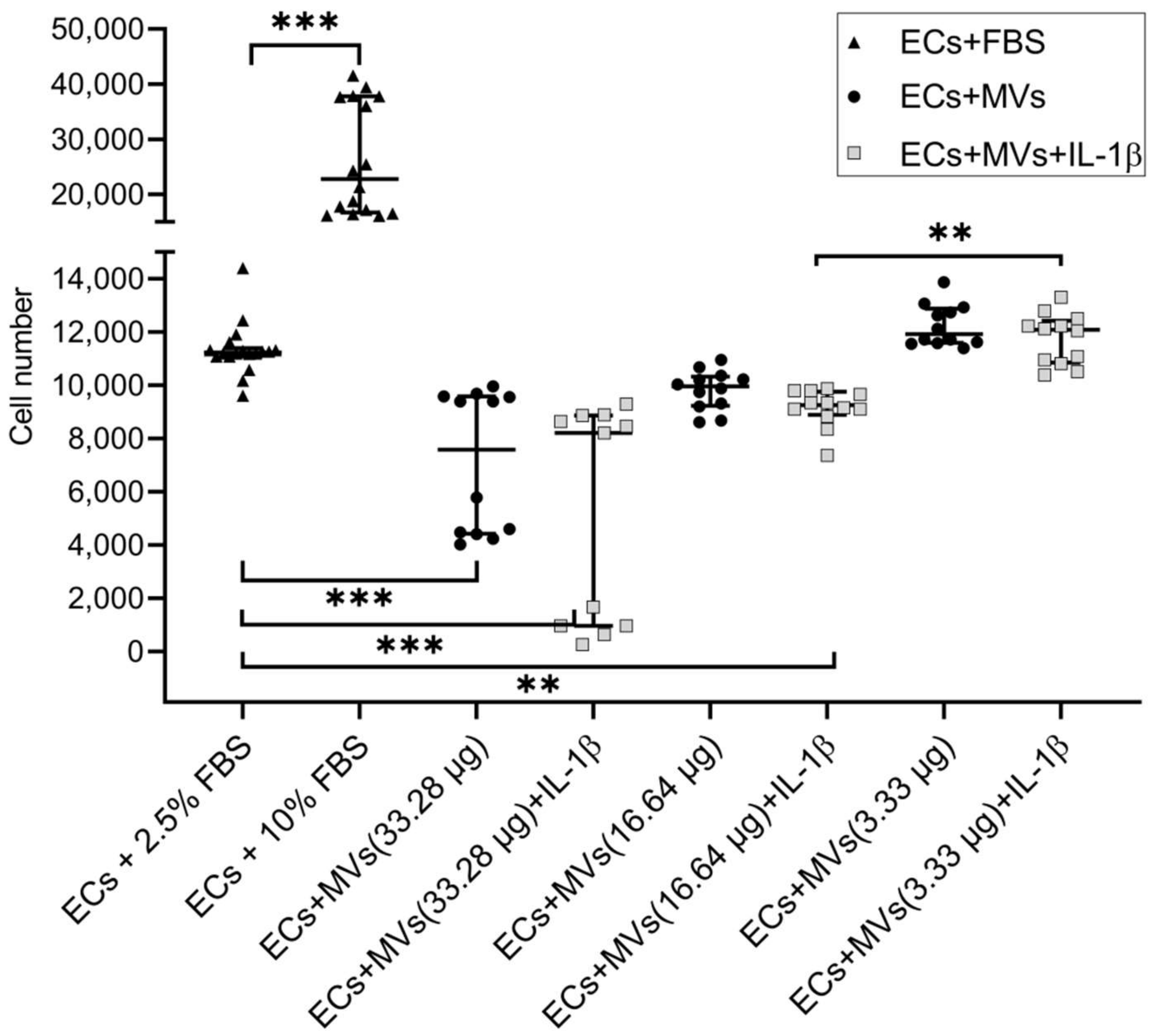
2.1. Effects of MVs Derived from NK Cells of the NK-92 Cell Line Stimulated with IL-1β on the Proliferative Activity of Endothelial Cells of the EA.hy926 Cell Line
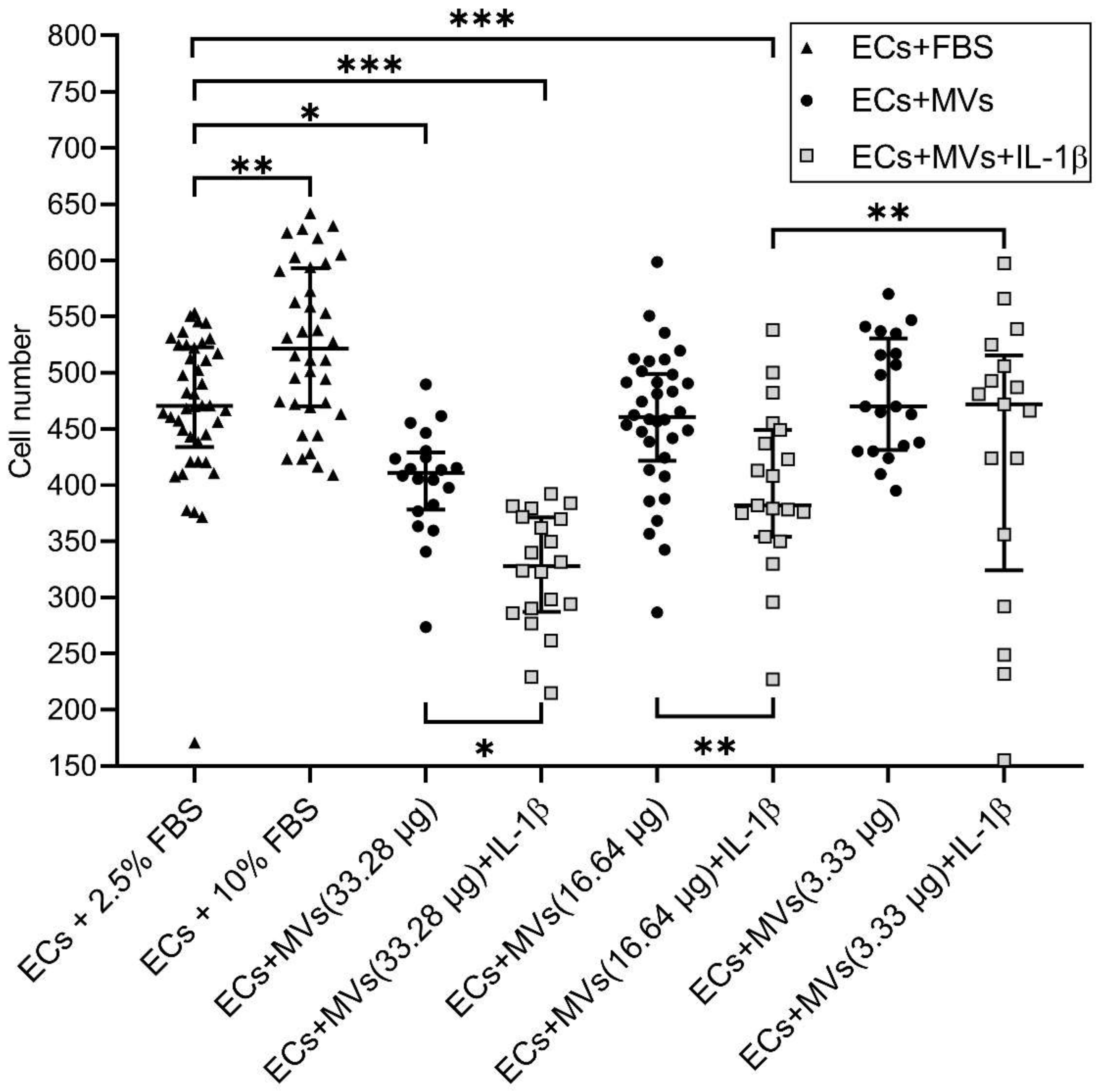
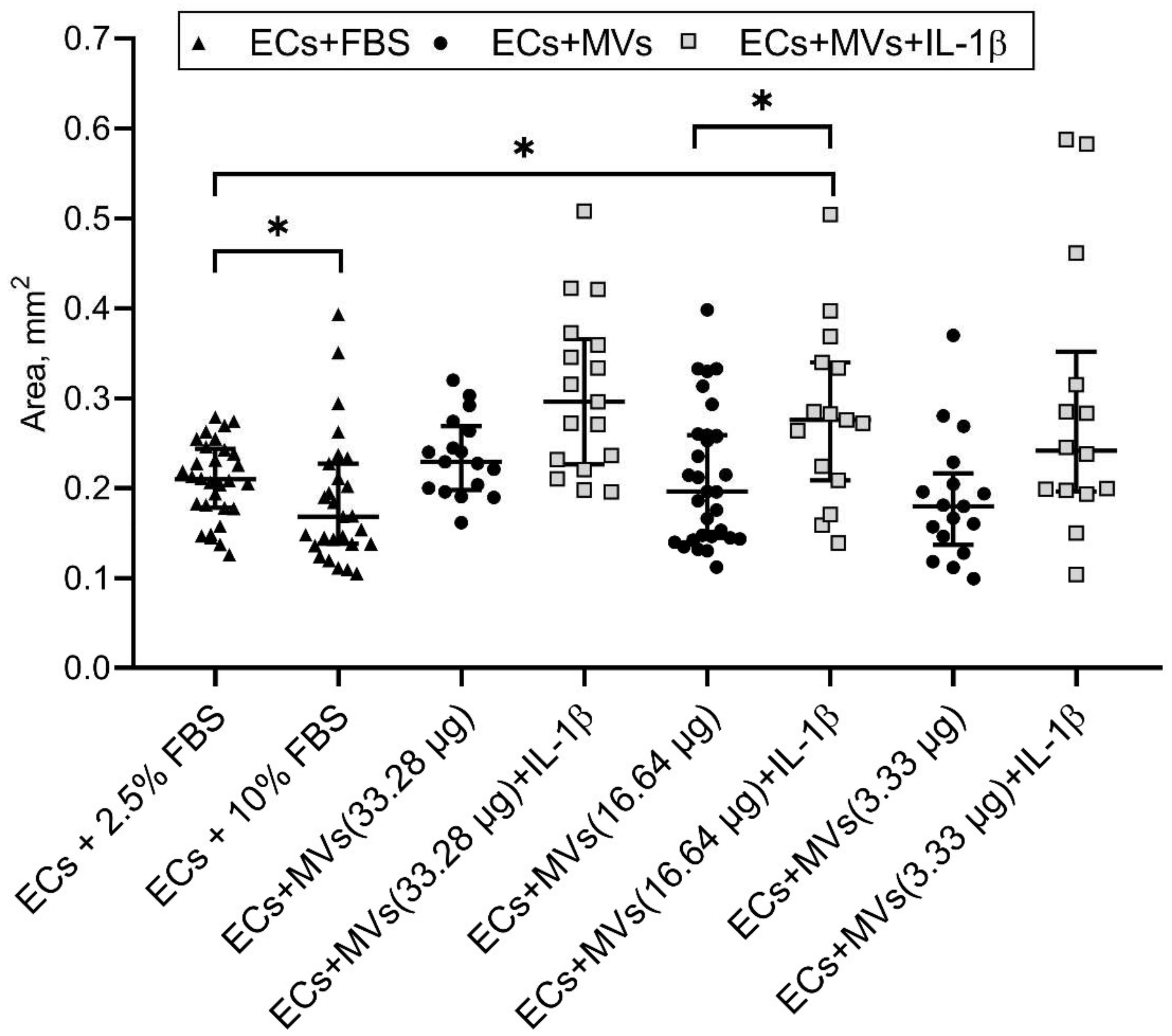
2.2. Effects of MVs Derived from NK Cells of the NK-92 Cell Line Stimulated with IL-1β on the Migratory Activity of Endothelial Cells of the EA.hy926 Cell Line
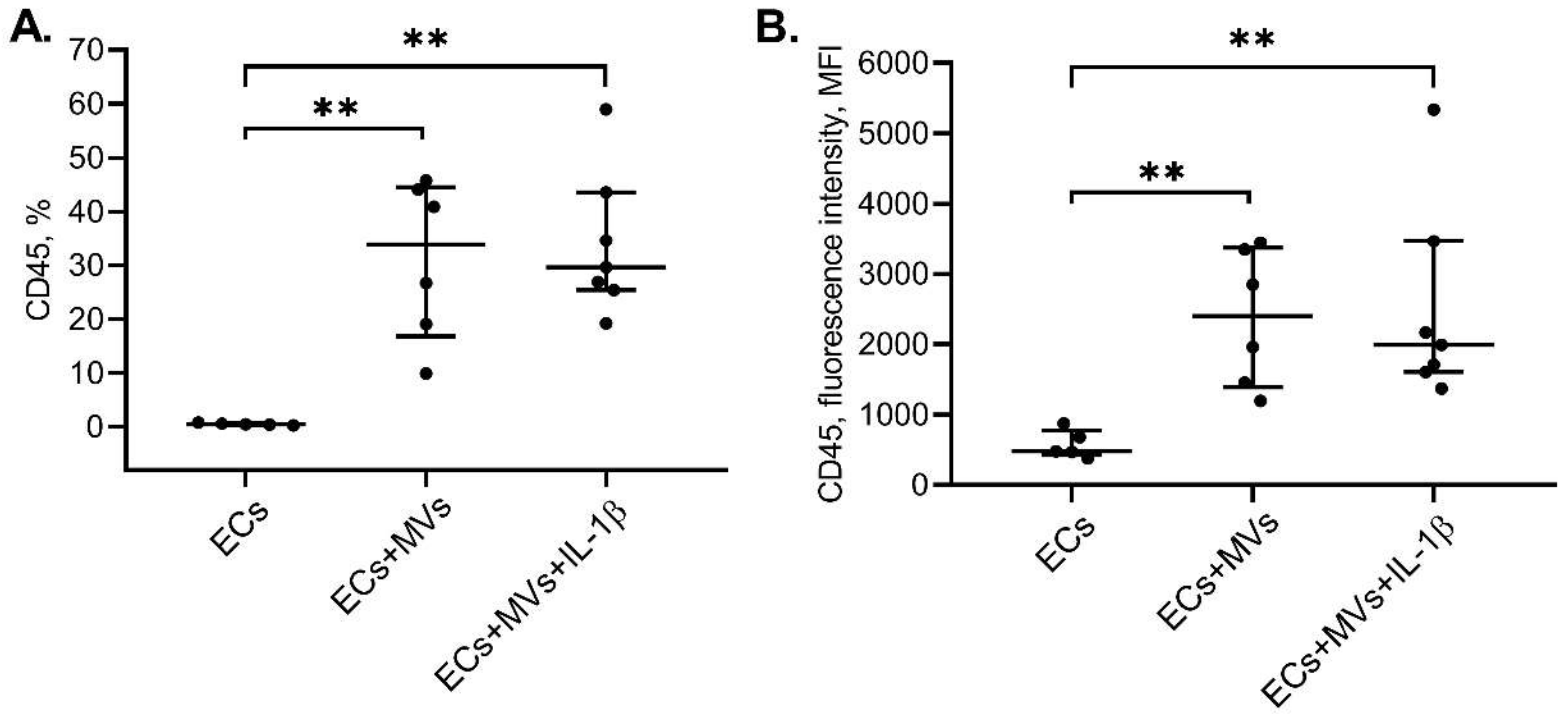
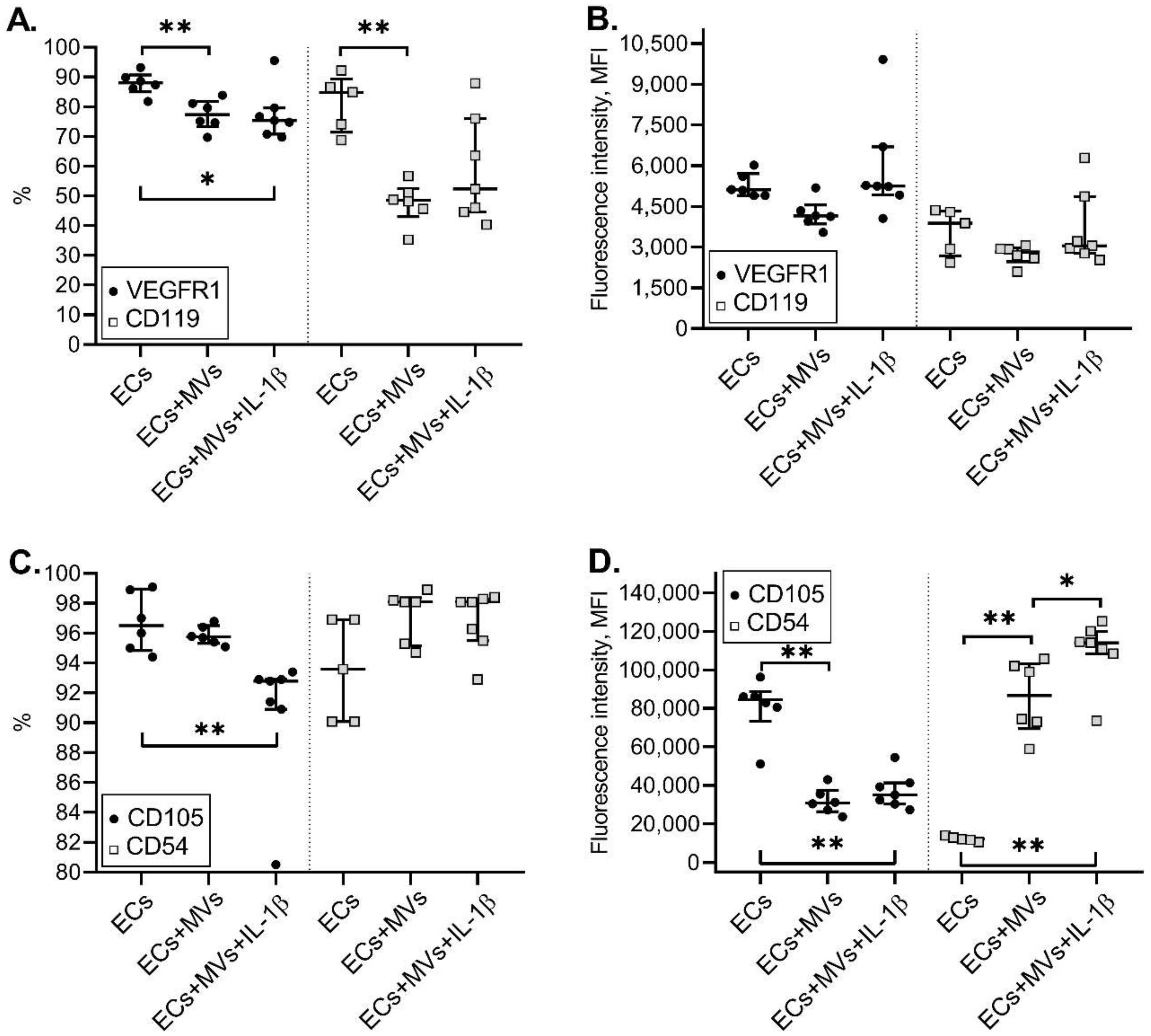
2.3. Effects of MVs Derived from NK-92 Cells Stimulated with IL-1β on the Phenotype of ECs of the Ea.hy926 Cell Line
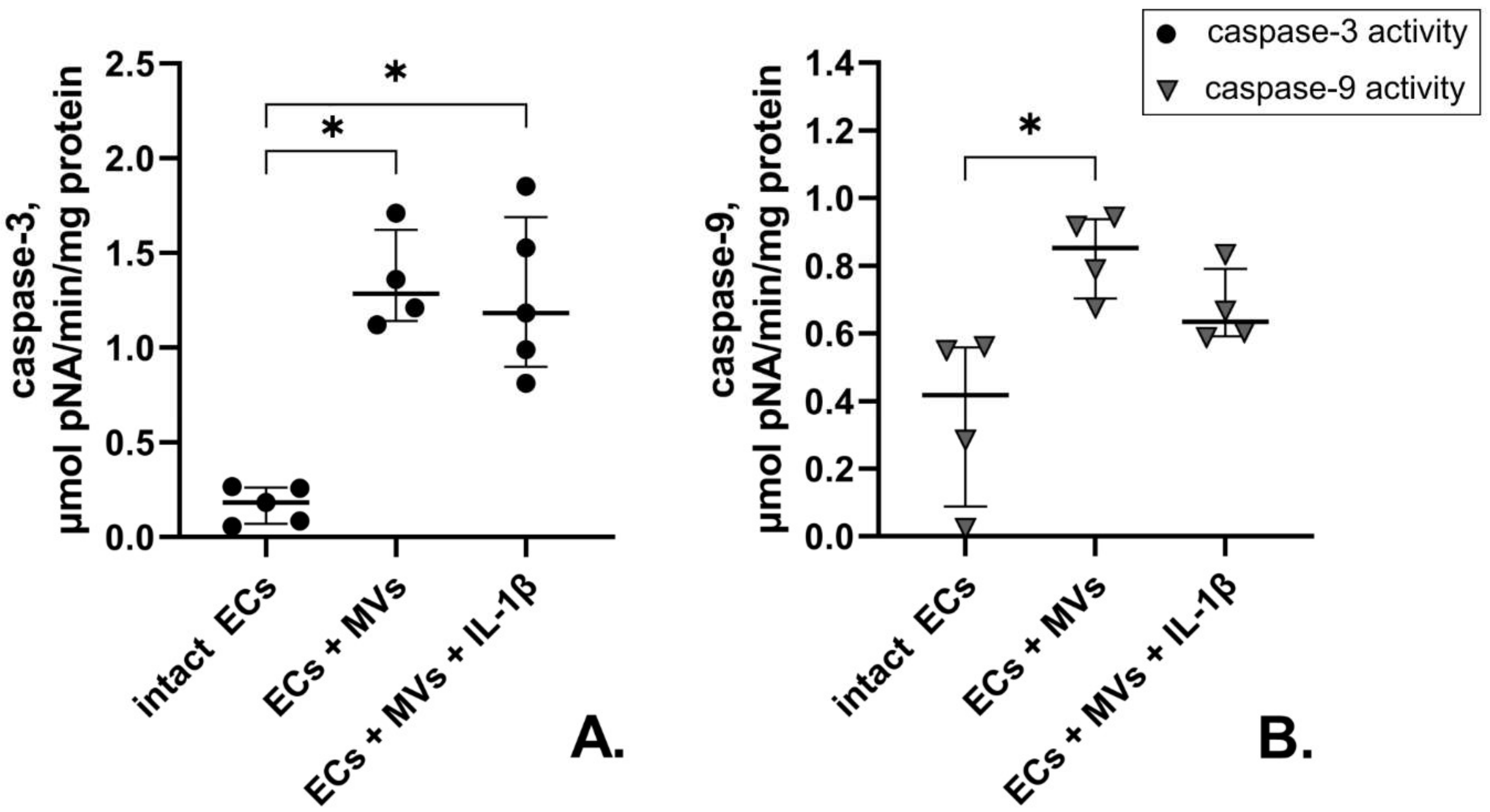
2.4. Effects of MVs Derived from Cells of the NK-92 Cell Line Stimulated with IL-1β on Caspase-3 and Caspase-9 Activity in Endothelial Cells of the EA.hy926 Cell Line
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. MV Isolation
4.3. MV Analysis by Dynamic Light Scattering
4.4. Analysis of Total Protein Content of Cell and MV Lysates
4.5. Preparation of MV Dilutions
4.6. Evaluation of the Effect of MVs Derived from Cells of the NK-92 Cell Line on the Migratory Activity of Endothelial Cells of the EA.hy926 Cell Line
4.7. Evaluation of the Effect of MVs Derived from Cells of the NK-92 Cell Line on the Proliferative Activity of Endothelial Cells of the EA.Hy92 Cell Line
4.8. Evaluation of the Effect of MVs Derived from Cells of the NK-92 Cell Line on the Phenotype of Endothelial Cells of the Ea.hy926 Cell Line
4.9. Evaluation of the Eeffect of MVs Derived from Cells of the NK-92 Cell Line on Caspase Activity in ECs of the Ea.Hy926 Cell Line
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Antonyak, M.A.; Cerione, R.A. Microvesicles as mediators of intercellular communication in cancer. Methods Mol. Biol. 2014, 1165, 147–173. [Google Scholar] [CrossRef] [PubMed]
- Sedgwick, A.E.; D’Souza-Schorey, C. The biology of extracellular microvesicles. Traffic 2018, 19, 319–327. [Google Scholar] [CrossRef]
- Akers, J.C.; Gonda, D.; Kim, R.; Carter, B.S.; Chen, C.C. Biogenesis of extracellular vesicles (EV): Exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J. Neuro-Oncol. 2013, 113, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ciardiello, C.; Cavallini, L.; Spinelli, C.; Yang, J.; Reis-Sobreiro, M.; De Candia, P.; Minciacchi, V.R.; Di Vizio, D. Focus on Extracellular Vesicles: New Frontiers of Cell-to-Cell Communication in Cancer. Int. J. Mol. Sci. 2016, 17, 175. [Google Scholar] [CrossRef] [Green Version]
- Pegtel, D.M.; Peferoen, L.; Amor, S. Extracellular vesicles as modulators of cell-to-cell communication in the healthy and diseased brain. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokolov, D.I.; Ovchinnikova, O.M.; Korenkov, D.A.; Viknyanschuk, A.N.; Benken, K.A.; Onokhin, K.V.; Selkov, S.A. Influence of peripheral blood microparticles of pregnant women with preeclampsia on the phenotype of monocytes. Transl. Res. J. Lab. Clin. Med. 2016, 170, 112–123. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasgupta, S.K.; Abdel-Monem, H.; Niravath, P.; Le, A.; Bellera, R.V.; Langlois, K.; Nagata, S.; Rumbaut, R.E.; Thiagarajan, P. Lactadherin and clearance of platelet-derived microvesicles. Blood 2009, 113, 1332–1339. [Google Scholar] [CrossRef] [Green Version]
- Mause, S.F.; Weber, C. Microparticles: Protagonists of a novel communication network for intercellular information exchange. Circ. Res. 2010, 107, 1047–1057. [Google Scholar] [CrossRef]
- D’Souza-Schorey, C.; Clancy, J.W. Tumor-derived microvesicles: Shedding light on novel microenvironment modulators and prospective cancer biomarkers. Genes Dev. 2012, 26, 1287–1299. [Google Scholar] [CrossRef] [Green Version]
- Kalra, H.; Drummen, G.P.C.; Mathivanan, S. Focus on Extracellular Vesicles: Introducing the Next Small Big Thing. Int. J. Mol. Sci. 2016, 17, 170. [Google Scholar] [CrossRef] [Green Version]
- Tricarico, C.; Clancy, J.; D’Souza-Schorey, C. Biology and biogenesis of shed microvesicles. Small GTPases 2017, 8, 220–232. [Google Scholar] [CrossRef] [Green Version]
- Martinez de Lizarrondo, S.; Roncal, C.; Calvayrac, O.; Rodríguez, C.; Varo, N.; Purroy, A.; Lorente, L.; Rodríguez, J.A.; Doeuvre, L.; Hervás-Stubbs, S.; et al. Synergistic Effect of Thrombin and CD40 Ligand on Endothelial Matrix Metalloproteinase-10 Expression and Microparticle Generation In Vitro and In Vivo. Arter. Thromb. Vasc. Biol. 2012, 32, 1477–1487. [Google Scholar] [CrossRef] [Green Version]
- Del Conde, I.; Shrimpton, C.N.; Thiagarajan, P.; López, J.A. Tissue-factor–bearing microvesicles arise from lipid rafts and fuse with activated platelets to initiate coagulation. Blood 2005, 106, 1604–1611. [Google Scholar] [CrossRef]
- Pluskota, E.; Woody, N.M.; Szpak, D.; Ballantyne, C.M.; Soloviev, D.A.; Simon, D.I.; Plow, E.F. Expression, activation, and function of integrin alphaMbeta2 (Mac-1) on neutrophil-derived microparticles. Blood 2008, 112, 2327–2335. [Google Scholar] [CrossRef] [Green Version]
- Bernimoulin, M.; Waters, E.K.; Foy, M.; Steele, B.M.; Sullivan, M.; Falet, H.; Walsh, M.T.; Barteneva, N.; Geng, J.-G.; Hartwig, J.H.; et al. Differential stimulation of monocytic cells results in distinct populations of microparticles. J. Thromb. Haemost. 2009, 7, 1019–1028. [Google Scholar] [CrossRef] [Green Version]
- Keerthikumar, S.; Gangoda, L.; Liem, M.; Fonseka, P.; Atukorala, I.; Ozcitti, C.; Mechler, A.; Adda, C.G.; Ang, C.-S.; Mathivanan, S. Proteogenomic analysis reveals exosomes are more oncogenic than ectosomes. Oncotarget 2015, 6, 15375–15396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratajczak, J.; Miękus, K.; Kucia, M.; Zhang, J.; Reca, R.; Dvorak, P.; Ratajczak, M.Z. Embryonic stem cell-derived microvesicles reprogram hematopoietic progenitors: Evidence for horizontal transfer of mRNA and protein delivery. Leukemia 2006, 20, 847–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, C.T., Jr.; Kurre, P. Vesicle Trafficking and RNA Transfer Add Complexity and Connectivity to Cell–Cell Communication. Cancer Res. 2013, 73, 3200–3205. [Google Scholar] [CrossRef] [Green Version]
- Waldenström, A.; Gennebäck, N.; Hellman, U.; Ronquist, G. Cardiomyocyte Microvesicles Contain DNA/RNA and Convey Biological Messages to Target Cells. PLoS ONE 2012, 7, e34653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hugel, B.; Martínez, M.C.; Kunzelmann, C.; Freyssinet, J.-M. Membrane Microparticles: Two Sides of the Coin. Physiology 2005, 20, 22–27. [Google Scholar] [CrossRef]
- Kreger, B.T.; Dougherty, A.L.; Greene, K.S.; Cerione, R.A.; Antonyak, M.A. Microvesicle Cargo and Function Changes upon Induction of Cellular Transformation. J. Biol. Chem. 2016, 291, 19774–19785. [Google Scholar] [CrossRef] [Green Version]
- Piccin, A.; Murphy, W.G.; Smith, O.P. Circulating microparticles: Pathophysiology and clinical implications. Blood Rev. 2007, 21, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Budaj, M.; Poljak, Z.; Ďuriš, I.; Kaško, M.; Imrich, R.; Kopáni, M.; Maruščáková, L.; Hulín, I. Microparticles: A component of various diseases. Pol. Arch. Med. Wewn. 2012, 122 (Suppl. S1), 24–29. [Google Scholar] [CrossRef] [Green Version]
- Cooper, M.A.; Fehniger, T.; Caligiuri, M.A. The biology of human natural killer-cell subsets. Trends Immunol. 2001, 22, 633–640. [Google Scholar] [CrossRef]
- Thiruchelvam, U.; Wingfield, M.; O’Farrelly, C. Natural Killer Cells: Key Players in Endometriosis. Am. J. Reprod. Immunol. 2015, 74, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Chazara, O.; Xiong, S.; Moffett, A. Maternal KIR and fetal HLA-C: A fine balance. J. Leukoc. Biol. 2011, 90, 703–716. [Google Scholar] [CrossRef]
- Male, V.; Sharkey, A.; Masters, L.; Kennedy, P.R.; Farrell, L.E.; Moffett, A. The effect of pregnancy on the uterine NK cell KIR repertoire. Eur. J. Immunol. 2011, 41, 3017–3027. [Google Scholar] [CrossRef] [Green Version]
- EL Costa, H.; Tabiasco, J.; Berrebi, A.; Parant, O.; Aguerre-Girr, M.; Piccinni, M.-P.; Le Bouteiller, P. Effector functions of human decidual NK cells in healthy early pregnancy are dependent on the specific engagement of natural cytotoxicity receptors. J. Reprod. Immunol. 2009, 82, 142–147. [Google Scholar] [CrossRef]
- Klingemann, H. Challenges of cancer therapy with natural killer cells. Cytotherapy 2015, 17, 245–249. [Google Scholar] [CrossRef]
- Jong, A.Y.; Wu, C.-H.; Li, J.; Sun, J.; Fabbri, M.; Wayne, A.S.; Seeger, R.C. Large-scale isolation and cytotoxicity of extracellular vesicles derived from activated human natural killer cells. J. Extracell. Vesicles 2017, 6, 1294368. [Google Scholar] [CrossRef] [Green Version]
- Shoae-Hassani, A.; Hamidieh, A.A.; Behfar, M.; Mohseni, R.; Mortazavi-Tabatabaei, S.A.; Asgharzadeh, S. NK Cell–derived Exosomes From NK Cells Previously Exposed to Neuroblastoma Cells Augment the Antitumor Activity of Cytokine-activated NK Cells. J. Immunother. 2017, 40, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Timonen, T. Natural killer cells: Endothelial interactions, migration, and target cell recognition. J. Leukoc. Biol. 1997, 62, 693–701. [Google Scholar] [CrossRef] [Green Version]
- Farag, S.S.; Caligiuri, M.A. Human natural killer cell development and biology. Blood Rev. 2006, 20, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Naruse, K.; Lash, G.E.; Bulmer, J.N.; Innes, B.A.; Otun, H.A.; Searle, R.F.; Robson, S.C. The Urokinase Plasminogen Activator (uPA) System in Uterine Natural Killer Cells in the Placental Bed During Early Pregnancy. Placenta 2009, 30, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Lash, G.E.; Robson, S.C.; Bulmer, J.N. Review: Functional role of uterine natural killer (uNK) cells in human early pregnancy decidua. Placenta 2010, 31, S87–S92. [Google Scholar] [CrossRef]
- Rakocevic, J.; Orlic, D.; Mitrovic-Ajtic, O.; Tomasevic, M.; Dobric, M.; Zlatic, N.; Milasinovic, D.; Stankovic, G.; Ostojić, M.; Labudovic-Borovic, M. Endothelial cell markers from clinician’s perspective. Exp. Mol. Pathol. 2017, 102, 303–313. [Google Scholar] [CrossRef]
- Aird, W.C. Phenotypic Heterogeneity of the Endothelium: I. Structure, function, and mechanisms. Circ. Res. 2007, 100, 158–173. [Google Scholar] [CrossRef]
- Gerber, H.-P.; McMurtrey, A.; Kowalski, J.; Yan, M.; Keyt, B.A.; Dixit, V.; Ferrara, N. Vascular Endothelial Growth Factor Regulates Endothelial Cell Survival through the Phosphatidylinositol 3′-Kinase/Akt Signal Transduction Pathway. Requirement for Flk-1/KDR activation. J. Biol. Chem. 1998, 273, 30336–30343. [Google Scholar] [CrossRef] [Green Version]
- Ashkar, A.A.; Di Santo, J.P.; Croy, B.A. Interferon gamma Contributes to Initiation of Uterine Vascular Modification, Decidual Integrity, and Uterine Natural Killer Cell Maturation during Normal Murine Pregnancy. J. Exp. Med. 2000, 192, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Murphy, S.P.; Tayade, C.; Ashkar, A.A.; Hatta, K.; Zhang, J.; Croy, B.A. Interferon Gamma in Successful Pregnancies. Biol. Reprod. 2009, 80, 848–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markova, K.L.; Mikhailova, V.A.; Korenevsky, A.V.; Milyutina, Y.P.; Rodygina, V.V.; Aleksandrova, E.P.; Markov, A.S.; Balabas, O.A.; Selkov, S.A.; Sokolov, D.I. Microvesicles produced by natural killer cells of the NK-92 cell line affect the phenotype and functions of endothelial cells of the EA.Hy926 cell line. Med. Immunol. 2020, 22, 249–268. [Google Scholar] [CrossRef]
- Elkabets, M.; Ribeiro, V.S.G.; Dinarello, C.A.; Ostrand-Rosenberg, S.; Di Santo, J.P.; Apte, R.N.; Vosshenrich, C.A.J. IL-1beta regulates a novel myeloid-derived suppressor cell subset that impairs NK cell development and function. Eur. J. Immunol. 2010, 40, 3347–3357. [Google Scholar] [CrossRef]
- Van Der Pol, E.; Coumans, F.; Varga, Z.; Krumrey, M.; Nieuwland, R. Innovation in detection of microparticles and exosomes. J. Thromb. Haemost. 2013, 11, 36–45. [Google Scholar] [CrossRef] [Green Version]
- Van Der Pol, E.; Coumans, F.A.W.; Grootemaat, A.E.; Gardiner, C.; Sargent, I.L.; Harrison, P.; Sturk, A.; van Leeuwen, T.G.; Nieuwland, R. Particle size distribution of exosomes and microvesicles determined by transmission electron microscopy, flow cytometry, nanoparticle tracking analysis, and resistive pulse sensing. J. Thromb. Haemost. 2014, 12, 1182–1192. [Google Scholar] [CrossRef] [PubMed]
- Klingemann, H.; Boissel, L.; Toneguzzo, F. Natural Killer Cells for Immunotherapy—Advantages of the NK-92 Cell Line over Blood NK Cells. Front. Immunol. 2016, 7, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulcahy, L.A.; Pink, R.C.; Carter, D.R.F. Routes and mechanisms of extracellular vesicle uptake. J. Extracell. Vesicles 2014, 3, 24641. [Google Scholar] [CrossRef] [Green Version]
- Rozmyslowicz, T.; Majka, M.; Kijowski, J.; Murphy, S.L.; Conover, D.O.; Poncz, M.; Ratajczak, J.; Gaulton, G.N.; Ratajczak, M.Z. Platelet- and megakaryocyte-derived microparticles transfer CXCR4 receptor to CXCR4-null cells and make them susceptible to infection by X4-HIV. Aids 2003, 17, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Christianson, H.C.; Svensson, K.J.; van Kuppevelt, T.H.; Li, J.-P.; Belting, M. Cancer cell exosomes depend on cell-surface heparan sulfate proteoglycans for their internalization and functional activity. Proc. Natl. Acad. Sci. USA 2013, 110, 17380–17385. [Google Scholar] [CrossRef] [Green Version]
- Sokolov, D.I.; Markova, K.L.; Mikhailova, V.A.; Vyazmina, L.P.; Milyutina, Y.P.; Kozyreva, A.R.; Zhdanova, A.A.; Malygina, D.A.; Onokhin, K.V.; Ivanova, A.N.; et al. Phenotypic and Functional Characteristics of Microvesicles Produced by Natural Killer Cells. Med. Immunol. 2019, 21, 669–688. [Google Scholar] [CrossRef]
- Bevilacqua, M.P.; Pober, J.S.; Wheeler, M.E.; Cotran, R.S.; Gimbrone, M.A., Jr. Interleukin 1 acts on cultured human vascular endothelium to increase the adhesion of polymorphonuclear leukocytes, monocytes, and related leukocyte cell lines. J. Clin. Investig. 1985, 76, 2003–2011. [Google Scholar] [CrossRef]
- Kawakami, A.; Hida, A.; Yamasaki, S.; Miyashita, T.; Nakashima, K.; Tanaka, F.; Ida, H.; Furuyama, M.; Migita, K.; Origuchi, T.; et al. Modulation of the expression of membrane-bound CD54 (mCD54) and soluble form of CD54 (sCD54) in endothelial cells by glucosyl transferase inhibitor: Possible role of ceramide for the shedding of mCD54. Biochem. Biophys. Res. Commun. 2002, 296, 26–31. [Google Scholar] [CrossRef]
- van de Stolpe, A.; van der Saag, P.T. Intercellular adhesion molecule-1. J. Mol. Med. (Berl.) 1996, 74, 13–33. [Google Scholar] [CrossRef] [PubMed]
- Lugini, L.; Cecchetti, S.; Huber, V.; Luciani, F.; Macchia, G.; Spadaro, F.; Paris, L.; Abalsamo, L.; Colone, M.; Molinari, A.; et al. Immune surveillance properties of human NK cell-derived exosomes. J. Immunol. 2012, 189, 2833–2842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Susanto, O.; Trapani, J.A.; Brasacchio, D. Controversies in granzyme biology. Tissue Antigens 2012, 80, 477–487. [Google Scholar] [CrossRef]
- Chen, P.-L.; Easton, A. Apoptotic Phenotype Alters the Capacity of Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand to Induce Human Vascular Endothelial Activation. J. Vasc. Res. 2008, 45, 111–122. [Google Scholar] [CrossRef]
- Gong, J.H.; Maki, G.; Klingemann, H.G. Characterization of a human cell line (NK-92) with phenotypical and functional characteristics of activated natural killer cells. Leukemia 1994, 8, 652–658. [Google Scholar]
- Komatsu, F.; Kajiwara, M. Relation of natural killer cell line NK-92-mediated cytolysis (NK-92-lysis) with the surface markers of major histocompatibility complex class I antigens, adhesion molecules, and Fas of target cells. Oncol. Res. 1998, 10, 483–489. [Google Scholar]
- Thornhill, M.H.; Li, J.; Haskard, D.O. Leucocyte Endothelial Cell Adhesion: A Study comparing Human Umbilical Vein Endothelial Cells and the Endothelial Cell Line EA-hy-926. Scand. J. Immunol. 1993, 38, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Edgell, C.J.; McDonald, C.C.; Graham, J.B. Permanent cell line expressing human factor VIII-related antigen established by hybridization. Proc. Natl. Acad. Sci. USA 1983, 80, 3734–3737. [Google Scholar] [CrossRef] [Green Version]
- Riesbeck, K.; Billström, A.; Tordsson, J.; Brodin, T.; Kristensson, K.; Dohlsten, M. Endothelial Cells Expressing an Inflammatory Phenotype Are Lysed by Superantigen-Targeted Cytotoxic T Cells. Clin. Diagn. Lab. Immunol. 1998, 5, 675–682. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Kaslan, M.; Lee, S.H.; Yao, J.; Gao, Z. Progress in Exosome Isolation Techniques. Theranostics 2017, 7, 789–804. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Greening, D.W.; Zhu, H.-J.; Takahashi, N.; Simpson, R.J. Extracellular vesicle isolation and characterization: Toward clinical application. J. Clin. Investig. 2016, 126, 1152–1162. [Google Scholar] [CrossRef] [Green Version]
- Simak, J.; Gelderman, M.P.; Yu, H.; Wright, V.; Baird, A.E. Circulating endothelial microparticles in acute ischemic stroke: A link to severity, lesion volume and outcome. J. Thromb. Haemost. 2006, 4, 1296–1302. [Google Scholar] [CrossRef]
- Korenevskii, A.V.; Milyutina, Y.P.; Zhdanova, A.A.; Pyatygina, K.M.; Sokolov, D.I.; Sel’Kov, S.A. Mass-Spectrometric Analysis of Proteome of Microvesicles Produced by NK-92 Natural Killer Cells. Bull. Exp. Biol. Med. 2018, 165, 564–571. [Google Scholar] [CrossRef]
- Markova, K.L.; Kozyreva, A.R.; Gorshkova, A.A.; Aleksandrova, E.P.; Berezkina, M.E.; Mikhailova, V.A.; Ivanova, A.N.; Kaputkina, S.Y.; Onokhin, K.V.; Benken, K.A.; et al. Methodological Approaches to Assessing the Size and Morphology of Microvesicles of Cell Lines. Bull. Exp. Biol. Med. 2020, 169, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Evans-Osses, I.; Reichembach, L.H.; Ramirez, M.I. Exosomes or microvesicles? Two kinds of extracellular vesicles with different routes to modify protozoan-host cell interaction. Parasitol. Res. 2015, 114, 3567–3575. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Markov, A.S.; Markova, K.L.; Sokolov, D.I.; Selkov, S.A. Registration Certificate No. 2019612366 for Computer Program “MarkMigration”. Available online: https://www.fips.ru/registers-doc-view/fips_servlet?DB=EVM&DocNumber=2019612366&TypeFile=html (accessed on 18 February 2021).
- Gojova, A.; Barakat, A.I. Vascular endothelial wound closure under shear stress: Role of membrane fluidity and flow-sensitive ion channels. J. Appl. Physiol. 2005, 98, 2355–2362. [Google Scholar] [CrossRef] [Green Version]
- Si, Y.; Chu, H.; Zhu, W.; Xiao, T.; Shen, X.; Fu, Y.; Xu, R.; Jiang, H. Concentration-dependent effects of rapamycin on proliferation, migration and apoptosis of endothelial cells in human venous malformation. Exp. Ther. Med. 2018, 16, 4595–4601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waters, W.R.; Harkins, K.R.; Wannemuehler, M.J. Five-color flow cytometric analysis of swine lymphocytes for detection of proliferation, apoptosis, viability, and phenotype. Cytometry 2002, 48, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Philpott, N.J.; Scopes, J.; Marsh, J.C.; Gordon-Smith, E.C.; Gibson, F.M. Increased apoptosis in aplastic anemia bone marrow progenitor cells: Possible pathophysiologic significance. Exp. Hematol. 1995, 23, 1642–1648. [Google Scholar] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Markova, K.; Mikhailova, V.; Milyutina, Y.; Korenevsky, A.; Sirotskaya, A.; Rodygina, V.; Tyshchuk, E.; Grebenkina, P.; Simbirtsev, A.; Selkov, S.; et al. Effects of Microvesicles Derived from NK Cells Stimulated with IL-1β on the Phenotype and Functional Activity of Endothelial Cells. Int. J. Mol. Sci. 2021, 22, 13663. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413663
Markova K, Mikhailova V, Milyutina Y, Korenevsky A, Sirotskaya A, Rodygina V, Tyshchuk E, Grebenkina P, Simbirtsev A, Selkov S, et al. Effects of Microvesicles Derived from NK Cells Stimulated with IL-1β on the Phenotype and Functional Activity of Endothelial Cells. International Journal of Molecular Sciences. 2021; 22(24):13663. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413663
Chicago/Turabian StyleMarkova, Kseniia, Valentina Mikhailova, Yulia Milyutina, Andrey Korenevsky, Anastasia Sirotskaya, Veronika Rodygina, Elizaveta Tyshchuk, Polina Grebenkina, Andrey Simbirtsev, Sergey Selkov, and et al. 2021. "Effects of Microvesicles Derived from NK Cells Stimulated with IL-1β on the Phenotype and Functional Activity of Endothelial Cells" International Journal of Molecular Sciences 22, no. 24: 13663. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413663