
Extracellular Vesicles Mediate Communication between Endothelial and Vascular Smooth Muscle Cells

 , and
, and
Abstract
:1. Introduction
2. Results
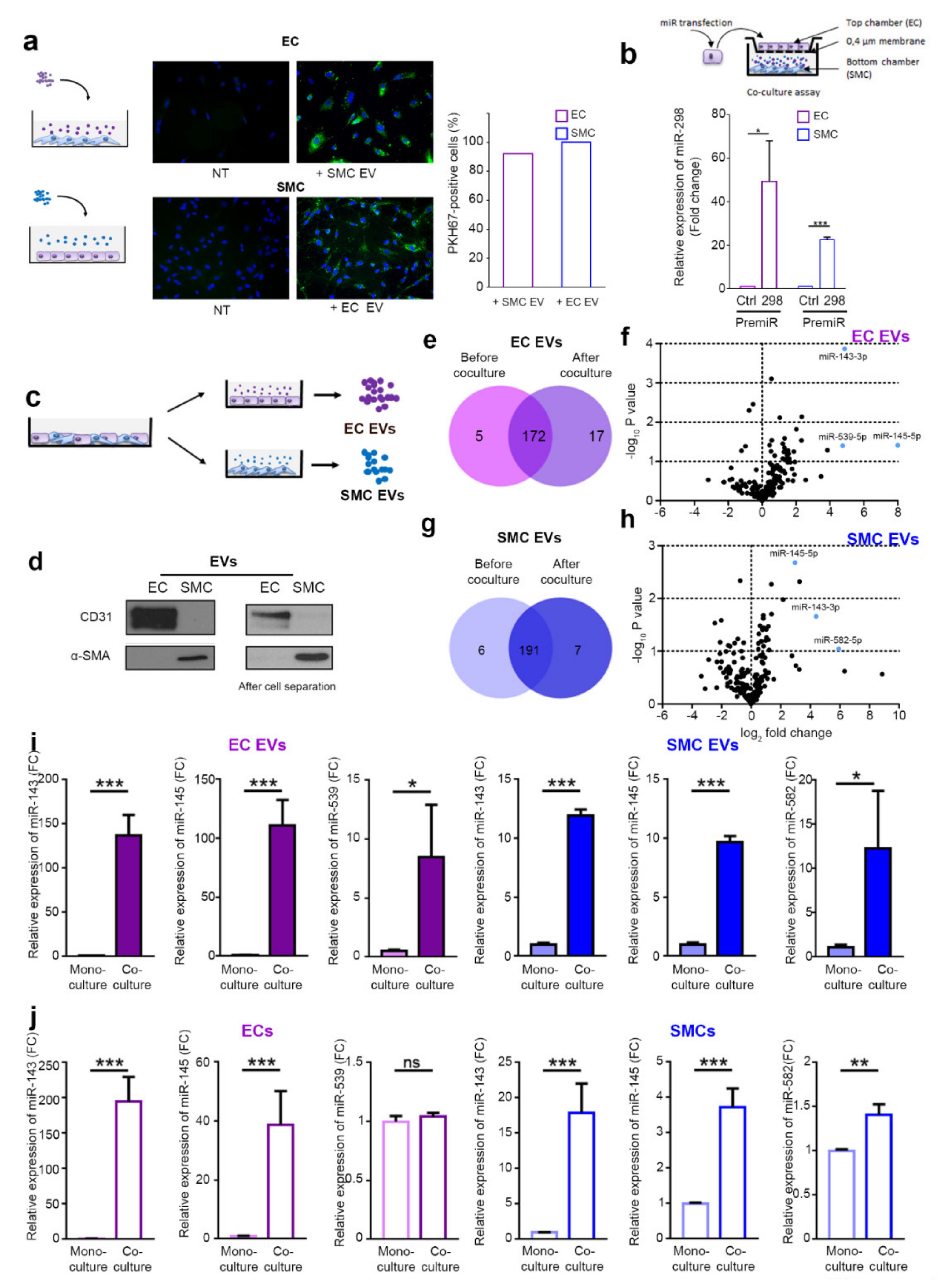
2.1. ECs and SMCs Are Able to Communicate via EVs
2.2. The microRNAs Cargo of EVs Is Modified by the Coculture
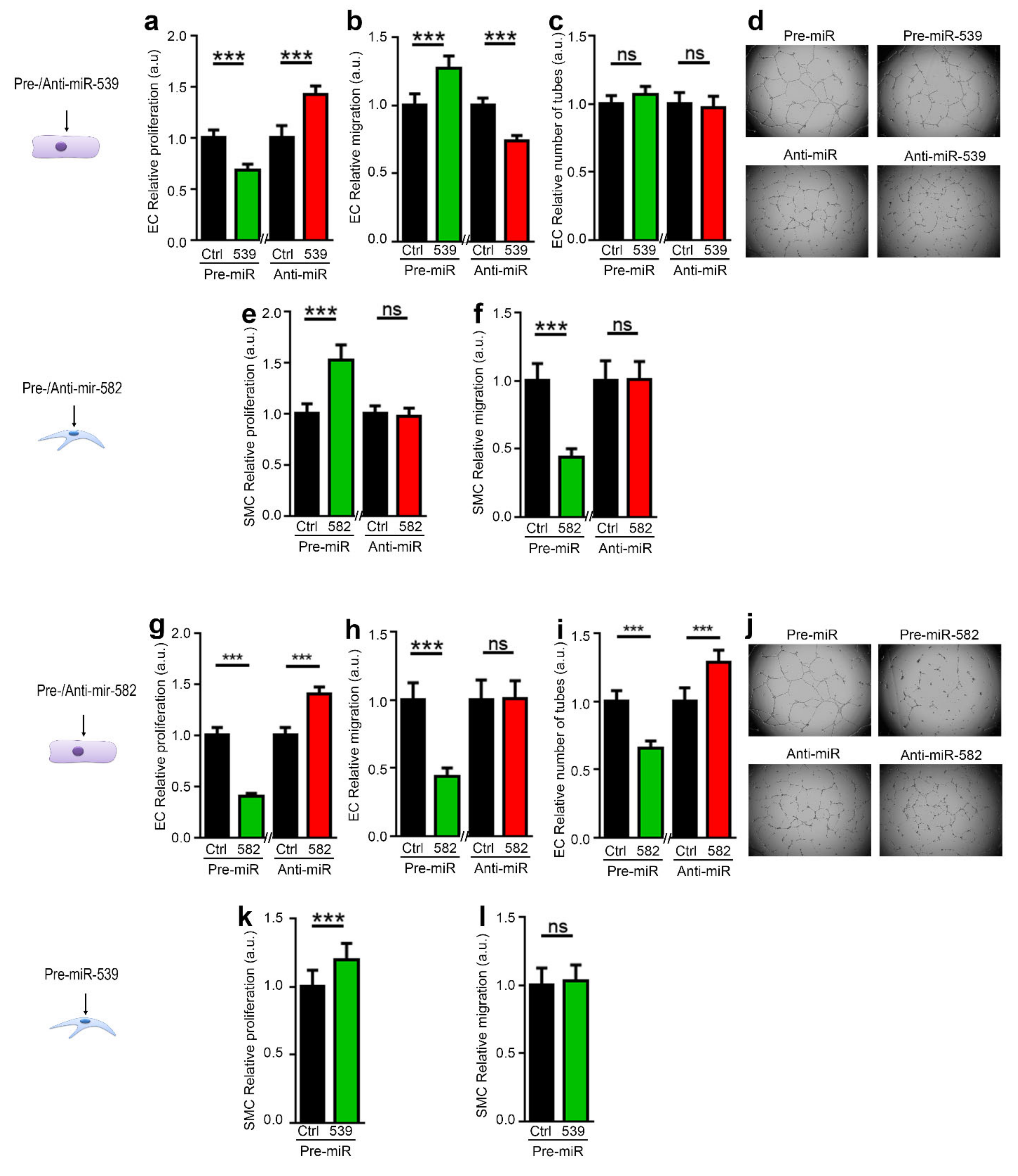
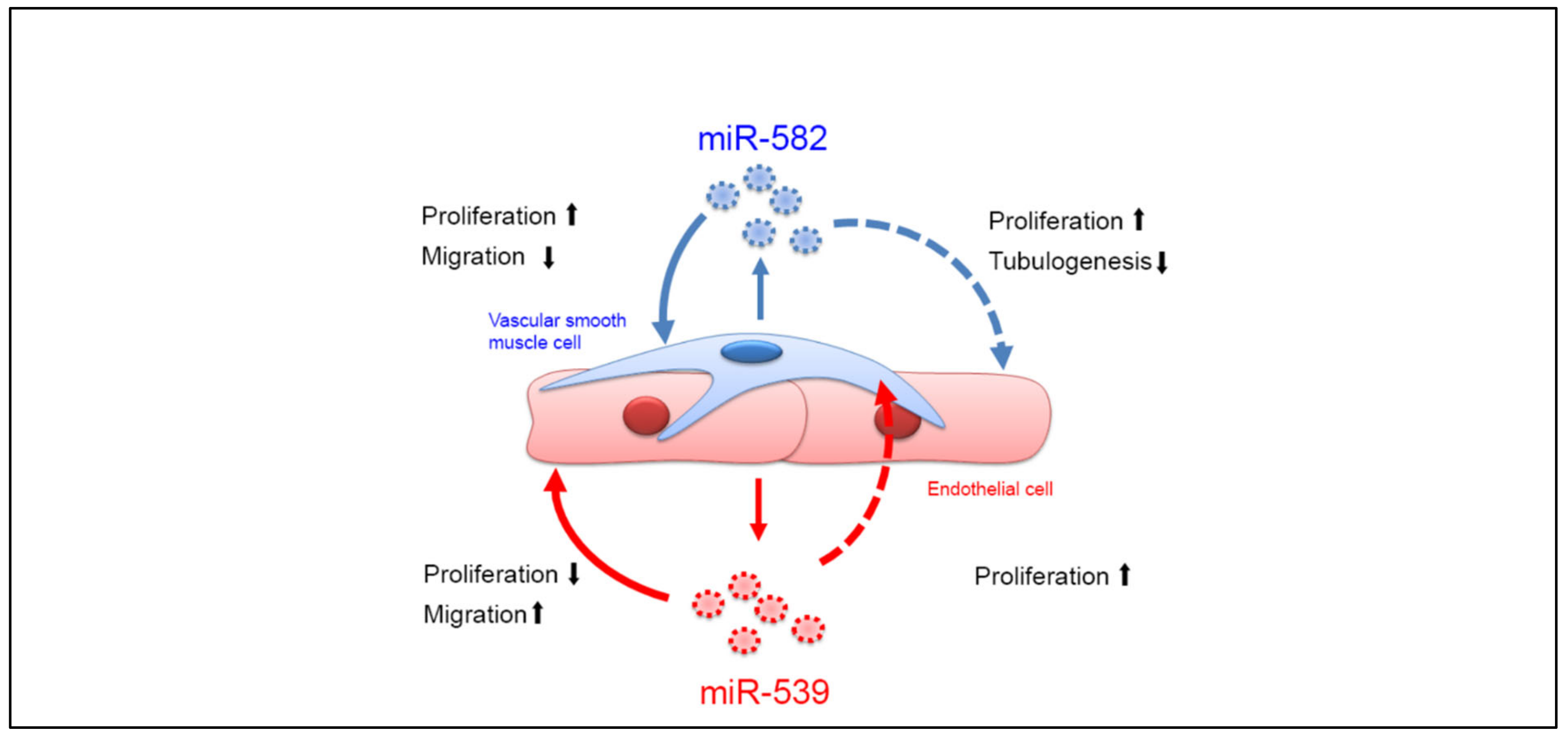
2.3. miR-539 and miR-582 Modulate ECs and SMCs Functions In Vitro
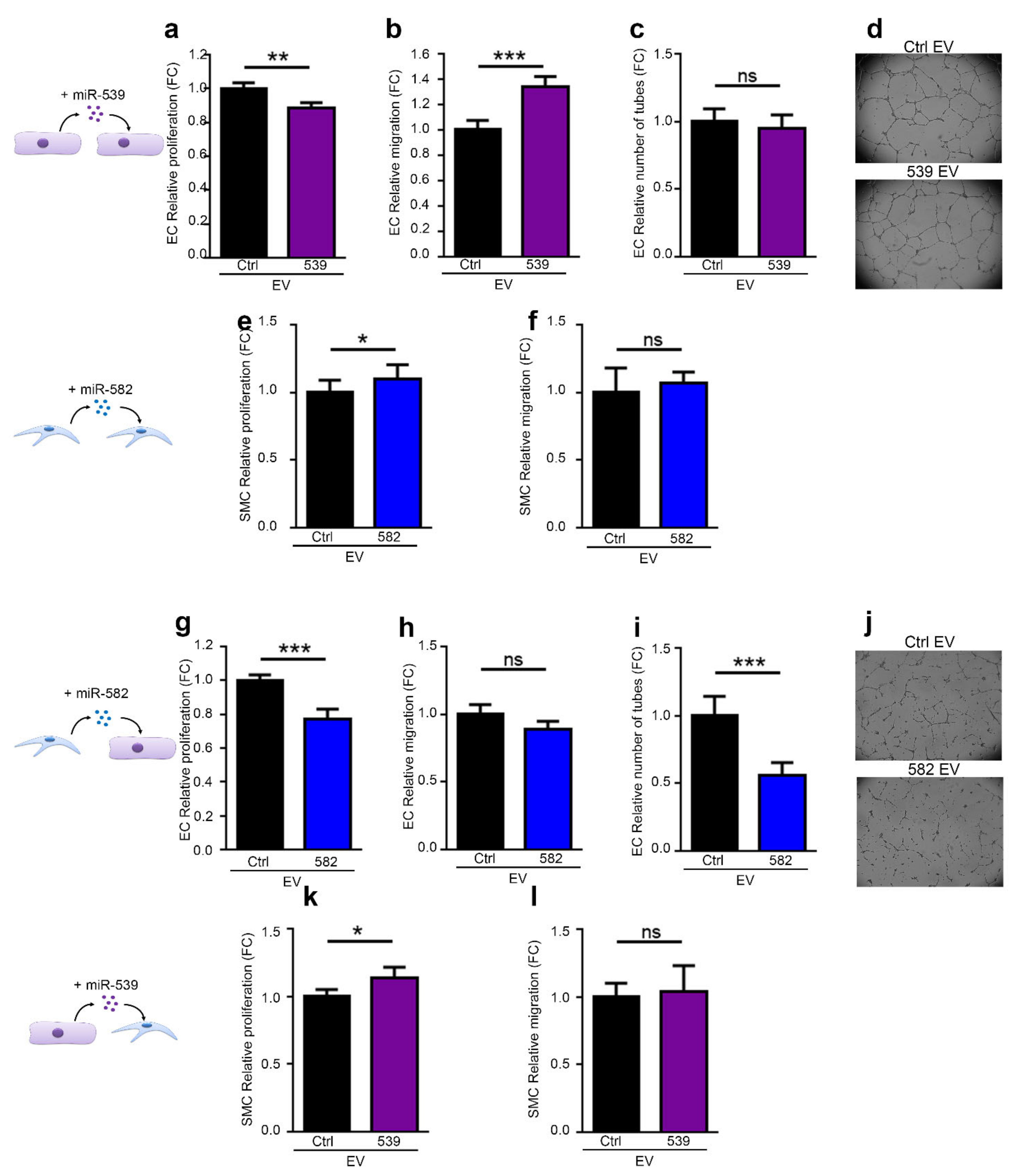
2.4. miR-539 and miR-582 Can Be Transferred via EVs and Modulate ECs Functions In Vitro
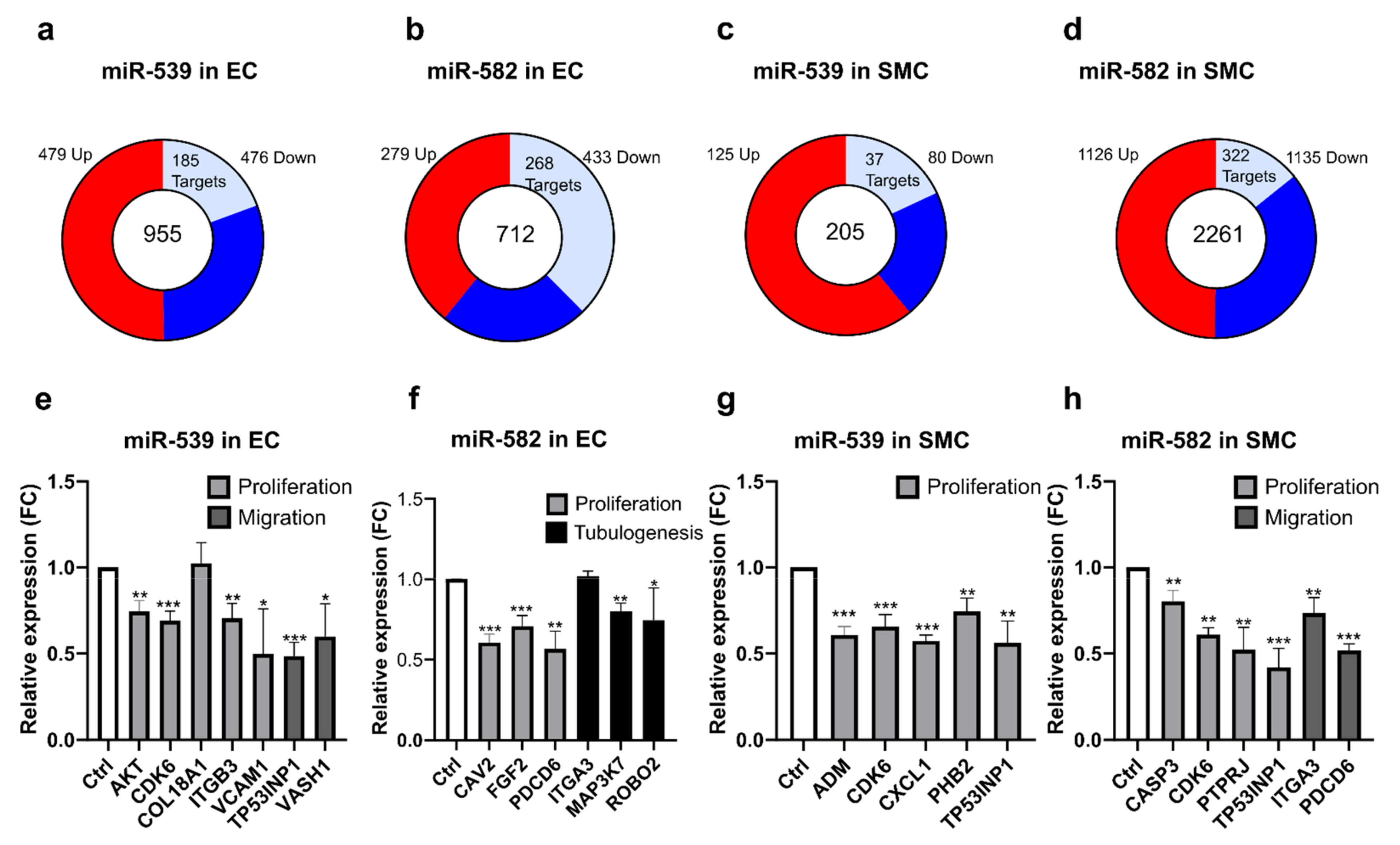
2.5. miR-539 and miR-582 Target Many Genes in ECs and SMCs
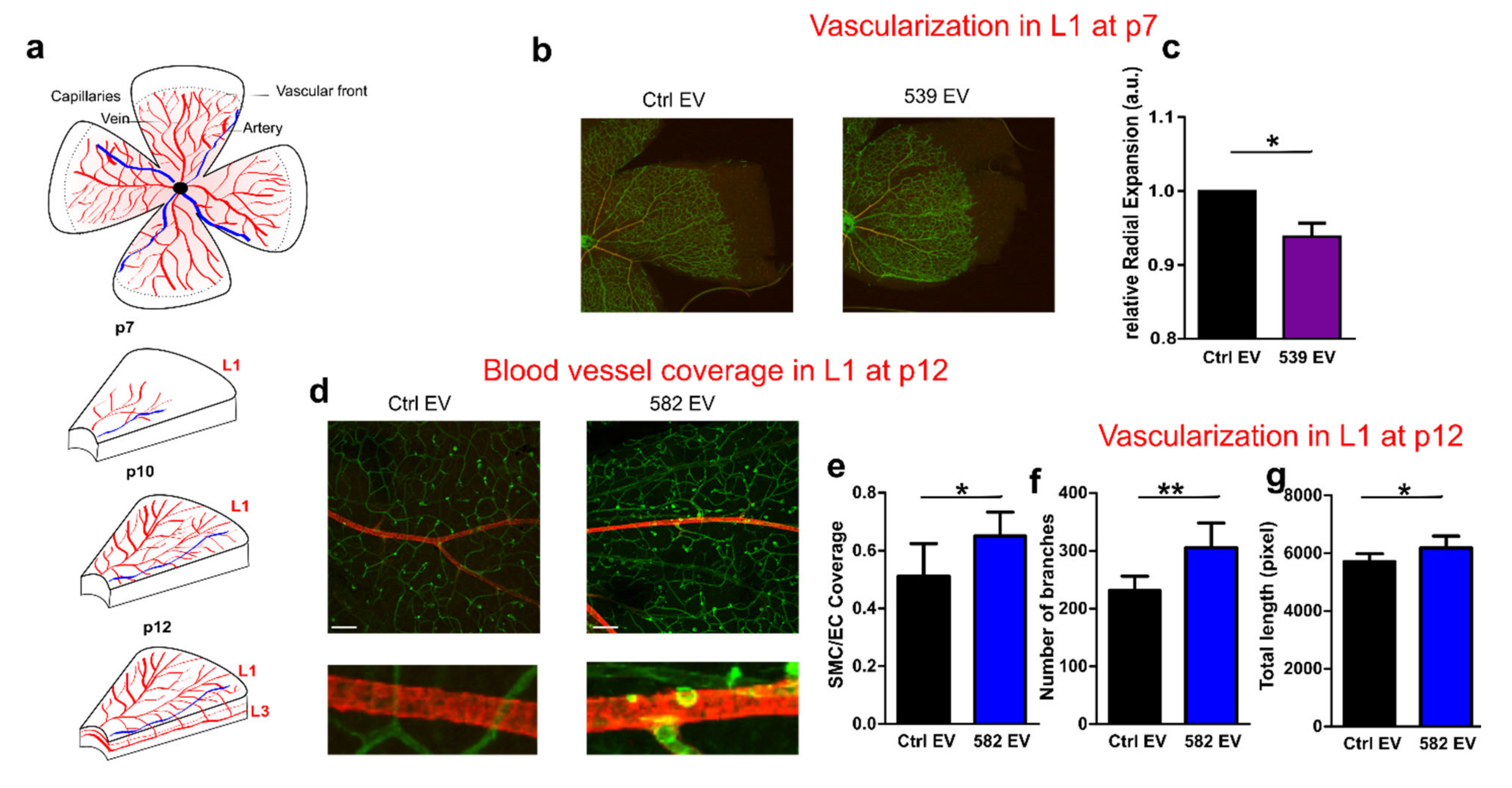
2.6. miR-539 and miR-582 Participate in Blood Vessel Maturation
3. Discussion
4. Material and Methods
4.1. Cell Culture
4.2. EVs Isolation
4.3. PHK67 Labelling of EVs
4.4. Western Blot Analysis
4.5. Dynamic Light Scattering
4.6. Electron Microscopy of Whole-Mounted Immuno-Labelled EVs
4.7. Flow Cytometry
4.8. Cells Transfections and Treatments
4.9. Transwell Coculture Assay
4.10. RNAse Treatment of EVs
4.11. Coculture and microRNA Profiling of EVs
4.12. miRNA Expression Analysis
4.13. Proliferation Assays
4.14. Migration Assay for SMCs in Boyden Chambers
4.15. Migration Healing Assay of ECs
4.16. Tubulogenesis Assay
4.17. Next-Generation Sequencing
4.18. mRNA Expression Analysis
4.19. Sequences of qPCR Primers

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Forward Primer | Reverse Primer |
|---|---|---|
| ADM | ATGAAGCTGGTTTCCGTCG | GACATCCGCAGTTCCCTCTT |
| AKT1 | AGCGACGTGGCTATTGTGAAG | GCCATCATTCTTGAGGAGGAAGT |
| CASP3 | CATGGAAGCGAATCAATGGACT | CTGTACCAGACCGAGATGTCA |
| CAV2 | AAGACCTGCCTAATGGTTCTGC | CTCGTACACAATGGAGCAATGAT |
| CDK6 | TGCCCGCATCTATAGTTTCCA | AGCCAACACTCCAGAGATCCAC |
| CXCL1 | CTTGCCTCAATCCTGCATC | CCTTCTGGTCAGTTGGATTTG |
| FGF2 | CACATTTAGAAGCCAGTAATCT | CCCGACGGCCGAG TTGAC |
| ITGA3 | TGTGGCTTGGAGTGACTGTG | TCATTGCCTCGCACGTAGC |
| ITGB3 | AGTAACCTGCGGATTGGCTTC | GTCACCTGGTCAGTTAGCGT |
| MAP3K7 | ATTGTAGAGCTTCGGCAGTTATC | CTGTAAACACCAACTCATTGCG |
| PDCD6 | ATGGCCGCCTACTCTTACC | TCCTGTCTTTATCGACCCTCTG |
| PHB2 | GTGCGCGAATCTGTGTTCAC | GATAATGGGGTACTGGAACCAAG |
| PTPRJ | GGCACCCCTAGTCCAATTCC | TCCCATTAGATCCTTGTTCAGGT |
| ROBO2 | GTTTGTGTTGCGAGGAACTATCT | GTTTTGTCGGAAGTCATCTCGTA |
| TP53INP1 | TTCCTCCAACCAAGAACCAGA | GCTCAGTAGGTGACTCTTCACT |
| VASH1 | GGTGGGCTACCTGTGGATG | CACTCGGTATGGGGATCTTGG |
| VCAM-1 | GAGACCACCCCAGAATCTAGATA | ACTTCCTGTCTGCATCCTCCA |
4.20. Retinal Murine Neovascularization Model
4.21. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Potente, M.; Gerhardt, H.; Carmeliet, P. Basic and therapeutic aspects of angiogenesis. Cell 2011, 146, 873–887. [Google Scholar] [CrossRef] [Green Version]
- Lilly, B. We have contact: Endothelial cell-smooth muscle cell interactions. Physiology 2014, 29, 234–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaengel, K.; Genové, G.; Armulik, A.; Betsholtz, C. Endothelial-mural cell signaling in vascular development and angiogenesis. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 630–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweeney, M.; Foldes, G. It Takes Two: Endothelial-Perivascular Cell Cross-Talk in Vascular Development and Disease. Front. Cardiovasc. Med. 2018, 5, 154. [Google Scholar] [CrossRef]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Théry, C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, Secretion, and Intercellular Interactions of Exosomes and Other Extracellular Vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2019, 8, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosaka, N.; Kogure, A.; Yamamoto, T.; Urabe, F.; Usuba, W.; Prieto-Vila, M.; Ochiya, T. Exploiting the message from cancer: The diagnostic value of extracellular vesicles for clinical applications. Exp. Mol. Med. 2019, 51, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tkach, M.; Thery, C. Communication by Extracellular Vesicles: Where We Are and Where We Need to Go. Cell 2016, 164, 1226–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, N.; Whiteside, T.L. Potential roles of tumor-derived exosomes in angiogenesis. Expert Opin. Ther. Targets 2018, 22, 409–417. [Google Scholar] [CrossRef]
- Maia, J.; Caja, S.; Moraes, M.C.S.; Couto, N.; Costa-Silva, B. Exosome-Based Cell-Cell Communication in the Tumor Microenvironment. Front. Cell Dev. Biol. 2018, 6, 18. [Google Scholar] [CrossRef]
- Bovy, N.; Blomme, B.; Frères, P.; Dederen, S.; Nivelles, O.; Lion, M.; Carnet, O.; Martial, J.A.; Noël, A.; Thiry, M.; et al. Endothelial exosomes contribute to the antitumor response during breast cancer neoadjuvant chemotherapy via microRNA transfer. Oncotarget 2015, 6, 10253–10266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Zhang, L.; Wang, S.; Han, Q.; Zhao, R.C. Exosomes secreted by mesenchymal stem cells promote endothelial cell angiogenesis by transferring miR-125a. J. Cell Sci. 2016, 129, 2182–2189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Balkom, B.W.M.; De Jong, O.G.; Smits, M.; Brummelman, J.; Den Ouden, K.; De Bree, P.M.; Van Eijndhoven, M.A.J.; Pegtel, D.M.; Stoorvogel, W.; Urdinger, T.; et al. Endothelial cells require miR-214 to secrete exosomes that suppress senescence and induce angiogenesis in human and mouse endothelial cells. Blood 2017, 121, 3997–4007. [Google Scholar] [CrossRef] [Green Version]
- Kapustin, A.N.; Shanahan, C.M. Emerging roles for vascular smooth muscle cell exosomes in calcification and coagulation. J. Physiol. 2016, 594, 2905–2914. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Blanco, F.J.; Stevens, H.; Lu, R.; Caudrillier, A.; McBride, M.; McClure, J.D.; Grant, J.; Thomas, M.; Frid, M.; et al. MicroRNA-143 Activation Regulates Smooth Muscle and Endothelial Cell Crosstalk in Pulmonary Arterial Hypertension. Circ. Res. 2015, 117, 870–883. [Google Scholar] [CrossRef] [PubMed]
- Climent, M.; Quintavalle, M.; Miragoli, M.; Chen, J.; Condorelli, G.; Elia, L. TGFβ triggers miR-143/145 transfer from smooth muscle cells to endothelial cells, thereby modulating vessel stabilization. Circ. Res. 2015, 116, 1753–1764. [Google Scholar] [CrossRef] [Green Version]
- Hergenreider, E.; Heydt, S.; Tréguer, K.; Boettger, T.; Horrevoets, A.J.G.; Zeiher, A.M.; Scheffer, M.P.; Frangakis, A.S.; Yin, X.; Mayr, M.; et al. Atheroprotective communication between endothelial cells and smooth muscle cells through miRNAs. Nat. Cell Biol. 2012, 14, 249–256. [Google Scholar] [CrossRef]
- Lin, X.; He, Y.; Hou, X.; Zhang, Z.; Wang, R.; Wu, Q. Endothelial Cells Can Regulate Smooth Muscle Cells in Contractile Phenotype through the miR-206/ARF6&NCX1/Exosome Axis. PLoS ONE 2016, 11, e0152959. [Google Scholar] [CrossRef]
- Zheng, B.; Yin, W.; Suzuki, T.; Zhang, X.; Zhang, Y.; Song, L.; Jin, L.; Zhan, H.; Zhang, H.; Li, J.; et al. Exosome-Mediated miR-155 Transfer from Smooth Muscle Cells to Endothelial Cells Induces Endothelial Injury and Promotes Atherosclerosis. Mol. Ther. 2017, 25, 1279–1294. [Google Scholar] [CrossRef] [Green Version]
- Kowal, J.; Arras, G.; Colombo, M.; Jouve, M.; Morath, J.P.; Primdal-Bengtson, B.; Dingli, F.; Loew, D.; Tkach, M.; Théry, C. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc. Natl. Acad. Sci. USA 2016, 113, E968–E977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rust, R.; Grönnert, L.; Dogançay, B.; Schwab, M.E. A Revised View on Growth and Remodeling in the Retinal Vasculature. Sci. Rep. 2019, 9, 3263. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.P.; Kim, E.Y.; Badr, C.E.; Weissleder, R.; Mempel, T.R.; Tannous, B.A.; Breakefield, X.O. Visualization and tracking of tumour extracellular vesicle delivery and RNA translation using multiplexed reporters. Nat. Commun. 2015, 6, 7029. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nabet, B.Y.; Qiu, Y.; Shabason, J.E.; Wu, T.J.; Yoon, T.; Kim, B.C.; Benci, J.L.; DeMichele, A.M.; Tchou, J.; Marcotrigiano, J.; et al. Exosome RNA Unshielding Couples Stromal Activation to Pattern Recognition Receptor Signaling in Cancer. Cell 2017, 170, 352–366.e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, H.; Wang, W. MicroRNA-539 suppresses osteosarcoma cell invasion and migration in vitro and targeting Matrix metallopeptidase-8. Int. J. Clin. Exp. Pathol. 2015, 8, 8075–8082. [Google Scholar]
- Uchino, K.; Takeshita, F.; Takahashi, R.; Kosaka, N.; Fujiwara, K.; Naruoka, H.; Sonoke, S.; Yano, J.; Sasaki, H.; Nozawa, S.; et al. Therapeutic Effects of MicroRNA-582-5p and -3p on the Inhibition of Bladder Cancer Progression. Mol. Ther. 2013, 21, 610–619. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Zhang, Y.; Wu, X. 786-0 renal cancer cell line-derived exosomes promote 786-0 cell migration and invasion in vitro. Oncol. Lett. 2014, 7, 1576–1580. [Google Scholar] [CrossRef] [Green Version]
- Willms, E.; Johansson, H.J.; Mäger, I.; Lee, Y.; Blomberg, K.E.M.; Sadik, M.; Alaarg, A.; Smith, C.I.E.; Lehtiö, J.; El Andaloussi, S.; et al. Cells release subpopulations of exosomes with distinct molecular and biological properties. Sci. Rep. 2016, 6, 22519. [Google Scholar] [CrossRef]
- Bordanaba-Florit, G.; Royo, F.; Kruglik, S.G.; Falcón-Pérez, J.M. Using single-vesicle technologies to unravel the heterogeneity of extracellular vesicles. Nat. Protoc. 2021, 16, 3163–3185. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Moita, C.; van Niel, G.; Kowal, J.; Vigneron, J.; Benaroch, P.; Manel, N.; Moita, L.F.; Théry, C.; Raposo, G. Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles. J. Cell Sci. 2013, 126, 5553–5565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Deun, J.; Mestdagh, P.; Sormunen, R.; Cocquyt, V.; Vermaelen, K.; Vandesompele, J.; Bracke, M.; De Wever, O.; Hendrix, A. The impact of disparate isolation methods for extracellular vesicles on downstream RNA profiling. J. Extracell. Vesicles 2014, 3, 24858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mateescu, B.; Kowal, E.J.K.; van Balkom, B.W.M.; Bartel, S.; Bhattacharyya, S.N.; Buzás, E.I.; Buck, A.H.; de Candia, P.; Chow, F.W.N.; Das, S.; et al. Obstacles and opportunities in the functional analysis of extracellular vesicle RNA–An ISEV position paper. J. Extracell. Vesicles 2017, 6, 1286095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seux, M.; Peuget, S.; Montero, M.; Siret, C.; Rigot, V.; Clerc, P.; Gigoux, V.; Pellegrino, E.; Pouyet, L.; Guessan, P.; et al. TP53INP1 decreases pancreatic cancer cell migration by regulating SPARC expression. Oncogene 2011, 30, 3049–3061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, K.; Hasegawa, Y.; Yamashita, H.; Shimizu, K.; Ding, Y.; Abe, M.; Ohta, H.; Imagawa, K.; Hojo, K.; Maki, H.; et al. Vasohibin as an endothelium-derived negative feedback regulator of angiogenesis. J. Clin. Investig. 2004, 114, 898–907. [Google Scholar] [CrossRef] [Green Version]
- Somanath, P.R.; Razorenova, O.V.; Chen, J.; Byzova, T.V. Akt1 in endothelial cell and angiogenesis. Cell Cycle 2006, 5, 512–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conway, E.M.; Collen, D.; Carmeliet, P. Molecular mechanisms of blood vessel growth. Cardiovasc. Res. 2001, 49, 507–521. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; Vo-Ransdell, C.; Abel, B.; Willoughby, C.; Jang, S.; Sowa, G. Caveolin-2 is a negative regulator of anti-proliferative function and signaling of transforming growth factor beta in endothelial cells. Am. J. Physiol. Cell Physiol. 2011, 301, C1161–C1174. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zuo, D.; Yuan, Y.; Yang, X.; Hong, Z.; Zhang, R. MicroRNA-183 promotes cell proliferation via regulating programmed cell death 6 in pediatric acute myeloid leukemia. J. Cancer Res. Clin. Oncol. 2016, 143, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Zippel, N.; Malik, R.A.; Frömel, T.; Popp, R.; Bess, E.; Strilic, B.; Wettschureck, N.; Fleming, I.; Fisslthaler, B. Transforming growth factor-β-activated kinase 1 regulates angiogenesis via AMP-activated protein kinase-α1 and redox balance in endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 2792–2799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morioka, S.; Inagaki, M.; Komatsu, Y.; Mishina, Y.; Matsumoto, K.; Ninomiya-Tsuji, J. TAK1 kinase signaling regulates embryonic angiogenesis by modulating endothelial cell survival and migration. Blood 2012, 120, 3846–3857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rama, N.; Dubrac, A.; Mathivet, T.; Ní Chárthaigh, R.-A.; Genet, G.; Cristofaro, B.; Pibouin-Fragner, L.; Ma, L.; Eichmann, A.; Chédotal, A. Slit2 signaling through Robo1 and Robo2 is required for retinal neovascularization. Nat. Med. 2015, 21, 483–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cartland, S.; Genner, S.; Zahoor, A.; Kavurma, M. Comparative Evaluation of TRAIL, FGF-2 and VEGF-A-Induced Angiogenesis In Vitro and In Vivo. Int. J. Mol. Sci. 2016, 17, 2025. [Google Scholar] [CrossRef] [Green Version]
- Otto, T.; Sicinski, P. The kinase-independent, second life of CDK6 in transcription. Cancer Cell 2013, 24, 141–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Feng, Y.; Li, Z.; Tang, C. The effect of adrenomedullin and proadrenomedullin N-terminal 20 peptide on angiotensin II induced vascular smooth muscle cell proliferation. Iran. J. Basic Med. Sci. 2016, 19, 49–54. [Google Scholar]
- Tang, R.; Zhang, G.; Chen, S.-Y. Smooth Muscle Cell Proangiogenic Phenotype Induced by Cyclopentenyl Cytosine Promotes Endothelial Cell Proliferation and Migration. J. Biol. Chem. 2016, 291, 26913–26921. [Google Scholar] [CrossRef] [Green Version]
- Merkwirth, C.; Dargazanli, S.; Tatsuta, T.; Geimer, S.; Löwer, B.; Wunderlich, F.T.; Von Kleist-Retzow, J.C.; Waisman, A.; Westermann, B.; Langer, T. Prohibitins control cell proliferation and apoptosis by regulating OPA1-dependent cristae morphogenesis in mitochondria. Genes Dev. 2008, 22, 476–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Long, B.; Zhou, L.-Y.; Liu, F.; Zhou, Q.-Y.; Liu, C.-Y.; Fan, Y.-Y.; Li, P.-F. CARL lncRNA inhibits anoxia-induced mitochondrial fission and apoptosis in cardiomyocytes by impairing miR-539-dependent PHB2 downregulation. Nat. Commun. 2014, 5, 3596. [Google Scholar] [CrossRef] [Green Version]
- Woo, M.; Hakem, R.; Furlonger, C.; Hakem, A.; Duncan, G.S.; Sasaki, T.; Bouchard, D.; Lu, L.; Wu, G.E.; Paige, C.J.; et al. Caspase-3 regulates cell cycle in B cells: A consequence of substrate specificity. Nat. Immunol. 2003, 4, 1016–1022. [Google Scholar] [CrossRef]
- Smart, C.E.; Askarian Amiri, M.E.; Wronski, A.; Dinger, M.E.; Crawford, J.; Ovchinnikov, D.A.; Vargas, A.C.; Reid, L.; Simpson, P.T.; Song, S.; et al. Expression and function of the protein tyrosine phosphatase receptor J (PTPRJ) in normal mammary epithelial cells and breast tumors. PLoS ONE 2012, 7, e40742. [Google Scholar] [CrossRef] [Green Version]
- Kurozumi, A.; Goto, Y.; Matsushita, R.; Fukumoto, I.; Kato, M.; Nishikawa, R.; Sakamoto, S.; Enokida, H.; Nakagawa, M.; Ichikawa, T.; et al. Tumor-suppressive microRNA-223 inhibits cancer cell migration and invasion by targeting ITGA3/ITGB1 signaling in prostate cancer. Cancer Sci. 2016, 107, 84–94. [Google Scholar] [CrossRef] [Green Version]
- Su, D.; Xu, H.; Feng, J.; Gao, Y.; Gu, L.; Ying, L.; Katsaros, D.; Yu, H.; Xu, S.; Qi, M. PDCD6 is an independent predictor of progression free survival in epithelial ovarian cancer. J. Transl. Med. 2012, 10, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, F.; Yang, J.; Xu, Y.; Guan, S. MiR-539 Targets MMP-9 to Regulate the Permeability of Blood–Brain Barrier in Ischemia/Reperfusion Injury of Brain. Neurochem. Res. 2018, 43, 2260–2267. [Google Scholar] [CrossRef]
- Feng, Y.; Wang, J.; Yuan, Y.; Zhang, X.; Shen, M.; Yuan, F. MiR-539-5p inhibits experimental choroidal neovascularization by targeting CXCR7. FASEB J. 2018, 32, 1626–1639. [Google Scholar] [CrossRef] [Green Version]
- Quinn, J.F.; Patel, T.; Wong, D.; Das, S.; Freedman, J.E.; Laurent, L.C.; Carter, B.S.; Hochberg, F.; Van Keuren-jensen, K.; Huentelman, M.; et al. Extracellular RNAs: Development as biomarkers of human disease. J. Extracell. Vesicles 2015, 4, 27495. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, E.A.; Nachman, R.L.; Becker, C.G.; Minick, C.R. Culture of human endothelial cells derived from umbilical veins. Identification by morphologic and immunologic criteria. J. Clin. Investig. 1973, 52, 2745–2756. [Google Scholar] [CrossRef] [PubMed]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Gerhardt, H.; Golding, M.; Fruttiger, M.; Ruhrberg, C.; Lundkvist, A.; Abramsson, A.; Jeltsch, M.; Mitchell, C.; Alitalo, K.; Shima, D.; et al. VEGF guides angiogenic sprouting utilizing endothelial tip cell filopodia. J. Cell Biol. 2003, 161, 1163–1177. [Google Scholar] [CrossRef]






| GO ID | Biological Processes | Matched Genes in the Network | Genes in the Networks | False Discovery Rate |
|---|---|---|---|---|
| miR-539 in ECs | ||||
| GO:0030336 | negative regulation of cell migration | 28 | 237 | 0.00093 |
| GO:0008284 | positive regulation of cell population proliferation | 69 | 878 | 0.002 |
| miR-582 in ECs | ||||
| GO:0035295 | tube development | 59 | 793 | 1.37 × 105 |
| GO:0008284 | positive regulation of cell population proliferation | 58 | 878 | 0.00035 |
| GO:0001568 | blood vessel development | 44 | 464 | 1.49 × 106 |
| GO:0001525 | angiogenesis | 30 | 297 | 4.72 × 105 |
| miR-539 in SMCs | ||||
| GO:0008285 | negative regulation of cell population proliferation | 18 | 669 | 0.0247 |
| GO:0030334 | regulation of cell migration | 20 | 753 | 0.0194 |
| GO:0045765 | regulation of angiogenesis | 14 | 277 | 0.0044 |
| miR-582 in SMCs | ||||
| GO:0008285 | negative regulation of cell population proliferation | 103 | 669 | 0.0016 |
| GO:0030335 | positive regulation of cell migration | 81 | 452 | 0.00015 |
| GO:1901342 | regulation of vasculature development | 58 | 305 | 0.00067 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fontaine, M.; Herkenne, S.; Ek, O.; Paquot, A.; Boeckx, A.; Paques, C.; Nivelles, O.; Thiry, M.; Struman, I. Extracellular Vesicles Mediate Communication between Endothelial and Vascular Smooth Muscle Cells. Int. J. Mol. Sci. 2022, 23, 331. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010331
Fontaine M, Herkenne S, Ek O, Paquot A, Boeckx A, Paques C, Nivelles O, Thiry M, Struman I. Extracellular Vesicles Mediate Communication between Endothelial and Vascular Smooth Muscle Cells. International Journal of Molecular Sciences. 2022; 23(1):331. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010331
Chicago/Turabian StyleFontaine, Marie, Stéphanie Herkenne, Olivier Ek, Alicia Paquot, Amandine Boeckx, Cécile Paques, Olivier Nivelles, Marc Thiry, and Ingrid Struman. 2022. "Extracellular Vesicles Mediate Communication between Endothelial and Vascular Smooth Muscle Cells" International Journal of Molecular Sciences 23, no. 1: 331. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010331