
Silencing of ATP Synthase β Impairs Egg Development in the Leafhopper Scaphoideus titanus, Vector of the Phytoplasma Associated with Grapevine Flavescence Dorée

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. RNAi Machinery of Scaphoideus titanus
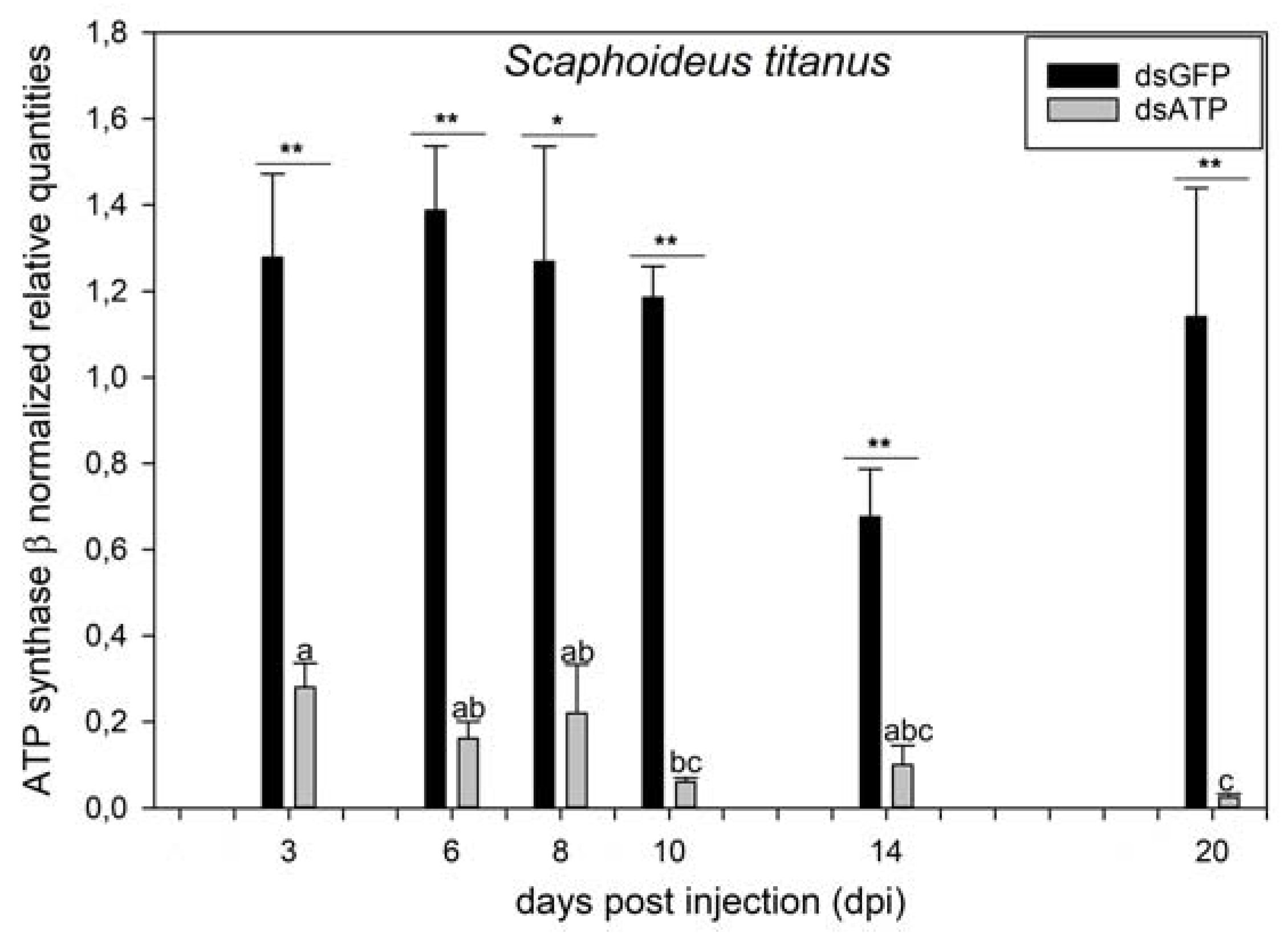
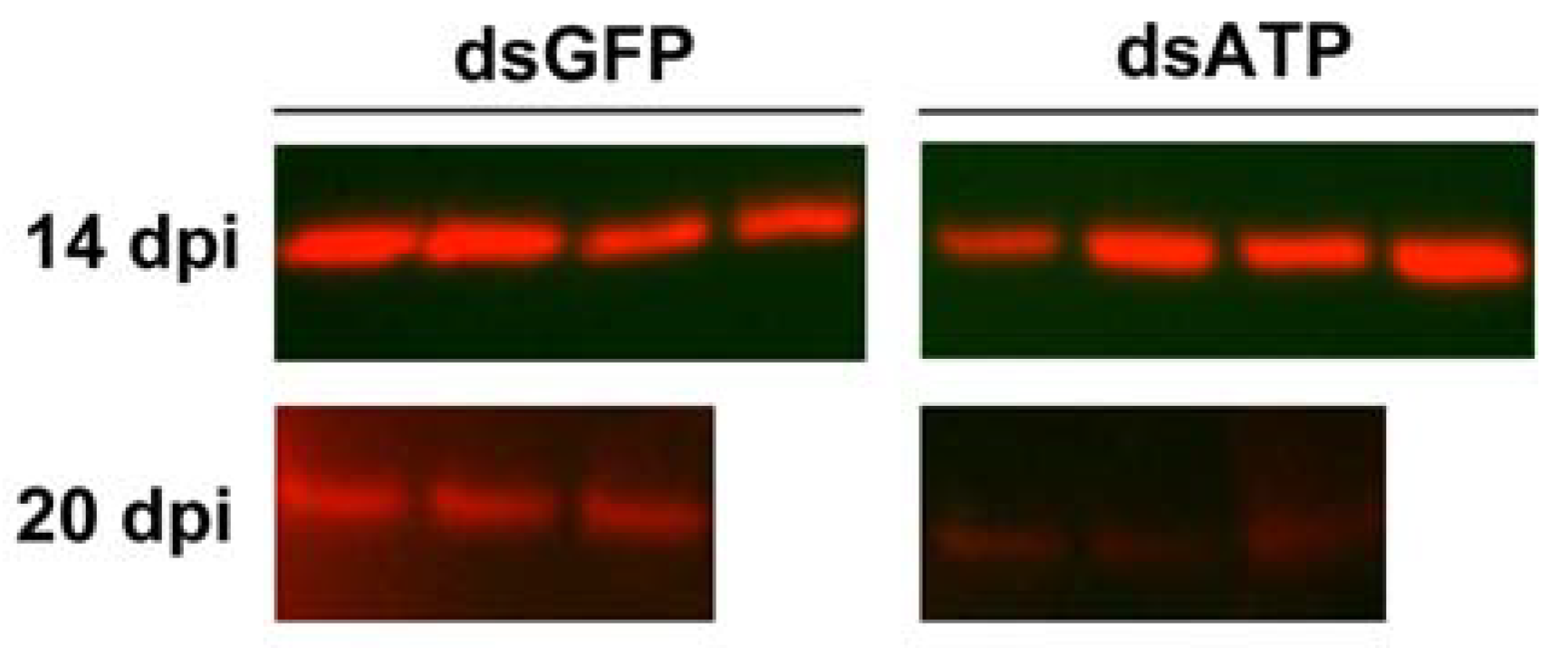
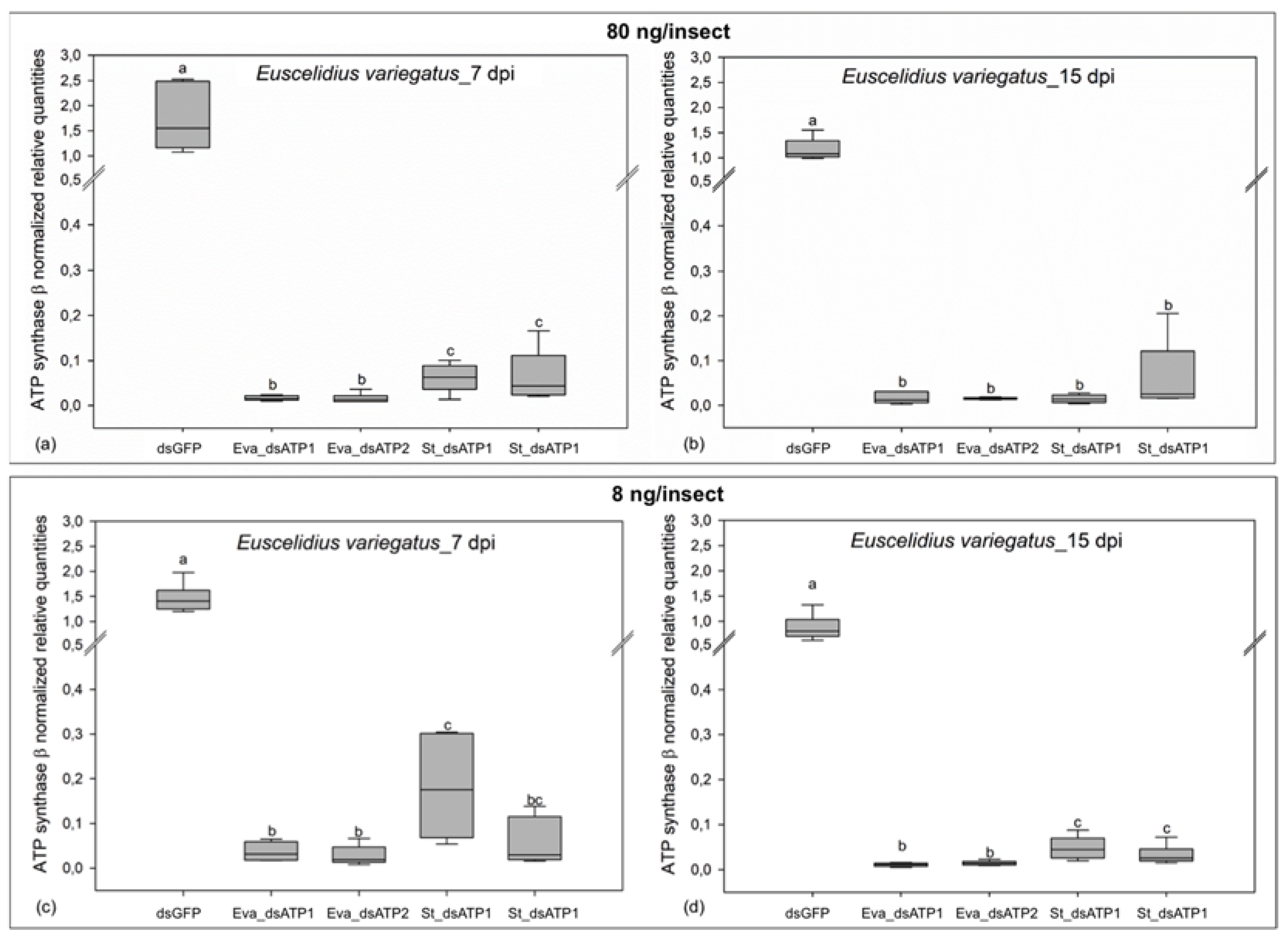
2.2. ATP Synthase β Is Silenced at Transcript and Protein Levels in Scaphoideus titanus
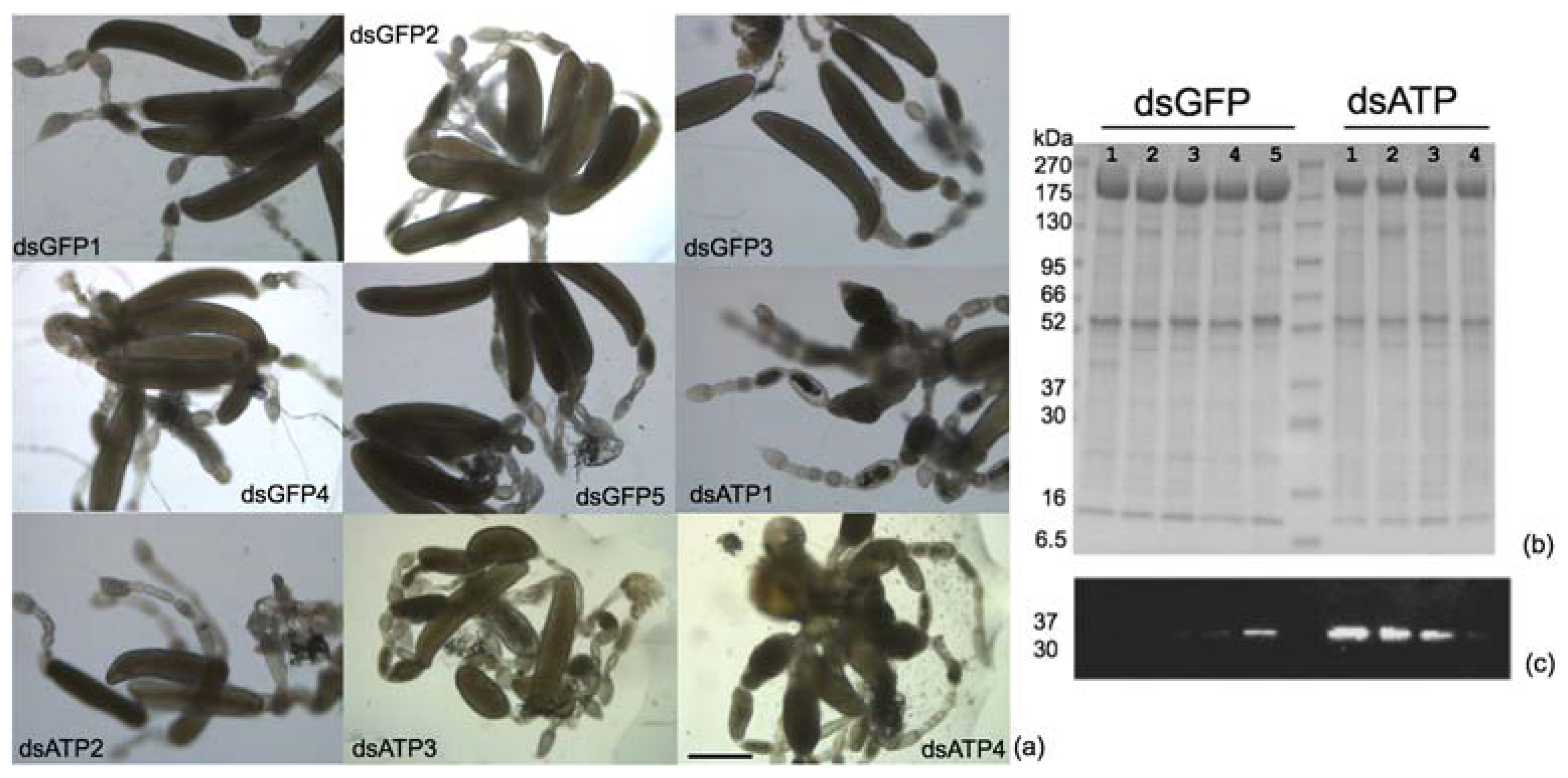
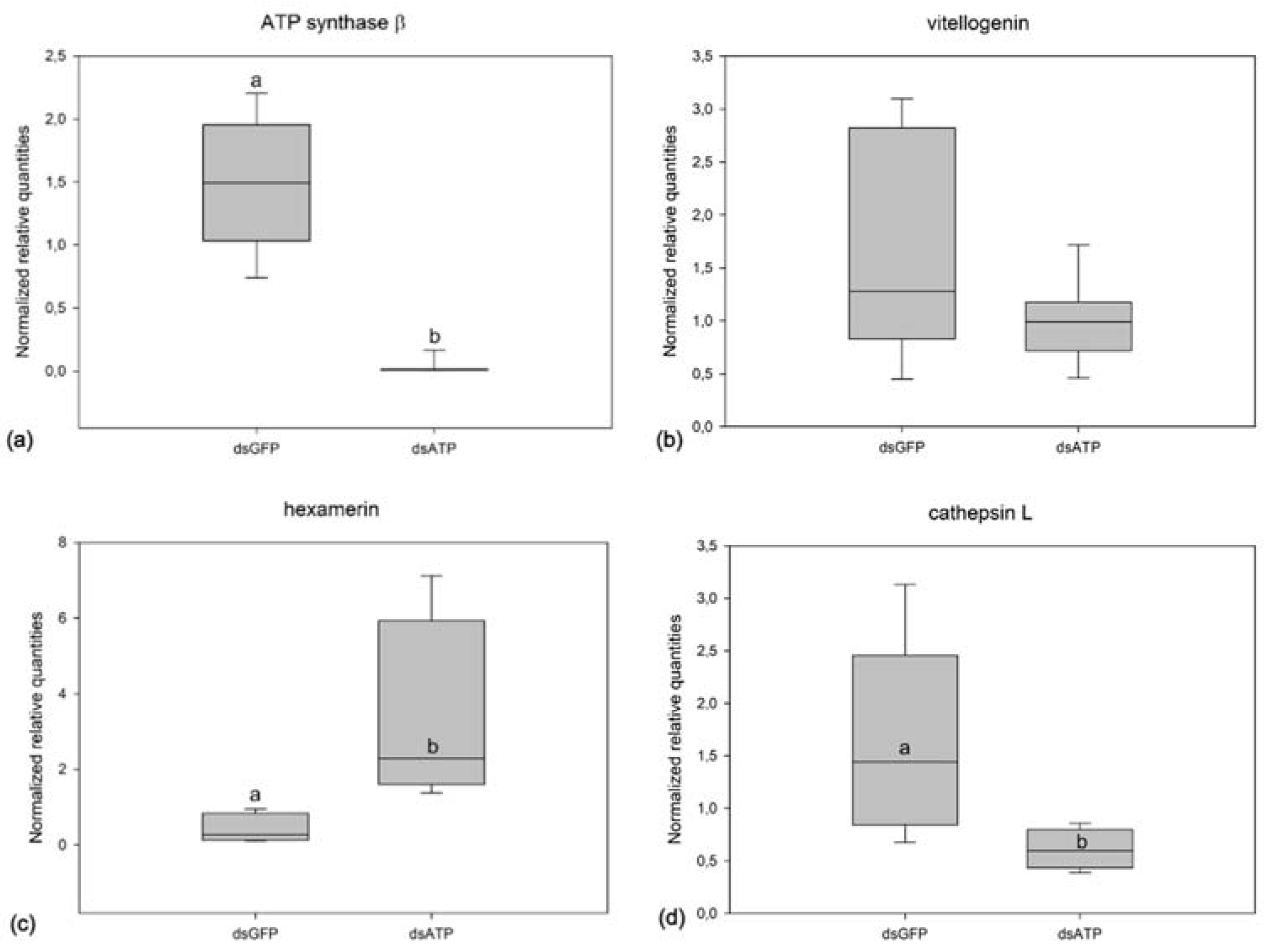
2.3. Effects of ATP Synthase β Silencing on Survival Rate and Egg Maturation in Scaphoideus titanus
2.4. ATP Synthase β Sequence Analysis
2.5. Specificity of dsRNAs on Non-Target Species
3. Discussion
4. Materials and Methods
4.1. Insect Rearing
4.2. Synthesis and Delivery of dsRNA
4.3. Ovary Observation and Progeny Collection
4.4. RNA Extraction, cDNA Synthesis, and Gene Expression Analysis
4.5. Protein Gels and Western Blots
4.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Bertin, S.; Guglielmino, C.R.; Karam, N.; Gomulski, L.M.; Malacrida, A.R.; Gasperi, G. Diffusion of the Nearctic Leafhopper Scaphoideus titanus Ball in Europe: A Consequence of Human Trading Activity. Genetica 2007, 131, 275–285. [Google Scholar] [CrossRef]
- Papura, D.; Burban, C.; van Helden, M.; Giresse, X.; Nusillard, B.; Guillemaud, T.; Kerdelhué, C. Microsatellite and Mitochondrial Data Provide Evidence for a Single Major Introduction for the Neartic Leafhopper Scaphoideus titanus in Europe. PLoS ONE 2012, 7, e36882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA Panel on Plant Health PLH. Scientific Opinion on Pest Categorisation of Grapevine Flavescence Dorée: Grapevine Flavescence Dorée Pest Categorisation. EFSA J. 2014, 12, 3851. [Google Scholar] [CrossRef]
- EFSA Panel on Plant Health PLH; Jeger, M.; Bragard, C.; Caffier, D.; Candresse, T.; Chatzivassiliou, E.; Dehnen-Schmutz, K.; Gilioli, G.; Jaques Miret, J.A.; MacLeod, A.; et al. Risk to Plant Health of Flavescence Dorée for the EU Territory. EFSA J. 2016, 14, 4603. [Google Scholar] [CrossRef]
- EFSA; Tramontini, S.; Delbianco, A.; Vos, S. Pest Survey Card on Flavescence Dorée Phytoplasma and Its Vector Scaphoideus Titanus. EFSA Support. Publ. 2020, 17, 1909E. [Google Scholar] [CrossRef]
- Kirdat, K.; Tiwarekar, B.; Thorat, V.; Sathe, S.; Shouche, Y.; Yadav, A. ‘Candidatus Phytoplasma Sacchari’, a Novel Taxon Associated with Sugarcane Grassy Shoot (SCGS) Disease. Int. J. Syst. Evol. Microbiol. 2021, 71, 4591. [Google Scholar] [CrossRef]
- Rodrigues Jardim, B.; Kinoti, W.M.; Tran-Nguyen, L.T.T.; Gambley, C.; Rodoni, B.; Constable, F.E. “Candidatus Phytoplasma Stylosanthis”, a Novel Taxon with a Diverse Host Range in Australia, Characterised Using Multilocus Sequence Analysis of 16S RRNA, SecA, Tuf, and Rp Genes. Int. J. Syst. Evol. Microbiol. 2021, 71, 4589. [Google Scholar] [CrossRef]
- Zhao, Y.; Wei, W.; Davis, R.E.; Lee, I.-M.; Bottner-Parker, K.D. The Agent Associated with Blue Dwarf Disease in Wheat Represents a New Phytoplasma Taxon, “Candidatus Phytoplasma Tritici”. Int. J. Syst. Evol. Microbiol. 2021, 71, 4604. [Google Scholar] [CrossRef]
- Jones, L.M.; Pease, B.; Perkins, S.L.; Constable, F.E.; Kinoti, W.M.; Warmington, D.; Allgood, B.; Powell, S.; Taylor, P.; Pearce, C.; et al. “Candidatus Phytoplasma Dypsidis”, a Novel Taxon Associated with a Lethal Wilt Disease of Palms in Australia. Int. J. Syst. Evol. Microbiol. 2021, 71, 4818. [Google Scholar] [CrossRef] [PubMed]
- Marcone, C. Molecular Biology and Pathogenicity of Phytoplasmas. Ann. Appl. Biol. 2014, 165, 199–221. [Google Scholar] [CrossRef]
- Tomkins, M.; Kliot, A.; Marée, A.F.; Hogenhout, S.A. A Multi-Layered Mechanistic Modelling Approach to Understand How Effector Genes Extend beyond Phytoplasma to Modulate Plant Hosts, Insect Vectors and the Environment. Curr. Opin. Plant Biol. 2018, 44, 39–48. [Google Scholar] [CrossRef]
- Trivellone, V. An Online Global Database of Hemiptera -Phytoplasma-Plant Biological Interactions. Biodivers. Data J. 2019, 7, e32910. [Google Scholar] [CrossRef] [PubMed]
- Caudwell, A.; Kuszala, C.; Larrue, J.; Bachelier, J. Transmission de La Flavescence Dorée de La Fève à La Fève Par Des Cicadelles Des Genres Euscelis et Euscelidius. Ann. Phytopathol. 1972, 1572, 181–189. [Google Scholar]
- Weintraub, P.G.; Beanland, L. Insect Vectors of Phytoplasmas. Annu. Rev. Entomol. 2006, 51, 91–111. [Google Scholar] [CrossRef] [PubMed]
- Conti, M.; D’Agostino, G.; Casetta, A.; Mela, L. Some Characteristics of Chrysanthemum Yellows Disease. Acta Horticult. 1988, 234, 129–136. [Google Scholar] [CrossRef]
- Christiaens, O.; Whyard, S.; Vélez, A.M.; Smagghe, G. Double-Stranded RNA Technology to Control Insect Pests: Current Status and Challenges. Front. Plant Sci. 2020, 11, 451. [Google Scholar] [CrossRef]
- Lim, F.-H.; Rasid, O.A.; As’wad, A.W.M.; Vadamalai, G.; Parveez, G.K.A.; Wong, M.-Y. The Future Is Now: Revolution of RNA-Mediated Gene Silencing in Plant Protection against Insect Pests and Diseases. Plant Biotechnol. Rep. 2020, 14, 643–662. [Google Scholar] [CrossRef]
- Liu, S.; Jaouannet, M.; Dempsey, D.A.; Imani, J.; Coustau, C.; Kogel, K.-H. RNA-Based Technologies for Insect Control in Plant Production. Biotechnol. Adv. 2020, 39, 107463. [Google Scholar] [CrossRef]
- Zhu, K.Y.; Palli, S.R. Mechanisms, Applications, and Challenges of Insect RNA Interference. Annu. Rev. Entomol. 2020, 65, 293–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taning, C.N.; Arpaia, S.; Christiaens, O.; Dietz-Pfeilstetter, A.; Jones, H.; Mezzetti, B.; Sabbadini, S.; Sorteberg, H.; Sweet, J.; Ventura, V.; et al. RNA-based Biocontrol Compounds: Current Status and Perspectives to Reach the Market. Pest Manag. Sci. 2020, 76, 841–845. [Google Scholar] [CrossRef]
- Zotti, M.; dos Santos, E.A.; Cagliari, D.; Christiaens, O.; Taning, C.N.T.; Smagghe, G. RNA Interference Technology in Crop Protection against Arthropod Pests, Pathogens and Nematodes. Pest Manag. Sci. 2018, 74, 1239–1250. [Google Scholar] [CrossRef]
- Kanakala, S.; Ghanim, M. RNA Interference in Insect Vectors for Plant Viruses. Viruses 2016, 8, 329. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Redinbaugh, M.G.; Michel, A.P. Molecular Interactions and Immune Responses between Maize Fine Streak Virus and the Leafhopper Vector Graminella Nigrifrons through Differential Expression and RNA Interference: Virus Defense and RNAi in G. Nigrifons. Insect Mol. Biol. 2015, 24, 391–401. [Google Scholar] [CrossRef]
- Xue, W.-H.; Liu, Y.-L.; Jiang, Y.-Q.; He, S.-F.; Wang, Q.-Q.; Yang, Z.-N.; Xu, H.-J. Molecular Characterization of Insulin-like Peptides in the Brown Planthopper, Nilaparvata lugens (Hemiptera: Delphacidae). Insect Mol. Biol. 2020, 29, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, Y.; Hattori, M. Gene Silencing by Parental RNA Interference in the Green Rice Leafhopper, Nephotettix cincticeps (Hemiptera: Cicadellidae): Parental RNAi in Nephotettix Cincticeps. Arch. Insect Biochem. Physiol. 2016, 91, 152–164. [Google Scholar] [CrossRef]
- Abbà, S.; Galetto, L.; Ripamonti, M.; Rossi, M.; Marzachì, C. RNA Interference of Muscle Actin and ATP Synthase Beta Increases Mortality of the Phytoplasma Vector Euscelidius variegatus. Pest Manag. Sci. 2019, 75, 1425–1434. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Peng, Y.; Zhang, H.; Wang, K.; Zhao, C.; Zhu, G.; Reddy Palli, S.; Han, Z. Off-Target Effects of RNAi Correlate with the Mismatch Rate between DsRNA and Non-Target MRNA. RNA Biol. 2021, 18, 1747–1759. [Google Scholar] [CrossRef] [PubMed]
- Arora, A.K.; Chung, S.H.; Douglas, A.E. Non-Target Effects of DsRNA Molecules in Hemipteran Insects. Genes 2021, 12, 407. [Google Scholar] [CrossRef]
- Papadopoulou, N.; Devos, Y.; Álvarez-Alfageme, F.; Lanzoni, A.; Waigmann, E. Risk Assessment Considerations for Genetically Modified RNAi Plants: EFSA’s Activities and Perspective. Front. Plant Sci. 2020, 11, 445. [Google Scholar] [CrossRef] [Green Version]
- Galetto, L.; Abbà, S.; Rossi, M.; Ripamonti, M.; Palmano, S.; Bosco, D.; Marzachì, C. Silencing of ATP Synthase β Reduces Phytoplasma Multiplication in a Leafhopper Vector. J. Insect Physiol. 2021, 128, 104176. [Google Scholar] [CrossRef]
- Galetto, L.; Ripamonti, M.; Abbà, S.; Rossi, M.; Manfredi, M.; Bosco, D.; Marzachì, C. Silencing of ATP Synthase β Induces Female Sterility in a Leafhopper Phytoplasma Vector. Entomol. General. 2021, 41, 497–510. [Google Scholar] [CrossRef]
- Galetto, L.; Bosco, D.; Balestrini, R.; Genre, A.; Fletcher, J.; Marzachì, C. The Major Antigenic Membrane Protein of “Candidatus Phytoplasma Asteris” Selectively Interacts with ATP Synthase and Actin of Leafhopper Vectors. PLoS ONE 2011, 6, e22571. [Google Scholar] [CrossRef] [Green Version]
- Rashidi, M.; Galetto, L.; Bosco, D.; Bulgarelli, A.; Vallino, M.; Veratti, F.; Marzachì, C. Role of the Major Antigenic Membrane Protein in Phytoplasma Transmission by Two Insect Vector Species. BMC Microbiol. 2015, 15, 193. [Google Scholar] [CrossRef] [Green Version]
- Ottati, S.; Chiapello, M.; Galetto, L.; Bosco, D.; Marzachì, C.; Abbà, S. New Viral Sequences Identified in the Flavescence Dorée Phytoplasma Vector Scaphoideus titanus. Viruses 2020, 12, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowling, D.; Pauli, T.; Donath, A.; Meusemann, K.; Podsiadlowski, L.; Petersen, M.; Peters, R.S.; Mayer, C.; Liu, S.; Zhou, X.; et al. Phylogenetic Origin and Diversification of RNAi Pathway Genes in Insects. Genome Biol. Evol. 2017, 8, 3784–3793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joga, M.R.; Zotti, M.J.; Smagghe, G.; Christiaens, O. RNAi Efficiency, Systemic Properties, and Novel Delivery Methods for Pest Insect Control: What We Know so Far. Front. Physiol. 2016, 7, 553. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, A.; Anfora, G.; Lucchi, A.; Lanzo, F.; Virant-Doberlet, M.; Mazzoni, V. Exploitation of Insect Vibrational Signals Reveals a New Method of Pest Management. PLoS ONE 2012, 7, e32954. [Google Scholar] [CrossRef] [PubMed]
- Tacoli, F.; Mori, N.; Pozzebon, A.; Cargnus, E.; Da Vià, S.; Zandigiacomo, P.; Duso, C.; Pavan, F. Control of Scaphoideus Titanus with Natural Products in Organic Vineyards. Insects 2017, 8, 129. [Google Scholar] [CrossRef] [Green Version]
- Gonella, E.; Crotti, E.; Mandrioli, M.; Daffonchio, D.; Alma, A. Asaia Symbionts Interfere with Infection by Flavescence Dorée Phytoplasma in Leafhoppers. J. Pest Sci. 2018, 91, 1033–1046. [Google Scholar] [CrossRef]
- Arraes, F.B.M.; Martins-de-Sa, D.; Noriega Vasquez, D.D.; Melo, B.P.; Faheem, M.; de Macedo, L.L.P.; Morgante, C.V.; Barbosa, J.A.R.G.; Togawa, R.C.; Moreira, V.J.V.; et al. Dissecting Protein Domain Variability in the Core RNA Interference Machinery of Five Insect Orders. RNA Biol. 2020, 18, 1653–1681. [Google Scholar] [CrossRef]
- Bucher, G.; Scholten, J.; Klingler, M. Parental RNAi in Tribolium (Coleoptera). Curr. Biol. 2002, 12, R85–R86. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.C.; Brown, S.J.; Tomoyasu, Y. Larval RNAi in Drosophila? Dev. Genes Evol. 2008, 218, 505–510. [Google Scholar] [CrossRef]
- Guan, R.-B.; Li, H.-C.; Fan, Y.-J.; Hu, S.-R.; Christiaens, O.; Smagghe, G.; Miao, X.-X. A Nuclease Specific to Lepidopteran Insects Suppresses RNAi. J. Biol. Chem. 2018, 293, 6011–6021. [Google Scholar] [CrossRef] [Green Version]
- Jaubert-Possamai, S.; Le Trionnaire, G.; Bonhomme, J.; Christophides, G.K.; Rispe, C.; Tagu, D. Gene Knockdown by RNAi in the Pea Aphid Acyrthosiphon pisum. BMC Biotechnol. 2007, 7, 63. [Google Scholar] [CrossRef] [PubMed]
- Coleman, A.D.; Wouters, R.H.M.; Mugford, S.T.; Hogenhout, S.A. Persistence and Transgenerational Effect of Plant-Mediated RNAi in Aphids. J. Exp. Bot. 2015, 66, 541–548. [Google Scholar] [CrossRef] [Green Version]
- Paim, R.M.M.; Araujo, R.N.; Lehane, M.J.; Gontijo, N.F.; Pereira, M.H. Long-Term Effects and Parental RNAi in the Blood Feeder Rhodnius prolixus (Hemiptera; Reduviidae). Insect Biochem. Mol. Biol. 2013, 43, 1015–1020. [Google Scholar] [CrossRef]
- Bogenhagen, D.F.; Haley, J.D. Pulse-Chase SILAC–Based Analyses Reveal Selective Oversynthesis and Rapid Turnover of Mitochondrial Protein Components of Respiratory Complexes. J. Biol. Chem. 2020, 295, 2544–2554. [Google Scholar] [CrossRef] [Green Version]
- Martins, J.R.; Anhezini, L.; Dallacqua, R.P.; Simões, Z.L.P.; Bitondi, M.M.G. A Honey Bee Hexamerin, HEX 70a, Is Likely to Play an Intranuclear Role in Developing and Mature Ovarioles and Testioles. PLoS ONE 2011, 6, e29006. [Google Scholar] [CrossRef] [Green Version]
- Hatle, J.D.; Borst, D.W.; Eskew, M.R.; Juliano, S.A. Maximum Titers of Vitellogenin and Total Hemolymph Protein Occur during the Canalized Phase of Grasshopper Egg Production. Physiol. Biochem. Zool. 2001, 74, 885–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hathaway, M.; Hatle, J.; Li, S.; Ding, X.; Barry, T.; Hong, F.; Wood, H.; Borst, D. Characterization of Hexamerin Proteins and Their MRNAs in the Adult Lubber Grasshopper: The Effects of Nutrition and Juvenile Hormone on Their Levels. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2009, 154, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.L.; Telfer, W.H. Storage Hexamer Utilization in Two Lepidopterans: Differences Correlated with the Timing of Egg Formation. J. Insect Sci. 2001, 1, 2. [Google Scholar] [CrossRef]
- Wheeler, D.E.; Buck, N.A. A Role for Storage Proteins in Autogenous Reproduction in Aedes atropalpus. J. Insect Physiol. 1996, 42, 961–966. [Google Scholar] [CrossRef]
- Saikhedkar, N.; Summanwar, A.; Joshi, R.; Giri, A. Cathepsins of Lepidopteran Insects: Aspects and Prospects. Insect Biochem. Mol. Biol. 2015, 64, 51–59. [Google Scholar] [CrossRef]
- Waniek, P.J.; Pacheco Costa, J.E.; Jansen, A.M.; Costa, J.; Araújo, C.A.C. Cathepsin L of Triatoma brasiliensis (Reduviidae, Triatominae): Sequence Characterization, Expression Pattern and Zymography. J. Insect Physiol. 2012, 58, 178–187. [Google Scholar] [CrossRef] [Green Version]
- Irles, P.; Bellés, X.; Piulachs, M.D. Identifying Genes Related to Choriogenesis in Insect Panoistic Ovaries by Suppression Subtractive Hybridization. BMC Genom. 2009, 10, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.; Chen, L.; Zhang, F.; Wu, P.; Li, E.; Qin, J. Molecular Characterization of Cathepsin L CDNA and Its Expression during Oogenesis and Embryogenesis in the Oriental River Prawn Macrobrachium nipponense (Palaemonidae). Genet. Mol. Res. 2013, 12, 5215–5225. [Google Scholar] [CrossRef] [PubMed]
- Trivellone, V.; Ripamonti, M.; Angelini, E.; Filippin, L.; Rossi, M.; Marzachí, C.; Galetto, L. Evidence Suggesting Interactions between Immunodominant Membrane Protein Imp of Flavescence Dorée Phytoplasma and Protein Extracts from Distantly Related Insect Species. J. Appl. Microbiol. 2019, 127, 1801–1813. [Google Scholar] [CrossRef]
- Leyva, J.A.; Bianchet, M.A.; Amzel, L.M. Understanding ATP Synthesis: Structure and Mechanism of the F1-ATPase. Mol. Membr. Biol. 2003, 20, 27–33. [Google Scholar] [CrossRef]
- Bachman, P.M.; Bolognesi, R.; Moar, W.J.; Mueller, G.M.; Paradise, M.S.; Ramaseshadri, P.; Tan, J.; Uffman, J.P.; Warren, J.; Wiggins, B.E.; et al. Characterization of the Spectrum of Insecticidal Activity of a Double-Stranded RNA with Targeted Activity against Western Corn Rootworm (Diabrotica virgifera LeConte). Transgenic Res. 2013, 22, 1207–1222. [Google Scholar] [CrossRef] [Green Version]
- Whyard, S.; Singh, A.D.; Wong, S. Ingested Double-Stranded RNAs Can Act as Species-Specific Insecticides. Insect Biochem. Mol. Biol. 2009, 39, 824–832. [Google Scholar] [CrossRef]
- Jain, R.G.; Robinson, K.E.; Fletcher, S.J.; Mitter, N. RNAi-Based Functional Genomics in Hemiptera. Insects 2020, 11, 557. [Google Scholar] [CrossRef] [PubMed]
- Mitter, N.; Worrall, E.A.; Robinson, K.E.; Li, P.; Jain, R.G.; Taochy, C.; Fletcher, S.J.; Carroll, B.J.; Lu, G.Q.; Xu, Z.P. Clay Nanosheets for Topical Delivery of RNAi for Sustained Protection against Plant Viruses. Nature Plants 2017, 3, 16207. [Google Scholar] [CrossRef]
- Kunte, N.; McGraw, E.; Bell, S.; Held, D.; Avila, L.-A. Prospects, Challenges and Current Status of RNAi through Insect Feeding. Pest Manag. Sci. 2020, 76, 26–41. [Google Scholar] [CrossRef]
- Yu, X.; Killiny, N. RNA Interference-Mediated Control of Asian Citrus Psyllid, the Vector of the Huanglongbing Bacterial Pathogen. Trop. Plant Pathol. 2020, 45, 298–305. [Google Scholar] [CrossRef]
- Yan, S.; Ren, B.; Zeng, B.; Shen, J. Improving RNAi Efficiency for Pest Control in Crop Species. BioTechniques 2020, 68, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Bennett, M.; Deikman, J.; Hendrix, B.; Iandolino, A. Barriers to Efficient Foliar Uptake of DsRNA and Molecular Barriers to DsRNA Activity in Plant Cells. Front. Plant Sci. 2020, 11, 816. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.K.B.; Hunter, W.B.; Park, A.L.; Gundersen-Rindal, D.E. Double Strand RNA Delivery System for Plant-Sap-Feeding Insects. PLoS ONE 2017, 12, e0171861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrade, E.C.; Hunter, W.B. RNAi Feeding Bioassay: Development of a Non-Transgenic Approach to Control Asian Citrus Psyllid and Other Hemipterans. Entomol. Exp. Appl. 2017, 162, 389–396. [Google Scholar] [CrossRef]
- Pampolini, F.; Rodrigues, T.B.; Leelesh, R.S.; Kawashima, T.; Rieske, L.K. Confocal Microscopy Provides Visual Evidence and Confirms the Feasibility of DsRNA Delivery to Emerald Ash Borer through Plant Tissues. J. Pest Sci. 2020, 93, 1143–1153. [Google Scholar] [CrossRef]
- Chuche, J.; Sauvion, N.; Thiéry, D. Mixed Xylem and Phloem Sap Ingestion in Sheath-Feeders as Normal Dietary Behavior: Evidence from the Leafhopper Scaphoideus Titanus. J. Insect Physiol. 2017, 102, 62–72. [Google Scholar] [CrossRef]
- Ripamonti, M.; Maron, F.; Cornara, D.; Marzachì, C.; Fereres, A.; Bosco, D. Scaphoideus titanus Ball Feeding Behaviour on Three Grapevine Cultivars with Different Susceptibilities to Flavescence Dorée. bioRxiv 2021. [Google Scholar] [CrossRef]
- Lett, J.-M.; Granier, M.; Grondin, M.; Turpin, P.; Molinaro, F.; Chiroleu, F.; Peterschmitt, M.; Reynaud, B. Electrical Penetration Graphs from Cicadulina Mbila on Maize, the Fine Structure of Its Stylet Pathways and Consequences for Virus Transmission Efficiency. Entomol. Exp. Appl. 2001, 101, 93–109. [Google Scholar] [CrossRef]
- Stafford, C.A.; Walker, G.P. Characterization and Correlation of DC Electrical Penetration Graph Waveforms with Feeding Behavior of Beet Leafhopper, Circulifer tenellus. Entomol. Exp. Appl. 2009, 130, 113–129. [Google Scholar] [CrossRef]
- Carpane, P.; Wayadande, A.; Backus, E.; Dolezal, W.; Fletcher, J. Characterization and Correlation of New Electrical Penetration Graph Waveforms for the Corn Leafhopper (Hemiptera: Cicadellidae). Ann. Entom. Soc. Amer. 2011, 104, 515–525. [Google Scholar] [CrossRef] [Green Version]
- Trębicki, P.; Tjallingii, W.F.; Harding, R.M.; Rodoni, B.C.; Powell, K.S. EPG Monitoring of the Probing Behaviour of the Common Brown Leafhopper Orosius orientalis on Artificial Diet and Selected Host Plants. Arthropod-Plant Inte. 2012, 6, 405–415. [Google Scholar] [CrossRef] [Green Version]
- Mezzetti, B.; Smagghe, G.; Arpaia, S.; Christiaens, O.; Dietz-Pfeilstetter, A.; Jones, H.; Kostov, K.; Sabbadini, S.; Opsahl-Sorteberg, H.-G.; Ventura, V.; et al. RNAi: What Is Its Position in Agriculture? J. Pest Sci. 2020, 93, 1125–1130. [Google Scholar] [CrossRef]
- Lindbo, J.A. TRBO: A High-Efficiency Tobacco Mosaic Virus RNA-Based Overexpression Vector. Plant Physiol. 2007, 145, 1232–1240. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, S.; Oshima, K.; Kakizawa, S.; Arashida, R.; Jung, H.-Y.; Yamaji, Y.; Nishigawa, H.; Ugaki, M.; Namba, S. Interaction between the Membrane Protein of a Pathogen and Insect Microfilament Complex Determines Insect-Vector Specificity. Proc. Natl. Acad. Sci. USA 2006, 103, 4252–4257. [Google Scholar] [CrossRef] [Green Version]
- Candiano, G.; Bruschi, M.; Musante, L.; Santucci, L.; Ghiggeri, G.M.; Carnemolla, B.; Orecchia, P.; Zardi, L.; Righetti, P.G. Blue Silver: A Very Sensitive Colloidal Coomassie G-250 Staining for Proteome Analysis. Electrophoresis 2004, 25, 1327–1333. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Names | Scaphoideus titanus | Euscelidius variegatus1 | Percent Identity | Query Coverage |
|---|---|---|---|---|
| AGO1 | MZ161181 | GFTU01004640.1 | 99 | 99 |
| AGO2 | MZ161183 2 | GFTU01008426.1 2 | 85 | 92 |
| AGO3 | MZ161185 | GFTU01016377.1 2 | 86 | 84 |
| Dicer1 | MZ161180 2 | GFTU01004888.1 2 | 90 | 99 |
| Dicer2 | MZ161186 | GFTU01010822.1 | 70 | 99 |
| Drosha | MZ161187 | GFTU01002931.1 2 | 67 | 71 |
| Loquacious | MZ161182 | GFTU01010148.1 | 84 | 100 |
| Pasha | MZ161189 | GFTU01010211.1 | 80 | 97 |
| Piwi | MZ161188 | GFTU01014046.1 | 88 | 100 |
| R2D2 | MZ161184 | GFTU01005523.1 | 65 | 99 |
| Insect Species | Order | Percent Identity with St_dsATP1 | Percent Identity with St_dsATP2 |
|---|---|---|---|
| Euscelidius variegatus | Hemiptera | 87.63 | 88.72 |
| Metcalfa pruinosa | 84.79 | 66.15 | |
| Halyomorpha halys | 83.12 | 66.67 | |
| Frankliniella occidentalis | Thysanoptera | 86.88 | 55.90 |
| Helicoverpa armigera | Lepidoptera | 86.88 | 51.85 |
| Ostrinia furnacalis | 84.79 | 44.62 | |
| Nasonia vitripennis | Hymenoptera | 85.42 | 55.19 |
| Apis mellifera | 79.38 | 58.62 | |
| Locusta migratoria | Orthoptera | 85.00 | 54.26 |
| Dendroctonus ponderosae | Coleoptera | 84.58 | 51.55 |
| Zootermopsis nevadensis | Dictyoptera | 82.92 | 55.38 |
| Drosophila melanogaster | Diptera | 81.25 | 47.02 |
| dsRNA Injected | Dose of dsRNA (ng/insect) | N° F0 Parental Insects | N° F1 Offspring | Mean Ratio of Offspring/Parental Female | ||
|---|---|---|---|---|---|---|
| Males | Females | Nymphs | Adults | |||
| dsGFP | 80 | 2 | 12 | 80 | 2 | 6.83 |
| 8 | 5 | 18 | 128 | 18 | 8.11 | |
| Eva_dsATP1 | 80 | 1 | 6 | 0 | 0 | - |
| 8 | 4 | 9 | 0 | 5 | 0.55 | |
| Eva_dsATP2 | 80 | 1 | 5 | 0 | 0 | - |
| 8 | 4 | 6 | 0 | 0 | - | |
| St_dsATP1 | 80 | 5 | 8 | 0 | 0 | - |
| 8 | 4 | 8 | 0 | 4 | 0.50 | |
| St_dsATP2 | 80 | 1 | 8 | 0 | 0 | - |
| 8 | 2 | 3 | 2 | 0 | 0.67 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ripamonti, M.; Cerone, L.; Abbà, S.; Rossi, M.; Ottati, S.; Palmano, S.; Marzachì, C.; Galetto, L. Silencing of ATP Synthase β Impairs Egg Development in the Leafhopper Scaphoideus titanus, Vector of the Phytoplasma Associated with Grapevine Flavescence Dorée. Int. J. Mol. Sci. 2022, 23, 765. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23020765
Ripamonti M, Cerone L, Abbà S, Rossi M, Ottati S, Palmano S, Marzachì C, Galetto L. Silencing of ATP Synthase β Impairs Egg Development in the Leafhopper Scaphoideus titanus, Vector of the Phytoplasma Associated with Grapevine Flavescence Dorée. International Journal of Molecular Sciences. 2022; 23(2):765. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23020765
Chicago/Turabian StyleRipamonti, Matteo, Luca Cerone, Simona Abbà, Marika Rossi, Sara Ottati, Sabrina Palmano, Cristina Marzachì, and Luciana Galetto. 2022. "Silencing of ATP Synthase β Impairs Egg Development in the Leafhopper Scaphoideus titanus, Vector of the Phytoplasma Associated with Grapevine Flavescence Dorée" International Journal of Molecular Sciences 23, no. 2: 765. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23020765