
A Brief Review of In Vitro Models for Injury and Regeneration in the Peripheral Nervous System


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
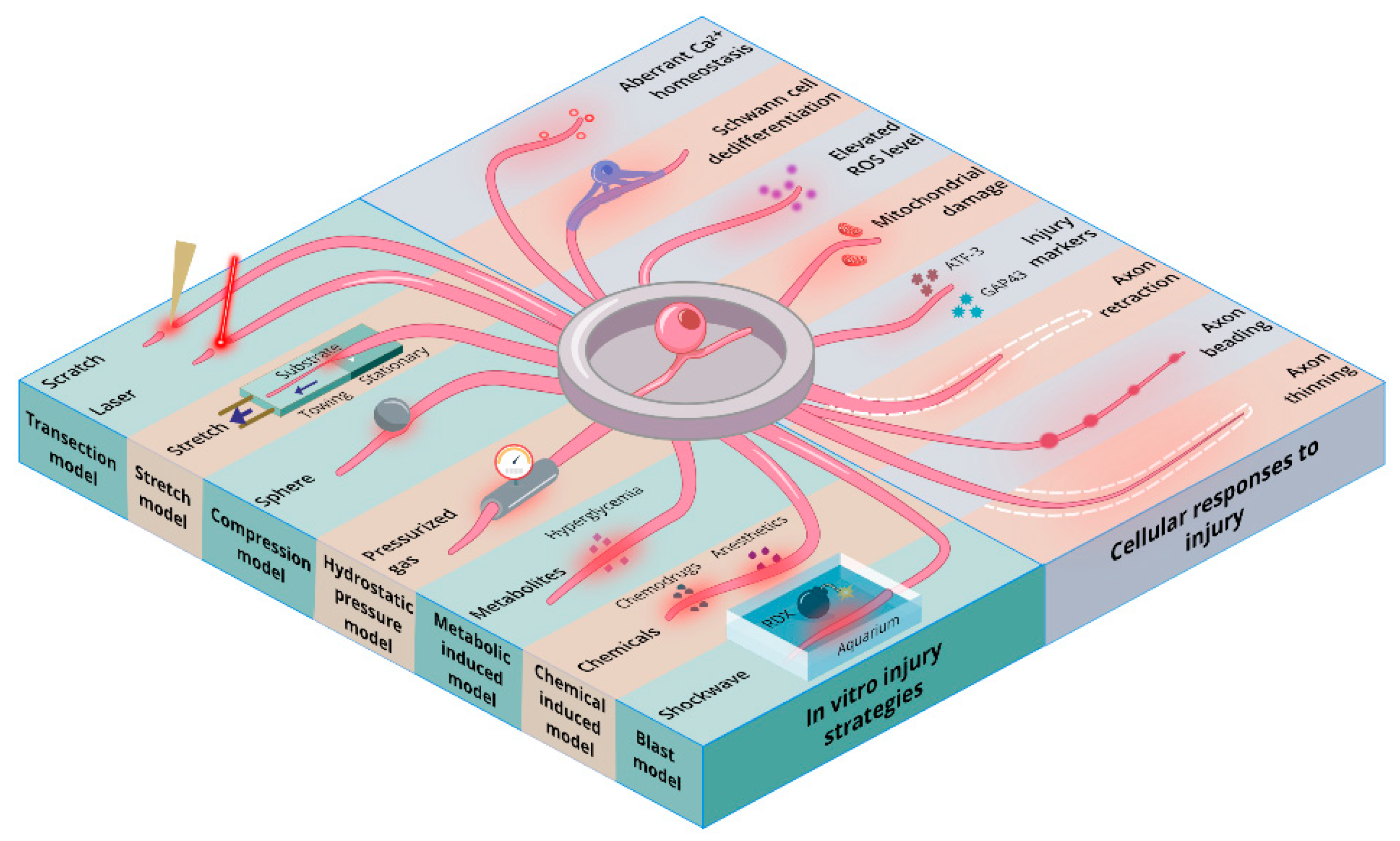
1.1. In Vitro Nerve Injury: A Window to Observe Cellular Mechanisms
1.2. PNI-Associated Cellular Changes Observed In Vitro
2. Transection Injury Models
2.1. The Campenot Chamber Models
2.2. Microfluidic Chamber Models
2.3. Forceful Trituration
2.4. Tear on Cell Monolayer
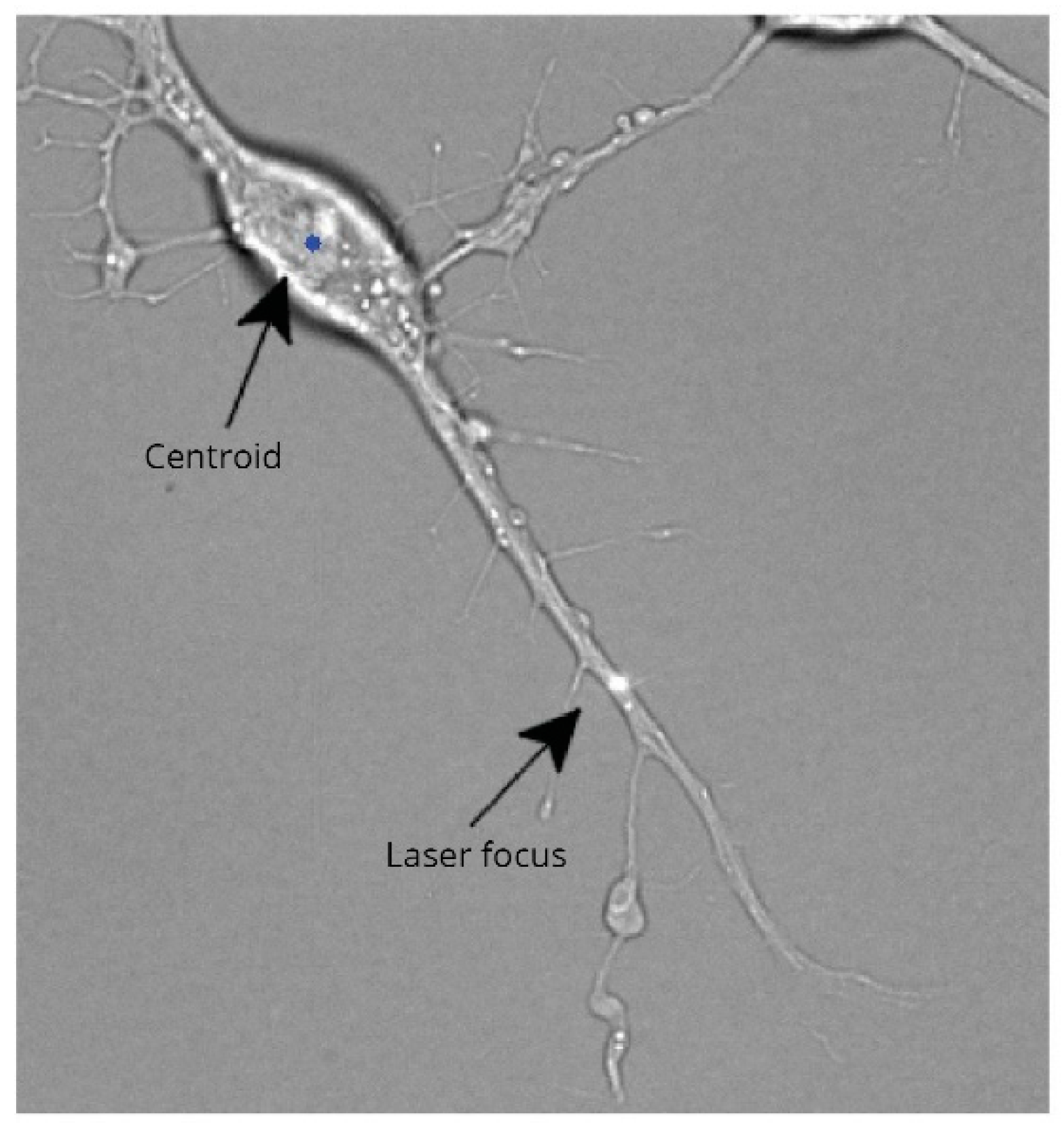
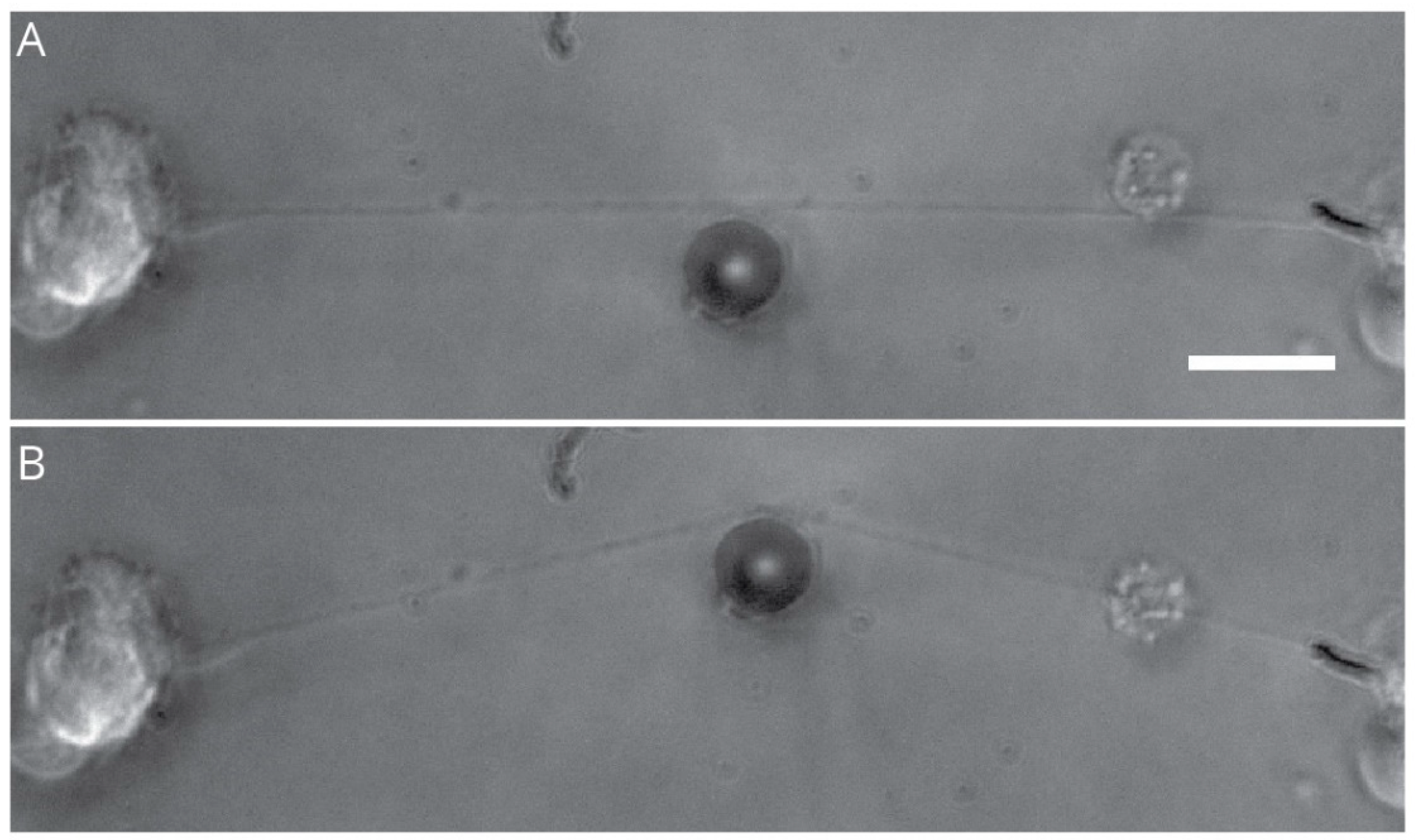
2.5. Laser-Induced Transection
2.6. Advantages
- Compartmentalization of peripheral neuron cultures provides robust separation of axon and soma that morphologically reflects a more in vivo arrangement.
- The spatial separation also allows for the application of consistent injury events, while the cellular response can be easily visualized and quantified using conventional approaches.
- In addition to being cost-effective, this model also allows for the performance of various biochemical assays that require higher volumes of injured cells.
- The use of laser-induced transection provides precision and accuracy for single-cell studies.
2.7. Disadvantages
- Though the compartmental models are quite easy to set up, these devices are prone to fluid leak that may warrant repetition of experiments.
- There may be difficulty in the assembly of the device for advanced live-cell microscopy, and necessary modifications may have to be incorporated.
- Single-cell studies using laser-induced injuries will need advanced expertise for set-up and may not be possible in a typical laboratory setting.
- It is quite challenging to limit glial cells within compartments, and eventually, they tend to migrate into all compartments.
3. Axonal Stretch Injury Model
3.1. Advantages
- Direct and real-time measurement of controlled mechanical strain.
- The cellular response is reproducible, and the principles behind stretching mechanisms are quite simple.
3.2. Disadvantages
- Difficulty in mimicking various injury dynamics as seen in real-life conditions, where nerves may be exposed to stretching and twisting force.
- Techniques such as incorporating a PDMS membrane within microfluidic systems requires experience and advanced skills.
- Sum total of tensile strength of individual axons, being lower than the tensile strength of an entire nerve, makes in vitro models less relatable to in vivo observations.
4. Compression Induced-Injury Models
4.1. Advantages
- Reasonable similarity of in-vitro-induced cellular changes compared to in vivo experiments.
4.2. Disadvantages
- Model offers limited insight into the biomechanics of the nerve injury because of the difficulty in measuring the strain produced at the site of injury and the rate of application of the strain.
- Since the biomechanical force of injury varies, it is difficult to predict and control the lesions formed and control which axons are affected and which are spared.
5. Hydrostatic Pressure Models
5.1. Pressurized Chambers
5.2. Hydrostatic Fluid Columns
5.3. Syringe Pump System
5.4. Advantages
- Cells can be subjected to a wide variety of controlled hydrostatic pressure ranges.
- In vitro platform helps in understanding the underlying mechanisms of CNS injuries, glaucoma, and cardiovascular research.
5.5. Disadvantages
- There may be design complexity with the necessity of a pump system in a standard cell culture set-up.
- Fluid flow may induce shear stress in cells.
6. Metabolic Nerve Injury Models
6.1. Advantages
- Inexpensive and easy to perform.
- Can be utilized to study the effect of hyperglycemia on specific cell types of the peripheral nervous system.
6.2. Disadvantages
- Pathogenesis of diabetes-induced nerve injury goes beyond merely the hyperglycemia condition. Hence, an in vitro model of hyperglycemic growth media may not exactly mimic the multi-component involvement of metabolic disorders as observed in vivo.
7. Chemically Induced Nerve Injury Models
7.1. Advantages
- Effect of chemotherapeutics on non-neuronal cells and its subsequent effect on neurons can be studied separately.
7.2. Disadvantages
- Caution to be exercised when analyzing results from CIPN in vitro models, mainly because cancer, being a multi-component disease, will invariably have unknown variables that may be missed.
8. Blast-Induced Injury Models
8.1. Advantages
- Functional or biochemical measures can be taken after injury at multiple time points.
8.2. Disadvantages
- Limitations in stimulating shock wave under special circumstances such as trenches, cabin, and underwater explosions.
- Experiments with compounds such as RDX may require additional regulatory approvals and specialized safety oversight.
9. Challenges and Future Perspectives
10. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Qian, C.; Zhou, F.-Q. Updates and challenges of axon regeneration in the mammalian central nervous system. J. Mol. Cell Biol. 2020, 12, 798–806. [Google Scholar] [CrossRef]
- Rosso, G.; Guck, J. Mechanical changes of peripheral nerve tissue microenvironment and their structural basis during development. APL Bioeng. 2019, 3, 036107. [Google Scholar] [CrossRef] [Green Version]
- Park, S.E.; Ahn, J.; Jeong, H.-E.; Youn, I.; Huh, D.; Chung, S. A Three-dimensional in vitro model of the peripheral nervous system. NPG Asia Mater. 2021, 13, 2. [Google Scholar] [CrossRef]
- Babetto, E.; Wong, K.M.; Beirowski, B. A glycolytic shift in schwann cells supports injured axons. Nat. Neurosci. 2020, 23, 1215–1228. [Google Scholar] [CrossRef] [PubMed]
- Fawcett, J.W. The struggle to make CNS axons regenerate: Why has it been so difficult? Neurochem. Res. 2020, 45, 144–158. [Google Scholar] [CrossRef] [Green Version]
- Fawcett, J.W.; Verhaagen, J. Intrinsic determinants of axon regeneration. Devel. Neurobio. 2018, 78, 890–897. [Google Scholar] [CrossRef]
- Hussain, G.; Wang, J.; Rasul, A.; Anwar, H.; Qasim, M.; Zafar, S.; Aziz, N.; Razzaq, A.; Hussain, R.; de Aguilar, J.L.G.; et al. Current status of therapeutic approaches against peripheral nerve injuries: A detailed story from injury to recovery. Int. J. Biol. Sci. 2020, 16, 116–134. [Google Scholar] [CrossRef] [Green Version]
- Heary, K.O.; Wong, A.W.K.; Lau, S.C.L.; Dengler, J.; Thompson, M.R.; Crock, L.W.; Novak, C.B.; Philip, B.A.; Mackinnon, S.E. Quality of life and psychosocial factors as predictors of pain relief following nerve surgery. Hand 2020, 155894472091121. [Google Scholar] [CrossRef] [PubMed]
- Houshyar, S.; Bhattacharyya, A.; Shanks, R. Peripheral nerve conduit: Materials and structures. ACS Chem. Neurosci. 2019, 10, 3349–3365. [Google Scholar] [CrossRef]
- Gopalakrishnan-Prema, V.; Mohanan, A.; Shivaram, S.B.; Madhusudanan, P.; Raju, G.; Menon, D.; Shankarappa, S.A. Electrical stimulation of co-woven nerve conduit for peripheral neurite differentiation. Biomed. Mater. 2020, 15, 065015. [Google Scholar] [CrossRef]
- Raza, C.; Riaz, H.A.; Anjum, R.; Shakeel, N. ul A. Repair strategies for injured peripheral nerve: Review. Life Sci. 2020, 243, 117308. [Google Scholar] [CrossRef]
- Isaacs, J.; Cochran, A.R. Nerve transfers for peripheral nerve injury in the upper limb: A case-based review. Bone Jt. J. 2019, 101, 124–131. [Google Scholar] [CrossRef]
- Mohanty, C.; Bhat, D.; Devi, B. Use of animal models in peripheral nerve surgery and research. Neurol. India 2019, 67, 100. [Google Scholar] [CrossRef]
- Vela, F.; Martínez-Chacón, G.; Ballestín, A.; Campos, J.; Sánchez-Margallo, F.; Abellán, E. Animal models used to study direct peripheral nerve repair: A systematic review. Neural. Regen. Res. 2020, 15, 491. [Google Scholar] [CrossRef] [PubMed]
- Montanino, A.; Kleiven, S. Utilizing a structural mechanics approach to assess the primary effects of injury loads onto the axon and its components. Front. Neurol. 2018, 9, 643. [Google Scholar] [CrossRef] [PubMed]
- Romeo-Guitart, D.; Casas, C. Network-centric medicine for peripheral nerve injury: Treating the whole to boost endogenous mechanisms of neuroprotection and regeneration. Neural. Regen. Res. 2019, 14, 1122. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Cho, Y. Experimental model systems for understanding human axonal injury responses. Int. J. Mol. Sci. 2021, 22, 474. [Google Scholar] [CrossRef]
- Rigoni, M.; Montecucco, C. Animal models for studying motor axon terminal paralysis and recovery. J. Neurochem. 2017, 142, 122–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, R.M.; Post, A.; Hoshizaki, B.; Ramesh, K.T. A multiscale computational approach to estimating axonal damage under inertial loading of the head. J. Neurotrauma 2013, 30, 102–118. [Google Scholar] [CrossRef]
- Höke, A. Mechanisms of disease: What factors limit the success of peripheral nerve regeneration in humans? Nat. Clin. Pract. Neurol. 2006, 2, 448–454. [Google Scholar] [CrossRef]
- Navarro, X.; Vivó, M.; Valero-Cabré, A. Neural plasticity after peripheral nerve injury and regeneration. Prog. Neurobiol. 2007, 82, 163–201. [Google Scholar] [CrossRef]
- Elsayed, H.; Faroni, A.; Ashraf, M.R.; Osuji, J.; Wunderley, L.; Zhang, L.; Elsobky, H.; Mansour, M.; Zidan, A.S.; Reid, A.J. Development and characterisation of an in vitro model of wallerian degeneration. Front. Bioeng. Biotechnol. 2020, 8, 784. [Google Scholar] [CrossRef] [PubMed]
- Jessen, K.R.; Arthur-Farraj, P. Repair schwann cell update: Adaptive reprogramming, EMT, and stemness in regenerating nerves. Glia 2019, 67, 421–437. [Google Scholar] [CrossRef]
- Pellegatta, M.; Taveggia, C. The complex work of proteases and secretases in wallerian degeneration: Beyond neuregulin-1. Front. Cell. Neurosci. 2019, 13, 93. [Google Scholar] [CrossRef] [PubMed]
- Modrak, M.; Talukder, M.A.H.; Gurgenashvili, K.; Noble, M.; Elfar, J.C. Peripheral Nerve Injury and Myelination: Potential Therapeutic Strategies. J. Neurosci. Res. 2020, 98, 780–795. [Google Scholar] [CrossRef]
- Renner, M.; Stute, G.; Alzureiqi, M.; Reinhard, J.; Wiemann, S.; Schmid, H.; Faissner, A.; Dick, H.B.; Joachim, S.C. Optic nerve degeneration after retinal ischemia/reperfusion in a rodent model. Front. Cell. Neurosci. 2017, 11, 254. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, Y.; Li, Y.; Segal, R.A. A Mechanistic understanding of axon degeneration in chemotherapy-induced peripheral neuropathy. Front. Neurosci. 2017, 11, 481. [Google Scholar] [CrossRef]
- Girouard, M.P.; Simas, T.; Hua, L.; Morquette, B.; Khazaei, M.R.; Unsain, N.; Johnstone, A.D.; Rambaldi, I.; Sanz, R.L.; Di Raddo, M.E.; et al. Collapsin response mediator protein 4 (Crmp4) facilitates wallerian degeneration and axon regeneration following sciatic nerve injury. Eneuro 2020, 7, 1–17. [Google Scholar] [CrossRef]
- Rodemer, W.; Selzer, M.E. Role of axon resealing in retrograde neuronal death and regeneration after spinal cord injury. Neural Regen. Res. 2019, 14, 399–404. [Google Scholar] [CrossRef]
- Canty, A.J.; Jackson, J.S.; Huang, L.; Trabalza, A.; Bass, C.; Little, G.; Tortora, M.; Khan, S.; De Paola, V. In vivo imaging of injured cortical axons reveals a rapid onset form of wallerian degeneration. BMC Biol. 2020, 18, 170. [Google Scholar] [CrossRef]
- Kerstein, P.C.; Patel, K.M.; Gomez, T.M. Calpain-mediated proteolysis of talin and FAK regulates adhesion dynamics necessary for axon guidance. J. Neurosci. 2017, 37, 1568–1580. [Google Scholar] [CrossRef] [Green Version]
- Blanquie, O.; Bradke, F. Cytoskeleton dynamics in axon regeneration. Curr. Opin. Neurobiol. 2018, 51, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Gu, C. Rapid and reversible development of axonal varicosities: A new form of neural plasticity. Front. Mol. Neurosci. 2021, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.E. A view from the ending: Axonal dieback and regeneration following SCI. Neurosci. Lett. 2017, 652, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Jukkola, P.; Wang, Q.; Esparza, T.; Zhao, Y.; Brody, D.; Gu, C. Polarity of varicosity initiation in central neuron mechanosensation. J. Cell Biol. 2017, 216, 2179–2199. [Google Scholar] [CrossRef] [Green Version]
- Shrirao, A.B.; Kung, F.H.; Omelchenko, A.; Schloss, R.S.; Boustany, N.N.; Zahn, J.D.; Yarmush, M.L.; Firestein, B.L. Microfluidic platforms for the study of neuronal injury in vitro. Biotechnol. Bioeng. 2018, 115, 815–830. [Google Scholar] [CrossRef]
- Wahane, S.; Halawani, D.; Zhou, X.; Zou, H. Epigenetic regulation of axon regeneration and glial activation in injury responses. Front. Genet. 2019, 10, 640. [Google Scholar] [CrossRef]
- Nagendran, T.; Larsen, R.S.; Bigler, R.L.; Frost, S.B.; Philpot, B.D.; Nudo, R.J.; Taylor, A.M. Distal axotomy enhances retrograde presynaptic excitability onto injured pyramidal neurons via trans-synaptic signaling. Nat. Commun. 2017, 8, 625. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, C.; Gan, C.; Zhao, K.; Chen, J.; Song, J.; Lei, T. A Precise, controllable in vitro model for diffuse axonal injury through uniaxial stretch injury. Front. Neurosci. 2019, 13, 1063. [Google Scholar] [CrossRef] [Green Version]
- Nagendran, T.; Taylor, A.M. Unique axon-to-soma signaling pathways mediate dendritic spine loss and hyper-excitability post-axotomy. Front. Cell. Neurosci. 2019, 13, 431. [Google Scholar] [CrossRef] [Green Version]
- Davies, A.J.; Kim, H.W.; Gonzalez-Cano, R.; Choi, J.; Back, S.K.; Roh, S.E.; Johnson, E.; Gabriac, M.; Kim, M.S.; Lee, J.; et al. Natural killer cells degenerate intact sensory afferents following nerve injury. Cell 2019, 176, 716–728. [Google Scholar] [CrossRef] [Green Version]
- Davies, A.J.; Rinaldi, S.; Costigan, M.; Oh, S.B. Cytotoxic immunity in peripheral nerve injury and pain. Front. Neurosci. 2020, 14, 142. [Google Scholar] [CrossRef]
- Boerboom, A.; Dion, V.; Chariot, A.; Franzen, R. Molecular mechanisms involved in schwann cell plasticity. Front. Mol. Neurosci. 2017, 10, 38. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.J.; Xu, R.J.; Ju, X.; Wang, W.H.; Luo, Z.P.; Liu, C.M.; Yang, L.; Li, B.; Chen, J.Q.; Meng, B.; et al. The telomerase reverse transcriptase (TERT) and P53 regulate mammalian PNS and CNS axon regeneration downstream of c-Myc. Biorxiv 2019, 39, 9107–9118. [Google Scholar] [CrossRef]
- Campenot, R.B. Local control of neurite development by nerve growth factor. Proc. Natl. Acad. Sci. USA 1977, 74, 4516–4519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campenot, R.B.; Lund, K.; Mok, S.A. Production of compartmented cultures of rat sympathetic neurons. Nat. Protoc. 2009, 4, 1869–1887. [Google Scholar] [CrossRef]
- Tierney, W.; Vicino, I.; Sun, S.; Chiu, W.; Engel, E.; Taylor, M.; Hogue, I. Methods and applications of campenot trichamber neuronal cultures for the study of neuroinvasive viruses. Life Sci. 2021. [Google Scholar] [CrossRef]
- Fantuzzo, J.A.; Hart, R.P.; Zahn, J.D.; Pang, Z.P. Compartmentalized devices as tools for investigation of human brain network dynamics: Compartmentalized devices to study human brain network dynamics. Dev. Dyn. 2019, 248, 65–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klusch, A.; Gorzelanny, C.; Reeh, P.W.; Schmelz, M.; Petersen, M.; Sauer, S.K. Local NGF and GDNF levels modulate morphology and function of porcine DRG neurites, in vitro. PLoS ONE 2018, 13, e0203215. [Google Scholar] [CrossRef]
- Bertrand, J.; Winton, M.J.; Rodriguez-Hernandez, N.; Campenot, R.B.; McKerracher, L. Application of Rho antagonist to neuronal cell bodies promotes neurite growth in compartmented cultures and regeneration of retinal ganglion cell axons in the optic nerve of adult rats. J. Neurosci. 2005, 25, 1113–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimpinski, K.; Campenot, R.B.; Mearow, K. Effects of the neurotrophins nerve growth factor, neurotrophin-3, and brain-derived neurotrophic factor (BDNF) on neurite growth from adult sensory neurons in compartmented cultures. J. Neurobiol. 1997, 33, 395–410. [Google Scholar] [CrossRef]
- MacInnis, B.L.; Senger, D.L.; Campenot, R.B. Spatial requirements for TrkA kinase activity in the support of neuronal survival and axon growth in rat sympathetic neurons. Neuropharmacology 2003, 45, 995–1010. [Google Scholar] [CrossRef]
- Ohtake, Y.; Matsuhisa, K.; Kaneko, M.; Kanemoto, S.; Asada, R.; Imaizumi, K.; Saito, A. Axonal activation of the unfolded protein response promotes axonal regeneration following peripheral nerve injury. Neuroscience 2018, 375, 34–48. [Google Scholar] [CrossRef] [PubMed]
- Jocher, G.; Mannschatz, S.H.; Offterdinger, M.; Schweigreiter, R. Microfluidics of small-population neurons allows for a precise quantification of the peripheral axonal growth state. Front. Cell. Neurosci. 2018, 12, 166. [Google Scholar] [CrossRef] [Green Version]
- Katiyar, N.; Raju, G.; Madhusudanan, P.; Gopalakrishnan-Prema, V.; Shankarappa, S.A. Neuronal delivery of nanoparticles via nerve fibres in the skin. Sci. Rep. 2021, 11, 2566. [Google Scholar] [CrossRef]
- Atmaramani, R.; Veeramachaneni, S.; Mogas, L.V.; Koppikar, P.; Black, B.J.; Hammack, A.; Pancrazio, J.J.; Granja-Vazquez, R. Investigating the function of adult DRG neuron axons using an in vitro microfluidic culture system. Micromachines 2021, 12, 1317. [Google Scholar] [CrossRef]
- Madhusudanan, P.; Jerard, C.; Katiyar, N.; Raju, G.; Shankarappa, S.A. Effect of gold nanoparticle treated dorsal root ganglion cells on peripheral neurite differentiation. Toxicol. Vitr. 2021, 74, 105175. [Google Scholar] [CrossRef] [PubMed]
- Roselló-Busquets, C.; de la Oliva, N.; Martínez-Mármol, R.; Hernaiz-Llorens, M.; Pascual, M.; Muhaisen, A.; Navarro, X.; del Valle, J.; Soriano, E. Cholesterol depletion regulates axonal growth and enhances central and peripheral nerve regeneration. Front. Cell. Neurosci. 2019, 13, 40. [Google Scholar] [CrossRef] [Green Version]
- Yong, Y.; Hughes, C.; Deppmann, C. A microfluidic culture platform to assess axon degeneration. Methods Mol. Biol. 2020, 83–86. [Google Scholar] [CrossRef]
- Osaki, T.; Uzel, S.G.M.; Kamm, R.D. Microphysiological 3D model of amyotrophic lateral sclerosis (ALS) from human IPS-derived muscle cells and optogenetic motor neurons. Sci. Adv. 2018, 4, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Spijkers, X.M.; Pasteuning-Vuhman, S.; Dorleijn, J.C.; Vulto, P.; Wevers, N.R.; Pasterkamp, R.J. A Directional 3D neurite outgrowth model for studying motor axon biology and disease. Sci. Rep. 2021, 11, 2080. [Google Scholar] [CrossRef]
- Vysokov, N.; McMahon, S.B.; Raouf, R. The role of NaV channels in synaptic transmission after axotomy in a microfluidic culture platform. Sci. Rep. 2019, 9, 12915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.N.; Michel, U.; Lenz, C.; Friedel, C.C.; Köster, S.; D’Hedouville, Z.; Tönges, L.; Urlaub, H.; Bähr, M.; Lingor, P.; et al. Calpain-mediated cleavage of collapsin response mediator protein-2 drives acute axonal degeneration. Sci. Rep. 2016, 6, 37050. [Google Scholar] [CrossRef]
- Sala-Jarque, J.; Mesquida-Veny, F.; Badiola-Mateos, M.; Samitier, J.; Hervera, A.; Del Río, J.A. Neuromuscular activity induces paracrine signaling and triggers axonal regrowth after injury in microfluidic lab-on-chip devices. Cells 2020, 9, 302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tecoma, E.S.; Monyer, H.; Goldberg, M.P.; Choi, D.W. Traumatic neuronal injury in vitro is attenuated by NMDA antagonists. Neuron 1989, 2, 1541–1545. [Google Scholar] [CrossRef]
- Payette, D.J.; Xie, J.; Shirwany, N.; Guo, Q. Exacerbation of apoptosis of cortical neurons following traumatic brain injury in Par-4 transgenic mice. Int. J. Clin. Exp. Pathol. 2008, 1, 44–56. [Google Scholar] [PubMed]
- Datar, A.; Ameeramja, J.; Bhat, A.; Srivastava, R.; Mishra, A.; Bernal, R.; Prost, J.; Callan-Jones, A.; Pullarkat, P.A. The roles of microtubules and membrane tension in axonal beading, retraction, and atrophy. Biophys. J. 2019, 117, 880–891. [Google Scholar] [CrossRef]
- Üstün, R.; Oğuz, E.K.; Şeker, A.; Korkaya, H. Thymoquinone protects DRG neurons from axotomy-induced cell death. Neurol. Res. 2018, 40, 930–937. [Google Scholar] [CrossRef]
- Gross, G.W.; Lucas, J.H.; Higgins, M.L. Laser microbeam surgery: Ultrastructural changes associated with neurite transection in culture. J. Neurosci. 1983, 3, 1979–1993. [Google Scholar] [CrossRef] [Green Version]
- Cengiz, N.; Öztürk, G.; Erdoǧan, E.; Him, A.; Oǧuz, E.K. Consequences of neurite transection in vitro. J. Neurotrauma 2012, 29, 2465–2474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunik, D.; Dion, C.; Ozaki, T.; Levin, L.A.; Costantino, S. Laser-based single-axon transection for high-content axon injury and regeneration studies. PLoS ONE 2011, 6, e26832. [Google Scholar] [CrossRef] [Green Version]
- Shao, X.; You, R.; Hui, T.H.; Fang, C.; Gong, Z.; Yan, Z.; Chang, R.C.C.; Shenoy, V.B.; Lin, Y. Tension- and adhesion-regulated retraction of injured axons. Biophys. J. 2019, 117, 193–202. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, M.; Tao, X.; Xu, Z.; Zheng, Y.; Zhu, M.; Zhang, L.; Qiao, J.; Gao, L. Difference of acute dissociation and 1-day culture on the electrophysiological properties of rat dorsal root ganglion neurons. J. Physiol. Biochem. 2018, 74, 207–221. [Google Scholar] [CrossRef]
- Wall, E.J.; Massie, J.B.; Kwan, M.K.; Rydevik, B.L.; Myers, R.R.; Garfin, S.R. experimental stretch neuropathy: Changes in nerve conduction under tension. J. Bone Jt. Surg. Br. Vol. 1992, 74, 126–129. [Google Scholar] [CrossRef] [PubMed]
- Loverde, J.R.; Pfister, B.J. Developmental axon stretch stimulates neuron growth while maintaining normal electrical activity, intracellular calcium flux, and somatic morphology. Front. Cell. Neurosci. 2015, 9, 308. [Google Scholar] [CrossRef] [Green Version]
- Umansky, D.; Midha, R. Commentary: Rapid-stretch injury to peripheral nerves: Histologic results. Neurosurgery 2019, 86, nyz196. [Google Scholar] [CrossRef] [Green Version]
- Mahan, M.A.; Yeoh, S.; Monson, K.; Light, A. Rapid stretch injury to peripheral nerves: Biomechanical results. Neurosurgery 2019, 85, E137–E144. [Google Scholar] [CrossRef] [PubMed]
- Yap, Y.C.; King, A.E.; Guijt, R.M.; Jiang, T.; Blizzard, A.; Breadmore, M.C.; Dickson, T.C. Mild and repetitive very mild axonal stretch injury triggers cystoskeletal mislocalization and growth cone collapse. PLoS ONE 2017, 12, e0176997. [Google Scholar] [CrossRef]
- Zhou, M.; Hu, M.; He, S.; Li, B.; Liu, C.; Min, J.; Hong, L. Effects of RSC96 schwann cell-derived exosomes on proliferation, senescence, and apoptosis of dorsal root ganglion cells in vitro. Med. Sci. Monit. 2018, 24, 7841–7849. [Google Scholar] [CrossRef] [PubMed]
- Aomura, S.; Nakadate, H.; Kaneko, Y.; Nishimura, A.; Willinger, R. Stretch-induced functional disorder of axonal transport in the cultured rat cortex neuron. Integr. Mol. Med. 2016, 3, 654–660. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.M.; Rzigalinski, B.A.; Willoughby, K.A.; Sitterding, H.A.; Ellis, E.F. Stretch-induced injury alters mitochondrial membrane potential and cellular ATP in cultured astrocytes and neurons. J. Neurochem. 2000, 74, 1951–1960. [Google Scholar] [CrossRef] [PubMed]
- Ellis, E.F.; Mckinney, J.S.; Willoughby, K.A.; Liang, S.; Povlishock, J.T. A new model for rapid stretch-induced injury of cells in culture: Characterization of the model using astrocytes. J. Neurotrauma 1995, 12, 325–339. [Google Scholar] [CrossRef]
- Bianchi, F.; Malboubi, M.; George, J.H.; Jerusalem, A.; Thompson, M.S.; Ye, H. Ion current and action potential alterations in peripheral neurons subject to uniaxial strain. J. Neurosci. Res. 2019, 97, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Katanosaka, K.; Takatsu, S.; Mizumura, K.; Naruse, K.; Katanosaka, Y. TRPV2 is required for mechanical nociception and the stretch-evoked response of primary sensory neurons. Sci. Rep. 2018, 8, 16782. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.W.; Cheng, C.M.; LeDuc, P.R.; Chen, C.C. Understanding sensory nerve mechanotransduction through localized elastomeric matrix control. PLoS ONE 2009, 4, e4293. [Google Scholar] [CrossRef]
- Tavalin, S.J.; Ellis, E.F.; Satin, L.S. Mechanical perturbation of cultured cortical neurons reveals a stretch-induced delayed depolarization. J. Neurophysiol. 1995, 74, 2767–2773. [Google Scholar] [CrossRef]
- Chetta, J.; Kye, C.; Shah, S.B. Cytoskeletal dynamics in response to tensile loading of mammalian axons. Cytoskeleton 2010, 67, 650–665. [Google Scholar] [CrossRef]
- Mutalik, S.P.; Joseph, J.; Pullarkat, P.A.; Ghose, A. Cytoskeletal mechanisms of axonal contractility. Biophys. J. 2018, 115, 713–724. [Google Scholar] [CrossRef] [Green Version]
- Loverde, J.R.; Ozoka, V.C.; Aquino, R.; Lin, L.; Pfister, B.J. Live imaging of axon stretch growth in embryonic and adult neurons. J. Neurotrauma 2011, 28, 2389–2403. [Google Scholar] [CrossRef]
- Tsantoulas, C.; Farmer, C.; Machado, P.; Baba, K.; McMahon, S.B.; Raouf, R. Probing functional properties of nociceptive axons using a microfluidic culture system. PLoS ONE 2013, 8, e80722. [Google Scholar] [CrossRef]
- Kampanis, V.; Tolou-Dabbaghian, B.; Zhou, L.; Roth, W.; Puttagunta, R. Cyclic stretch of either PNS or CNS located nerves can stimulate neurite outgrowth. Cells 2020, 10, 32. [Google Scholar] [CrossRef] [PubMed]
- Mahan, M.A. Nerve stretching: A history of tension. J. Neurosurg. 2020, 132, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, K.S.; Struzyna, L.A.; Das, S.; Cullen, D.K. Stretch growth of motor axons in custom mechanobioreactors to generate long-projecting axonal constructs. J. Tissue Eng. Regen. Med. 2019, 13, 2040–2054. [Google Scholar] [CrossRef] [PubMed]
- Gladman, S.J.; Ward, R.E.; Michael-Titus, A.T.; Knight, M.M.; Priestley, J.V. The effect of mechanical strain or hypoxia on cell death in subpopulations of rat dorsal root ganglion neurons in vitro. Neuroscience 2010, 171, 577–587. [Google Scholar] [CrossRef]
- Ji, J.; Tyurina, Y.Y.; Tang, M.; Feng, W.; Stolz, D.B.; Clark, R.S.B.; Meaney, D.F.; Kochanek, P.M.; Kagan, V.E.; Bayír, H. Mitochondrial injury after mechanical stretch of cortical neurons in vitro: Biomarkers of apoptosis and selective peroxidation of anionic phospholipids. J. Neurotrauma 2012, 29, 776–788. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Liu, Y.; Yang, D.; Yuan, F.; Ding, J.; Chen, H.; Tian, H. Sesamin protects SH-SY5Y cells against mechanical stretch injury and promoting cell survival. BMC Neurosci. 2017, 18, 57. [Google Scholar] [CrossRef] [Green Version]
- Mutalik, S.P.; Ghose, A. Axonal cytomechanics in neuronal development. J. Biosci. 2020, 45, 64. [Google Scholar] [CrossRef]
- Faktorovich, S.; Filatov, A.; Rizvi, Z. Common compression neuropathies. Clin. Geriatr. Med. 2021, 37, 241–252. [Google Scholar] [CrossRef]
- Lee, E.Y.; Lim, A.Y.T. Nerve compression in the upper limb. Clin. Plast. Surg. 2019, 46, 285–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menorca, R.M.G.; Fussell, T.S.; Elfar, J.C. Peripheral nerve trauma: Mechanisms of injury and recovery. Hand Clin. 2013, 29, 317–330. [Google Scholar] [CrossRef] [Green Version]
- Tapadia, M.; Mozaffar, T.; Gupta, R. Compressive neuropathies of the upper extremity: Update on pathophysiology, classification, and electrodiagnostic findings. J. Hand Surg. 2010, 35, 668–677. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, O.; Atsuta, Y.; Iwahara, T.; Muramoto, T.; Watakabe, M.; Takemitsu, Y. The effects of mechanical compression and hypoxia on nerve root and dorsal root ganglia: An analysis of ectopic firing using an in vitro model. Spine 1996, 21, 2089–2094. [Google Scholar] [CrossRef]
- Amir, R.; Kocsis, J.D.; Devor, M. Multiple interacting sites of ectopic spike electrogenesis in primary sensory neurons. J. Neurosci. 2005, 25, 2576–2585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.; LaMotte, R.H. Multiple sites for generation of ectopic spontaneous activity in neurons of the chronically compressed dorsal root ganglion. J. Neurosci. 2007, 27, 14059–14068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.; Greenquist, K.W.; LaMotte, R.H. Inflammatory mediators enhance the excitability of chronically compressed dorsal root ganglion neurons. J. Neurophysiol. 2006, 95, 2098–2107. [Google Scholar] [CrossRef] [Green Version]
- Ye, Z.; Wang, Y.; Quan, X.; Li, J.; Hu, X.; Huang, J.; Luo, Z. Effects of mechanical force on cytoskeleton structure and calpain-induced apoptosis in rat dorsal root ganglion neurons in vitro. PLoS ONE 2012, 7, e52183. [Google Scholar] [CrossRef]
- Ewan, E.E.; Avraham, O.; Carlin, D.; Gonçalves, T.M.; Zhao, G.; Cavalli, V. Ascending dorsal column sensory neurons respond to spinal cord injury and downregulate genes related to lipid metabolism. Sci. Rep. 2021, 11, 374. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.; Reid, A.; Verkhratsky, A.; Magnaghi, V.; Faroni, A. Gene Expression changes in dorsal root ganglia following peripheral nerve injury: Roles in inflammation, cell death and nociception. Neural Regen. Res. 2019, 14, 939–947. [Google Scholar] [CrossRef]
- 1Wang, T.; Hurwitz, O.; Shimada, S.G.; Qu, L.; Fu, K.; Zhang, P.; Ma, C.; LaMotte, R.H. Chronic compression of the dorsal root ganglion enhances mechanically evoked pain behavior and the activity of cutaneous nociceptors in mice. PLoS ONE 2015, 10, e0137512. [Google Scholar] [CrossRef]
- Zhao, J.Y.; Liang, L.; Gu, X.; Li, Z.; Wu, S.; Sun, L.; Atianjoh, F.E.; Feng, J.; Mo, K.; Jia, S.; et al. DNA methyltransferase DNMT3a contributes to neuropathic pain by repressing kcna2 in primary afferent neurons. Nat. Commun. 2017, 8, 14712. [Google Scholar] [CrossRef]
- Myers, K.A.; Rattner, J.B.; Shrive, N.G.; Hart, D.A.; Myers, K.A.; Rattner, J.B.; Shrive, N.G.; Hart, D.A. Hydrostatic pressure sensation in cells: Integration into the tensegrity model. Biochem. Cell Biol. 2007, 85, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Frieboes, L.R.; Gupta, R. An in-vitro traumatic model to evaluate the response of myelinated cultures to sustained hydrostatic compression injury. J. Neurotrauma 2009, 26, 2245–2256. [Google Scholar] [CrossRef] [Green Version]
- Quan, X.; Guo, K.; Wang, Y.; Huang, L.; Chen, B.; Ye, Z.; Luo, Z. Mechanical compression insults induce nanoscale changes of membrane-skeleton arrangement which could cause apoptosis and necrosis in dorsal root ganglion neurons. Biosci. Biotechnol. Biochem. 2014, 78, 1631–1639. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Tao, R.; Wang, M.; Tian, J.; Genin, G.M.; Lu, T.J.; Xu, F. Regulation of cell behavior by hydrostatic pressure. Appl. Mech. Rev. 2019, 71, 40803. [Google Scholar] [CrossRef]
- Ask, P.; Levitan, H.; Robinson, P.J.; Rapoport, S.I. Peripheral nerve as an osmometer: Role of endoneurial capillaries in frog sciatic nerve. Am. J. Physiol. Cell Physiol. 1983, 244, C75–C81. [Google Scholar] [CrossRef]
- Lim, T.K.Y.; Shi, X.Q.; Johnson, J.M.; Rone, M.B.; Antel, J.P.; David, S.; Zhang, J. Peripheral nerve injury induces persistent vascular dysfunction and endoneurial hypoxia, contributing to the genesis of neuropathic pain. J. Neurosci. 2015, 35, 3346–3359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizisin, A.P.; Weerasuriya, A. Homeostatic Regulation of the endoneurial microenvironment during development, aging and in response to trauma, disease and toxic insult. Acta Neuropathol. 2011, 121, 291–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, R.R.; Rydevik, B.L.; Heckman, H.M.; Powell, H.C. Proximodistal gradient in endoneurial fluid pressure. Exp. Neurol. 1988, 102, 368–370. [Google Scholar] [CrossRef]
- Sinar, E.J.; Mendelow, A.D.; Graham, D.I.; Teasdale, G.M. Experimental intracerebral hemorrhage: Effects of a temporary mass lesion. J. Neurosurg. 1987, 66, 568–576. [Google Scholar] [CrossRef]
- Tök, L.; Naziroʇlu, M.; Uʇuz, A.C.; Tök, Ö. Elevated hydrostatic pressures induce apoptosis and oxidative stress through mitochondrial membrane depolarization in PC12 neuronal cells: A cell culture model of glaucoma. J. Recept. Signal Transduct. 2014, 34, 410–416. [Google Scholar] [CrossRef]
- Lei, Y.; Rajabi, S.; Pedrigi, R.M.; Overby, D.R.; Thomas Read, A.; Ross Ethier, C. In vitro models for glaucoma research: Effects of hydrostatic pressure. Investig. Ophthalmol. Vis. Sci. 2011, 52, 6329–6339. [Google Scholar] [CrossRef]
- Mandal, A.; Shahidullah, M.; Delamere, N.A. Hydrostatic pressure-induced release of stored calcium in cultured rat optic nerve head astrocytes. Investig. Ophthalmol. Vis. Sci. 2010, 51, 3129–3138. [Google Scholar] [CrossRef] [Green Version]
- Guo, T.; Bian, Z.; Trocki, K.; Chen, L.; Zheng, G.; Feng, B. Optical recording reveals topological distribution of functionally classified colorectal afferent neurons in intact lumbosacral DRG. Physiol. Rep. 2019, 7, e14097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahlberg, B.; Ghuman, H.; Liu, J.R.; Modo, M. Ex vivo biomechanical characterization of syringe-needle ejections for intracerebral cell delivery. Sci. Rep. 2018, 8, 9194. [Google Scholar] [CrossRef] [PubMed]
- Pastores, G.M. Neuropathic gaucher disease. Wien. Med. Wochenschr. 2010, 160, 605–608. [Google Scholar] [CrossRef] [PubMed]
- Feldman, E.L.; Callaghan, B.C.; Pop-Busui, R.; Zochodne, D.W.; Wright, D.E.; Bennett, D.L.; Bril, V.; Russell, J.W.; Viswanathan, V. Diabetic neuropathy. Nat. Rev. Dis. Primers 2019, 5, 41. [Google Scholar] [CrossRef]
- Yang, H.; Sloan, G.; Ye, Y.; Wang, S.; Duan, B.; Tesfaye, S.; Gao, L. New perspective in diabetic neuropathy: From the periphery to the brain, a call for early detection, and precision medicine. Front. Endocrinol. 2020, 10, 929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagihashi, S.; Mizukami, H.; Sugimoto, K. Mechanism of diabetic neuropathy: Where are we now and where to go? J. Diabetes Investig. 2011, 2, 18–32. [Google Scholar] [CrossRef] [Green Version]
- Koshimura, K.; Tanaka, J.; Murakami, Y.; Kato, Y. Effect of high concentration of glucose on dopamine release from pheochromocytoma-12 cells. Metabolism 2003, 52, 922–926. [Google Scholar] [CrossRef]
- Koshimura, K.; Tanaka, J.; Murakami, Y.; Kato, Y. Involvement of nitric oxide in glucose toxicity on differentiated pc12 cells: Prevention of glucose toxicity by tetrahydrobiopterin, a cofactor for nitric oxide synthase. Neurosci. Res. 2002, 43, 31–38. [Google Scholar] [CrossRef]
- Shi, Y.; Liang, X.; Zhang, H.; Sun, Q.; Wu, Q.; Qu, L. Combination of quercetin, cinnamaldehyde and hirudin protects rat dorsal root ganglion neurons against high glucose-induced injury through Nrf-2/HO-1 activation and NF-ΚB inhibition. Chin. J. Integr. Med. 2017, 23, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, S.; Xue, J.; Wei, Z.; Ao, P.; Shen, B.; Ding, L. CGRP reduces apoptosis of DRG cells induced by high-glucose oxidative stress injury through PI3K/AKT induction of heme oxygenase-1 and Nrf-2 expression. Oxidative Med. Cell. Longev. 2019, 2019, 1–9. [Google Scholar] [CrossRef]
- Peeraer, E.; van Lutsenborg, A.; Verheyen, A.; de Jongh, R.; Nuydens, R.; Meert, T.F. Pharmacological evaluation of rat dorsal root ganglion neurons as an in vitro model for diabetic neuropathy. J. Pain Res. 2011, 4, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Zherebitskaya, E.; Akude, E.; Smith, D.R.; Fernyhough, P. Development of selective axonopathy in adult sensory neurons isolated from diabetic rats: Role of glucose-induced oxidative stress. Diabetes 2009, 58, 1356–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zherebitskaya, E.; Schapansky, J.; Akude, E.; Smith, D.R.; van der Ploeg, R.; Solovyova, N.; Verkhratsky, A.; Fernyhough, P. Sensory neurons derived from diabetic rats have diminished internal Ca2+ stores linked to impaired re-uptake by the endoplasmic reticulum. ASN Neuro. 2012, 4, e00072. [Google Scholar] [CrossRef] [PubMed]
- Sango, K.; Saito, H.; Takano, M.; Tokashiki, A.; Inoue, S.; Horie, H. Cultured adult animal neurons and schwann cells give us new insights into diabetic neuropathy. Curr. Diabetes Rev. 2006, 2, 169–183. [Google Scholar] [CrossRef]
- Jones, M.R.; Urits, I.; Wolf, J.; Corrigan, D.; Colburn, L.; Peterson, E.; Williamson, A.; Viswanath, O. Drug-induced peripheral neuropathy: A narrative review. Curr. Clin. Pharmacol. 2020, 15, 38–48. [Google Scholar] [CrossRef]
- Popescu, C. Severe acute axonal neuropathy induced by ciprofloxacin: A case report. Case Rep. Neurol. 2018, 10, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Shankarappa, S.A.; Sagie, I.; Tsui, J.H.; Chiang, H.H.; Stefanescu, C.; Zurakowski, D.; Kohane, D.S. Duration and local toxicity of sciatic nerve blockade with coinjected site 1 sodium-channel blockers and quaternary lidocaine derivatives. Reg. Anesth. Pain Med. 2012, 37, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Verlinde, M.; Hollmann, M.W.; Stevens, M.F.; Hermanns, H.; Werdehausen, R.; Lirk, P. Local anesthetic-induced neurotoxicity. Int. J. Mol. Sci. 2016, 17, 339. [Google Scholar] [CrossRef]
- Hewson, D.W.; Bedforth, N.M.; Hardman, J.G. Peripheral nerve injury arising in anaesthesia practice. Anaesthesia 2018, 73, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Wen, X.; Li, Y.; Liu, X.; Sun, C.; Lin, J.; Zhang, W.; Wu, Y.; Wang, X. Roles of CaMKIIβ in the neurotoxicity induced by ropivacaine hydrochloride in dorsal root ganglion. Artif. Cells Nanomed. Biotechnol. 2019, 47, 2948–2956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staff, N.P.; Fehrenbacher, J.C.; Caillaud, M.; Damaj, M.I.; Segal, R.A.; Rieger, S. Pathogenesis of paclitaxel-induced peripheral neuropathy: A current review of in vitro and in vivo findings using rodent and human model systems. Exp. Neurol. 2020, 324, 113121. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, M.; Gorgun, M.F.; Englander, E.W. Translesion SYnthesis DNA polymerase kappa is indispensable for DNA repair synthesis in cisplatin Exposed dorsal root ganglion neurons. Mol. Neurobiol. 2018, 55, 2506–2515. [Google Scholar] [CrossRef] [PubMed]
- Triarico, S.; Romano, A.; Attinà, G.; Capozza, M.A.; Maurizi, P.; Mastrangelo, S.; Ruggiero, A. Vincristine-induced peripheral neuropathy (VIPN) in pediatric tumors: Mechanisms, risk factors, strategies of prevention and treatment. Int. J. Mol. Sci. 2021, 22, 4112. [Google Scholar] [CrossRef]
- Baldewig, M.; Goldbaum, O.; Richter-Landsberg, C.; Weyland, A.; Bantel, C. Short-term incubation of gabapentin or pregabalin does not affect chemically induced injury in neuronal cell models in vitro. JPR 2018, 11, 1181–1190. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Hamre, J.; Eldridge, S.; Behrsing, H.P.; Cutuli, F.M.; Mussio, J.; Davis, M. Multiparametric image analysis of rat dorsal root ganglion cultures to evaluate peripheral neuropathy- inducing chemotherapeutics. Toxicol. Sci. 2017, 156, 275–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eldridge, S.; Scuteri, A.; Jones, E.M.C.; Cavaletti, G.; Guo, L.; Glaze, E. Considerations for a reliable in vitro model of chemotherapy-induced peripheral neuropathy. Toxics 2021, 9, 300. [Google Scholar] [CrossRef]
- Eldridge, S.; Guo, L.; Hamre, J. A comparative review of chemotherapy-induced peripheral neuropathy in in vivo and in vitro models. Toxicol. Pathol. 2020, 48, 190–201. [Google Scholar] [CrossRef]
- Silva, A.; Wang, Q.; Wang, M.; Ravula, S.K.; Glass, J.D. Evidence for direct axonal toxicity in vincristine neuropathy. J. Peripher. Nerv. Syst. 2006, 11, 211–216. [Google Scholar] [CrossRef]
- Yang, I.H.; Siddique, R.; Hosmane, S.; Thakor, N.; Höke, A. Compartmentalized microfluidic culture platform to study mechanism of paclitaxel-induced axonal degeneration. Exp. Neurol. 2009, 218, 124–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geisler, S.; Doan, R.A.; Cheng, G.C.; Cetinkaya-Fisgin, A.; Huang, S.X.; Höke, A.; Milbrandt, J.; DiAntonio, A. Vincristine and bortezomib use distinct upstream mechanisms to activate a common SARM1-dependent axon degeneration program. JCI Insight 2019, 4, e129920. [Google Scholar] [CrossRef]
- Imai, S.; Koyanagi, M.; Azimi, Z.; Nakazato, Y.; Matsumoto, M.; Ogihara, T.; Yonezawa, A.; Omura, T.; Nakagawa, S.; Wakatsuki, S.; et al. Taxanes and platinum derivatives impair schwann cells via distinct mechanisms. Sci. Rep. 2017, 7, 5947. [Google Scholar] [CrossRef] [Green Version]
- Werdehausen, R.; Braun, S.; Essmann, F.; Schulze-Osthoff, K.; Walczak, H.; Lipfert, P.; Stevens, M.F. Lidocaine induces apoptosis via the mitochondrial pathway independently of death receptor signaling. Anesthesiology 2007, 107, 136–143. [Google Scholar] [CrossRef]
- El-Boghdadly, K.; Pawa, A.; Chin, K.J. Local anesthetic systemic toxicity: Current perspectives. Local Reg. Anesth. 2018, 11, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermanns, H.; Hollmann, M.W.; Stevens, M.F.; Lirk, P.; Brandenburger, T.; Piegeler, T.; Werdehausen, R. Molecular mechanisms of action of systemic lidocaine in acute and chronic pain: A narrative review. Br. J. Anaesth. 2019, 123, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.-J.; Kim, H.Y.; Ahn, J.-H. Neurotoxicity of local anesthetics in dentistry. J. Dent. Anesth. Pain Med. 2020, 20, 55. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, K. Downregulation of microRNA-33-5p protected bupivacaine-induced apoptosis in murine dorsal root ganglion neurons through GDNF. Neurotox. Res. 2019, 35, 860–866. [Google Scholar] [CrossRef]
- Guo, Z.; Liu, Y.; Cheng, M. Resveratrol Protects bupivacaine-induced neuro-apoptosis in dorsal root ganglion neurons via activation on tropomyosin receptor kinase A. Biomed. Pharmacother. 2018, 103, 1545–1551. [Google Scholar] [CrossRef]
- Brull, R.; Hadzic, A.; Reina, M.A.; Barrington, M.J. Pathophysiology and etiology of nerve injury following peripheral nerve blockade. Reg. Anesth. Pain Med. 2015, 40, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Yang, S.; Luo, C. TNF-alpha and TNF-R1 regulate bupivacaine-induced apoptosis in spinal cord dorsal root ganglion neuron. Eur. J. Pharmacol. 2018, 833, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.E.; Garner, J. Pathophysiology of primary blast injury. J. R. Army Med. Corps 2019, 165, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Antonine Jerusalem, M.D. Continuum modelling of neuronal cell under blast loading. Acta Biomater. 2012, 8, 3360–3371. [Google Scholar] [CrossRef] [Green Version]
- Cernak, I. Understanding blast-induced neurotrauma: How far have we come? Concussion 2017, 2, CNC42. [Google Scholar] [CrossRef] [Green Version]
- Hicks, R.R.; Fertig, S.J.; Desrocher, R.E.; Koroshetz, W.J.; Pancrazio, J.J. Neurological effects of blast injury. J. Trauma Inj. Infect. Crit. Care 2010, 68, 1257–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suneson, A.; Hansson, H.A.; Lycke, E.; Seeman, T. Pressure wave injuries to rat dorsal root ganglion cells in culture caused by high-energy missiles. J. Trauma Inj. Infect. Crit. Care 1989, 29, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Zander, N.E.; Piehler, T.; Boggs, M.E.; Banton, R.; Benjamin, R. In vitro studies of primary explosive blast loading on neurons. J. Neurosci. Res. 2015, 93, 1353–1363. [Google Scholar] [CrossRef]
- Chen, P.; Piao, X.; Bonaldo, P. Role of macrophages in wallerian degeneration and axonal regeneration after peripheral nerve injury. Acta Neuropathol. 2015, 130, 605–618. [Google Scholar] [CrossRef]
- Madhusudanan, P.; Reade, S.; Shankarappa, S.A. Neuroglia as targets for drug delivery systems: A review. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 667–679. [Google Scholar] [CrossRef]
- Sanders, V.M.; Jones, K.J. Role of immunity in recovery from a peripheral nerve injury. J. Neuroimmune Pharmacol. 2006, 1, 11–19. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varier, P.; Raju, G.; Madhusudanan, P.; Jerard, C.; Shankarappa, S.A. A Brief Review of In Vitro Models for Injury and Regeneration in the Peripheral Nervous System. Int. J. Mol. Sci. 2022, 23, 816. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23020816
Varier P, Raju G, Madhusudanan P, Jerard C, Shankarappa SA. A Brief Review of In Vitro Models for Injury and Regeneration in the Peripheral Nervous System. International Journal of Molecular Sciences. 2022; 23(2):816. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23020816
Chicago/Turabian StyleVarier, Parvathi, Gayathri Raju, Pallavi Madhusudanan, Chinnu Jerard, and Sahadev A. Shankarappa. 2022. "A Brief Review of In Vitro Models for Injury and Regeneration in the Peripheral Nervous System" International Journal of Molecular Sciences 23, no. 2: 816. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23020816