
Kidney Injury Causes Accumulation of Renal Sodium That Modulates Renal Lymphatic Dynamics

 , , ,
, , ,  , and
, and 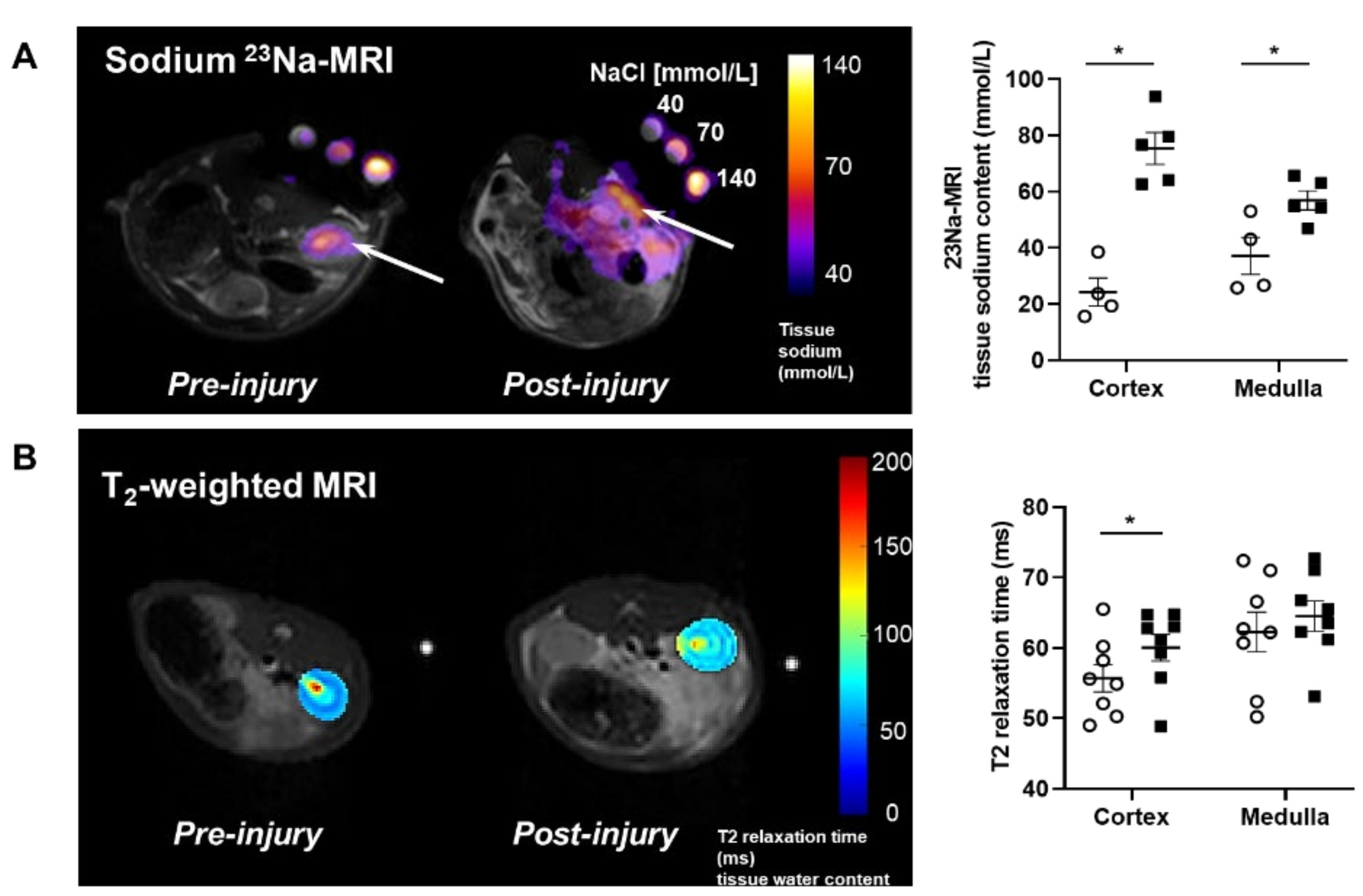{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
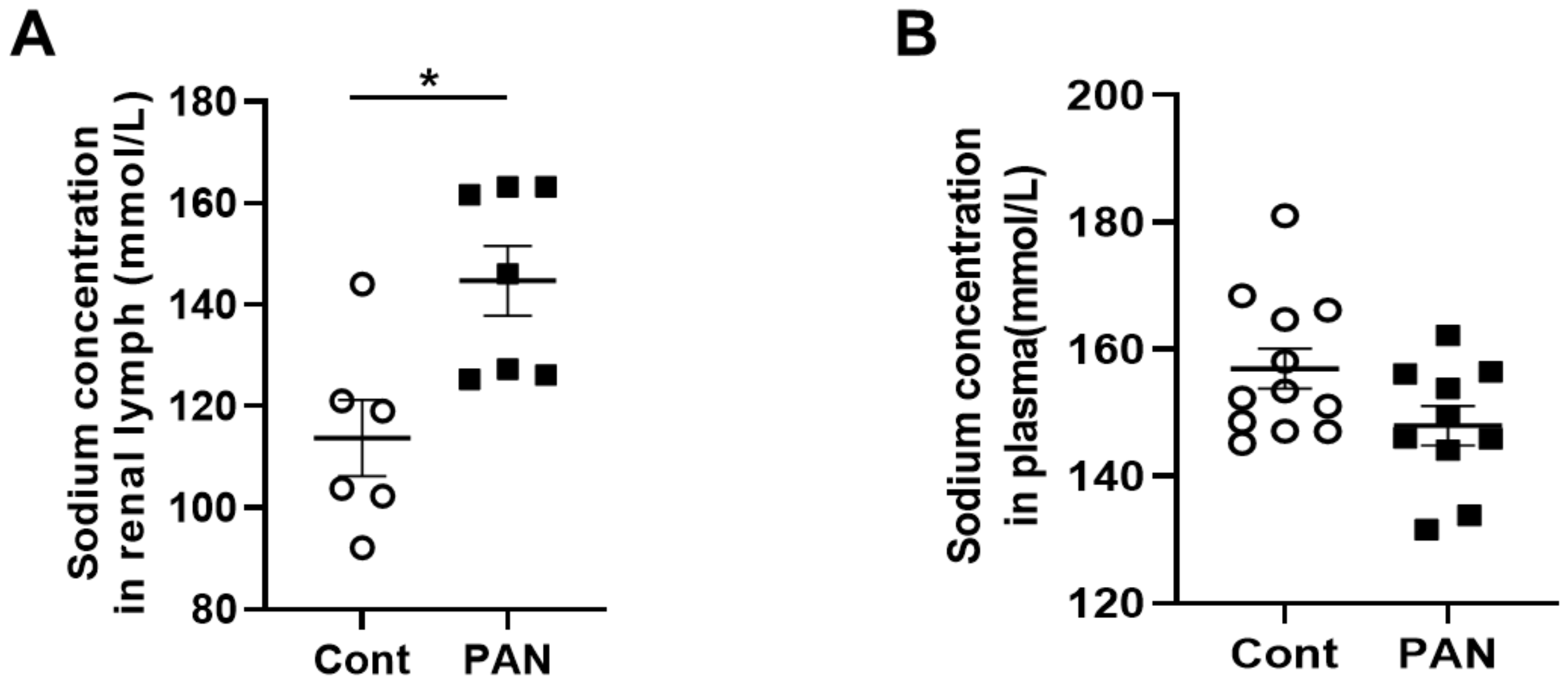
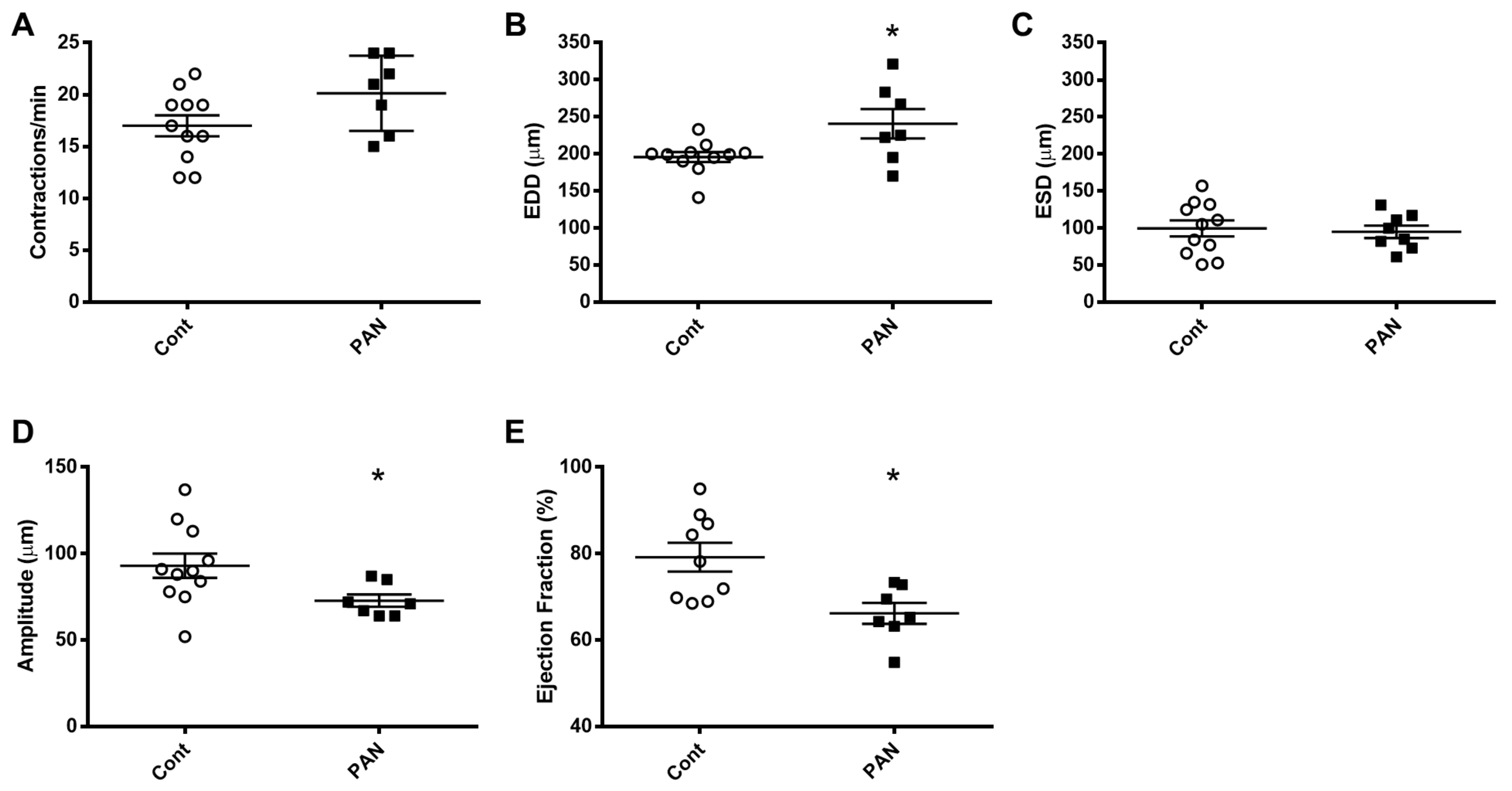
2.1. Proteinuric Kidney Injury Increased Renal Sodium
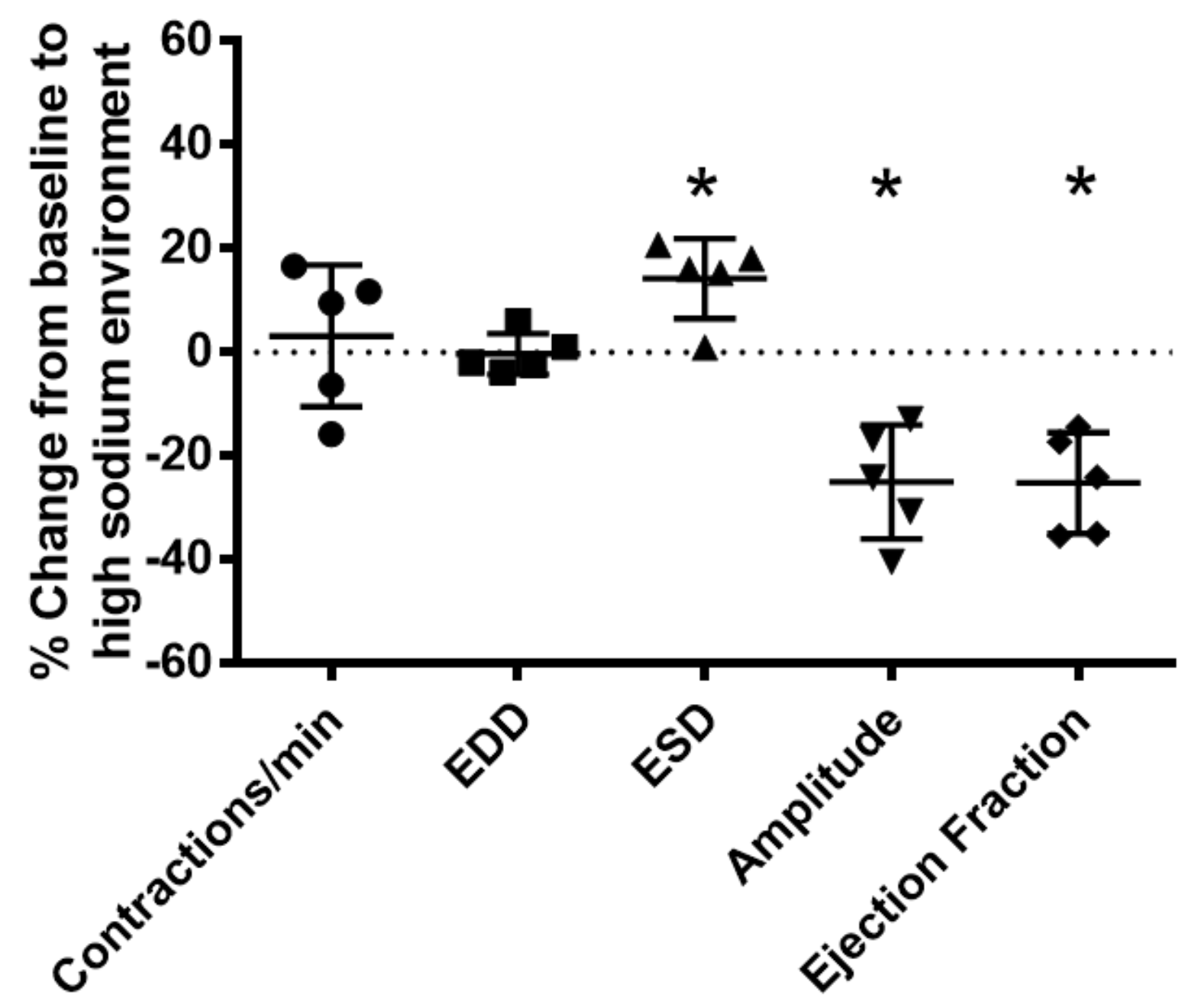
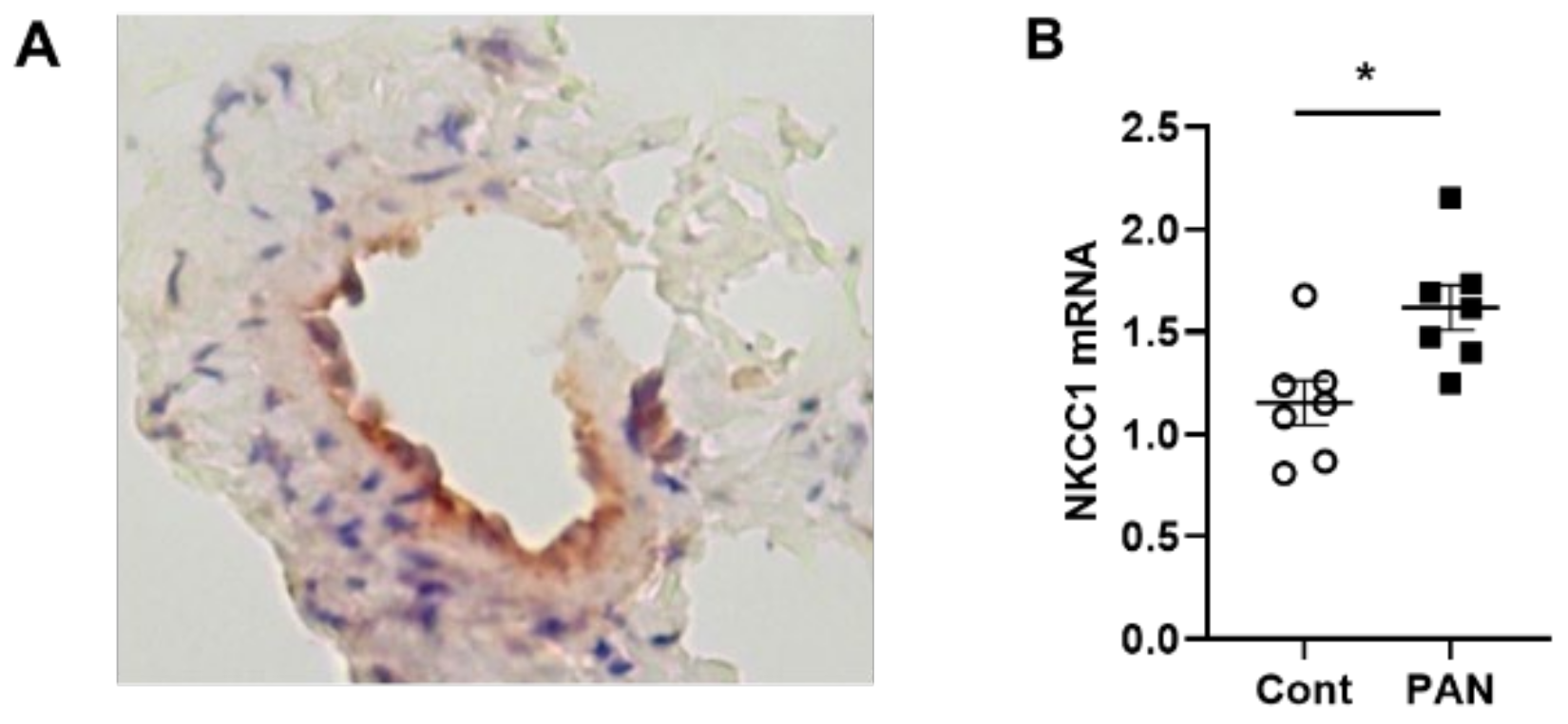
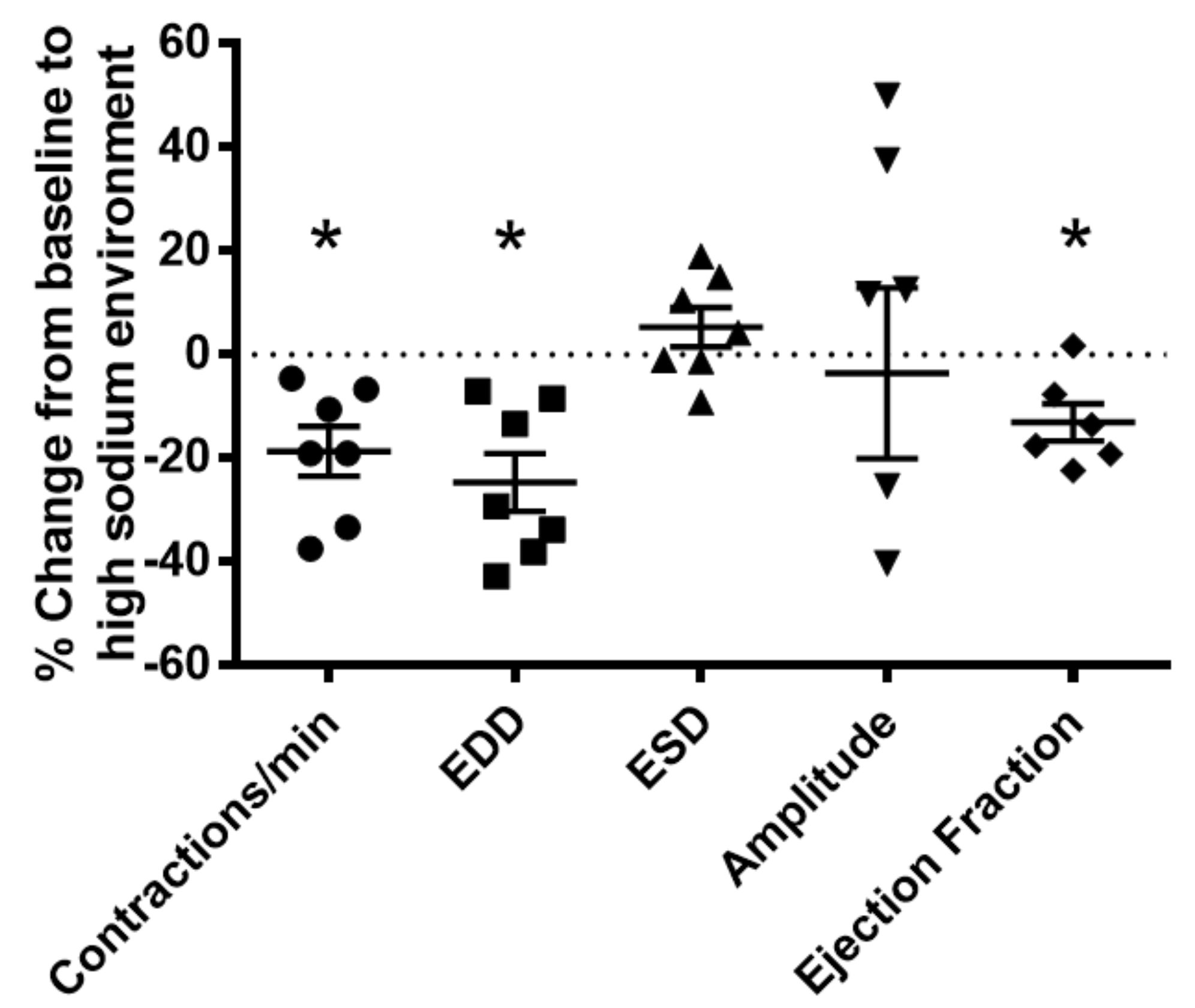
2.2. High Sodium Environment Changed Contractility of Renal Lymphatic Vessels Involving the NKCC1 Transporter
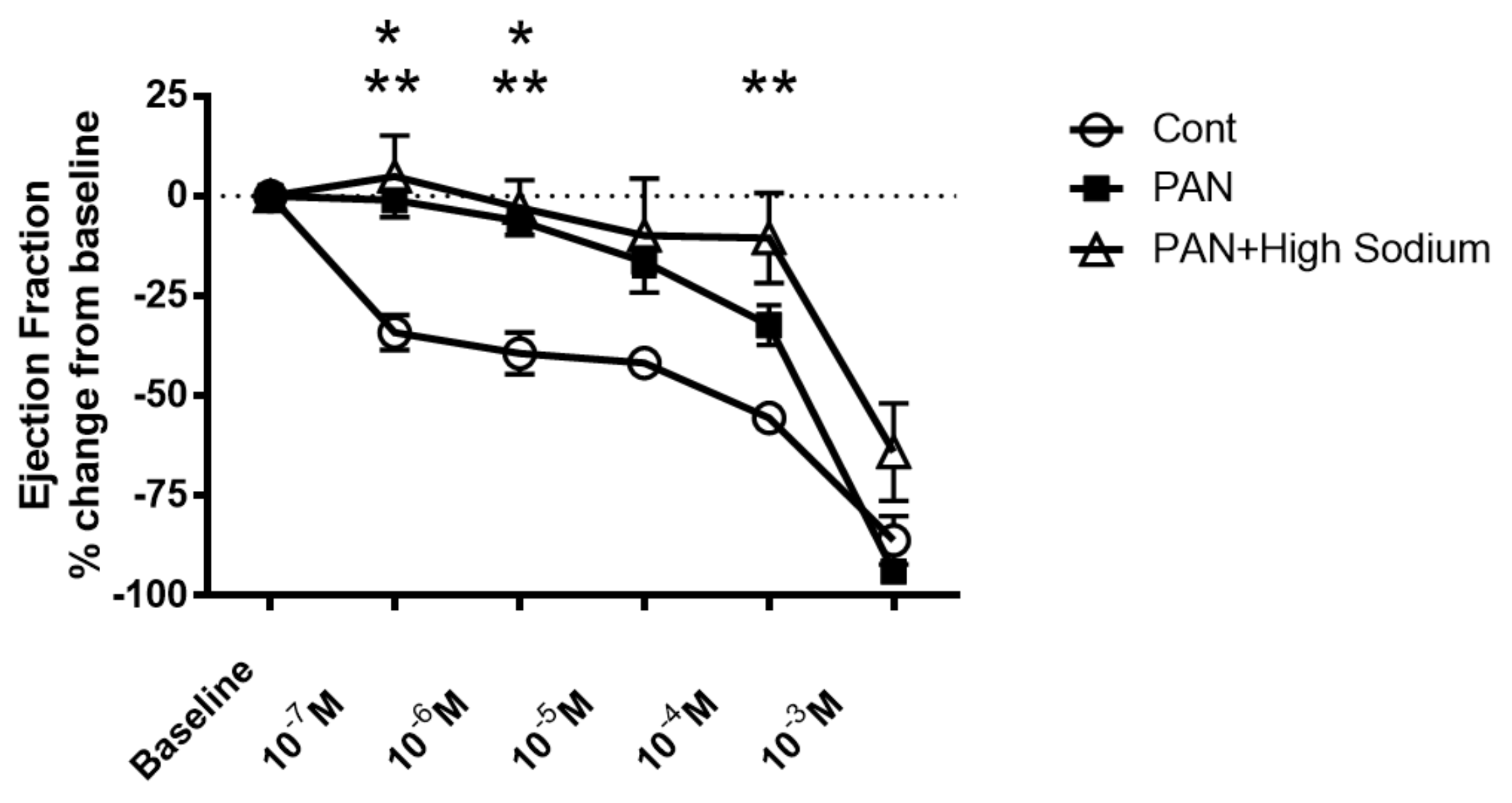
2.3. Kidney Injury Diminished the Lymphatic Vascular Response to a High-Sodium Environment and NKCC Inhibition by Furosemide
3. Discussion
4. Materials and Methods
4.1. Animal Experiments
4.2. Magnetic Resonance Imaging Acquisition
Image Analysis
4.3. Serum and Lymph Sodium Analysis
4.4. Measurement of Lymphatic Vessels Contractility
Buffers
4.5. Reverse Transcription Quantitative Real-Time PCR
4.6. Immunohistochemical Staining
4.7. Cell Culture
4.8. Western Blot
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ellison, D.H.; Welling, P. Insights into Salt Handling and Blood Pressure. N. Engl. J. Med. 2021, 385, 1981–1993. [Google Scholar] [CrossRef] [PubMed]
- Mullins, L.; Ivy, J.; Ward, M.; Tenstad, O.; Wiig, H.; Kitada, K.; Manning, J.; Rakova, N.; Muller, D.; Mullins, J. Abnormal Neonatal Sodium Handling in Skin Precedes Hypertension in the SAME Rat. Pflugers Arch. 2021, 473, 897–910. [Google Scholar] [CrossRef] [PubMed]
- Rabelink, T.J.; Rotmans, J.I. Salt Is Getting Under Our Skin. Nephrol. Dial. Transplant. 2009, 24, 3282–3283. [Google Scholar] [CrossRef] [PubMed]
- Wiig, H.; Schröder, A.; Neuhofer, W.; Jantsch, J.; Kopp, C.; Karlsen, T.V.; Boschmann, M.; Goss, J.; Bry, M.; Rakova, N.; et al. Immune Cells Control Skin Lymphatic Electrolyte Homeostasis and Blood Pressure. J. Clin. Investig. 2013, 123, 2803–2815. [Google Scholar] [CrossRef] [PubMed]
- Matthay, M.A.; Zemans, R.L.; Zimmerman, G.A.; Arabi, Y.M.; Beitler, J.R.; Mercat, A.; Herridge, M.; Randolph, A.G.; Calfee, C.S. Acute Respiratory Distress Syndrome. Nat. Rev. Dis. Primers 2019, 5, 18. [Google Scholar] [CrossRef]
- Rossitto, G.; Mary, S.; Chen, J.Y.; Boder, P.; Chew, K.S.; Neves, K.B.; Alves, R.L.; Montezano, A.C.; Welsh, P.; Petrie, M.C.; et al. Tissue Sodium Excess Is Not Hypertonic and Reflects Extracellular Volume Expansion. Nat. Commun. 2020, 11, 4222. [Google Scholar] [CrossRef]
- Abouelkheir, G.R.; Upchurch, B.D.; Rutkowski, J.M. Lymphangiogenesis: Fuel, Smoke, or Extinguisher of Inflammation’s Fire? Exp. Biol. Med. (Maywood) 2017, 242, 884–895. [Google Scholar] [CrossRef]
- Donnan, M.D.; Kenig-Kozlovsky, Y.; Quaggin, S.E. The Lymphatics in Kidney Health and Disease. Nat. Rev. Nephrol. 2021, 17, 655–675. [Google Scholar] [CrossRef]
- Hasegawa, S.; Nakano, T.; Torisu, K.; Tsuchimoto, A.; Eriguchi, M.; Haruyama, N.; Masutani, K.; Tsuruya, K.; Kitazono, T. Vascular Endothelial Growth Factor-C Ameliorates Renal Interstitial Fibrosis through Lymphangiogenesis in Mouse Unilateral Ureteral Obstruction. Lab. Investig. 2017, 97, 1439–1452. [Google Scholar] [CrossRef]
- Hwang, S.D.; Song, J.H.; Kim, Y.; Lim, J.H.; Kim, M.Y.; Kim, E.N.; Hong, Y.A.; Chung, S.; Choi, B.S.; Kim, Y.S.; et al. Inhibition of Lymphatic Proliferation by the Selective VEGFR-3 Inhibitor SAR131675 Ameliorates Diabetic Nephropathy in db/db Mice. Cell Death Dis. 2019, 10, 219. [Google Scholar] [CrossRef]
- Kerjaschki, D.; Huttary, N.; Raab, I.; Regele, H.; Bojarski-Nagy, K.; Bartel, G.; Kröber, S.M.; Greinix, H.; Rosenmaier, A.; Karlhofer, F.; et al. Lymphatic Endothelial Progenitor Cells Contribute to De Novo Lymphangiogenesis in Human Renal Transplants. Nat. Med. 2006, 12, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Pei, G.; Yao, Y.; Yang, Q.; Wang, M.; Wang, Y.; Wu, J.; Wang, P.; Li, Y.; Zhu, F.; Yang, J.; et al. Lymphangiogenesis in Kidney and Lymph Node Mediates Renal Inflammation and Fibrosis. Sci. Adv. 2019, 5, eaaw5075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazdani, S.; Poosti, F.; Kramer, A.B.; Mirković, K.; Kwakernaak, A.J.; Hovingh, M.; Slagman, M.C.; Sjollema, K.A.; de Borst, M.H.; Navis, G.; et al. Proteinuria Triggers Renal Lymphangiogenesis Prior to the Development of Interstitial Fibrosis. PLoS ONE 2012, 7, e50209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarjou, A.; Black, L.M.; Bolisetty, S.; Traylor, A.M.; Bowhay, S.A.; Zhang, M.Z.; Harris, R.C.; Agarwal, A. Dynamic Signature of Lymphangiogenesis During Acute Kidney Injury and Chronic Kidney Disease. Lab. Investig. 2019, 99, 1376–1388. [Google Scholar] [CrossRef]
- Solari, E.; Marcozzi, C.; Negrini, D.; Moriondo, A. Lymphatic Vessels and Their Surroundings: How Local Physical Factors Affect Lymph Flow. Biology (Basel) 2020, 9, 463. [Google Scholar] [CrossRef] [PubMed]
- Scallan, J.P.; Bouta, J.P.; Rahimi, H.; Kenney, H.M.; Ritchlin, C.T.; Davis, C.T.; Schwarz, E.M. Ex vivo Demonstration of Functional Deficiencies in Popliteal Lymphatic Vessels From TNF-Transgenic Mice With Inflammatory Arthritis. Front. Physiol 2021, 12, 745096. [Google Scholar] [CrossRef]
- Shelton, E.L.; Yang, H.C.; Zhong, J.; Salzman, M.M.; Kon, V. Renal Lymphatic Vessel Dynamics. Am. J. Physiol. Renal. Physiol. 2020, 319, F1027–F1036. [Google Scholar] [CrossRef]
- Dormans, T.P.; Pickkers, P.; Russel, F.G.; Smits, P. Vascular Effects of Loop Diuretics. Cardiovasc. Res. 1996, 32, 988–997. [Google Scholar] [CrossRef]
- Karlsen, T.V.; Nikpey, E.; Han, J.; Reikvam, T.; Rakova, N.; Castorena-Gonzalez, J.A.; Davis, M.J.; Titze, J.M.; Tenstad, O.; Wiig, H. High-Salt Diet Causes Expansion of the Lymphatic Network and Increased Lymph Flow in Skin and Muscle of Rats. Arter. Thromb. Vasc. Biol. 2018, 38, 2054–2064. [Google Scholar] [CrossRef] [Green Version]
- Balasubbramanian, D.; Baranwal, G.; Clark, M.C.; Goodlett, B.L.; Mitchell, B.M.; Rutkowski, J.M. Kidney-specific lymphangiogenesis increases sodium excretion and lowers blood pressure in mice. J. Hypertens. 2020, 38, 874–885. [Google Scholar] [CrossRef]
- Delpire, E.; Gagnon, K.B. Na(+)-K(+)-2cl(−) Cotransporter (Nkcc) Physiological Function in Nonpolarized Cells and Transporting Epithelia. Compr. Physiol. 2018, 8, 871–901. [Google Scholar] [CrossRef]
- Zeniya, M.; Sohara, E.; Kita, S.; Iwamoto, T.; Susa, K.; Mori, T.; Oi, K.; Chiga, M.; Takahashi, D.; Yang, S.S.; et al. Dietary Salt Intake Regulates WNK3-SPAK-NKCC1 Phosphorylation Cascade in Mouse Aorta through Angiotensin II. Hypertension 2013, 62, 872–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldwin, S.N.; Sandow, S.L.; Mondéjar-Parreño, G.; Stott, J.B.; Greenwood, I.A. K(V)7 Channel Expression and Function within Rat Mesenteric Endothelial Cells. Front. Physiol. 2020, 11, 598779. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wu, X.; Mao, Y.; Liu, C.; Wu, Y.; Tang, J.; Zhao, K.; Li, P. Nitric Oxide Alleviated High Salt-Induced Cardiomyocyte Apoptosis and Autophagy Independent of Blood Pressure in Rats. Front. Cell Dev. Biol. 2021, 9, 646575. [Google Scholar] [CrossRef] [PubMed]
- Madelin, G.; Lee, J.S.; Regatte, R.R.; Jerschow, A. Sodium MRI: Methods and Applications. Prog. Nucl. Magn. Reson. Spectrosc. 2014, 79, 14–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katz, Y.J.; Cockett, A.; Moor, R.S. Elevation of Inferior Vena Cava Pressure and Thoracic Lymph and Urine Flow. Circ. Res. 1959, 7, 118–122. [Google Scholar] [CrossRef] [Green Version]
- Lebrie, S.J.; Mayerson, H.S. Influence of Elevated Venous Pressure on Flow and Composition of Renal Lymph. Am. J. Physiol. 1960, 198, 1037–1040. [Google Scholar] [CrossRef]
- Baranwal, G.; Creed, H.A.; Black, L.M.; Auger, A.; Quach, A.M.; Vegiraju, R.; Eckenrode, H.E.; Agarwal, A.; Rutkowski, J.M. Expanded Renal Lymphatics Improve Recovery Following Kidney Injury. Physiol Rep. 2021, 9, e15094. [Google Scholar] [CrossRef]
- Zhao, X.; Lai, G.; Tu, J.; Liu, S.; Zhao, Y. Crosstalk between Phosphorylation and Ubiquitination Is Involved in High Salt-Induced WNK4 Expression. Exp. Ther. Med. 2021, 21, 133. [Google Scholar] [CrossRef]
- Akar, F.; Jiang, G.; Paul, R.J.; O’Neill, W.C. Contractile Regulation of the Na(+)-K(+)-2cl(−) Cotransporter in Vascular Smooth Muscle. Am. J. Physiol. Cell Physiol. 2001, 281, C579–C584. [Google Scholar] [CrossRef]
- Akar, F.; Skinner, E.; Klein, J.D.; Jena, M.; Paul, R.J.; O’Neill, W.C. Vasoconstrictors and Nitrovasodilators Reciprocally Regulate the Na+-K+-2cl- Cotransporter in Rat Aorta. Am. J. Physiol. 1999, 276, C1383–C1390. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.; Agollah, G.D.; Sevick-Muraca, E.M.; Chan, W. Altered Lymphatic Function and Architecture in Salt-Induced Hypertension Assessed by Near-Infrared Fluorescence Imaging. J. Biomed. Opt. 2012, 17, 080504. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, R.; Isshiki, M.; Ono, N.; Nishimoto, M.; Fujita, T. A High-Salt Diet Differentially Modulates Mechanical Activity of Afferent and Efferent Collecting Lymphatics in Murine Iliac Lymph Nodes. Lymphat. Res. Biol. 2015, 13, 85–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hueper, K.; Gutberlet, M.; Brasen, J.H.; Jang, M.S.; Thorenz, A.; Chen, R.; Hertel, B.; Barrmeyer, A.; Schmidbauer, M.; Meier, M.; et al. Multiparametric Functional Mri: Non-Invasive Imaging of Inflammation and Edema Formation after Kidney Transplantation in Mice. PLoS ONE 2016, 11, e0162705. [Google Scholar] [CrossRef] [PubMed]
- Schley, G.; Jordan, J.; Ellmann, S.; Rosen, S.; Eckardt, K.U.; Uder, M.; Willam, C.; Bauerle, T. Multiparametric Magnetic Resonance Imaging of Experimental Chronic Kidney Disease: A Quantitative Correlation Study with Histology. PLoS ONE 2018, 13, e0200259. [Google Scholar] [CrossRef] [Green Version]
- Scallan, J.P.; Zawieja, S.D.; Castorena-Gonzalez, J.A.; Davis, M.J. Lymphatic Pumping: Mechanics, Mechanisms and Malfunction. J. Physiol. 2016, 594, 5749–5768. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Shelton, E.L.; Crescenzi, R.; Colvin, D.C.; Kirabo, A.; Zhong, J.; Delpire, E.J.; Yang, H.-C.; Kon, V. Kidney Injury Causes Accumulation of Renal Sodium That Modulates Renal Lymphatic Dynamics. Int. J. Mol. Sci. 2022, 23, 1428. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031428
Liu J, Shelton EL, Crescenzi R, Colvin DC, Kirabo A, Zhong J, Delpire EJ, Yang H-C, Kon V. Kidney Injury Causes Accumulation of Renal Sodium That Modulates Renal Lymphatic Dynamics. International Journal of Molecular Sciences. 2022; 23(3):1428. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031428
Chicago/Turabian StyleLiu, Jing, Elaine L. Shelton, Rachelle Crescenzi, Daniel C. Colvin, Annet Kirabo, Jianyong Zhong, Eric J. Delpire, Hai-Chun Yang, and Valentina Kon. 2022. "Kidney Injury Causes Accumulation of Renal Sodium That Modulates Renal Lymphatic Dynamics" International Journal of Molecular Sciences 23, no. 3: 1428. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031428