
Amniotic LPS-Induced Apoptosis in the Fetal Brain Is Suppressed by Vaginal LPS Preconditioning but Is Promoted by Continuous Ischemic Reperfusion


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
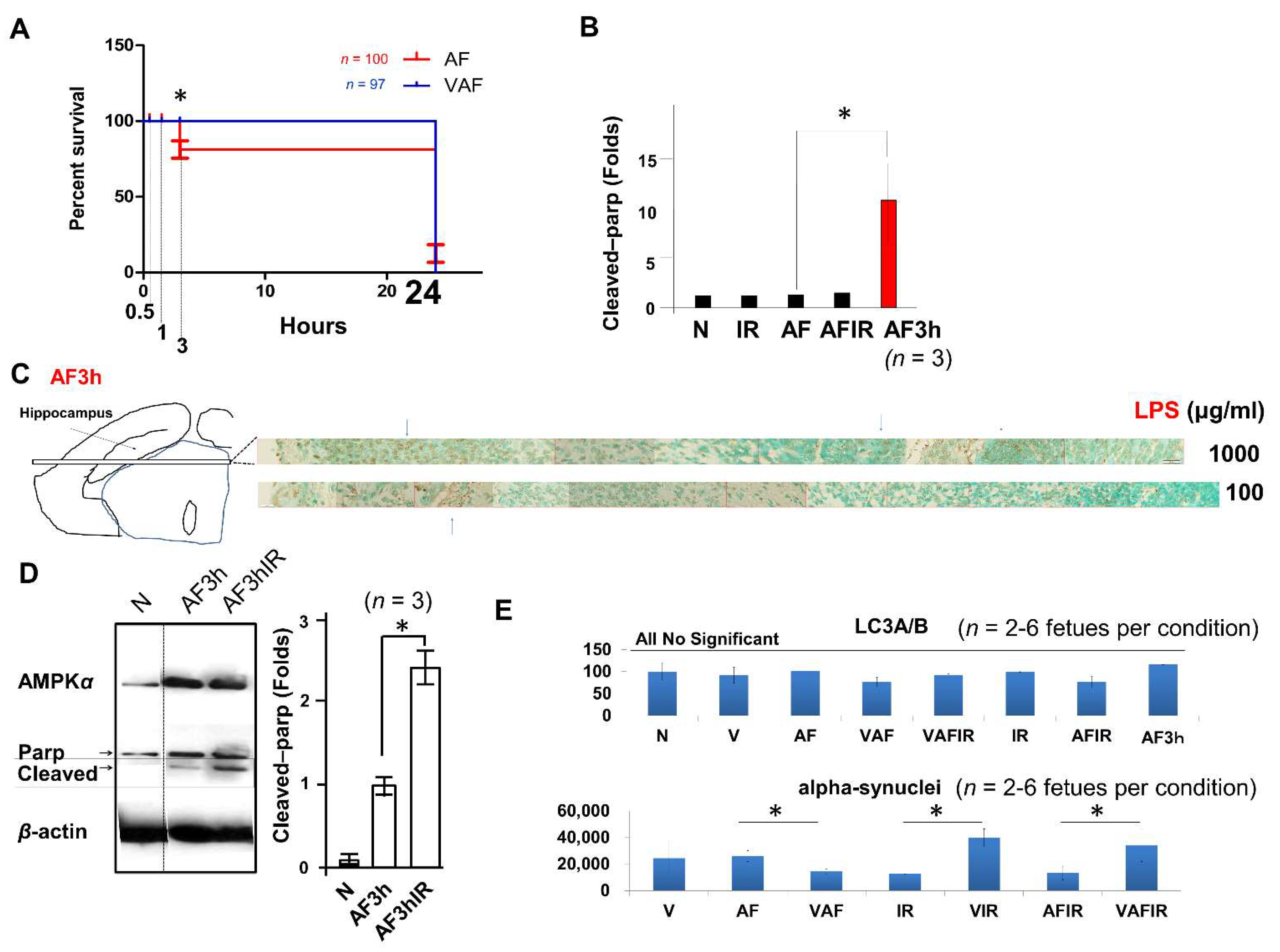
2.1. Amniotic LPS-Induced Apoptotic Cell Death Is Promoted by Continuous Ischemic Reperfusion
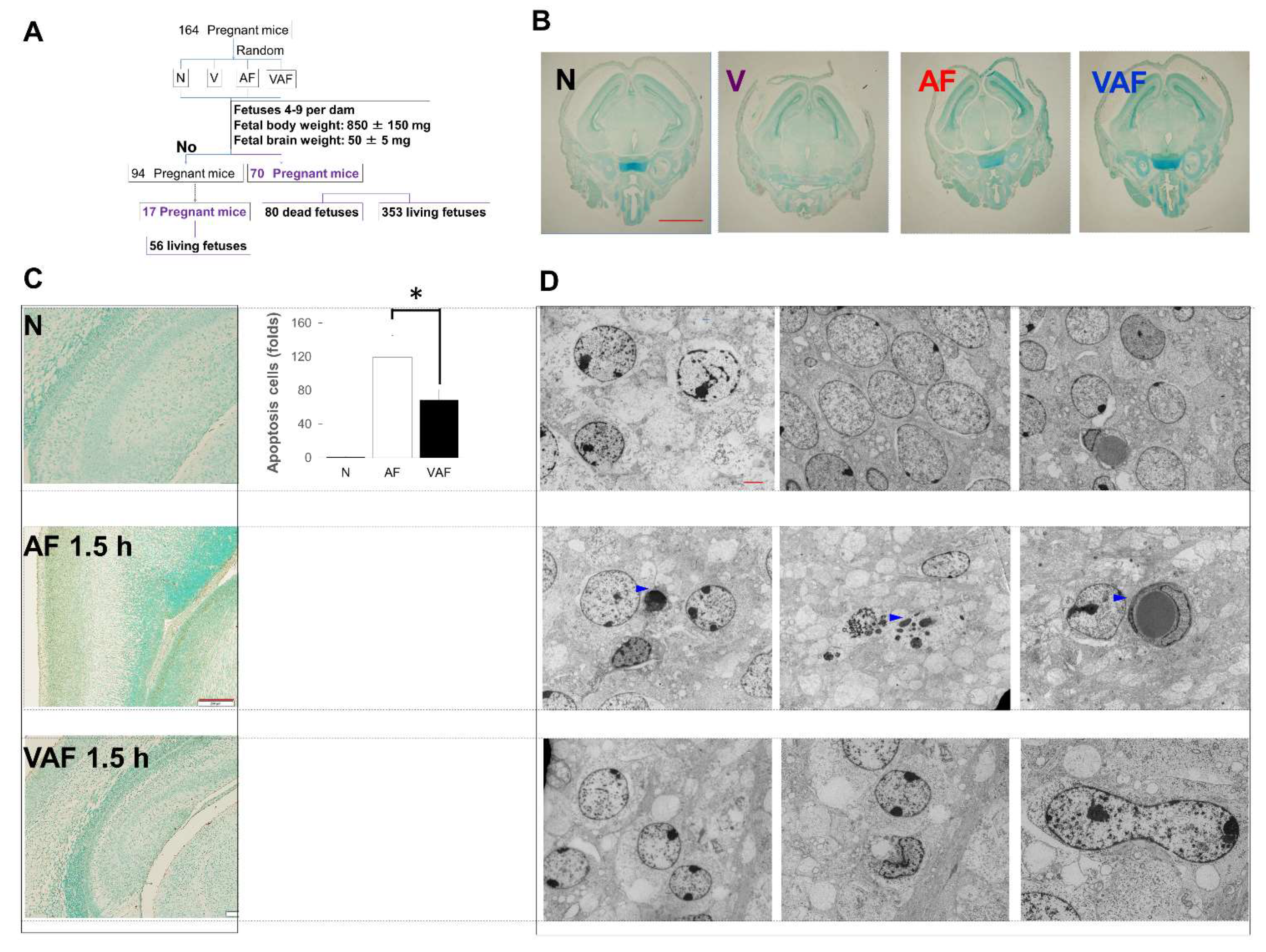
2.2. Amniotic LPS-Induced Apoptotic Cell Death Is Inhibited by Vaginal LPS Preconditioning
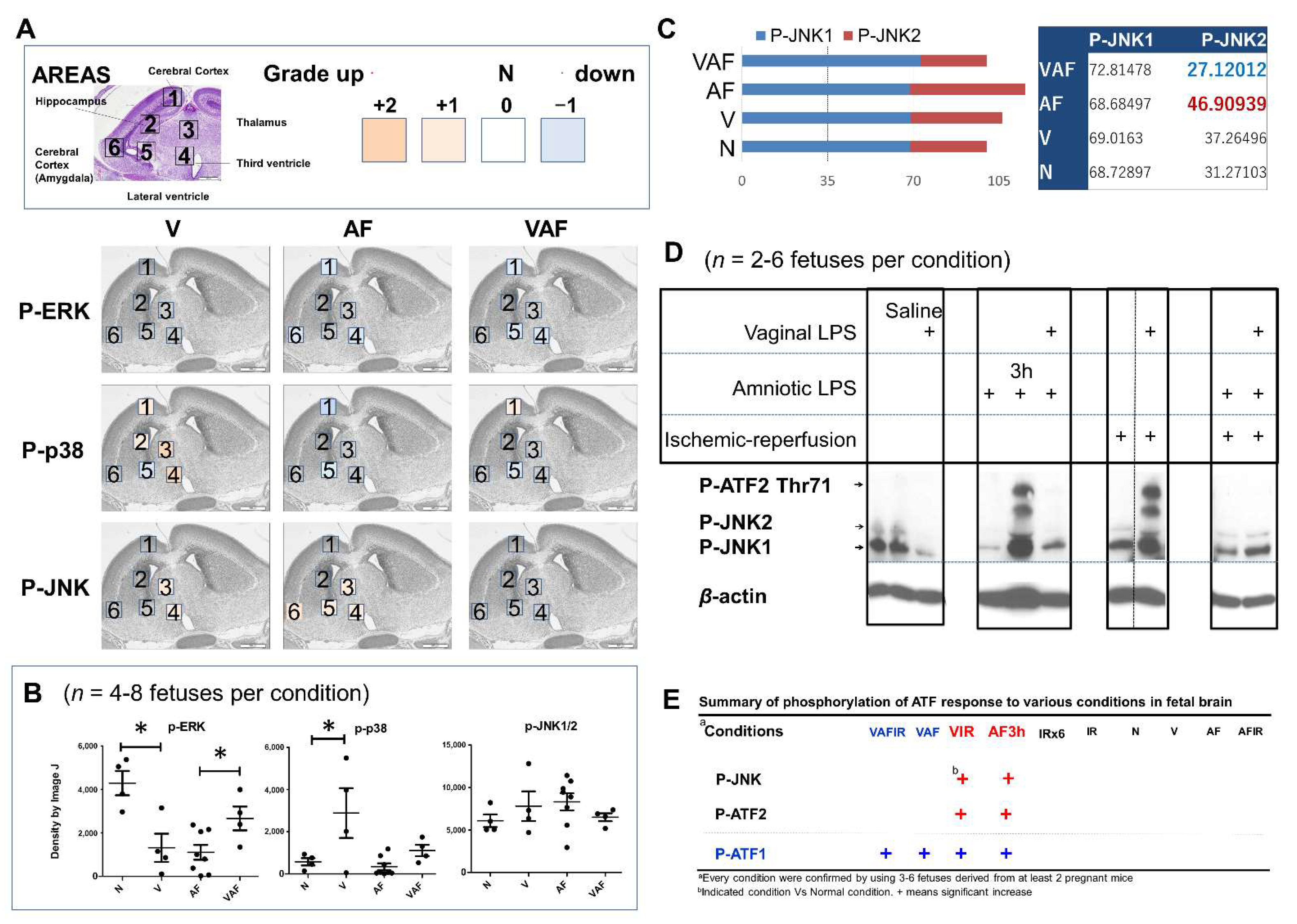
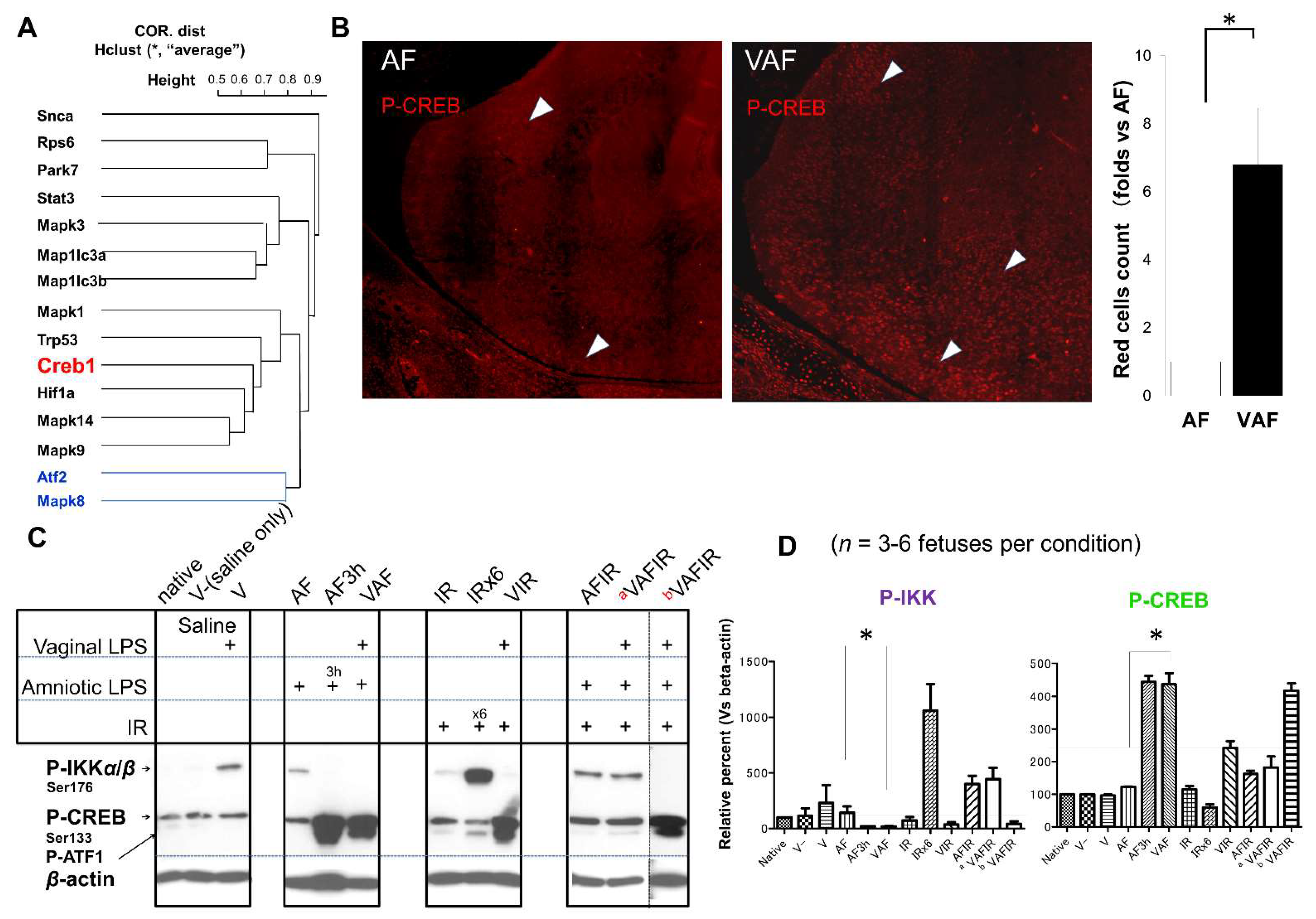
2.3. Vaginal LPS Preconditioning Rescues the Amniotic LPS-Induced Apoptosis through Cell-Signaling Transduction Pathway and Transcriptional Factors
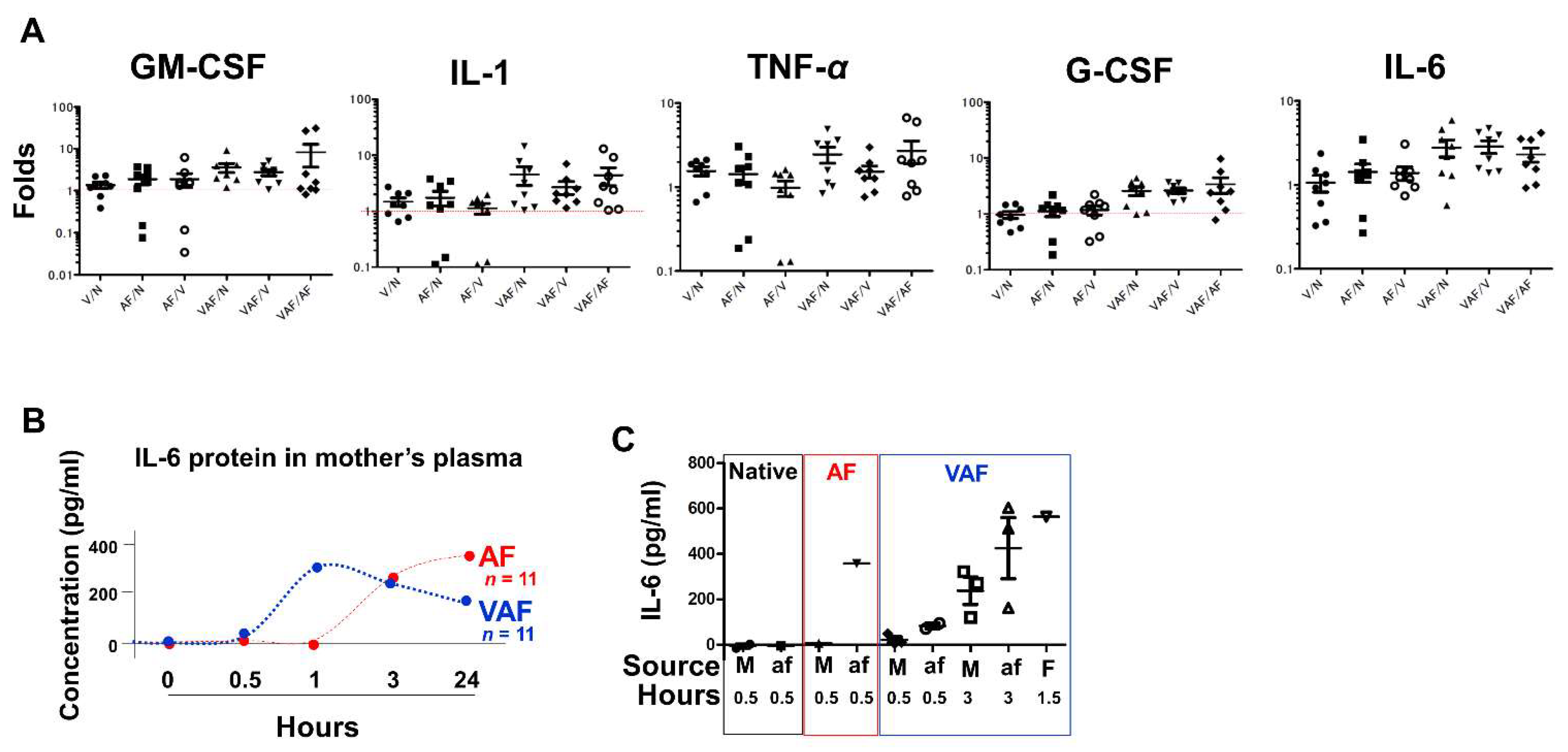
2.4. Vaginal LPS Preconditioning Results in an Earlier Inflammatory Response to Amniotic LPS including IL-6 Releasing in Fetal Brain
3. Discussion
4. Materials and Methods
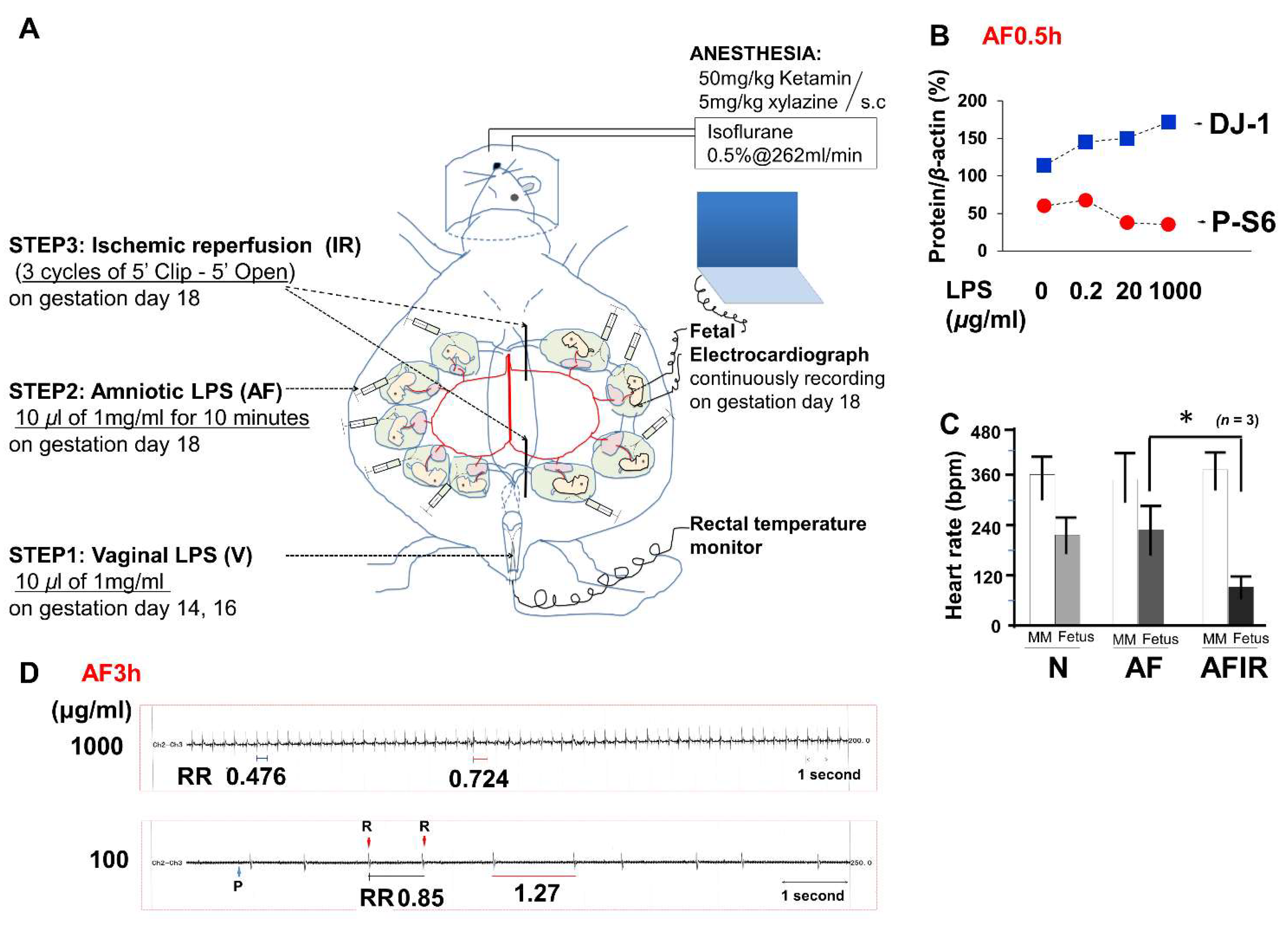
4.1. Mouse Model
- Native(N): no treatment
- Vaginal (V) LPS
- Amniotic fluid (AF) LPS
- Ischemic reperfusion (IR)
- Vaginal LPS plus amniotic fluid LPS (VAF)
- Vaginal LPS plus ischemic reperfusion (VIR)
- Amniotic fluid LPS plus ischemic reperfusion (AFIR)
- Vaginal LPS plus amniotic fluid LPS plus ischemic reperfusion (VAFIR)
- Step 1. On both GD14 and GD16, vaginal LPS was administered (10 μL of 1 mg/mL LPS, 95% confidence interval was 0.44 ± 0.14 mg/kg per pregnant mouse, n = 12 pregnant mice) using a Pipetman (Gilson, Inc., Middleton, WI, USA).
- Step 2. The temperature and humidity were controlled at 36 °C ± 1 °C and 60% ± 5%, respectively, during the cesarean section (Supplementary Figure S1A–I). All surgical tools were autoclaved at 120 °C for 20 min. GD18 pregnant mice were anesthetized with subcutaneous 50 mg/kg ketamine (Ketalar 500 mg®; Daiichi-Sankyo, Tokyo, Japan) and 5 mg/kg xylazine (Rompun™ 2% solution for injection; Bayer AG, Leverkusen, Germany) and maintained on inhalational isoflurane (Forane® 0.5%, 260 mL/min; AbbVie Inc., Chicago, IL, USA). After scrubbing in and putting on surgical gloves, the fetal electrocardiography (FECG) sensor was cleaned using 100% ethanol for 10 min before surgery (Supplementary Figure S2). Povidone iodine (7.5%) was used as an antiseptic on the skin of the mice before dissection and for the sterilization of surgical instruments. After the abdominal operation, 10 μL of LPS solution (1 mg/mL, with 95% confidence interval: 11.43 ± 4.03 mg/kg per fetus, n = 12 fetuses derived from 6 dams) was injected into each hole of amniotic fluid using sterile needles (30G) and syringes (BD, Franklin Lakes, NJ, USA), and the wounds were closed within 5 min. Fetal brains were then collected after 3 h (AF3h; n = 6 pregnant mice), or 24 h (AF24h; n = 6 pregnant mice), from amniotic fluid LPS injection for a time-course pathology analysis. If necessary, the condition of the fetuses was continuously monitored using a FECG sensor until the endpoint of 3 h after LPS injection. After surgery, pregnant mice were returned to their cages for the indicated time. For surgery 24-h samples, a 12-h on/12-h off light cycle was used, and temperature and humidity were maintained at room temperature.
- Step 3. Thirty minutes (except 3h in AF3hIR) after the amniotic LPS injection, ischemic reperfusion was performed by clipping the uterine artery for 5 min with a release for 5 min and repeating this for three cycles. Specifically, repeated twice normal IR (3 cycles), six cycles of clipping and release were performed as a worse ischemic reperfusion condition (n = 3 pregnant mice). Electromyography (EMG) was used to measure uterine muscle contractions. One EMG cycle approximated to 1 min, including a 20-s contraction and 40-s relaxation. As described in previous reports [55,56], FECG is sufficiently sensitive to monitor asphyxia-like ischemic reperfusion (Figure 1C). However, in the early period of 30 min, only amniotic LPS either without or with vaginal LPS preconditioning was hard to detect based on raw FECG data.
4.2. Quantitative and Qualitative Analyses of Fetal Mouse Brains
- Number of fetuses in a dam: 4–9
- Fetal body weight: 850 ± 150 mg
- Fetal brain weight: 50 ± 5 mg
4.3. Western Blotting
4.4. Quantitative Real-Time Polymerase Chain Reaction
4.5. Enzyme-Linked Immunosorbent Assay
4.6. Immunohistochemical Scoring
4.7. Imaging Experiments
4.8. Terminal Deoxynucleotidyl Transferase dUTP Nick-End Labeling Assay
4.9. Transmission Electron Microscopy
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Faro, S. Bacterial Vaginitis. Clin. Obstet. Gynecol. 1991, 34, 582–586. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, R.S.; Romero, R.; Hillier, S.L.; Eschenbach, D.A.; Sweet, R.L. A Review of Premature Birth and Subclinical Infection. Am. J. Obstet. Gynecol. 1992, 166, 1515–1528. [Google Scholar] [CrossRef]
- Kimberlin, D.F.; Andrews, W.W. Bacterial Vaginosis: Association with Adverse Pregnancy Outcome. Semin. Perinatol. 1998, 22, 242–250. [Google Scholar] [CrossRef]
- Peebles, K.; Velloza, J.; Balkus, J.E.; McClelland, R.S.; Barnabas, R.V. High Global Burden and Costs of Bacterial Vaginosis: A Systematic Review and Meta-Analysis. Sex Transm. Dis. 2019, 46, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Shatrov, J.G.; Birch, S.C.; Lam, L.T.; Quinlivan, J.A.; McIntyre, S.; Mendz, G.L. Chorioamnionitis and Cerebral Palsy: A Meta-analysis. Obstet. Gynecol. 2010, 116, 387–392. [Google Scholar] [CrossRef]
- Soraisham, A.S.; Trevenen, C.; Wood, S.; Singhal, N.; Sauve, R. Histological Chorioamnionitis and Neurodevelopmental Outcome in Preterm Infants. J. Perinatol. 2012, 33, 70–75. [Google Scholar] [CrossRef] [Green Version]
- Berger, R.; Garnier, Y. Perinatal Brain Injury. J. Perinat. Med. 2000, 28, 261–285. [Google Scholar] [CrossRef]
- Brunel, H.; Girard, N.; Confort-Gouny, S.; Viola, A.; Chaumoitre, K.; D’Ercole, C.; Figarella-Branger, D.; Raybaud, C.; Cozzone, P.; Panuel, M. Fetal Brain Injury. J. Neuroradiol. 2004, 31, 123–137. [Google Scholar] [CrossRef]
- Baier, R.J. Genetics of Perinatal Brain Injury in the Preterm Infant. Front. Biosci. 2005, 11, 1371–1387. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Rousset, C.I.; Hagberg, H.; Mallard, C. Lipopolysaccharide-Induced Inflammation and Perinatal Brain Injury. Semin. Fetal Neonatal Med. 2006, 11, 343–353. [Google Scholar] [CrossRef]
- Burd, I.; Balakrishnan, B.; Kannan, S. Models of Fetal Brain Injury, Intrauterine Inflammation, and Preterm birth. Am. J. Reprod. Immunol. 2012, 67, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Hagberg, H.; Mallard, C.; Ferriero, D.M.; Vannucci, S.J.; Levison, S.W.; Vexler, Z.S.; Gressens, P. The Role of Inflammation in Perinatal Brain Injury. Nat. Rev. Neurol. 2015, 11, 192–208. [Google Scholar] [CrossRef] [PubMed]
- Domínguez Rubio, A.P.; Correa, F.; Aisemberg, J.; Dorfman, D.; Bariani, M.V.; Rosenstein, R.E.; Zorrilla Zubilete, M.; Franchi, A.M. Maternal Administration of Melatonin Exerts Short- and Long-Term Neuroprotective Effects on the Offspring from Lipopolysaccharide-Treated Mice. J. Pineal. Res. 2017, 63, e12439. [Google Scholar] [CrossRef] [PubMed]
- Paton, M.C.B.; Allison, B.J.; Li, J.; Fahey, M.C.; Sutherland, A.E.; Nitsos, I.; Bischof, R.J.; Dean, J.M.; Moss, T.J.M.; Polglase, G.R.; et al. Human Umbilical Cord Blood Therapy Protects Cerebral White Matter from Systemic LPS Exposure in Preterm Fetal Sheep. Dev. Neurosci. 2018, 40, 258–270. [Google Scholar] [CrossRef]
- McCartney, S.A.; Kapur, R.; Liggitt, H.D.; Baldessari, A.; Coleman, M.; Orvis, A.; Ogle, J.; Katz, R.; Rajagopal, L.; Adams Waldorf, K.M. Amniotic Fluid Interleukin 6 and Interleukin 8 Are Superior Predictors of Fetal Lung Injury Compared with Maternal or Fetal Plasma Cytokines or Placental Histopathology in a Nonhuman Primate Model. Am. J. Obstet. Gynecol. 2021, 225, e81–e89. [Google Scholar] [CrossRef]
- Cai, Z.; Pan, Z.L.; Pang, Y.; Evans, O.B.; Rhodes, P.G. Cytokine Induction in Fetal Rat Brains and Brain Injury in Neonatal Rats after Maternal Lipopolysaccharide Administration. Pediatr. Res. 2000, 47, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Kallapur, S.G.; Nitsos, I.; Moss, T.J.; Polglase, G.R.; Pillow, J.J.; Cheah, F.C.; Kramer, B.W.; Newnham, J.P.; Ikegami, M.; Jobe, A.H. IL-1 Mediates Pulmonary and Systemic Inflammatory Responses to Chorioamnionitis Induced by Lipopolysaccharide. Am. J. Respir. Crit. Care Med. 2009, 179, 955–961. [Google Scholar] [CrossRef] [Green Version]
- Duncan, J.R.; Cock, M.L.; Suzuki, K.; Scheerlinck, J.P.; Harding, R.; Rees, S.M. Chronic Endotoxin Exposure Causes Brain Injury in the Ovine Fetus in the Absence of Hypoxemia. J. Soc. Gynecol. Investig. 2006, 13, 87–96. [Google Scholar] [CrossRef]
- Rousset, C.I.; Chalon, S.; Cantagrel, S.; Bodard, S.; Andres, C.; Gressens, P.; Saliba, E. Maternal Exposure to LPS Induces Hypomyelination in the Internal Capsule and Programmed Cell Death in the Deep Gray Matter in Newborn Rats. Pediatr. Res. 2006, 59, 428–433. [Google Scholar] [CrossRef] [Green Version]
- Bell, M.J.; Hallenbeck, J.M.; Gallo, V. Determining the Fetal Inflammatory Response in an Experimental Model of Intrauterine Inflammation in Rats. Pediatr. Res. 2004, 56, 541–546. [Google Scholar] [CrossRef] [Green Version]
- Rounioja, S.; Räsänen, J.; Glumoff, V.; Ojaniemi, M.; Mäkikallio, K.; Hallman, M. Intra-Amniotic Lipopolysaccharide Leads to Fetal Cardiac Dysfunction. A Mouse Model for Fetal Inflammatory Response. Cardiovasc. Res. 2003, 60, 156–164. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Tang, Y.; Wang, H.; Chen, W.; Jiang, H. Curcumin Alleviates Lipopolysaccharide-Induced Neuroinflammation in Fetal Mouse Brain. Restor. Neurol. Neurosci. 2018, 36, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Hagberg, H.; Peebles, D.; Mallard, C. Models of White Matter Injury: Comparison of Infectious, Hypoxic-Ischemic, and Excitotoxic Insults. Ment. Retard. Dev. Disabil. Res. Rev. 2002, 8, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Silver, R.M.; Lohner, W.S.; Daynes, R.A.; Mitchell, M.D.; Branch, D.W. Lipopolysaccharide-Induced Fetal Death: The Role of Tumor-Necrosis Factor Alpha. Biol. Reprod. 1994, 50, 1108–1112. [Google Scholar] [CrossRef] [PubMed]
- Sharp, G.C.; Ma, H.; Saunders, P.T.; Norman, J.E. A Computational Model of Lipopolysaccharide-Induced Nuclear Factor Kappa B Activation: A Key Signalling Pathway in Infection-Induced Preterm Labour. PLoS ONE 2013, 8, e70180. [Google Scholar] [CrossRef]
- Mathai, S.; Booth, L.C.; Davidson, J.O.; Drury, P.P.; Fraser, M.; Jensen, E.C.; George, S.; Naylor, A.; Gunn, A.J.; Bennet, L. Acute on Chronic Exposure to Endotoxin in Preterm Fetal Sheep. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 304, R189–R197. [Google Scholar] [CrossRef] [Green Version]
- van den Heuij, L.G.; Mathai, S.; Davidson, J.O.; Lear, C.A.; Booth, L.C.; Fraser, M.; Gunn, A.J.; Bennet, L. Synergistic White Matter Protection with Acute-on-Chronic Endotoxin and Subsequent Asphyxia in Preterm Fetal Sheep. J. Neuroinflamm. 2014, 11, 89. [Google Scholar] [CrossRef] [Green Version]
- Dhillon, S.K.; Gunn, A.J.; Jung, Y.; Mathai, S.; Bennet, L.; Fraser, M. Lipopolysaccharide-Induced Preconditioning Attenuates Apoptosis and Differentially Regulates TLR4 and TLR7 Gene Expression after Ischemia in the Preterm Ovine Fetal Brain. Deve. Neurosci. 2015, 37, 497–514. [Google Scholar] [CrossRef]
- Lai, J.L.; Liu, Y.H.; Liu, C.; Qi, M.P.; Liu, R.N.; Zhu, X.F.; Zhou, Q.G.; Chen, Y.Y.; Guo, A.Z.; Hu, C.M. Indirubin Inhibits LPS-Induced Inflammation via TLR4 Abrogation Mediated by the NF-kB and MAPK Signaling Pathways. Inflammation 2016, 40, 1–12. [Google Scholar] [CrossRef]
- Li, X.; Peng, H.; Wu, J.; Xu, Y. Brain Natriuretic Peptide-Regulated Expression of Inflammatory Cytokines in Lipopolysaccharide (LPS)-Activated Macrophages via NF-κB and Mitogen Activated Protein Kinase (MAPK) Pathways. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 3119–3126. [Google Scholar] [CrossRef]
- Ferrer, I.; Blanco, R.; Carmona, M. Differential Expression of Active, Phosphorylation-Dependent MAP Kinases, MAPK/ERK, SAPK/JNK and p38, and Specific Transcription Factor Substrates Following Quinolinic Acid Excitotoxicity in the Rat. Brain Res. Mol. Brain Res. 2001, 94, 48–58. [Google Scholar] [CrossRef]
- Ferrer, I.; Friguls, B.; Dalfó, E.; Planas, A.M. Early Modifications in the Expression of Mitogen-Activated Protein Kinase (MAPK/ERK), Stress-Activated Kinases SAPK/JNK and p38, and Their Phosphorylated Substrates Following Focal Cerebral Ischemia. Acta Neuropathol. 2003, 105, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Zaman, K.; Ryu, H.; Hall, D.; O’Donovan, K.; Lin, K.I.; Miller, M.P.; Marquis, J.C.; Baraban, J.M.; Semenza, G.L.; Ratan, R.R. Protection from Oxidative Stress-Induced Apoptosis in Cortical Neuronal Cultures by Iron Chelators Is Associated with Enhanced DNA Binding of Hypoxia-Inducible Factor-1 and ATF-1/CREB and Increased Expression of Glycolytic Enzymes, p21(waf1/cip1), and Erythropoietin. J. Neurosci. Off. J. Soc. Neurosci. 1999, 19, 9821–9830. [Google Scholar] [CrossRef] [Green Version]
- Lonze, B.E.; Riccio, A.; Cohen, S.; Ginty, D.D. Apoptosis, Axonal Growth Defects, and Degeneration of Peripheral Neurons in Mice Lacking CREB. Neuron 2002, 34, 371–385. [Google Scholar] [CrossRef] [Green Version]
- Matt, T. Transcriptional Control of the Inflammatory Response: A Role for the CREB-Binding Protein (CBP). Acta Med. Aus. 2002, 29, 77–79. [Google Scholar] [CrossRef]
- Dragunow, M. CREB and Neurodegeneration. Front. Biosci. 2004, 9, 100–103. [Google Scholar] [CrossRef]
- Kitagawa, K. CREB and cAMP Response Element-Mediated Gene Expression in the Ischemic Brain. FEBS J. 2007, 274, 3210–3217. [Google Scholar] [CrossRef]
- Carloni, S.; Girelli, S.; Scopa, C.; Buonocore, G.; Longini, M.; Balduini, W. Activation of Autophagy and Akt/CREB Signaling Play an Equivalent Role in the Neuroprotective Effect of Rapamycin in Neonatal Hypoxia-Ischemia. Autophagy 2010, 6, 366–377. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y.W.; Zhang, S.J.; Hoffmann, T.; Bading, H. Increasing Levels of Wild-Type CREB up-Regulates Several Activity-Regulated Inhibitor of Death (AID) Genes and Promotes Neuronal Survival. BMC Neurosci. 2012, 13, 48. [Google Scholar] [CrossRef] [Green Version]
- Planeta, C.S.; Lepsch, L.B.; Alves, R.; Scavone, C. Influence of the Dopaminergic System, CREB, and Transcription Factor-κB on Cocaine Neurotoxicity. Braz. J. Med. Biol. Res. 2013, 46, 909–915. [Google Scholar] [CrossRef]
- Hu, Y.; Liu, M.Y.; Liu, P.; Dong, X.; Boran, A.D. Neuroprotective Effects of 3,6’-Disinapoyl Sucrose through Increased BDNF Levels and CREB Phosphorylation via the CaMKII and ERK1/2 Pathway. J. Mol. Neurosci. 2014, 53, 600–607. [Google Scholar] [CrossRef] [PubMed]
- Ko, H.R.; Ahn, S.Y.; Chang, Y.S.; Hwang, I.; Yun, T.; Sung, D.K.; Sung, S.I.; Park, W.S.; Ahn, J.Y. Human UCB-MSCs Treatment upon Intraventricular Hemorrhage Contributes to Attenuate Hippocampal Neuron Loss and Circuit Damage through BDNF-CREB Signaling. Stem Cell Res. Ther. 2018, 9, 326. [Google Scholar] [CrossRef] [PubMed]
- Olanow, C.W.; Brundin, P. Parkinson’s Disease and Alpha Synuclein: Is Parkinson’s Disease a Prion-Like Disorder? Mov. Disord. Off. J. Mov. Disord. Soc. 2013, 28, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.; Kordower, J.H. Lewy Body Pathology in Fetal Grafts. Ann. N. Y. Acad. Sci. 2010, 1184, 55–67. [Google Scholar] [CrossRef]
- Ebadi, M.; Brown-Borg, H.; El Refaey, H.; Singh, B.B.; Garrett, S.; Shavali, S.; Sharma, S.K. Metallothionein-Mediated Neuroprotection in Genetically Engineered Mouse Models of Parkinson’s Disease. Brain Res. Mol. Brain Res. 2005, 134, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Lozano, A.M.; Lang, A.E.; Hutchison, W.D.; Dostrovsky, J.O. New Developments in Understanding the Etiology of Parkinson’s Disease and in Its Treatment. Curr. Opin. Neurobiol. 1999, 8, 783–790. [Google Scholar] [CrossRef]
- Vázquez-Vélez, G.E.; Gonzales, K.A.; Revelli, J.P.; Adamski, C.J.; Alavi Naini, F.; Bajić, A.; Craigen, E.; Richman, R.; Heman-Ackah, S.M.; Wood, M.J.A.; et al. Doublecortin-Like Kinase 1 Regulates α-Synuclein Levels and Toxicity. J. Neurosci. Off. J. Soc. Neurosci. 2020, 40, 459–477. [Google Scholar] [CrossRef]
- Koshino, T.; Kimura, Y.; Kameyama, Y.; Takahashi, T.; Yasui, T.; Chisaka, H.; Sugawara, J.; Okamura, K. Fractal and Periodic Heart Rate Dynamics in Fetal Sheep: Comparison of Conventional and New Measures Based on Fractal Analysis. Am. J. Physiol. Heart Circ. Physiol. 2003, 284, H1858–H1864. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.; Kimura, Y.; Chida, S.; Ito, T.; Katayama, N.; Okamura, K.; Nakao, M. A Novel Extraction Method of Fetal Electrocardiogram from the Composite Abdominal Signal. IEEE Trans. Biomed. Eng. 2007, 54, 49–58. [Google Scholar] [CrossRef]
- Sethi, N.; Funamoto, K.; Ingbar, C.; Mass, P.; Moak, J.; Wakai, R.; Strasburger, J.; Donofrio, M.; Khandoker, A.; Kimura, Y.; et al. Noninvasive Fetal Electrocardiography in the Diagnosis of Long QT Syndrome: A Case Series. Fetal Diagn. Ther. 2020, 47, 711–716. [Google Scholar] [CrossRef]
- Widatalla, N.; Khandoker, A.; Kasahara, Y.; Kimura, Y. Detection of End of T-Wave in Fetal ECG Using Recurrence Plots. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. 2019, 2019, 2618–2621. [Google Scholar] [CrossRef] [PubMed]
- Stoker, T.B.; Greenland, J.C. Parkinson’s Disease: Pathogenesis and Clinical Aspects; Exon Publications: Brisbane, Australia, 2018. [Google Scholar]
- Sabapathy, K.; Wagner, E.F. JNK2: A Negative Regulator of Cellular Proliferation. Cell Cycle 2004, 3, 1520–1523. [Google Scholar] [CrossRef] [PubMed]
- Waetzig, V.; Czeloth, K.; Hidding, U.; Mielke, K.; Kanzow, M.; Brecht, S.; Goetz, M.; Lucius, R.; Herdegen, T.; Hanisch, U.K. c-Jun N-Terminal Kinases (JNKs) Mediate Pro-Inflammatory Actions of Microglia. Glia 2005, 50, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Ito, T.; Velayo, C.; Sato, T.; Iida, K.; Endo, M.; Funamoto, K.; Sato, N.; Yaegashi, N.; Kimura, Y. Intrauterine Ischemic Reperfusion Switches the Fetal Transcriptional Pattern from HIF-1α- to P53-Dependent Regulation in the Murine Brain. PLoS ONE 2014, 9, e110577. [Google Scholar] [CrossRef]
- Dong, Y.; Kimura, Y.; Ito, T.; Velayo, C.; Sato, T.; Sugibayashi, R.; Funamoto, K.; Hitomi, K.; Iida, K.; Endo, M.; et al. Vaginal LPS Changed Gene Transcriptional Regulation Response to Ischemic Reperfusion and Increased Vulnerability of Fetal Brain Hemorrhage. Biochem. Biophys. Res. Commun. 2015, 468, 228–233. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, Y.; Kimura, Y.; Yaegashi, N. Amniotic LPS-Induced Apoptosis in the Fetal Brain Is Suppressed by Vaginal LPS Preconditioning but Is Promoted by Continuous Ischemic Reperfusion. Int. J. Mol. Sci. 2022, 23, 1787. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031787
Dong Y, Kimura Y, Yaegashi N. Amniotic LPS-Induced Apoptosis in the Fetal Brain Is Suppressed by Vaginal LPS Preconditioning but Is Promoted by Continuous Ischemic Reperfusion. International Journal of Molecular Sciences. 2022; 23(3):1787. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031787
Chicago/Turabian StyleDong, Yupeng, Yoshitaka Kimura, and Nobuo Yaegashi. 2022. "Amniotic LPS-Induced Apoptosis in the Fetal Brain Is Suppressed by Vaginal LPS Preconditioning but Is Promoted by Continuous Ischemic Reperfusion" International Journal of Molecular Sciences 23, no. 3: 1787. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031787