
The Thermal Stability of the Collagen Triple Helix Is Tuned According to the Environmental Temperature

 ,
,
Abstract
:1. Introduction
2. Results
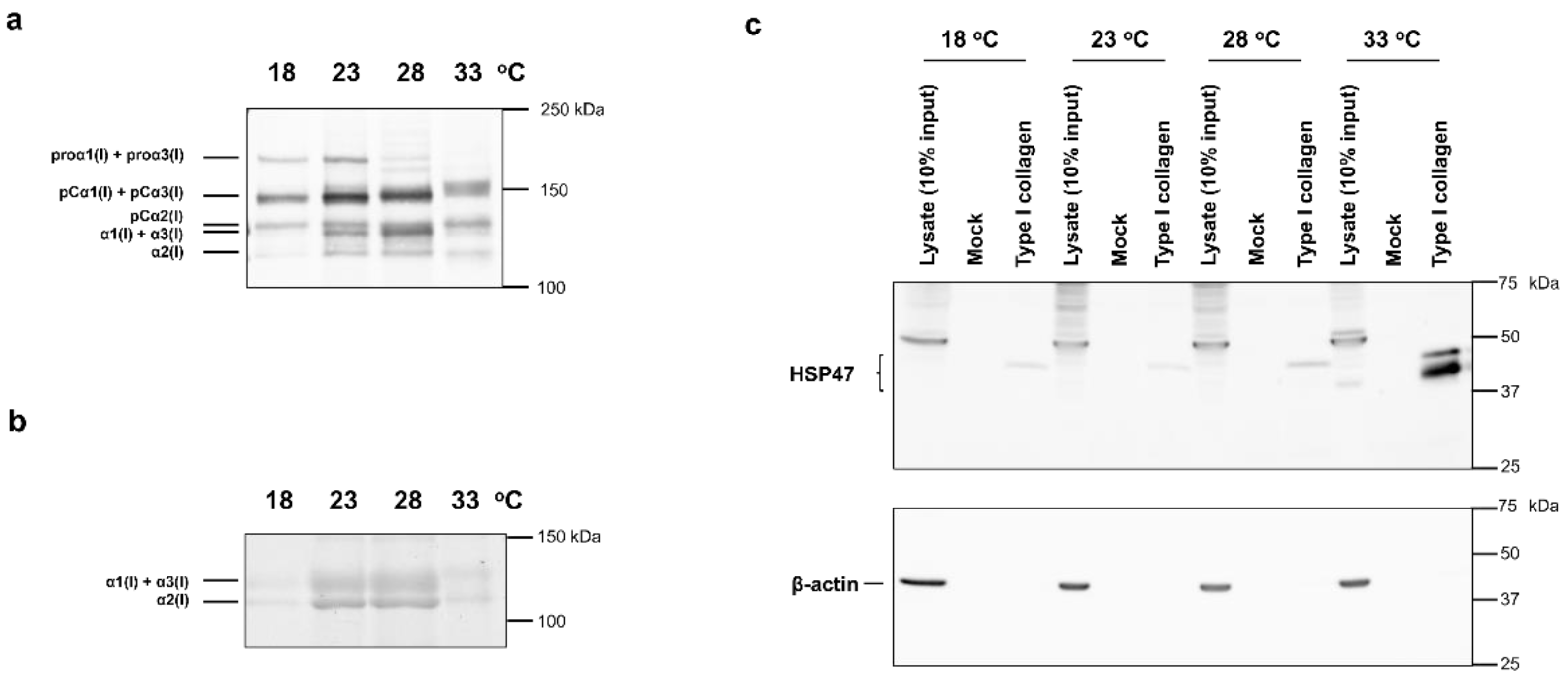
2.1. Western Blot Analysis of Proteins Expressed by ZF4 Cells Cultured at Different Temperatures
2.2. Quantification of Total Post-Translational Modifications of Collagen
2.3. Quantification of Site-Specific 3-Hyp Modification
2.4. α-Chain Ratios of Type I Collagen Secreted from ZF4 Cells
2.5. Thermal Stability of Collagen Triple-Helical Structure Secreted from ZF4 Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. SDS-PAGE Analysis of Pepsin-Treated Collagen Samples
4.3. Western Blotting Analysis of Culture Media Samples and Pull-Down Samples
4.4. Synthesis of Stable Isotope-Labeled Peptides
4.5. Quantification of Type I Collagen α-Chains by LC-MS
4.6. Quantification of Total Post-Translational Modifications in Collagen by LC-MS
4.7. Site-Specific Characterization of Prolyl 3-Hydroxylation in Zebrafish Type I Collagen by LC-MS
4.8. Calculation of the Amount Ratios of Possible Trimers in Type I Collagen
4.9. Measurement of the Melting Temperature of Collagen
4.10. Prediction of Relative Thermal Stability of Possible Isoforms of Zebrafish Type I Collagen
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ricard-Blum, S. The Collagen Family. Cold Spring Harb. Perspect. Biol. 2011, 3, a004978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg, R.A.; Prockop, D.J. The Thermal Transition of a Non-Hydroxylated Form of Collagen. Evidence for a Role for Hydroxyproline in Stabilizing the Triple-Helix of Collagen. Biochem. Biophys. Res. Commun. 1973, 52, 115–120. [Google Scholar] [CrossRef]
- Burjanadze, T.V. Hydroxyproline Content and Location in Relation to Collagen Thermal Stability. Biopolymers 1979, 18, 931–938. [Google Scholar] [CrossRef] [PubMed]
- Burjanadze, T.V. New Analysis of the Phylogenetic Change of Collagen Thermostability. Biopolymers 2000, 53, 523–528. [Google Scholar] [CrossRef]
- Taga, Y.; Tanaka, K.; Hattori, S.; Mizuno, K. In-Depth Correlation Analysis Demonstrates That 4-Hydroxyproline at the Yaa Position of Gly-Xaa-Yaa Repeats Dominantly Stabilizes Collagen Triple Helix. Matrix Biol. Plus 2021, 10, 100067. [Google Scholar] [CrossRef]
- Mizuno, K.; Peyton, D.H.; Hayashi, T.; Engel, J.; Bächinger, H.P. Effect of the -Gly-3(S)-Hydroxyprolyl-4(R)-Hydroxyprolyl- Tripeptide Unit on the Stability of Collagen Model Peptides. FEBS J. 2008, 275, 5830–5840. [Google Scholar] [CrossRef]
- Nokelainen, M.; Helaakoski, T.; Myllyharju, J.; Notbohm, H.; Pihlajaniemi, T.; Fietzek, P.P.; Kivirikko, K.I. Expression and Characterization of Recombinant Human Type II Collagens with Low and High Contents of Hydroxylysine and Its Glycosylated Forms. Matrix Biol. 1998, 16, 329–338. [Google Scholar] [CrossRef]
- Takuwa, A.; Yoshida, T.; Maruno, T.; Kawahara, K.; Mochizuki, M.; Nishiuchi, Y.; Kobayashi, Y.; Ohkubo, T. Ordered Self-Assembly of the Collagenous Domain of Adiponectin with Noncovalent Interactions via Glycosylated Lysine Residues. FEBS Lett. 2016, 590, 195–201. [Google Scholar] [CrossRef]
- Duan, R.; Konno, K.; Zhang, J.; Wang, S.; Yuan, C. Different Thermostability of Collagens from Scale of Carp (Cyprinus carpio) in Winter and Summer. J. Food Biochem. 2010, 34, 1275–1287. [Google Scholar] [CrossRef]
- Engel, J.; Prockop, D.J. The Zipper-like Folding of Collagen Triple Helices and the Effects of Mutations That Disrupt the Zipper. Annu. Rev. Biophys. Biophys. Chem. 1991, 20, 137–152. [Google Scholar] [CrossRef]
- Koide, T.; Nagata, K. Collagen Biosynthesis. In Collagen: Primer in Structure, Processing and Assembly; Brinckmann, J., Notbohm, H., Müller, P.K., Eds.; Topics in Current Chemistry; Springer: Berlin\Heidelberg, Germany, 2005; pp. 85–114. ISBN 978-3-540-31472-1. [Google Scholar]
- Myllyharju, J. Intracellular Post-Translational Modifications of Collagens. In Topics in Current Chemistry; Springer: New York, NY, USA, 2005; Volume 247, pp. 115–147. [Google Scholar]
- Forrester, A.; De Leonibus, C.; Grumati, P.; Fasana, E.; Piemontese, M.; Staiano, L.; Fregno, I.; Raimondi, A.; Marazza, A.; Bruno, G.; et al. A Selective ER-Phagy Exerts Procollagen Quality Control via a Calnexin-FAM134B Complex. EMBO J. 2019, 38, e99847. [Google Scholar] [CrossRef] [PubMed]
- Ishida, Y.; Kubota, H.; Yamamoto, A.; Kitamura, A.; Bächinger, H.P.; Nagata, K. Type I Collagen in Hsp47-null Cells Is Aggregated in Endoplasmic Reticulum and Deficient in N-Propeptide Processing and Fibrillogenesis. Mol. Biol. Cell 2006, 17, 2346–2355. [Google Scholar] [CrossRef] [PubMed]
- Koide, T.; Aso, A.; Yorihuzi, T.; Nagata, K. Conformational Requirements of Collagenous Peptides for Recognition by the Chaperone Protein HSP47. J. Biol. Chem. 2000, 275, 27957–27963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tasab, M.; Batten, M.R.; Bulleid, N.J. Hsp47: A Molecular Chaperone That Interacts with and Stabilizes Correctly-Folded Procollagen. EMBO J. 2000, 19, 2204–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagai, N.; Hosokawa, M.; Itohara, S.; Adachi, E.; Matsushita, T.; Hosokawa, N.; Nagata, K. Embryonic Lethality of Molecular Chaperone Hsp47 Knockout Mice Is Associated with Defects in Collagen Biosynthesis. J. Cell Biol. 2000, 150, 1499–1506. [Google Scholar] [CrossRef] [Green Version]
- Fujii, K.K.; Taga, Y.; Sakai, T.; Ito, S.; Hattori, S.; Nagata, K.; Koide, T. Lowering the Culture Temperature Corrects Collagen Abnormalities Caused by HSP47 Gene Knockout. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Drögemüller, C.; Becker, D.; Brunner, A.; Haase, B.; Kircher, P.; Seeliger, F.; Fehr, M.; Baumann, U.; Lindblad-Toh, K.; Leeb, T. A Missense Mutation in the SERPINH1 Gene in Dachshunds with Osteogenesis Imperfecta. PLOS Genet. 2009, 5, e1000579. [Google Scholar] [CrossRef] [Green Version]
- Christiansen, H.E.; Schwarze, U.; Pyott, S.M.; AlSwaid, A.; Al Balwi, M.; Alrasheed, S.; Pepin, M.G.; Weis, M.A.; Eyre, D.R.; Byers, P.H. Homozygosity for a Missense Mutation in SERPINH1, Which Encodes the Collagen Chaperone Protein HSP47, Results in Severe Recessive Osteogenesis Imperfecta. Am. J. Hum. Genet. 2010, 86, 389–398. [Google Scholar] [CrossRef] [Green Version]
- Cortemeglia, C.; Beitinger, T.L. Temperature Tolerances of Wild-Type and Red Transgenic Zebra Danios. Trans. Am. Fish. Soc. 2005, 134, 1431–1437. [Google Scholar] [CrossRef]
- Schaefer, J.; Ryan, A. Developmental Plasticity in the Thermal Tolerance of Zebrafish Danio Rerio. J. Fish Biol. 2006, 69, 722–734. [Google Scholar] [CrossRef]
- Morvan-Dubois, G.; Le Guellec, D.; Garrone, R.; Zylberberg, L.; Bonnaud, L. Phylogenetic Analysis of Vertebrate Fibrillar Collagen Locates the Position of Zebrafish α3(I) and Suggests an Evolutionary Link between Collagen α Chains and Hox Clusters. J. Mol. Evol. 2003, 57, 501–514. [Google Scholar] [CrossRef] [PubMed]
- Takita, K.K.; Fujii, K.K.; Ishii, K.; Koide, T. Structural Optimization of Cyclic Peptides That Efficiently Detect Denatured Collagen. Org. Biomol. Chem. 2019, 17, 7380–7387. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Li, L.; Li, Q.; He, X.; Cui, Z. Transcriptomic Characterization of Temperature Stress Responses in Larval Zebrafish. PLoS ONE 2012, 7, e37209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Wan, Y.; Shen, Z.; Liu, Y.; Li, D.; Li, L.; Tang, R.; Zhang, X. Molecular Characterization of Hsp47 in Grass Carp (Ctenopharyngodon Idella) and Its Correlation with Type I Collagen in Response to Fish Aerobic Exercise. Fishes 2021, 6, 17. [Google Scholar] [CrossRef]
- Taga, Y.; Kusubata, M.; Ogawa-Goto, K.; Hattori, S. Stable Isotope-Labeled Collagen: A Novel and Versatile Tool for Quantitative Collagen Analyses Using Mass Spectrometry. J. Proteome Res. 2014, 13, 3671–3678. [Google Scholar] [CrossRef]
- Taga, Y.; Kusubata, M.; Ogawa-Goto, K.; Hattori, S. Developmental Stage-Dependent Regulation of Prolyl 3-Hydroxylation in Tendon Type I Collagen. J. Biol. Chem. 2016, 291, 837–847. [Google Scholar] [CrossRef] [Green Version]
- Tonelli, F.; Cotti, S.; Leoni, L.; Besio, R.; Gioia, R.; Marchese, L.; Giorgetti, S.; Villani, S.; Gistelinck, C.; Wagener, R.; et al. Crtap and P3h1 Knock out Zebrafish Support Defective Collagen Chaperoning as the Cause of Their Osteogenesis Imperfecta Phenotype. Matrix Biol. 2020, 90, 40–60. [Google Scholar] [CrossRef]
- DiChiara, A.S.; Li, R.C.; Suen, P.H.; Hosseini, A.S.; Taylor, R.J.; Weickhardt, A.F.; Malhotra, D.; McCaslin, D.R.; Shoulders, M.D. A Cysteine-Based Molecular Code Informs Collagen C-Propeptide Assembly. Nat. Commun. 2018, 9, 4206. [Google Scholar] [CrossRef]
- Walker, D.R.; Hulgan, S.A.H.; Peterson, C.M.; Li, I.-C.; Gonzalez, K.J.; Hartgerink, J.D. Predicting the Stability of Homotrimeric and Heterotrimeric Collagen Helices. Nat. Chem. 2021, 13, 260–269. [Google Scholar] [CrossRef]
- Kawahara, K.; Yoshida, T.; Maruno, T.; Oki, H.; Ohkubo, T.; Koide, T.; Kobayashi, Y. Spatiotemporal Regulation of PEDF Signaling by Type I Collagen Remodeling. Proc. Natl. Acad. Sci. USA 2020, 117, 11450–11458. [Google Scholar] [CrossRef]
- Jalan, A.A.; Sammon, D.; Hartgerink, J.D.; Brear, P.; Stott, K.; Hamaia, S.W.; Hunter, E.J.; Walker, D.R.; Leitinger, B.; Farndale, R.W. Chain Alignment of Collagen I Deciphered Using Computationally Designed Heterotrimers. Nat. Chem. Biol. 2020, 16, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, S.A.; Harsch, M.; Murphy, L.; Rosenbloom, J. Effects of Temperature on Conformation, Hydroxylation, and Secretion of Chick Tendon Procollagen. J. Biol. Chem. 1974, 249, 4480–4486. [Google Scholar] [CrossRef]
- Murphy, L.; Rosenbloom, J. Evidence That Chick Tendon Procollagen Must Be Denatured to Serve as Substrate for Proline Hydroxylase. Biochem. J. 1973, 135, 249–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg, R.A.; Prockop, D.J. Purification of (14C) Protocollagen and Its Hydroxylation by Prolyl-Hydroxylase. Biochemistry 1973, 12, 3395–3401. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.G.; Phamluong, K.; Li, L.; Sun, M.; Cao, T.C.; Liu, P.S.; Modrusan, Z.; Sandoval, W.N.; Rangell, L.; Carano, R.A.D.; et al. Global Defects in Collagen Secretion in a Mia3/TANGO1 Knockout Mouse. J. Cell Biol. 2011, 193, 935–951. [Google Scholar] [CrossRef] [Green Version]
- Saito, K.; Chen, M.; Bard, F.; Chen, S.; Zhou, H.; Woodley, D.; Polischuk, R.; Schekman, R.; Malhotra, V. TANGO1 Facilitates Cargo Loading at Endoplasmic Reticulum Exit Sites. Cell 2009, 136, 891–902. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.; Kenny, S.J.; Hemmati, J.; Xu, K.; Schekman, R. TANGO1 and SEC12 Are Copackaged with Procollagen I to Facilitate the Generation of Large COPII Carriers. Proc. Natl. Acad. Sci. USA 2018, 115, E12255–E12264. [Google Scholar] [CrossRef] [Green Version]
- McCaughey, J.; Stevenson, N.L.; Cross, S.; Stephens, D.J. ER-to-Golgi Trafficking of Procollagen in the Absence of Large Carriers. J. Cell Biol. 2019, 218, 929–948. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, Y.; Ito, S.; Nagata, K.; Sakai, L.Y.; Bächinger, H.P. Intracellular Mechanisms of Molecular Recognition and Sorting for Transport of Large Extracellular Matrix Molecules. Proc. Natl. Acad. Sci. USA 2016, 113, E6036–E6044. [Google Scholar] [CrossRef] [Green Version]
- Torre-Blanco, A.; Adachi, E.; Hojima, Y.; Wootton, J.A.; Minor, R.R.; Prockop, D.J. Temperature-Induced Post-Translational Over-Modification of Type I Procollagen. Effects of Over-Modification of the Protein on the Rate of Cleavage by Procollagen N-Proteinase and on Self-Assembly of Collagen into Fibrils. J. Biol. Chem. 1992, 267, 2650–2655. [Google Scholar] [CrossRef]
- Kimura, S.; Zhu, X.-P.; Matsui, R.; Shijoh, M.; Takamizawa, S. Characterization of Fish Muscle Type I Collagen. J. Food Sci. 1988, 53, 1315–1318. [Google Scholar] [CrossRef]
- Saito, M.; Higuchi, T.; Uchida, N. Identification and Primary Structures of Eel Type I Collagen proα1, proα2 and proα3. Fish. Sci. 2014, 80, 1323–1335. [Google Scholar] [CrossRef]
- Saito, M.; Takenouchi, Y.; Kunisaki, N.; Kimura, S. Complete Primary Structure of Rainbow Trout Type I Collagen Consisting of α1(I)α2(I)α3(I) Heterotrimers. Eur. J. Biochem. 2001, 268, 2817–2827. [Google Scholar] [CrossRef]
- Duan, R.; Zhang, J.; Du, X.; Yao, X.; Konno, K. Properties of Collagen from Skin, Scale and Bone of Carp (Cyprinus carpio). Food Chem. 2009, 112, 702–706. [Google Scholar] [CrossRef]
- Notbohm, H.; Mosler, S.; Bodo, M.; Yang, C.; Lehmann, H.; Bätge, B.; Müller, P.K. Comparative Study on the Thermostability of Collagen I of Skin and Bone: Influence of Posttranslational Hydroxylation of Prolyl and Lysyl Residues. J. Protein Chem. 1992, 11, 635–643. [Google Scholar] [CrossRef]
- Jimenez, S.; Bashey, R. Identification of Collagen α1(I) Trimer in Embryonic Chick Tendons and Calvaria. Biochem. Biophys. Res. Commun. 1977, 78, 1354–1361. [Google Scholar] [CrossRef]
- Makareeva, E.; Han, S.; Vera, J.C.; Sackett, D.L.; Holmbeck, K.; Phillips, C.L.; Visse, R.; Nagase, H.; Leikin, S. Carcinomas Contain a Matrix Metalloproteinase-Resistant Isoform of Type I Collagen Exerting Selective Support to Invasion. Cancer Res. 2010, 70, 4366–4374. [Google Scholar] [CrossRef] [Green Version]
- Moro, L.; Smith, B.D. Identification of Collagen α1(I) Trimer and Normal Type I Collagen in a Polyoma Virus-Induced Mouse Tumor. Arch. Biochem. Biophys. 1977, 182, 33–41. [Google Scholar] [CrossRef]
- Shapiro, F.D.; Eyre, D.R. Collagen Polymorphism in Extracellular Matrix of Human Osteosarcoma. J. Natl. Cancer Inst. 1982, 69, 1009–1016. [Google Scholar] [CrossRef]
- Yamagata, S.; Yamagata, T. FBJ Virus-Induced Osteosarcoma Contains Type I, Type I Trimer, Type III as Well as Type V Collagens. J. Biochem. 1984, 96, 17–26. [Google Scholar] [CrossRef]
- Minafra, I.P.; Luparello, C.; Sciarrino, S.; Tomasino, R.M.; Minafra, S. Quantitative Determination of Collagen Types Present in the Ductal Infiltrating Carcinoma of Human Mammary Gland. Cell Biol. Int. Rep. 1985, 9, 291–296. [Google Scholar] [CrossRef]
- Kuznetsova, N.V.; McBride, D.J.; Leikin, S. Changes in Thermal Stability and Microunfolding Pattern of Collagen Helix Resulting from the Loss of α2(I) Chain in Osteogenesis Imperfecta Murine. J. Mol. Biol. 2003, 331, 191–200. [Google Scholar] [CrossRef]
- Miles, C.A.; Sims, T.J.; Camacho, N.P.; Bailey, A.J. The Role of the α2 Chain in the Stabilization of the Collagen Type I Heterotrimer: A Study of the Type I Homotrimer in oim Mouse Tissues. J. Mol. Biol. 2002, 321, 797–805. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Lawson, R.N.; Chughtai, M.S. Breast Cancer and Body Temperature. Can. Med. Assoc. J. 1963, 88, 68–70. [Google Scholar]
- Ishikawa, Y.; Taga, Y.; Zientek, K.; Mizuno, N.; Salo, A.M.; Semenova, O.; Tufa, S.F.; Keene, D.R.; Holden, P.; Mizuno, K.; et al. Type I and Type V Procollagen Triple Helix Uses Different Subsets of the Molecular Ensemble for Lysine Posttranslational Modifications in the rER. J. Biol. Chem. 2021, 296, 100453. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture Temperature | (α1)3 | [(α1)2α3] | [(α1)2α2] | [α1α2α3] | (α1)3 + [(α1)2α3] | (α1)3 + [(α1)2α2] |
|---|---|---|---|---|---|---|
| Average (SD) (%) | Average (SD) (%) | Average (SD) (%) | Average (SD) (%) | Average (SD) (%) | Average (SD) (%) | |
| 18 °C | 15.1 (3.1) − x max: 6.1 (x = 9.0) min: 0 (x = 15.1) | x max: 15.1 (x = 15.1) min: 9.0 (x = 9.0) | −9.0 (7.6) + x max: 6.1 (x = 15.1) min: 0 (x = 9.0) | 93.9 (5.4) − x max: 84.6 (x = 9.0) min: 78.5 (x = 15.1) | 15.1 (3.1) | 6.1 (5.4) |
| 23 °C | 9.4 (2.2) − y max: 9.4 (y = 0) min: 0 (y = 9.4) | y max: 9.4 (y = 9.4) min: 0 (y = 0) | 3.7 (4.5) + y max: 13.1 (y = 9.4) min: 3.7 (y = 0) | 86.9 (3.3) − y max: 86.9 (y = 0) min: 77.8 (y = 9.1) | 9.4 (2.2) | 13.1 (3.3) |
| 28 °C | 11.1 (3.5) − z max: 11.1 (z = 0) min: 0 (z = 11.1) | z max: 11.1 (z = 11.1) min: 0 (z = 0) | 2.4 (11.2) + z max: 13.5 (z = 11.1) min: 2.4 (z = 0) | 86.4 (7.8) − z max: 86.4 (z = 0) min: 75.3 (z = 11.1) | 11.1 (3.5) | 13.6 (7.8) |
| 33 °C | 29.9 (8.2) − w max: 29.9 (w = 0) min: 0 (w = 29.9) | w max: 29.9 (w = 29.9) min: 0 (w = 0) | 4.9 (15.4) + w max: 34.8 (w = 29.9) min: 4.9 (w = 0) | 65.2 (7.3) − w max: 65.2 (w = 0) min: 35.3 (w = 29.9) | 29.9 (8.2) | 34.8 (7.3) |
| α-Chain Composition | Chain Stagger | Virtual Relative Thermal Stability (°C) |
|---|---|---|
| (α1)3 | α1α1α1 | 40.7 |
| [(α1)2α3] | α1α1α3 | 39.9 |
| α3α1α1 | 39.7 | |
| α1α3α1 | 39.3 | |
| [(α1)2α2] | α2α1α1 | 39.0 |
| α1α1α2 | 38.8 | |
| [α1α2α3] | α2α3α1 | 38.0 |
| α2α1α3 | 37.9 | |
| α1α3α2 | 37.7 | |
| α3α1α2 | 37.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujii, K.K.; Taga, Y.; Takagi, Y.K.; Masuda, R.; Hattori, S.; Koide, T. The Thermal Stability of the Collagen Triple Helix Is Tuned According to the Environmental Temperature. Int. J. Mol. Sci. 2022, 23, 2040. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042040
Fujii KK, Taga Y, Takagi YK, Masuda R, Hattori S, Koide T. The Thermal Stability of the Collagen Triple Helix Is Tuned According to the Environmental Temperature. International Journal of Molecular Sciences. 2022; 23(4):2040. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042040
Chicago/Turabian StyleFujii, Kazunori K., Yuki Taga, Yusuke K. Takagi, Ryo Masuda, Shunji Hattori, and Takaki Koide. 2022. "The Thermal Stability of the Collagen Triple Helix Is Tuned According to the Environmental Temperature" International Journal of Molecular Sciences 23, no. 4: 2040. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042040