
NGF/TRKA Promotes ADAM17-Dependent Cleavage of P75 in Ovarian Cells: Elucidating a Pro-Tumoral Mechanism

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
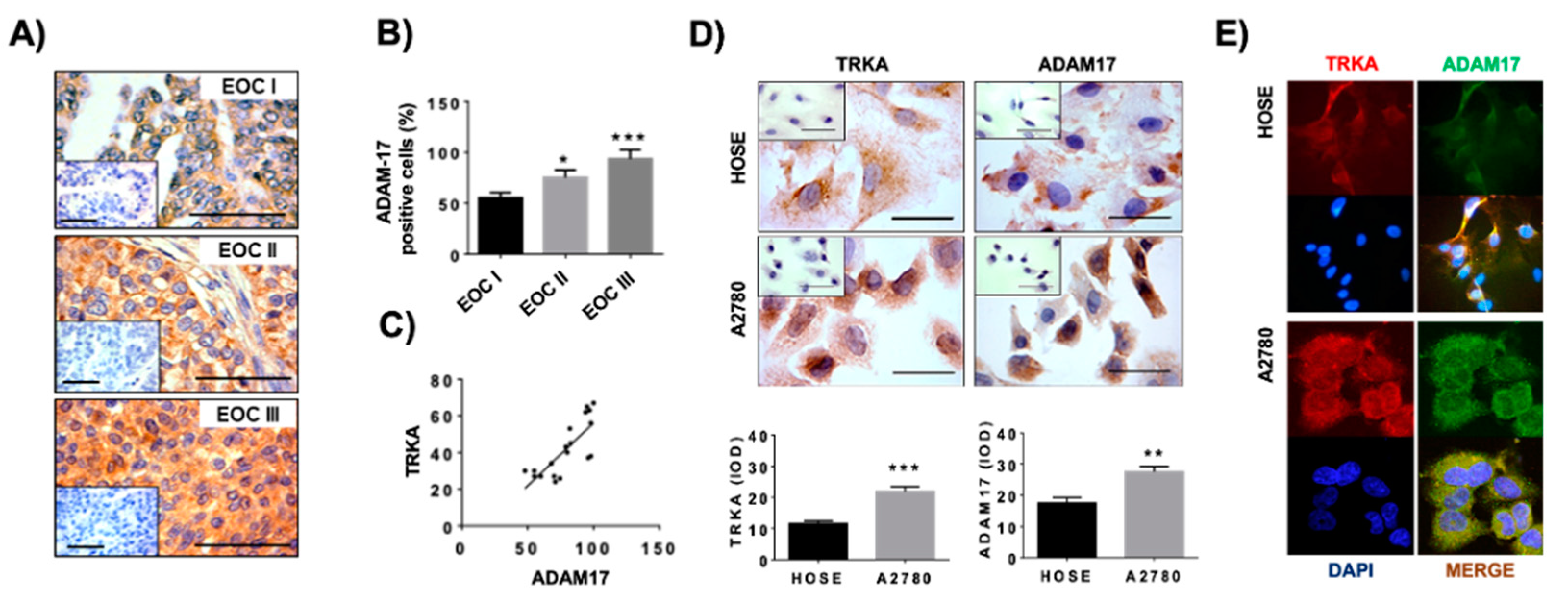
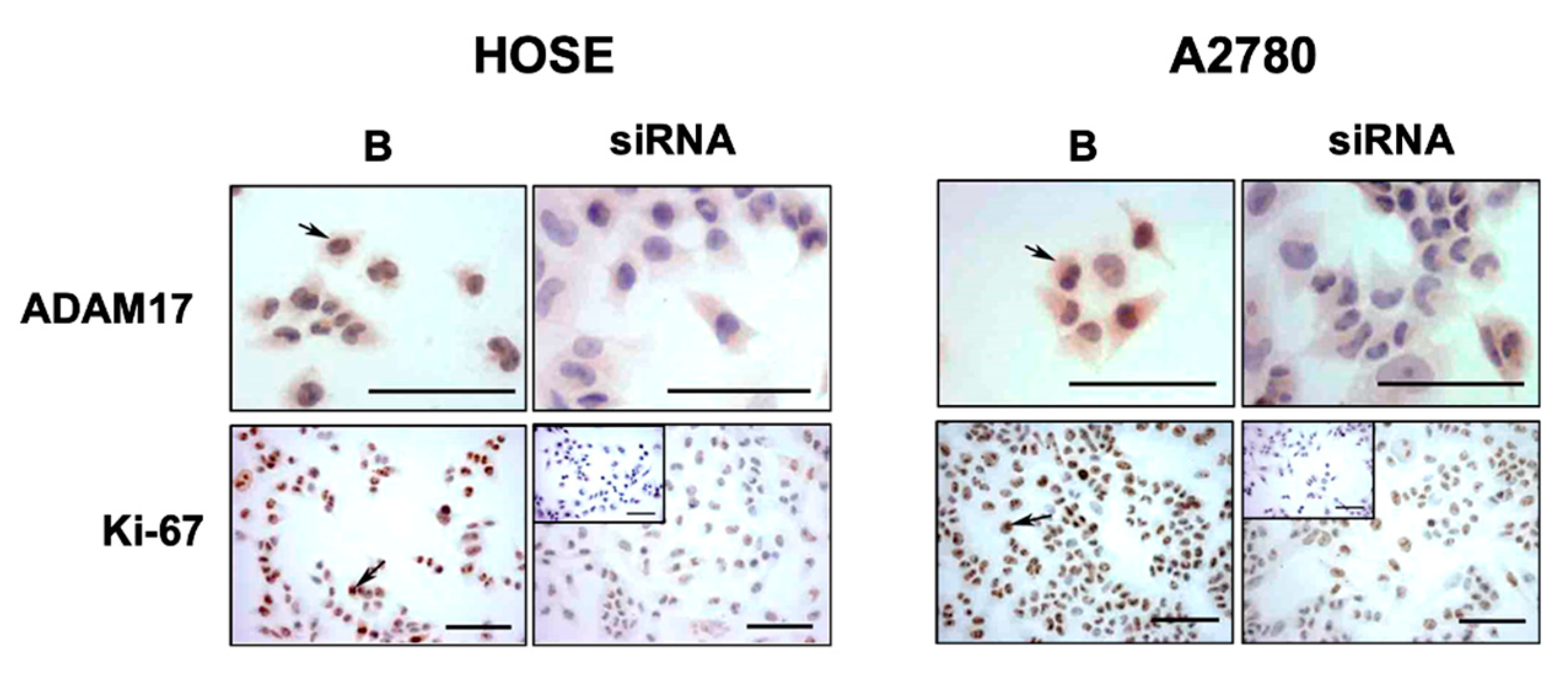
2.1. Detection of ADAM17 and TRKA in Epithelial Ovarian Cancer Biopsies and Ovarian Cells
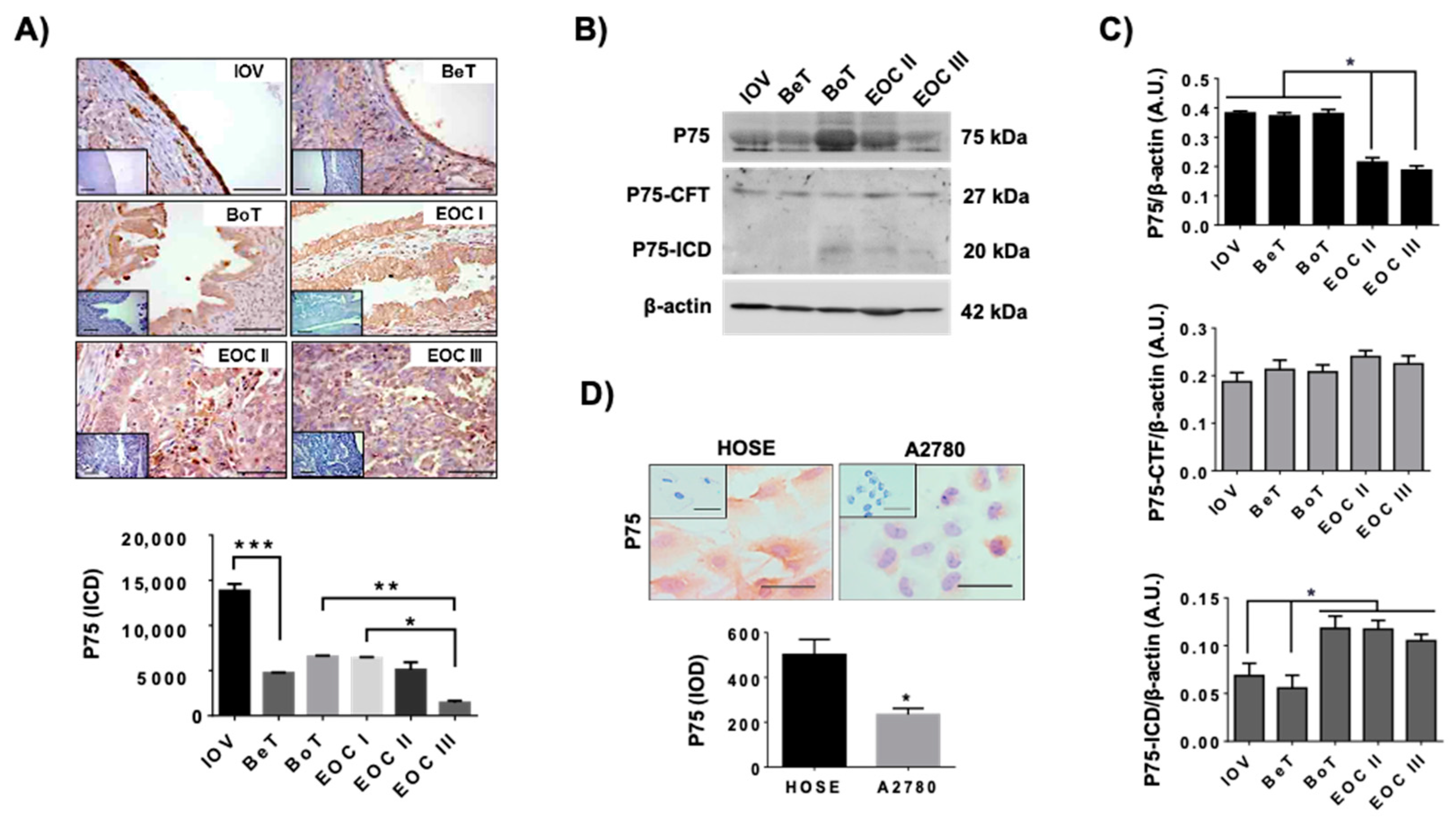
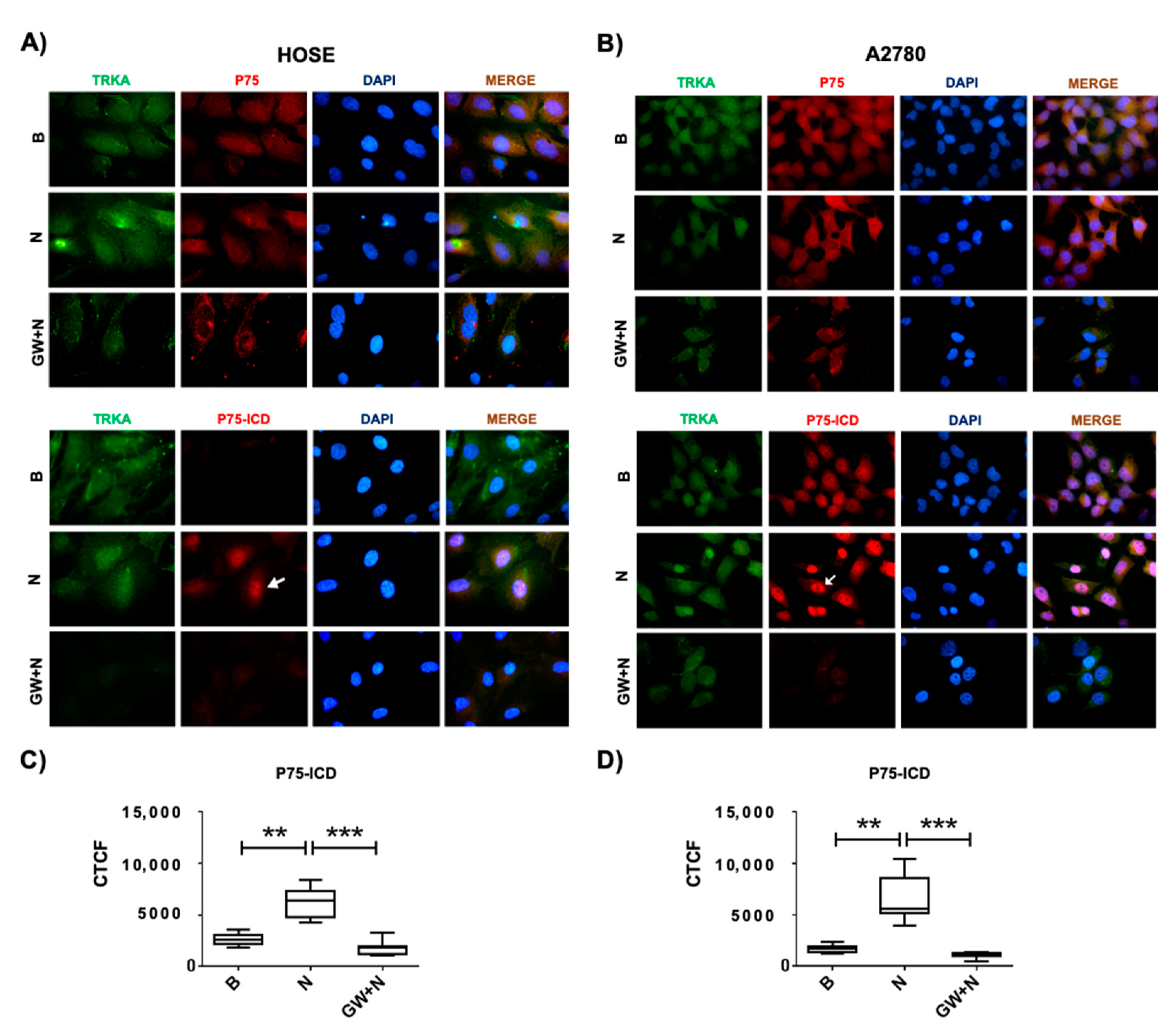
2.2. Levels of P75 during EOC Progression
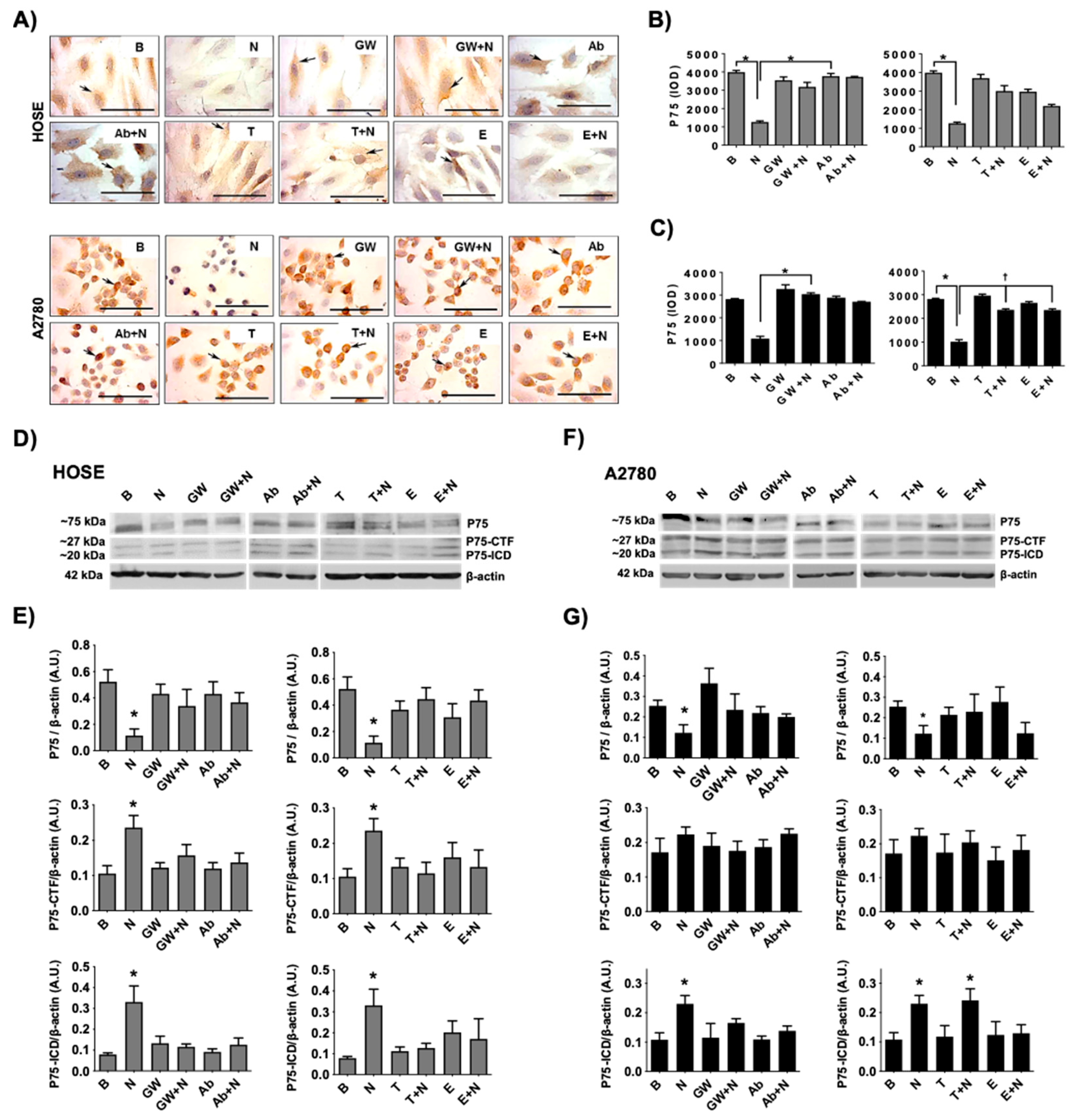
2.3. Levels of P75 and Its Fragments by NGF/TRKA in Human Ovarian Cell Lines
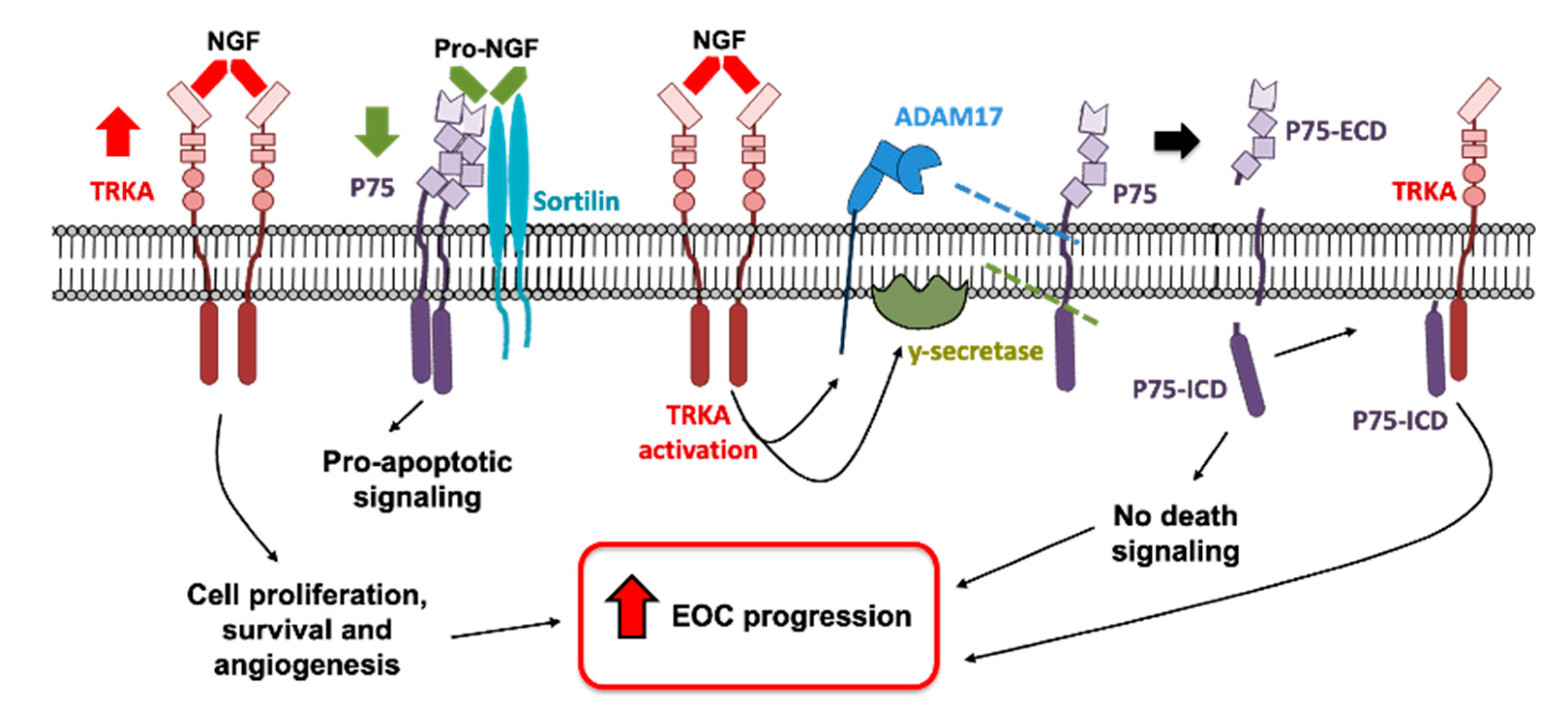
3. Discussion
4. Materials and Methods
4.1. Tissue Samples
4.2. Cell Culture and Treatments
4.3. Antibodies
4.4. Western Blotting
4.5. Immunohistochemistry
4.6. Immunocytochemistry
4.7. Statistical Evaluation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- American Cancer Society. Survival Rates for Ovarian Cancer. Available online: https://www.cancer.org/cancer/ovarian-cancer/detection-diagnosis-staging/survival-rates.html (accessed on 25 November 2021).
- Cancer Research, UK. Epithelial Ovarian Cancer. Available online: https://www.cancerresearchuk.org/about-cancer/ovarian-cancer/types/epithelial-ovarian-cancers/epithelial (accessed on 25 November 2021).
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Tapia, V.; Gabler, F.; Muñoz, M.; Yazigi, R.; Paredes, A.; Selman, A.; Vega, M.; Romero, C. Tyrosine kinase A receptor (trkA): A potential marker in epithelial ovarian cancer. Gynecol. Oncol. 2011, 121, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Campos, X.; Muñoz, Y.; Selman, A.; Yazigi, R.; Moyano, L.; Weinstein-Oppenheimer, C.; Lara, H.E.; Romero, C. Nerve growth factor and its high-affinity receptor trkA participate in the control of vascular endothelial growth factor expression in epithelial ovarian cancer. Gynecol. Oncol. 2007, 104, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Urzua, U.; Tapia, V.; Geraldo, M.P.; Selman, A.; Vega, M.; Romero, C. Nerve Growth Factor Stimulates Cellular Proliferation of Human Epithelial Ovarian Cancer. Horm. Metab. Res. 2012, 44, 656–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido, M.P.; Torres, I.; Vega, M.; Romero, C. Angiogenesis in Gynecological Cancers: Role of Neurotrophins. Front. Oncol. 2019, 9, 913. [Google Scholar] [CrossRef] [PubMed]
- Garrido, M.P.; Hurtado, I.; Valenzuela-Valderrama, M.; Salvatierra, R.; Hernández, A.; Vega, M.; Selman, A.; Quest, A.F.G.; Romero, C. NGF-Enhanced Vasculogenic Properties of Epithelial Ovarian Cancer Cells Is Reduced by Inhibition of the COX-2/PGE2 Signaling Axis. Cancers 2019, 11, 1970. [Google Scholar] [CrossRef] [Green Version]
- Garrido, M.P.; Torres, I.; Avila, A.; Chnaiderman, J.; Valenzuela-Valderrama, M.; Aramburo, J.; Oróstica, L.; Durán-Jara, E.; Lobos-Gonzalez, L.; Romero, C. NGF/TRKA Decrease miR-145-5p Levels in Epithelial Ovarian Cancer Cells. Int. J. Mol. Sci. 2020, 21, 7657. [Google Scholar] [CrossRef]
- Ichim, G.; Tauszig-Delamasure, S.; Mehlen, P. Neurotrophins and cell death. Exp. Cell Res. 2012, 318, 1221–1228. [Google Scholar] [CrossRef]
- Bassili, M.; Birman, E.; Schor, N.F.; Saragovi, H.U. Differential roles of Trk and P75 neurotrophin receptors in tumorigenesis and chemoresistance ex vivo and in vivo. Cancer Chemother. Pharmacol. 2010, 65, 1047–1056. [Google Scholar] [CrossRef] [Green Version]
- Vera, C.; Tapia, V.; Vega, M.; Romero, C. Role of nerve growth factor and its TRKA receptor in normal ovarian and epithelial ovarian cancer angiogenesis. J. Ovarian Res. 2014, 7, 82. [Google Scholar] [CrossRef]
- Heuer, J.G.; Fatemie-Nainie, S.; Wheeler, E.F.; Bothwell, M. Structure and developmental expression of the chicken NGF receptor. Dev. Biol. 1990, 137, 287–304. [Google Scholar] [CrossRef]
- Reichardt, L.F. Neurotrophin-regulated signalling pathways. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1545–1564. [Google Scholar] [CrossRef] [Green Version]
- Nykjaer, A.; Lee, R.; Teng, K.K.; Jansen, P.; Madsen, P.S.; Nielsen, M.S.; Jacobsen, C.; Kliemannel, M.; Schwarz, E.; Willnow, T.E.; et al. Sortilin is essential for proNGF-induced neuronal cell death. Nature 2004, 427, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Teng, H.K.; Teng, K.K.; Lee, R.; Wright, S.; Tevar, S.; Almeida, R.D.; Kermani, P.; Torkin, R.; Chen, Z.-Y.; Lee, F.S.; et al. ProBDNF Induces Neuronal Apoptosis via Activation of a Receptor Complex of P75NTR and Sortilin. J. Neurosci. 2005, 25, 5455–5463. [Google Scholar] [CrossRef]
- Khwaja, F.; Tabassum, A.; Allen, J.; Djakiew, D. The P75NTR tumor suppressor induces cell cycle arrest facilitating caspase mediated apoptosis in prostate tumor cells. Biochem. Biophys. Res. Commun. 2006, 341, 1184–1192. [Google Scholar] [CrossRef] [PubMed]
- Gravina, G.L.; Marampon, F.; Sanità, P.; Mancini, A.; Colapietro, A.; Scarsella, L.; Jitariuc, A.; Biordi, L.; Ficorella, C.; Festuccia, C. Increased expression and activity of P75NTR are crucial events in azacitidine-induced cell death in prostate cancer. Oncol. Rep. 2016, 36, 125–130. [Google Scholar] [CrossRef]
- Verbeke, S.; Meignan, S.; Lagadec, C.; Germain, E.; Hondermarck, H.; Adriaenssens, E.; Le Bourhis, X. Overexpression of P75NTR increases survival of breast cancer cells through p21waf1. Cell. Signal. 2010, 22, 1864–1873. [Google Scholar] [CrossRef]
- Jin, H.; Pan, Y.; Zhao, L.; Zhai, H.; Li, X.; Sun, L.; He, L.; Chen, Y.; Hong, L.; Du, Y.; et al. P75 Neurotrophin Receptor Suppresses the Proliferation of Human Gastric Cancer Cells. Neoplasia 2007, 9, 471–478. [Google Scholar] [CrossRef] [Green Version]
- Festuccia, C.; Gravina, G.L.; Muzi, P.; Pomante, R.; Ventura, L.; Ricevuto, E.; Vicentini, C.; Bologna, M. In vitro and in vivo effects of bicalutamide on the expression of TrkA and P75 neurotrophin receptors in prostate carcinoma. Prostate 2007, 67, 1255–1264. [Google Scholar] [CrossRef]
- Yoon, S.O.; Casaccia-Bonnefil, P.; Carter, B.; Chao, M.V. Competitive Signaling Between TrkA and P75 Nerve Growth Factor Receptors Determines Cell Survival. J. Neurosci. 1998, 18, 3273–3281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
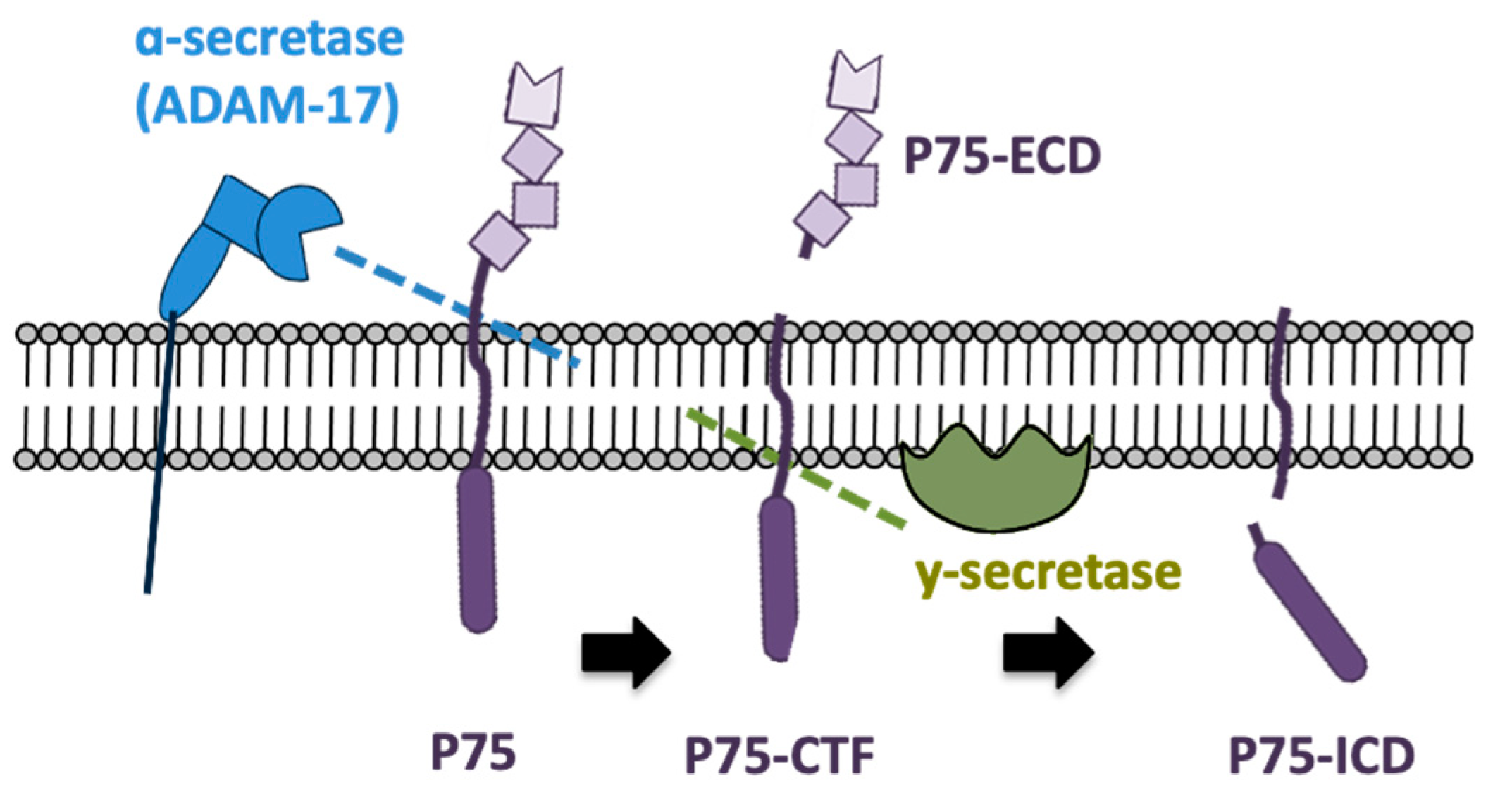
- Urra, S.; Escudero, C.A.; Ramos, P.; Lisbona, F.; Allende, E.; Covarrubias, P.; Parraguez, J.I.; Zampieri, N.; Chao, M.V.; Annaert, W.; et al. TrkA Receptor Activation by Nerve Growth Factor Induces Shedding of the P75 Neurotrophin Receptor Followed by Endosomal γ-Secretase-mediated Release of the P75 Intracellular Domain. J. Biol. Chem. 2007, 282, 7606–7615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, K.-M.; Tan, S.; Landman, N.; Petrova, K.; Murray, S.; Lewis, R.; Kim, P.K.; Kim, D.S.; Ryu, S.H.; Chao, M.V.; et al. Regulated Intramembrane Proteolysis of the P75 Neurotrophin Receptor Modulates Its Association with the TrkA Receptor. J. Biol. Chem. 2003, 278, 42161–42169. [Google Scholar] [CrossRef] [Green Version]
- Bronfman, F.C. Metalloproteases and γ-secretase: New membrane partners regulating P75 neurotrophin receptor signaling? J. Neurochem. 2007, 103, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Ni, S.-S.; Zhang, J.; Zhao, W.-L.; Dong, X.-C.; Wang, J.-L. ADAM17 is overexpressed in non-small cell lung cancer and its expression correlates with poor patient survival. Tumor Biol. 2013, 34, 1813–1818. [Google Scholar] [CrossRef] [PubMed]
- McGowan, P.M.; Ryan, B.M.; Hill, A.D.; McDermott, E.; O’Higgins, N.; Duffy, M.J. ADAM-17 Expression in Breast Cancer Correlates with Variables of Tumor Progression. Clin. Cancer Res. 2007, 13, 2335–2343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, X.; Yang, L.-Y.; Huang, G.-W.; Wang, W.; Lu, W.-Q. ADAM17 mRNA expression and pathological features of hepatocellular carcinoma. World J. Gastroenterol. 2004, 10, 2735–2739. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Ying, M.; Chen, G.; Lin, A.; Xie, Y.; Ohara, N.; Zhou, D. ADAM17 is associated with EMMPRIN and predicts poor prognosis in patients with uterine cervical carcinoma. Tumor Biol. 2014, 35, 7575–7586. [Google Scholar] [CrossRef]
- Aydin, D.; Bilici, A.; Yavuzer, D.; Kefeli, U.; Tan, A.; Ercelep, O.; Mert, A.; Yuksel, S.; Ozcelik, M.; Isik, D.; et al. Prognostic significance of ADAM17 expression in patients with gastric cancer who underwent curative gastrectomy. Clin. Transl. Oncol. 2015, 17, 604–611. [Google Scholar] [CrossRef]
- Rogmans, C.; Kuhlmann, J.D.; Hugendieck, G.; Link, T.; Arnold, N.; Weimer, J.P.; Flörkemeier, I.; Rambow, A.-C.; Lieb, W.; Maass, N.; et al. ADAM17—A Potential Blood-Based Biomarker for Detection of Early-Stage Ovarian Cancer. Cancers 2021, 13, 5563. [Google Scholar] [CrossRef]
- Hedemann, N.; Rogmans, C.; Sebens, S.; Wesch, D.; Reichert, M.; Schmidt-Arras, D.; Oberg, H.-H.; Pecks, U.; Van Mackelenbergh, M.; Weimer, J.; et al. ADAM17 inhibition enhances platinum efficiency in ovarian cancer. Oncotarget 2018, 9, 16043–16058. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, C.; Clementi, M.; Benitez, D.; Contreras, H.; Huidobro, C.; Castellón, E. Effect of GnRH analogs on the expression of TrkA and P75 neurotrophin receptors in primary cell cultures from human prostate adenocarcinoma. Prostate 2005, 65, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Tan, F.; Thiele, C.J.; Li, Z. Neurotrophin Signaling in Cancer. In Handbook of Neurotoxicity; Kostrzewa, R.M., Ed.; Springer: New York, NY, USA, 2014; pp. 1825–1847. [Google Scholar]
- Garrido, M.P.; Bruneau, N.; Vega, M.; Selman, A.; Tapia, J.C.; Romero, C. Follicle-stimulating hormone promotes nerve growth factor and vascular endothelial growth factor expression in epithelial ovarian cells. Histol Histopathol 2020, 35, 961–971. [Google Scholar] [CrossRef]
- Kanning, K.C.; Hudson, M.; Amieux, P.S.; Wiley, J.C.; Bothwell, M.; Schecterson, L.C. Proteolytic Processing of the P75 Neurotrophin Receptor and Two Homologs Generates C-Terminal Fragments with Signaling Capability. J. Neurosci. 2003, 23, 5425–5436. [Google Scholar] [CrossRef] [Green Version]
- Soslow, R.A. Histologic Subtypes of Ovarian Carcinoma: An overview. Int. J. Gynecol. Pathol. 2008, 27, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Khwaja, F.; Djakiew, D. Inhibition of cell-cycle effectors of proliferation in bladder tumor epithelial cells by the P75NTR tumor suppressor. Mol. Carcinog. 2003, 36, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Descamps, S.; Toillon, R.-A.; Adriaenssens, E.; Pawlowski, V.; Cool, S.M.; Nurcombe, V.; Le Bourhis, X.; Boilly, B.; Peyrat, J.-P.; Hondermarck, H. Nerve Growth Factor Stimulates Proliferation and Survival of Human Breast Cancer Cells through Two Distinct Signaling Pathways. J. Biol. Chem. 2001, 276, 17864–17870. [Google Scholar] [CrossRef] [Green Version]
- EL Yazidi-Belkoura, I.; Adriaenssens, E.; Dollé, L.; Descamps, S.; Hondermarck, H. Tumor Necrosis Factor Receptor-associated Death Domain Protein Is Involved in the Neurotrophin Receptor-mediated Antiapoptotic Activity of Nerve Growth Factor in Breast Cancer Cells. J. Biol. Chem. 2003, 278, 16952–16956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkhurst, C.N.; Zampieri, N.; Chao, M.V. Nuclear Localization of the P75 Neurotrophin Receptor Intracellular Domain. J. Biol. Chem. 2010, 285, 5361–5368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, B.; Lazarovici, P.; Ezersky, A.; Nesland, J.M.; Berner, A.; Risberg, B.; Tropé, C.G.; Kristensen, G.B.; Goscinski, M.; Van De Putte, G.; et al. Expression levels of the nerve growth factor receptors TrkA and P75 in effusions and solid tumors of serous ovarian carcinoma patients. Clin. Cancer Res. 2001, 7, 3457–3464. [Google Scholar] [PubMed]
- Matusica, D.; Skeldal, S.; Sykes, A.M.; Palstra, N.; Sharma, A.; Coulson, E.J. An Intracellular Domain Fragment of the P75 Neurotrophin Receptor (P75NTR) Enhances Tropomyosin Receptor Kinase A (TrkA) Receptor Function. J. Biol. Chem. 2013, 288, 11144–11154. [Google Scholar] [CrossRef] [Green Version]
- Skeldal, S.; Matusica, D.; Nykjaer, A.; Coulson, E.J. Proteolytic processing of the P75 neurotrophin receptor: A prerequisite for signalling? Neuronal life, growth and death signalling are crucially regulated by intra-membrane proteolysis and trafficking of P75(NTR). BioEssays 2011, 33, 614–625. [Google Scholar] [CrossRef] [PubMed]
- Zanin, J.P.; Montroull, L.E.; Volosin, M.; Friedman, W.J. The P75 Neurotrophin Receptor Facilitates TrkB Signaling and Function in Rat Hippocampal Neurons. Front. Cell. Neurosci. 2019, 13, 485. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-D.; Yuan, Y.; Liu, X.-H.; Gong, D.-J.; Bai, C.-G.; Wang, F.; Luo, J.-H.; Xu, Z.-Y. Self-renewal and chemotherapy resistance of P75NTR positive cells in esophageal squamous cell carcinomas. BMC Cancer 2009, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Wislet, S.; Vandervelden, G.; Rogister, B. From Neural Crest Development to Cancer and Vice Versa: How P75NTR and (Pro)neurotrophins Could Act on Cell Migration and Invasion? Front. Mol. Neurosci. 2018, 11, 244. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dang, C.; Ma, Q.; Shimahara, Y. Expression of nerve growth factor receptors and their prognostic value in human pancreatic cancer. Oncol. Rep. 2005, 14, 161–171. [Google Scholar] [PubMed]
- Krygier, S.; Djakiew, D. Molecular characterization of the loss of P75NTR expression in human prostate tumor cells. Mol. Carcinog. 2001, 31, 46–55. [Google Scholar] [CrossRef]
- Davidson, B.; Reich, R.; Lazarovici, P.; Flørenes, V.A.; Nielsen, S.; Nesland, J.M. Altered Expression and Activation of the Nerve Growth Factor Receptors TrkA and P75 Provide the First Evidence of Tumor Progression to Effusion in Breast Carcinoma. Breast Cancer Res. Treat. 2004, 83, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Moss, M.L.; Minond, D. Recent Advances in ADAM17 Research: A Promising Target for Cancer and Inflammation. Mediat. Inflamm. 2017, 2017, 9673537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, F.M.; Tape, C.J.; Jodrell, D.I.; Murphy, G. Anti-Tumour Effects of a Specific Anti-ADAM17 Antibody in an Ovarian Cancer Model In Vivo. PLoS ONE 2012, 7, e40597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedemann, N.; Herz, A.; Schiepanski, J.H.; Dittrich, J.; Sebens, S.; Dempfle, A.; Feuerborn, J.; Rogmans, C.; Tribian, N.; Flörkemeier, I.; et al. ADAM17 Inhibition Increases the Impact of Cisplatin Treatment in Ovarian Cancer Spheroids. Cancers 2021, 13, 2039. [Google Scholar] [CrossRef]
- Charkhchi, P.; Cybulski, C.; Gronwald, J.; Wong, F.O.; Narod, S.A.; Akbari, M.R. CA125 and Ovarian Cancer: A Comprehensive Review. Cancers 2020, 12, 3730. [Google Scholar] [CrossRef]
- Daoud, E.; Bodor, G. CA-125 concentrations in malignant and nonmalignant disease. Clin. Chem. 1991, 37, 1968–1974. [Google Scholar] [CrossRef]
- Hwang, H.C.; Clurman, B.E. Cyclin E in normal and neoplastic cell cycles. Oncogene 2005, 24, 2776–2786. [Google Scholar] [CrossRef] [Green Version]
- Maines-Bandiera, S.L.; Kruk, P.A.; Auersperg, N. Simian virus 40-transformed human ovarian surface epithelial cells escape normal growth controls but retain morphogenetic responses to extracellular matrix. Am. J. Obstet. Gynecol. 1992, 167, 729–735. [Google Scholar] [CrossRef]
- Hamilton, T.C.; Young, R.C.; Ozols, R.F. Experimental model systems of ovarian cancer: Applications to the design and evaluation of new treatment approaches. Semin. Oncol. 1984, 11, 285–298. [Google Scholar]
- Beaufort, C.M.; Helmijr, J.C.A.; Piskorz, A.M.; Hoogstraat, M.; Ruigrok-Ritstier, K.; Besselink, N.; Murtaza, M.; van Ijcken, W.F.; Heine, A.A.; Smid, M.; et al. Ovarian Cancer Cell Line Panel (OCCP): Clinical Importance of In Vitro Morphological Subtypes. PLoS ONE 2014, 9, e103988. [Google Scholar] [CrossRef] [PubMed]
- Auersperg, N.; Pan, J.; Grove, B.D.; Peterson, T.; Fisher, J.; Maines-Bandiera, S.; Somasiri, A.; Roskelley, C.D. E-cadherin induces mesenchymal-to-epithelial transition in human ovarian surface epithelium. Proc. Natl. Acad. Sci. USA 1999, 96, 6249–6254. [Google Scholar] [CrossRef] [Green Version]
- Tudrej, P.; Kujawa, K.A.; Cortez, A.J.; Lisowska, K.M. Characteristics of in Vivo Model Systems for Ovarian Cancer Studies. Diagnostics 2019, 9, 120. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garrido, M.P.; Vallejos, C.; Girardi, S.; Gabler, F.; Selman, A.; López, F.; Vega, M.; Romero, C. NGF/TRKA Promotes ADAM17-Dependent Cleavage of P75 in Ovarian Cells: Elucidating a Pro-Tumoral Mechanism. Int. J. Mol. Sci. 2022, 23, 2124. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042124
Garrido MP, Vallejos C, Girardi S, Gabler F, Selman A, López F, Vega M, Romero C. NGF/TRKA Promotes ADAM17-Dependent Cleavage of P75 in Ovarian Cells: Elucidating a Pro-Tumoral Mechanism. International Journal of Molecular Sciences. 2022; 23(4):2124. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042124
Chicago/Turabian StyleGarrido, Maritza P., Christopher Vallejos, Silvanna Girardi, Fernando Gabler, Alberto Selman, Fernanda López, Margarita Vega, and Carmen Romero. 2022. "NGF/TRKA Promotes ADAM17-Dependent Cleavage of P75 in Ovarian Cells: Elucidating a Pro-Tumoral Mechanism" International Journal of Molecular Sciences 23, no. 4: 2124. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042124