
The Roles of NOD-like Receptors in Innate Immunity in Otitis Media

 , , and
, , and
Abstract
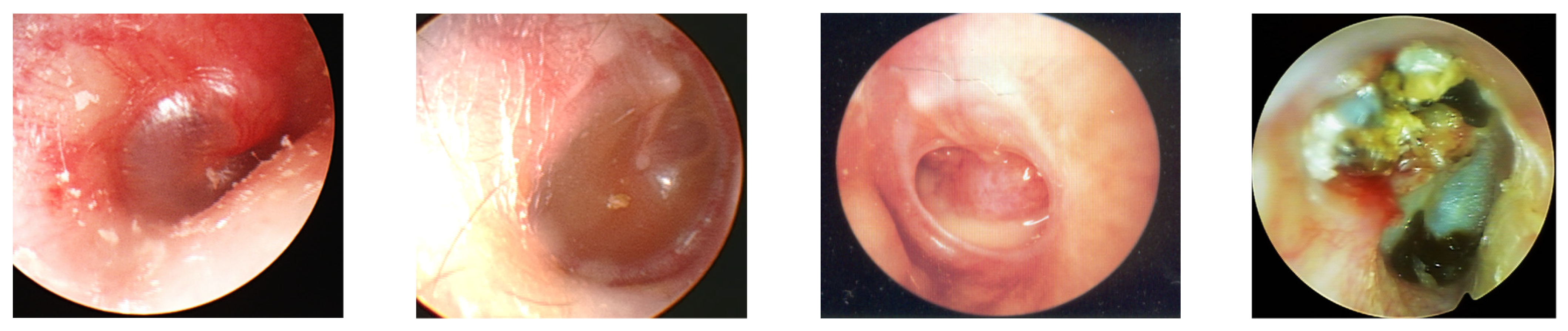
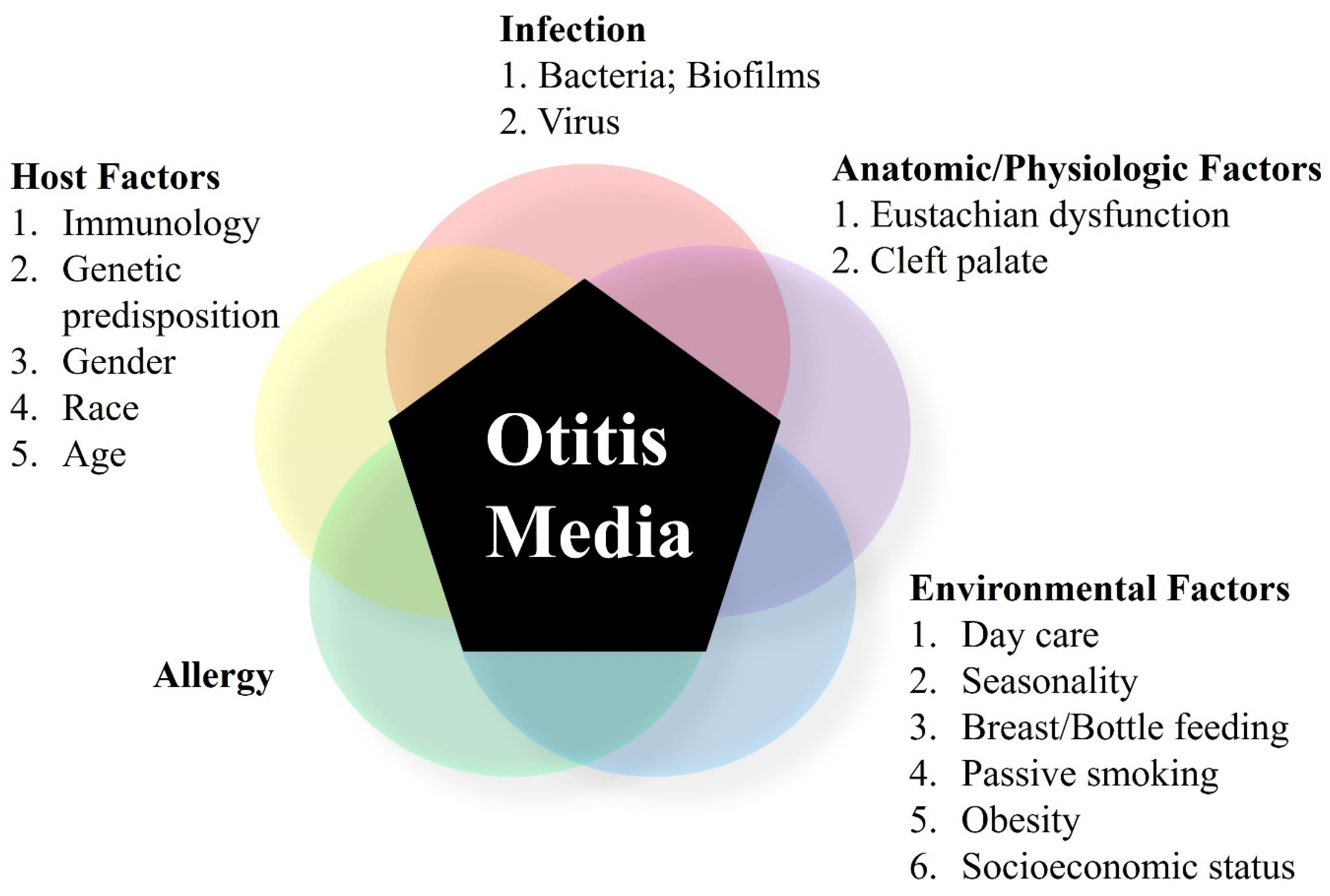
:1. Overview of Otitis Media
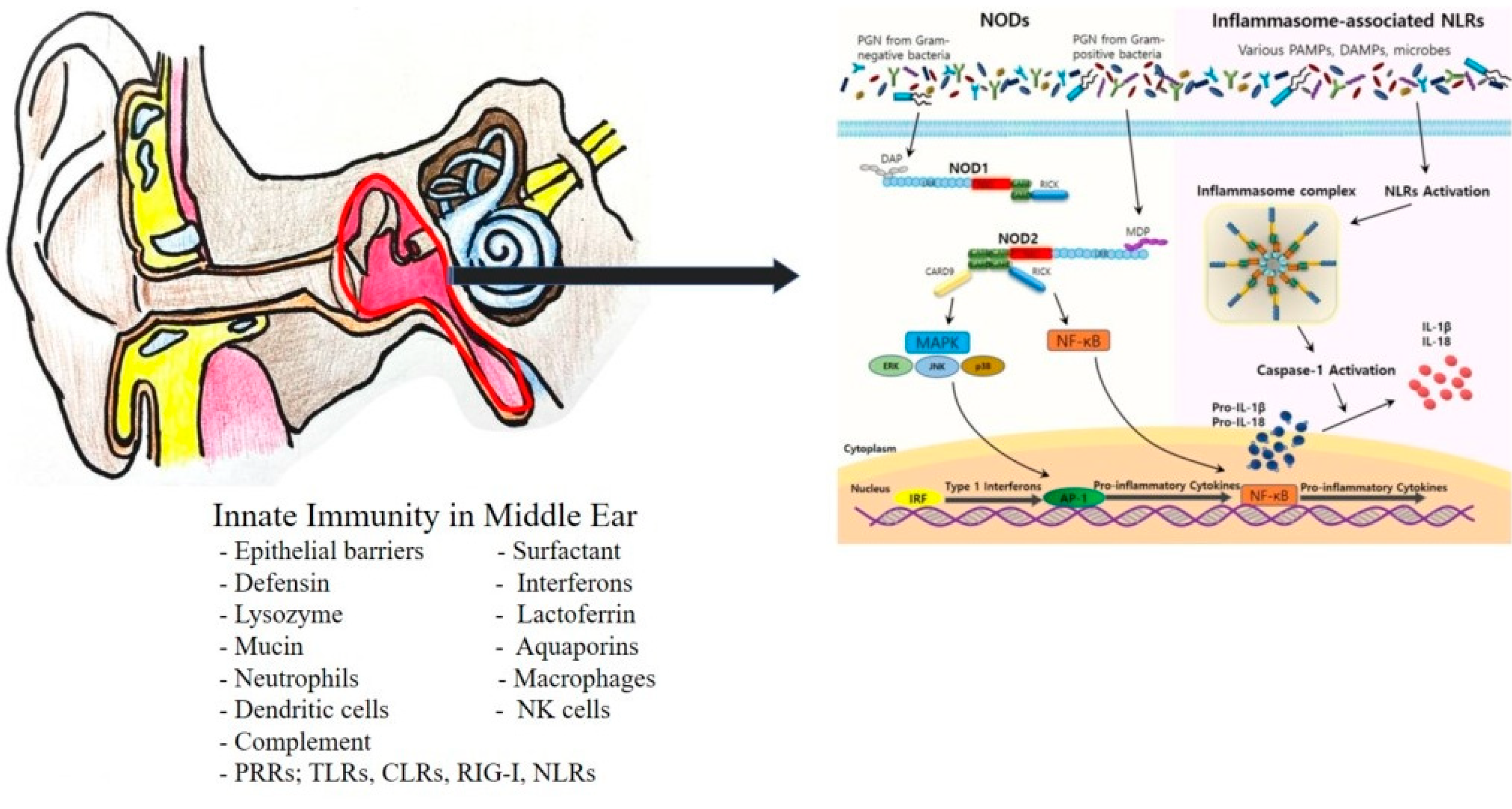
2. Innate Immunity in Otitis Media
3. NOD-like Receptors (NLRs) as Pattern-Recognition Receptors (PRRs)
4. NLRs and OM
- NLRA (A for acidic transactivating domain): CIITA
- NLRB (B for BIRs): NAIP
- NLRC (C for CARD): NOD1, NOD2, NLRC3, NLRC4, NLRC5
- NLRP (P for PYD): NLRP1, NLRP2, NLRP3, NLRP4, NLRP5, NLRP6, NLRP7, NLPR8, NLRP9, NLRP10, NLRP11, NLRP12, NLRP13, NLRP14
4.1. NLR1 and NLR2
4.2. NLRP3 (NLR Family Pyrin Domain Containing 3)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bluestone, C.D.; Klein, J.O. Otitis media and eustachian tube dysfunction. In Pediatric Otolaryngology, 4th ed.; Bluestone, C.D., Stool, S.E., Alper, C.M., Eds.; Saunders: Philadelphia, PA, USA, 2003; pp. 474–685. [Google Scholar]
- Rovers, M.M.; Schilder, A.G.; Zielhuis, G.A.; Rosenfeld, R.M. Otitis media. Lancet 2004, 363, 465–473. [Google Scholar] [CrossRef]
- Margaretha, L.C.; Ellen, M.M. Acute otitis media and otitis media with effusion. In Cummings Otolaryngology, 6th ed.; Paul, W.F., Bruce, H.H., Valerie, J.L., Robbins, J., Regan, T., Marci, L., Howard, W.F., Eds.; Elsevier: St Frisco, CO, USA, 2015; pp. 3019–3037. [Google Scholar]
- Yeo, S.G. Acute otitis media. In Korean Society of Otorhinolaryngology—Head and Neck Surgery, 3rd ed.; KoonJa: Seoul, Korea, 2018; pp. 363–383. [Google Scholar]
- Bluestone, C.D.; Alper, C.M.; Buchman, C.A.; Felding, J.U.; Ghadiali, S.N.; Hebda, P.A.; Sando, I.; Stangerup, S.E.; Swarts, J.D.; Takahashi, H. Recent advances in otitis media. Eustachian tube, middle ear, and mastoid anatomy, physiology, pathophysiology, and pathogenesis. Ann. Otol. Rhinol. Laryngol. 2005, 194, 16–30. [Google Scholar] [CrossRef]
- Hullegie, S.; Venekamp, R.P.; van Dongen, T.M.A.; Hay, A.D.; Moore, M.V.; Little, P.; Schilder, A.G.M.; Damoiseaux, R.A.M.J. Prevalence and Antimicrobial Resistance of Bacteria in Children with Acute Otitis Media and Ear Discharge: A Systematic Review. Pediatr. Infect. Dis. J. 2021, 40, 756–762. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, R.M.; Culpepper, L.; Yawn, B.; Mahoney, M.C. Otitis media with effusion clinical practice guideline. AAP, AAFP, AAO-HNS Subcommittee on Otitis Media with Effusion. Am. Fam. Physician 2004, 69, 2776–2779. [Google Scholar] [PubMed]
- Post, J.C.; Preston, R.A.; Aul, J.J.; Larkins-Pettigrew, M.; Rydquist-White, J.; Anderson, K.W.; Wadowsky, R.M.; Reagan, D.R.; Walker, E.S.; A Kingsley, L.; et al. Molecular analysis of bacterial pathogens in otitis media with effusion. JAMA 1995, 273, 1598–1604. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Tsuprun, V.; Kawano, H.; Paparella, M.M.; Zhang, Z.; Anway, R.; Ho, S.B. Characterization of mucins in human middle ear and Eustachian tube. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2001, 280, L1157–L1167. [Google Scholar] [CrossRef] [PubMed]
- Nokso-Koivisto, J.; Räty, R.; Blomqvist, S.; Kleemola, M.; Syrjänen, R.; Pitkäranta, A.; Kilpi, T.; Hovi, T. Presence of specific viruses in the middle ear fluids and respiratory secretions of young children with acute otitis media. J. Med Virol. 2004, 72, 241–248. [Google Scholar] [CrossRef]
- Scott-Brown’s Otorhinolaryngology, Head and Neck Surgery, 7th ed.; Gleeson, M.; Browning, G.G.; Burton, M.J.; Clark, R.; John, H.; Jones, N.S.; Lund, V.J.; Luxon, L.M.; Watkinson, J.C. (Eds.) Hodder Arnold: London, UK, 2007; Volume 3, pp. 3395–3452. [Google Scholar]
- Chole, R.A. Cellular and subcellular events of bone resorption in human and experimental cholesteatoma: The role of osteoclasts. Laryngoscope 1984, 94, 76–95. [Google Scholar] [CrossRef]
- van der Toom, H.F.E.; van der Schroeff, M.P.; Pauw, R.J. Single-Stage Mastoid Obliteration in Cholesteatoma Surgery and Recurrent and Residual Disease Rates: A Systematic Review. JAMA Otolaryngol. Head Neck Surg. 2018, 144, 440–446. [Google Scholar] [CrossRef]
- Budenz, C.L.; El-Kashlan, H.K.; Shelton, C.; Aygun, N.; Naparko, J.K. Complications of Temporal Bone Infection. In Cummings Otolaryngology Head and Neck Surgery, 6th ed.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 2160–2171. [Google Scholar]
- Lee, H.H. Role of Innate Immunity in Otitis Media. Korean J. Otorhinolaryngol.-Head Neck Surg. 2016, 59, 483–489. [Google Scholar] [CrossRef]
- Fornadley, J.A.; Burns, J.K. The effect of surfactant on eustachian tube function in a gerbil model of otitis media with effusion. Otolaryngol. Head Neck Surg. 1994, 110, 110–114. [Google Scholar] [CrossRef]
- Abdel-Razek, O.; Liu, T.; Chen, X.; Wang, Q.; Vanga, G.; Wang, G. Role of Surfactant Protein D in Experimental Otitis Media. J. Innate Immun. 2021, 13, 197–210. [Google Scholar] [CrossRef]
- Shin, D.J.; Gan-Undram, S.; Kim, S.J.; Jun, Y.J.; Im, G.J.; Jung, H.H. Expression of beta-defensins in the tubotympanum of experimental otitis media. Acta Otolaryngol. 2006, 126, 1040–1045. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.A.; McGillivary, G.; Bakaletz, L.O. Extracellular DNA within a Nontypeable Haemophilus influenzae-Induced Biofilm Binds Human Beta Defensin-3 and Reduces Its Antimicrobial Activity. J. Innate Immun. 2013, 5, 24–38. [Google Scholar] [CrossRef]
- Howie, V.; Pollard, R.B.; Kleyn, K.; Lawrence, B.; Peskuric, T.; Paucker, K.; Baron, S.; Hiatt, R.A.; Ottesen, E.A.; Sotomayor, Z.R.; et al. Presence of Interferon During Bacterial Otitis Media. J. Infect. Dis. 1982, 145, 811–814. [Google Scholar] [CrossRef]
- Lasisi, A.O.; Olayemi, O.; Arinola, O.G.; Omilabu, S.A. Interferon-gamma in suppurative otitis media: Significance of otorrhoea type and disease outcome. J. Laryngol. Otol. 2009, 123, 1103–1107. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Park, H.J.; Choung, Y.H.; Song, J.W. Distribution of Lysozyme- and Lactoferrin-Secreting Cells in Eustachian Tube of Normal BALB/c Mouse. Korean J. Otorhinolaryngol.-Head Neck Surg. 1999, 42, 811–815. [Google Scholar]
- Kawano, H.; Paparella, M.M.; Ho, S.B.; Schachern, P.A.; Morizono, N.; Le, C.T.; Lin, J. Identification of mucin gene (MUC5B) in middle ear mucosa of patients with chonic otitis media. In Abstracts of the Twenty first Midwinter Research Meeting; Lim, D.J., Ed.; Association for Research in Otolaryngology: St. Petersburg Beach, FL, USA, 1998; p. 123. [Google Scholar]
- Lim, D.J.; Chun, Y.M.; Lee, H.Y.; Moon, S.K.; Chang, K.H.; Li, J.D. Cell biology in relation to pathogens otitis media—A review. Vaccine 2000, 19 (Suppl. 1), S17–S25. [Google Scholar] [CrossRef]
- Kim, S.H.; Park, M.G. Decreased Expression of Aquaporins in Otitis Media: What about Chronic Otitis Media? Clin. Exp. Otorhinolaryngol. 2019, 12, 235–236. [Google Scholar] [CrossRef] [Green Version]
- Hong, W.; Juneau, R.A.; Pang, B.; Swords, W.E. Survival of Bacterial Biofilms within Neutrophil Extracellular Traps Promotes Nontypeable Haemophilus influenzae Persistence in the Chinchilla Model for Otitis Media. J. Innate Immun. 2009, 1, 215–224. [Google Scholar] [CrossRef]
- Davies, L.C.; Jenkins, S.J.; Allen, J.E.; Taylor, P.R. Tissue-resident macrophages. Nat. Immunol. 2013, 14, 986–995. [Google Scholar] [CrossRef]
- Hussl, B.; Egg, G.; Romani, N.; Kong, W.; Schrott-Fischer, A. Dendritic Cells in the Normal Human Tympanic Membrane. Ann. Otol. Rhinol. Laryngol. 1995, 104, 803–807. [Google Scholar] [CrossRef] [PubMed]
- Jecker, P.; Pabst, R.; Westermann, J. The Mucosa of the Middle Ear and Eustachian Tube in the Young Rat: Number of Granulocytes, Macrophages, Dendritic Cells, NK Cells and T and B Lymphocytes in Healthy Animals and during Otitis Media. Acta Oto-Laryngol. 1996, 116, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Tong, H.H.; Lambert, G.; Li, Y.X.; Thurman, J.M.; Stahl, G.L.; Douthitt, K.; Clancy, C.; He, Y.; Bowman, A.S. Deletion of the Complement C5a Receptor Alleviates the Severity of Acute Pneumococcal Otitis Media following Influenza A Virus Infection in Mice. PLoS ONE 2014, 9, e95160. [Google Scholar] [CrossRef] [Green Version]
- Närkiö-Mäkelä, M.; Teppo, A.M.; Meri, S. Complement C3 cleavage and cytokines interleukin-1beta and tumor necrosis factor-alpha in otitis media with effusion. Laryngoscope 2000, 110, 1745–1749. [Google Scholar]
- Kwon, J.A. Initiation of Plant Immune Response. Mol. Cell. Biol. News 2010, 1–4. Available online: http://www.ksmcb.or.kr (accessed on 20 December 2021).
- Gómez-Gómez, L.; Boller, T. FLS2: An LRR Receptor–like Kinase Involved in the Perception of the Bacterial Elicitor Flagellin in Arabidopsis. Mol. Cell 2000, 5, 1003–1011. [Google Scholar] [CrossRef]
- Fliegmann, J.; Mithofer, A.; Wanner, G.; Ebel, J. An ancient enzyme domain hidden in the putative betaglucan elicitor receptor of soybean may play an active part in the perception of pathogen-associated molecular patterns during broad host resistance. J. Biol. Chem. 2004, 279, 1132–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maruta, N.; Burdett, H.; Lim, B.Y.J.; Hu, X.; Desa, S.; Manik, M.K.; Kobe, B. Structural basis of NLR activation and innate immune signalling in plants. Immunogenetics 2022, 1–22. [Google Scholar] [CrossRef]
- Pei, G.; Dorhoi, A. NOD-Like Receptors: Guards of Cellular Homeostasis Perturbation during Infection. Int. J. Mol. Sci. 2021, 22, 6714. [Google Scholar] [CrossRef]
- El Kasmi, F. How activated NLRs induce anti-microbial defenses in plants. Biochem. Soc. Trans. 2021, 49, 2177–2188. [Google Scholar] [CrossRef] [PubMed]
- Danis, J.; Mellett, M. Nod-Like Receptors in Host Defence and Disease at the Epidermal Barrier. Int. J. Mol. Sci. 2021, 22, 4677. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.B.; Lima, C.A.D.; Vajgel, G.; Sandrin-Garcia, P. The Role of NLRP3 Inflammasome in Lupus Nephritis. Int. J. Mol. Sci. 2021, 22, 12476. [Google Scholar] [CrossRef]
- Feerick, C.L.; McKernan, D.P. DNA methyltransferase inhibitors increase NOD-like receptor activity and expression in a monocytic cell line. Immunopharmacol. Immunotoxicol. 2021, 1–11. [Google Scholar] [CrossRef]
- Takeuchi, O.; Akira, S. Pattern Recognition Receptors and Inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [Green Version]
- Mittal, R.; Kodiyan, J.; Gerring, R.; Mathee, K.; Li, J.-D.; Grati, M.; Liu, X.Z. Role of innate immunity in the pathogenesis of otitis media. Int. J. Infect. Dis. 2014, 29, 259–267. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Ryu, E.W.; Bin Kim, J.; Yeo, S.G. Clinical Approaches for Understanding the Expression Levels of Pattern Recognition Receptors in Otitis Media with Effusion. Clin. Exp. Otorhinolaryngol. 2011, 4, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Cha, S.H.; Kim, Y.I.; Byun, J.Y.; Park, M.S.; Yeo, S.G. Age-dependent changes in pattern recognition receptor and cytokine mRNA expression in children with otitis media with effusion. Int. J. Pediatr. Otorhinolaryngol. 2015, 79, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Cha, S.H.; Lee, H.Y.; Lee, S.K.; Chung, H.Y.; Yeo, J.H.; Kim, Y.I.; Yeo, S.G. Decreased pattern-recognition receptor-mediated cytokine mRNA expression in obese children with otitis media with effusion. Clin. Exp. Otorhinolaryngol. 2014, 7, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.I.; Oh, S.; Webster, P.; Lee, Y.J.; Lim, D.; Moon, S.K. NOD2/RICK-dependent β-defensin 2 regulation. PLoS ONE 2014, 9, e90933. [Google Scholar]
- Kaur, R.; Casey, J.; Pichichero, M. Differences in Innate Immune Response Gene Regulation in the Middle Ear of Children Who Are Otitis Prone and in Those Not Otitis Prone. Am. J. Rhinol. Allergy 2016, 30, e218–e223. [Google Scholar] [CrossRef] [Green Version]
- Granath, A.; Cardell, L.-O.; Uddman, R.; Harder, H. Altered Toll- and Nod-like receptor expression in human middle ear mucosa from patients with chronic middle ear disease. J. Infect. 2011, 63, 174–176. [Google Scholar] [CrossRef]
- Lee, J.; Leichtle, A.; Zuckerman, E.; Pak, K.; Spriggs, M.; Wasserman, S.I.; Kurabi, A. NOD1/NOD2-mediated recognition of non-typeable Haemophilus influenzae activates innate immunity during otitis media. Innate Immun. 2019, 25, 503–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.G.; Park, D.C.; Shim, J.S.; Jung, H.; Park, M.S.; Kim, Y.I.; Lee, J.W.; Yeo, S.G. TLR-9, NOD-1, NOD-2, RIG-I and immunoglobulins in recurrent otitis media with effusion. Int. J. Pediatr. Otorhinolaryngol. 2010, 74, 1425–1429. [Google Scholar] [CrossRef]
- Kariya, S.; Okano, M.; Zhao, P.; Kataoka, Y.; Yoshinobu, J.; Maeda, Y.; Ishihara, H.; Higaki, T.; Nishizaki, K. Activation of NLRP3 inflammasome in human middle ear cholesteatoma and chronic otitis media. Acta Oto-Laryngol. 2015, 136, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Kariya, S.; Okano, M.; Zhao, P.; Maeda, Y.; Kataoka, Y.; Higaki, T.; Noda, Y.; Makihara, S.; Nishizaki, K. NLRP3 inflammasome expression in lipopolysaccharide-induced otitis media. Acta Oto-Laryngol. 2018, 138, 1061–1065. [Google Scholar] [CrossRef] [PubMed]
- Kariya, S.; Okano, M.; Zhao, P.; Maeda, Y.; Kataoka, Y.; Higaki, T.; Makihara, S.; Nishihira, J.; Tachibana, T.; Nishizaki, K. Role of Macrophage Migration Inhibitory Factor in NLRP3 Inflammasome Expression in Otitis Media. Otol. Neurotol. 2020, 41, 364–370. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| NLR | Localization | Cell Types | Signaling Molecule/Adapter or Binder | Microorganism | Function |
|---|---|---|---|---|---|
| NOD1 | Cytoplasm | Monocytes DCs Macrophages T and B lymphocytes Intestinal epithelium | RIP2 ATGL16 | Clostridium difficile Gram-negative bacteria Helicobacter pylori Listeria monocytogenes Salmonella typhimurium Shigella flexneri Aspergillus fumigatus | NF-κB activation Autophagy |
| NOD2 | Cytoplasm | Monocytes DCs Macrophages T and B lymphocytes Myeloid cells Bronchial epithelial cells | RIP2 CARD9 | A. fumigatus Citrobacter rodentium Escherichia coli Enterococcus faecalis Gram-negative bacteria Helicobacter hepaticus L. monocytogenes S. typhimurium Candida parapsilosis Chitin | NF-κB activation Autophagy |
| NLRC3 | Negative TLR regulator | ||||
| NLRC4 | Macrophages | ASC-NAIPS caspase-1 | C. rodentium Legionella Pseudomonas Salmonella Shigella | Inflammasome formation | |
| NLRC5 | Virus | Transcription MHC class I-related genes | |||
| NLRX1 | ROS production Autophagy Negative regulator of TLR MAVS-dependent signaling | ||||
| NLRP1 | Leukocytes Epithelial cells | ASC-caspase-1 | Bacillus anthracis | Inflammasome formation | |
| NLRP2 | Inflammasome formation | ||||
| NLRP3 | Cytoplasm | Monocytes DCs Macrophages Neutrophils T and B lymphocytes Epithelial cells Myeloid cells | ASC Caspase-1 | Bacteria Listeria monocytogenes Neisseria gonorrhoeae Staphylococcus S. typhimurium A. fumigatus C. parapsilosis Cryptococcus neoformans Histoplasma capsulatum Malassezia spp. Paracoccidioides brasiliensis Sporothrix schenckii Viral RNA | Inflammasome formation |
| NLRP4 | Cytoplasm | DCs Macrophages | TBK1 | Candida albicans | Autophagy Negative regulation of NF-κB |
| NLRP6 | E. coli L. monocytogenes S. typhimurium | Inflammasome formation Negative TLR regulator | |||
| NLRP7 | Inflammasome formation | ||||
| NLRP10 | Cytoplasm | DCs Macrophages Epithelial cells T lymphocytes | ASC Caspase-1 | C. albicans | Negative regulation of NF-κB DC migration |
| NLRP12 | Myeloid cells | S. typhimurium Yersinia pestis | Negative TLR regulator | ||
| NAIP | Inflammasome formation | ||||
| NAIP2 | Inflammasome formation | ||||
| NAIP5 | Inflammasome formation | ||||
| AIM2 | ASC-caspase-1 | Francisella tularensis | |||
| CIITA | Lymphocytes Endothelial cells | MHCII regulation |
| Author. Year [Reference] | Associated Diseases | Study Design | Species and/or Sample | Detection Method | Target Gene(s) or Pathway(s) Associated with NLRs | Results/Conclusion |
|---|---|---|---|---|---|---|
| Shin K, et al., 2020 [53] | OM | Animal study | Mice | ELISA; immunohistochemistry | NLRP3 | IL-1β, NLRP3, ASC, and caspase-1 levels increased in LPS-treated wild-type mice; these increases were attenuated in LPS-treated Mif−/− mice. /Macrophage MIF plays an important role in the production of IL-1β and the NLRP3 inflammasome. |
| Lee J, et al., 2019 [49] | OM | Animal study | Mice | Histology; macrophage phagocytosis and NTHi-killing assay; DNA microarrays | NOD1, NOD2 | NOD1-KO mice appeared to have reduced macrophage enlistment with a delayed inflammatory response by neutrophils and prolonged mucosal hyperplasia, whereas NOD2-KO mice exhibited an overall reduction in the number of leukocytes recruited to the middle ear, leading to delayed bacterial clearance. /NODs play a role in the pathogenesis and recovery of OM, reinforcing the importance of innate immune signaling in the protective host response. |
| Kariva S, et al., 2018 [52] | AOM | Animal study | Mice | ELISA; immunohistochemistry. | NLRP3 | Trans-tympanic injection of LPS significantly upregulated IL-1β, NLRP3, ASC, and caspase-1 in the middle ear compared with that in control mice and induced NLRP3 inflammasome components in the middle ear. /The NLRP3 inflammasome may play an important role in the pathogenesis of OM. |
| Kaur R, et al., 2016 [47] | AOM | Prospective study | Human: middle ear fluid | Quantitative PCR | NLR | Changes in innate gene regulation in AOM, measured in middle ear fluid, were similar whether caused by S. pneumoniae or NTHi. The innate immune response in otitis-sensitive children differed from that of children who were not otitis prone. /Defects in innate responses in the middle ear likely contribute to otitis proneness. |
| Kariya S, et al., 2016 [51] | COM, Chole OM | Prospective study | Human: Middle ear tissue samples | RT-PCR; immunohistochemistry. | NLRP3 | NLRP3, ASC, and caspase-1 mRNA levels were significantly elevated in cholesteatoma and COM compared with that in normal controls. NLRP3, ASC, and caspase-1 protein were detected in infiltrating inflammatory cells in cholesteatoma and COM. /The NLRP3 inflammasome plays an important role in the pathogenesis of middle ear diseases. Modulation of inflammasome-mediated inflammation may be a novel therapeutic strategy for cholesteatoma and COM. |
| Kim SH, et al., 2015 [44] | OME | Prospective study | Human: Middle ear fluid | RT-PCR | NOD1 NOD2 | TLR-2, TLR-9, NOD-1, NOD-2, IL-1, IL-6, and TNFα mRNA expression levels in effusion fluid were significantly higher in children aged 0–2 and >7 years (p < 0.05 each) than in those in 2–4 and 4–7-year groups. TLR-4, TLR-5, TLR-9, and NOD-1 mRNA expression levels were significantly lower in culture-positive than culture-negative patients (p < 0.05 each). /PRR and cytokine mRNA expression levels differ by age in children with OME. |
| Woo JI, et al., 2014 [46] | OM | Animal study | Mice | RT-PCR; ELISA; transmission electron microscopy; luciferase assay and gene silencing | NOD1 NOD2 | NOD2 silencing inhibited NTHi-induced β-defensin 2 production in human middle ear epithelial cells, whereas NOD2 over-expression augmented it. NTHi-induced β-defensin 2 up-regulation was attenuated by cytochalasin D, an inhibitor of actin polymerization, and was enhanced by α-hemolysin, a pore-forming toxin. α-hemolysin-mediated enhancement of NTHi-induced β-defensin 2 up-regulation was blocked by silencing of NOD2. An NOD2 deficiency reduced inflammatory reactions in response to intratympanic inoculation of NTHi and inhibited NTHi clearance from the middle ear. /Cytoplasmic release of internalized NTHi is involved in the pathogenesis of NTHi infections, and NOD2-mediated β-defensin 2 regulation contributes to the protection against NTHi-induced OM. |
| Kim YJ, et al., 2014 [45] | OME | Prospective study | Human: Middle ear fluid | Quantitative PCR | NOD1 NOD2 | NOD2-mediated expression of IL-6, IL-12, and TNF-α mRNA was significantly lower in obese than non-obese children (p < 0.05). /PRR-mediated cytokine mRNA expression is lower in obese than non-obese children with OME. |
| Lee SY, et al., 2011 [43] | OME | Prospective study | Human: Middle ear fluid | Quantitative PCR | NOD1 NOD2 | PCR analyses showed that all effusion fluid samples collected from patients with OME expressed NOD1 and NOD2 mRNA. However, no differences in expression levels of PRRs in relation to characteristics of exudates, presence of bacteria, or frequencies of ventilation tube insertion were found. /Exudates of OME patients show expression of PRRs related to the innate immune response regardless of the characteristics of effusion fluid, presence of bacteria in exudates, or frequency of ventilation tube insertion. |
| Granath A, et al., 2011 [48] | CMED | Prospective study | Human: middle ear mucosa | Quantitative PCR; immunohistochemistry | NOD1 NOD2 NALP3 | In the first report of its kind, clear immunohistochemical staining for NOD2 and NALP3 was detected in the epithelium and lamina propria of the middle ear mucosa. NOD1 was not detected, and no staining for NOD2 or NALP3 was observed in negative controls. The functions of NLRs in AOM and CMED remain a matter of speculation. /CMED might have an infectious origin regardless of clinical appearance, and inborn individual differences in the ability to express PRRs are part of its pathogenesis. |
| Kim MG, et al., 2010 [50] | Recurrent OME | Prospective study | Human: Middle ear fluid | Quantitative PCR; ELISA. | NOD1 NOD2 | NOD1 mRNA levels were significantly lower in the otitis-prone than non-otitis-prone group. There was no correlation between immunoglobulin concentration and the expression of PRPs. /Decreased expression of PRRs is associated with increased susceptibility to OME. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
You, M.-W.; Kim, D.; Lee, E.-H.; Park, D.-C.; Lee, J.-M.; Kang, D.-W.; Kim, S.-H.; Yeo, S.-G. The Roles of NOD-like Receptors in Innate Immunity in Otitis Media. Int. J. Mol. Sci. 2022, 23, 2350. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042350
You M-W, Kim D, Lee E-H, Park D-C, Lee J-M, Kang D-W, Kim S-H, Yeo S-G. The Roles of NOD-like Receptors in Innate Immunity in Otitis Media. International Journal of Molecular Sciences. 2022; 23(4):2350. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042350
Chicago/Turabian StyleYou, Myung-Won, Dokyoung Kim, Eun-Hye Lee, Dong-Choon Park, Jae-Min Lee, Dae-Woong Kang, Sang-Hoon Kim, and Seung-Geun Yeo. 2022. "The Roles of NOD-like Receptors in Innate Immunity in Otitis Media" International Journal of Molecular Sciences 23, no. 4: 2350. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042350