
Phosphatidylcholine Enhances Homeostasis in Peach Seedling Cell Membrane and Increases Its Salt Stress Tolerance by Phosphatidic Acid

Abstract
:1. Introduction
2. Results
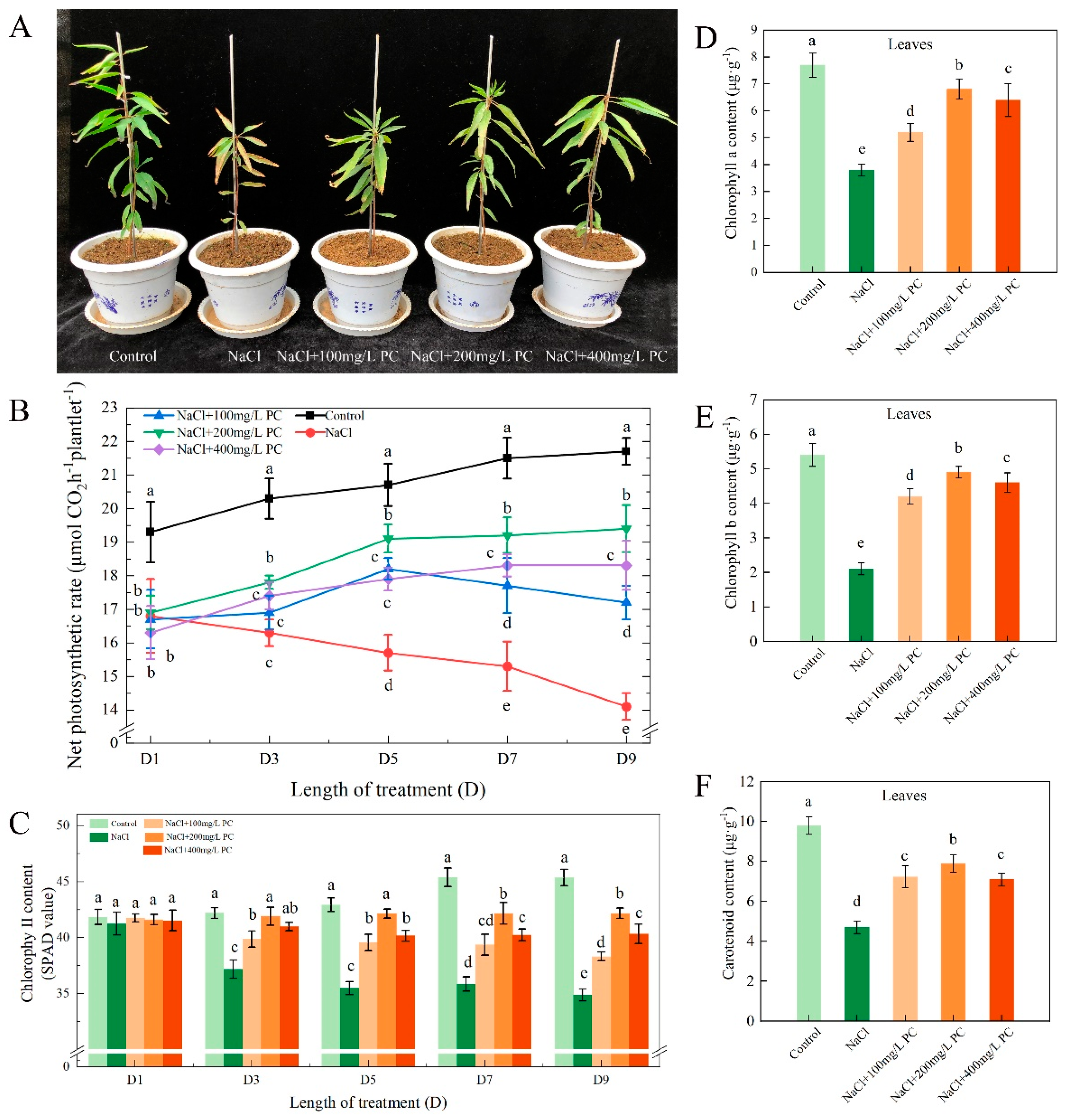
2.1. Exogenous Application of PC Improved the Net Photosynthetic Rate and Chlorophyll Content of Peach Leaves
2.2. Effect of PC on Stomatal Density and Size
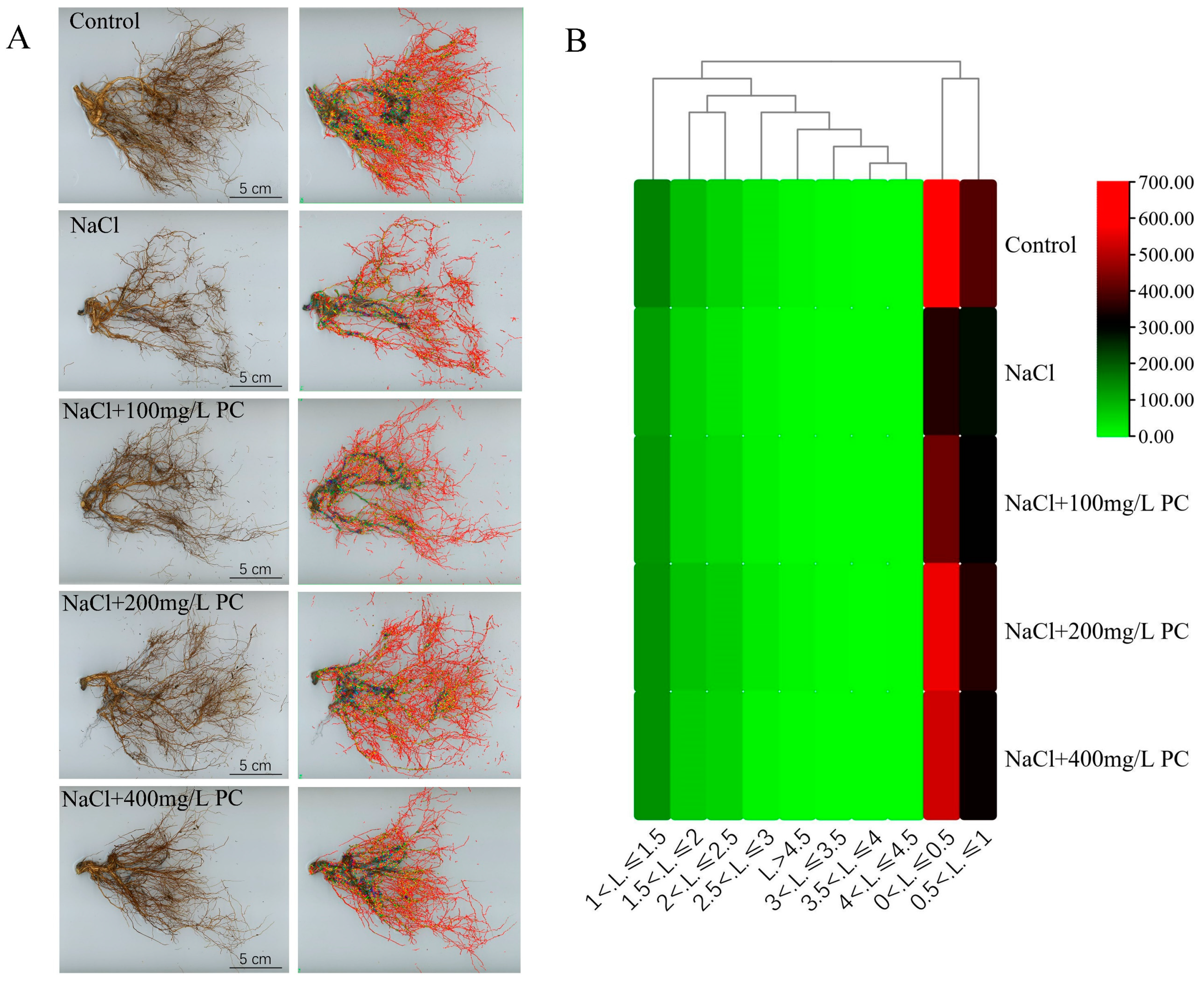
2.3. Structural Changes in the Peach Root System under Salt Stress
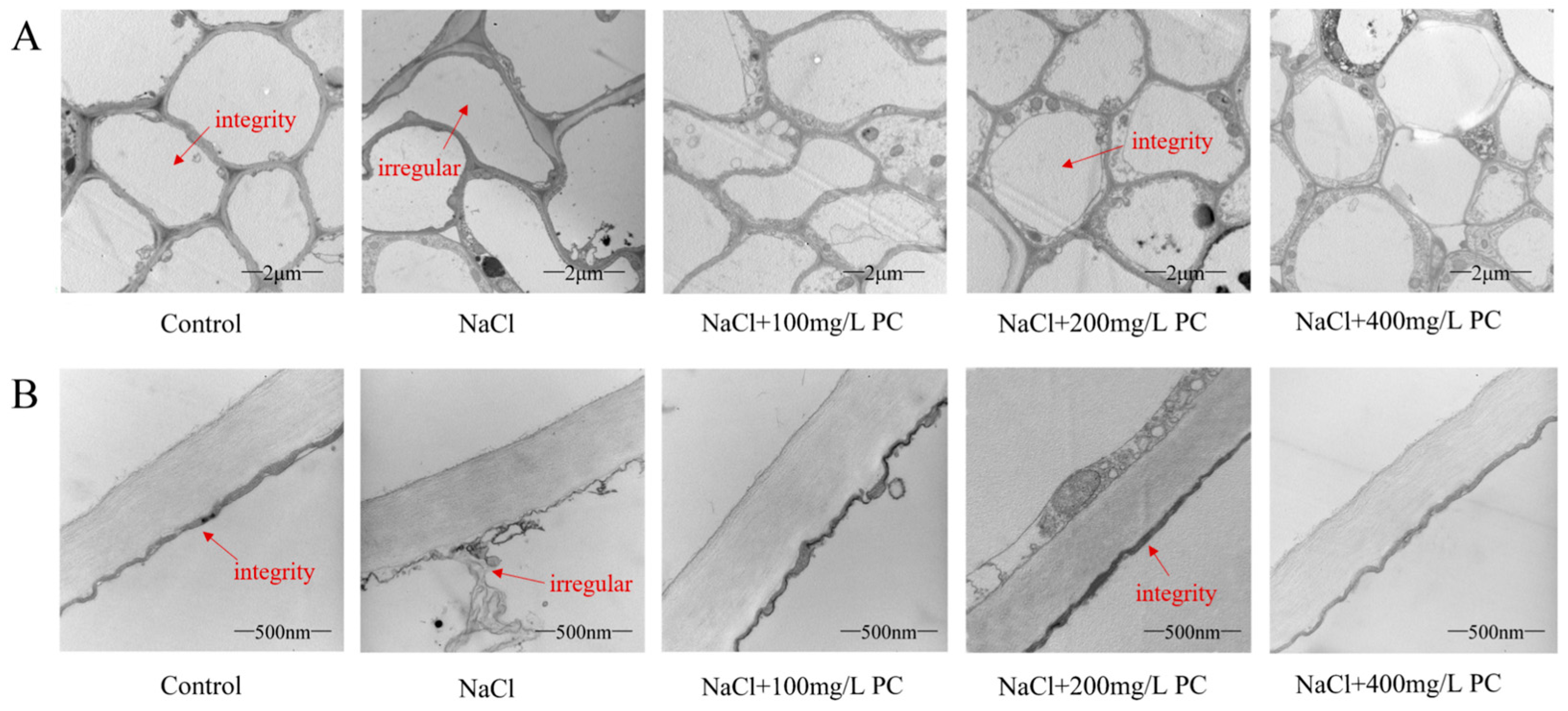
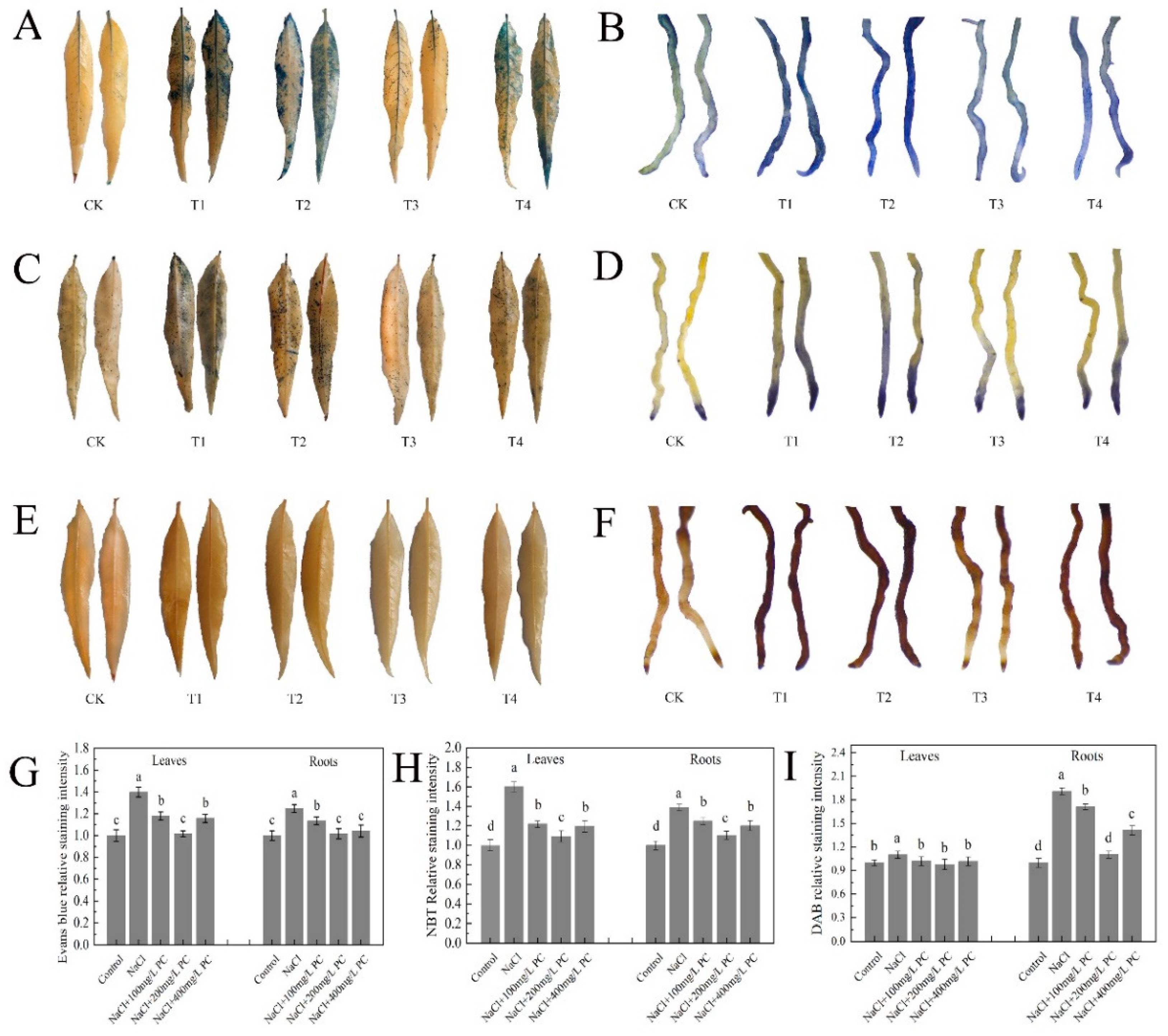
2.4. Root Cells and Cell Membrane Integrity of Peach Trees under Salt Stress Exposure
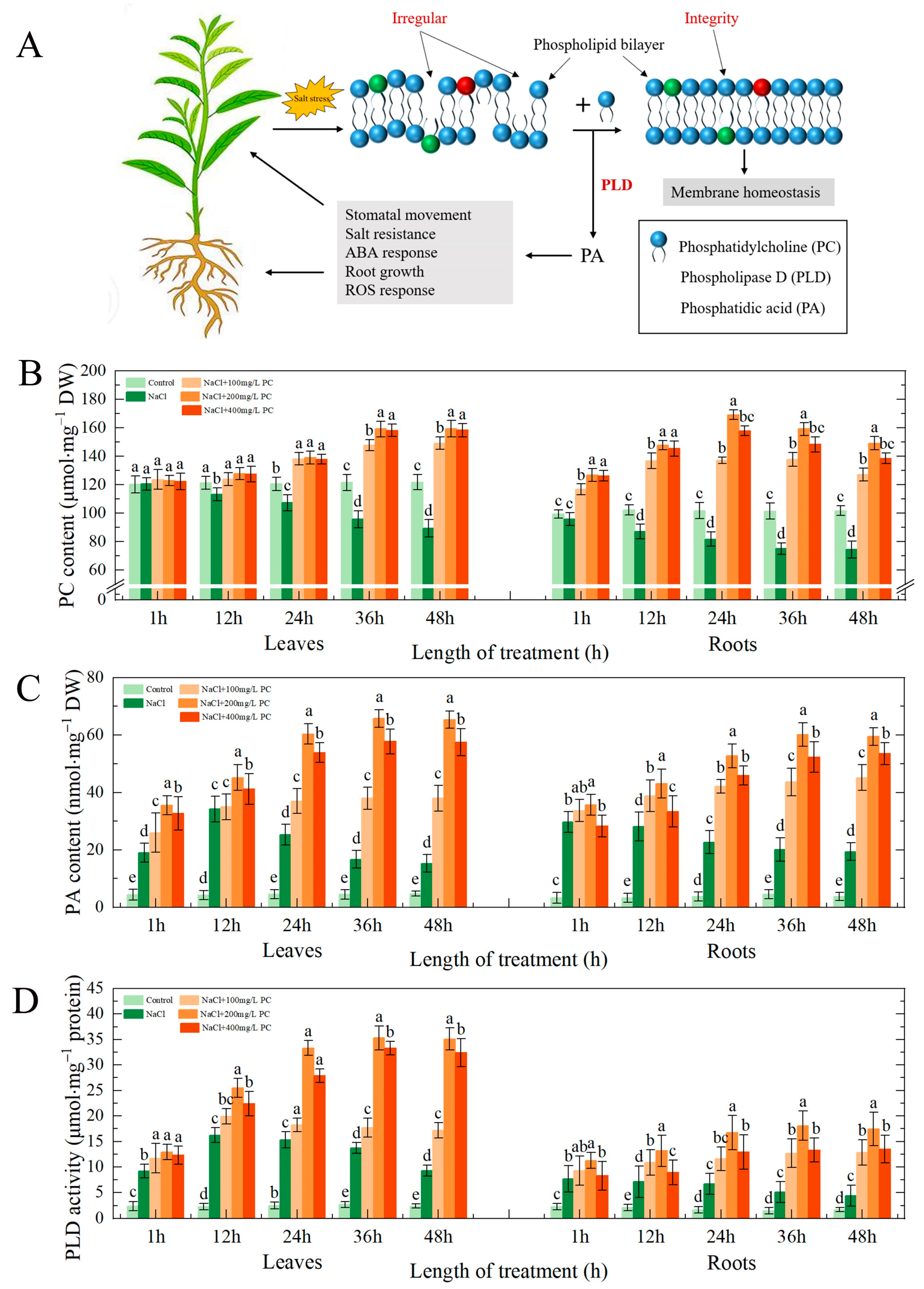
2.5. PC and PA Content, and PLD Activity in the Leaves and Roots under Salt Stress
2.6. Exogenous Application of PC Enhances Root Cell Activity
2.7. Electrolyte Leakage Rate and SOD, POD, MDA, Proline, and Soluble Sugar Contents
3. Discussion
3.1. PC, PA Contents, and PLD Activity in the Cells Exposed to Salt Stress
3.2. Effect of the Exogenous Application of PC on the Photosystem under Salt Stress Exposure
3.3. Effects of PC on the Root Structure, Growth, Cell Components, and Cell Membrane Integrity under Salt Stress
3.4. Application of PC on Maintaining Cell Activity under Salt Stress
3.5. Effect of PC Application on MDA, Proline, and Soluble Sugar Contents
4. Materials and Methods
4.1. Plant Material and Exogenous PC Treatment
4.2. PC and PA Content and PLD Enzyme Activity in Plants
4.3. Measurements of the Net Photosynthetic Rate and SPAD Values
4.4. Determination of Stomatal Area, Root Growth, and Root Cell Integrity
4.5. Measurements of Root Cell Activity, NBT and DAB Staining, and Electrolyte Leakage Rate
4.6. Determination of the Antioxidant system
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouhibi, C.; Attia, H.; Rebah, F.; Msilini, N.; Chebbi, M.; Aarrouf, J.; Urban, L.; Lachaal, M. Salt stress mitigation by seed priming with UV-C in lettuce plants: Growth, antioxidant activity and phenolic compounds. Plant Physiol. Biochem. 2014, 83, 126–133. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Lin, H. Signal transduction pathways in response to salt stress in plants. Plant Physiol. J. 2011, 47, 119–128. [Google Scholar]
- McMahon, H.T.; Gallop, J.L. Membrane curvature and mechanisms of dynamic cell membrane remodelling. Nature 2005, 438, 590–596. [Google Scholar] [CrossRef]
- Gigon, A.; Matos, A.-R.; Laffray, D.; Zuily-Fodil, Y.; Pham-Thi, A.-T. Effect of drought stress on lipid metabolism in the leaves of Arabidopsis thaliana (ecotype Columbia). Ann. Bot. 2004, 94, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Xing, S.; Guo, Q.; Zhao, M.; Zhang, J.; Gao, Q.; Wang, G.; Wang, W. Drought tolerance through over-expression of the expansin gene TaEXPB23 in transgenic tobacco. J. Plant Physiol. 2011, 168, 960–966. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, C.; Bedair, M.; Welti, R.; Sumner, L.W.; Baxter, I.; Wang, X. Suppression of phospholipase Dγs confers increased aluminum resistance in Arabidopsis thaliana. PLoS ONE 2011, 6, e28086. [Google Scholar] [CrossRef] [Green Version]
- Harwood, J.L. Recent advances in the biosynthesis of plant fatty acids. Biochim. Biophys. Acta Lipids Lipid Metab. 1996, 1301, 7–56. [Google Scholar] [CrossRef]
- Devaiah, S.P.; Roth, M.R.; Baughman, E.; Li, M.; Tamura, P.; Jeannotte, R.; Welti, R.; Wang, X. Quantitative profiling of polar glycerolipid species from organs of wild-type Arabidopsis and a Phospholipase Dα1 knockout mutant. Phytochemistry 2006, 67, 1907–1924. [Google Scholar] [CrossRef]
- Botella, C.; Jouhet, J.; Block, M.A. Importance of phosphatidylcholine on the chloroplast surface. Prog. Lipid Res. 2017, 65, 12–23. [Google Scholar] [CrossRef]
- Nakamura, Y.; Andrés, F.; Kanehara, K.; Liu, Y.-C.; Dörmann, P.; Coupland, G. Arabidopsis florigen FT binds to diurnally oscillating phospholipids that accelerate flowering. Nat. Commun. 2014, 5, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Qin, C.; Zhao, J.; Wang, X. Phospholipase Dα1-derived phosphatidic acid interacts with ABI1 phosphatase 2C and regulates abscisic acid signaling. Proc. Natl. Acad. Sci. USA 2004, 101, 9508–9513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Hong, Y.; Wang, X. Phospholipase D-and phosphatidic acid-mediated signaling in plants. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2009, 1791, 927–935. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Zhang, W.; Wang, X. Phospholipase D and phosphatidic acid signalling in plant response to drought and salinity. Plant Cell Environ. 2010, 33, 627–635. [Google Scholar] [CrossRef]
- Wang, X.; Devaiah, S.P.; Zhang, W.; Welti, R. Signaling functions of phosphatidic acid. Prog. Lipid Res. 2006, 45, 250–278. [Google Scholar] [CrossRef]
- Liu, Y.-C.; Lin, Y.-C.; Kanehara, K.; Nakamura, Y. A methyltransferase trio essential for phosphatidylcholine biosynthesis and growth. Plant Physiol. 2019, 179, 433–445. [Google Scholar] [CrossRef] [Green Version]
- Shimojima, M.; Watanabe, T.; Madoka, Y.; Koizumi, R.; Yamamoto, M.P.; Masuda, K.; Yamada, K.; Masuda, S.; Ohta, H. Differential regulation of two types of monogalactosyldiacylglycerol synthase in membrane lipid remodeling under phosphate-limited conditions in sesame plants. Front. Plant Sci. 2013, 4, 469. [Google Scholar] [CrossRef] [Green Version]
- McCord, J.M.; Fridovich, I. Superoxide dismutase: An enzymic function for erythrocuprein (hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar] [CrossRef]
- Ismail, A.; Takeda, S.; Nick, P. Life and death under salt stress: Same players, different timing? J. Exp. Bot. 2014, 65, 2963–2979. [Google Scholar] [CrossRef]
- Torres-Franklin, M.L.; Gigon, A.; De Melo, D.F.; Zuily-Fodil, Y.; Pham-Thi, A.T. Drought stress and rehydration affect the balance between MGDG and DGDG synthesis in cowpea leaves. Physiol. Plant. 2007, 131, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Magdy, M.; Mansour, F.; van Hasselt, P.R.; Kuiper, P.J. Plasma membrane lipid alterations induced by NaCl in winter wheat roots. Physiol. Plant. 1994, 92, 473–478. [Google Scholar] [CrossRef]
- Cramer, G.R.; Läuchli, A.; Polito, V.S. Displacement of Ca2+ by Na+ from the plasmalemma of root cells: A primary response to salt stress? Plant Physiol. 1985, 79, 207–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levitt, J. Responses of Plants to Environmental Stresses: Water, Radiation, Salt, and Other Stresses; Academic Press: Cambridge, MA, USA, 1980; Volume 2. [Google Scholar]
- Welti, R.; Li, W.; Li, M.; Sang, Y.; Biesiada, H.; Zhou, H.-E.; Rajashekar, C.; Williams, T.D.; Wang, X. Profiling membrane lipids in plant stress responses: Role of phospholipase Dα in freezing-induced lipid changes in Arabidopsis. J. Biol. Chem. 2002, 277, 31994–32002. [Google Scholar] [CrossRef] [Green Version]
- Kolesnikov, Y.; Nokhrina, K.; Kretynin, S.; Volotovski, I.; Martinec, J.; Romanov, G.; Kravets, V. Molecular structure of phospholipase D and regulatory mechanisms of its activity in plant and animal cells. Biochemistry 2012, 77, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.-H.; Lee, C.S.; Hwang, D.; Ryu, S.H. Understanding of the roles of phospholipase D and phosphatidic acid through their binding partners. Prog. Lipid Res. 2012, 51, 71–81. [Google Scholar] [CrossRef]
- Cruz-Ramírez, A.; Oropeza-Aburto, A.; Razo-Hernández, F.; Ramírez-Chávez, E.; Herrera-Estrella, L. Phospholipase DZ2 plays an important role in extraplastidic galactolipid biosynthesis and phosphate recycling in Arabidopsis roots. Proc. Natl. Acad. Sci. USA 2006, 103, 6765–6770. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.; Pan, X.; Welti, R.; Wang, X. Phospholipase Dα3 is involved in the hyperosmotic response in Arabidopsis. Plant Cell 2008, 20, 803–816. [Google Scholar] [CrossRef] [Green Version]
- Klimecka, M.; Szczegielniak, J.; Godecka, L.; Lewandowska-Gnatowska, E.; Dobrowolska, G.; Muszyńska, G. Regulation of wound-responsive calcium-dependent protein kinase from maize (ZmCPK11) by phosphatidic acid. Acta Biochim. Pol. 2011, 58, 589–595. [Google Scholar] [CrossRef]
- Testerink, C.; Munnik, T. Phosphatidic acid: A multifunctional stress signaling lipid in plants. Trends Plant Sci. 2005, 10, 368–375. [Google Scholar] [CrossRef]
- Hagio, M.; Gombos, Z.; Várkonyi, Z.; Masamoto, K.; Sato, N.; Tsuzuki, M.; Wada, H. Direct evidence for requirement of phosphatidylglycerol in photosystem II of photosynthesis. Plant Physiol. 2000, 124, 795–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, N.; Hagio, M.; Wada, H.; Tsuzuki, M. Environmental effects on acidic lipids of thylakoid membranes. Biochem. Soc. Trans. 2000, 28, 912–914. [Google Scholar] [CrossRef] [PubMed]
- Mizusawa, N.; Wada, H. The role of lipids in photosystem II. Biochim. Biophys. Acta Bioenerg. 2012, 1817, 194–208. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.J.; Kim, M.S.; Lee, J.K. Phosphatidylcholine is required for the efficient formation of photosynthetic membrane and B800-850 light-harvesting complex in Rhodobacter sphaeroides. J. Microbiol. Biotechnol. 2007, 17, 373–377. [Google Scholar] [PubMed]
- Jiang, Y.; Wu, K.; Lin, F.; Qu, Y.; Liu, X.; Zhang, Q. Phosphatidic acid integrates calcium signaling and microtubule dynamics into regulating ABA-induced stomatal closure in Arabidopsis. Planta 2014, 239, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Lin, F.; Mao, T.; Nie, J.; Yan, M.; Yuan, M.; Zhang, W. Phosphatidic acid regulates microtubule organization by interacting with MAP65-1 in response to salt stress in Arabidopsis. Plant Cell 2012, 24, 4555–4576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potocký, M.; Eliáš, M.; Profotová, B.; Novotná, Z.; Valentová, O.; Žárský, V. Phosphatidic acid produced by phospholipase D is required for tobacco pollen tube growth. Planta 2003, 217, 122–130. [Google Scholar] [CrossRef]
- Anthony, R.G.; Henriques, R.; Helfer, A.; Meszaros, T.; Rios, G.; Testerink, C.; Munnik, T.; Deák, M.; Koncz, C.; Bögre, L. A protein kinase target of a PDK1 signalling pathway is involved in root hair growth in Arabidopsis. EMBO J. 2004, 23, 572–581. [Google Scholar] [CrossRef]
- Li, M.; Qin, C.; Welti, R.; Wang, X. Double knockouts of phospholipases D ζ 1 and D ζ 2 in Arabidopsis affect root elongation during phosphate-limited growth but do not affect root hair patterning. Plant Physiol. 2006, 140, 761–770. [Google Scholar] [CrossRef] [Green Version]
- Wang, X. Lipid signaling. Curr. Opin. Plant Biol. 2004, 7, 329–336. [Google Scholar] [CrossRef]
- Hong, Y.; Devaiah, S.P.; Bahn, S.C.; Thamasandra, B.N.; Li, M.; Welti, R.; Wang, X. Phospholipase Dε and phosphatidic acid enhance Arabidopsis nitrogen signaling and growth. Plant J. 2009, 58, 376–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colla, G.; Rouphael, Y.; Jawad, R.; Kumar, P.; Rea, E.; Cardarelli, M. The effectiveness of grafting to improve NaCl and CaCl2 tolerance in cucumber. Sci. Hortic. 2013, 164, 380–391. [Google Scholar] [CrossRef]
- Aloni, B.; Cohen, R.; Karni, L.; Aktas, H.; Edelstein, M. Hormonal signaling in rootstock-scion interactions. Sci. Hortic. 2010, 127, 119–126. [Google Scholar] [CrossRef]
- Penella, C.; Landi, M.; Guidi, L.; Nebauer, S.G.; Pellegrini, E.; Bautista, A.S.; Remorini, D.; Nali, C.; López-Galarza, S.; Calatayud, A. Salt-tolerant rootstock increases yield of pepper under salinity through maintenance of photosynthetic performance and sinks strength. J. Plant Physiol. 2016, 193, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bewley, J.D. Physiological aspects of desiccation tolerance. Annu. Rev. Plant Biol. 1979, 30, 195–238. [Google Scholar] [CrossRef]
- Terashima, I.; Hanba, Y.T.; Tholen, D.; Niinemets, Ü. Leaf functional anatomy in relation to photosynthesis. Plant Physiol. 2011, 155, 108–116. [Google Scholar] [CrossRef] [Green Version]
- Hodge, A.; Berta, G.; Doussan, C.; Merchan, F.; Crespi, M. Plant root growth, architecture and function. Plant Soil 2009, 321, 153–187. [Google Scholar] [CrossRef]
- Zhu, J.-K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [Green Version]
- Flohe, L.; Günzler, W.; Schock, H. Glutathione peroxidase: A selenoenzyme. FEBS Lett. 1973, 32, 132–134. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-J.; Zhang, S.-S.; Tian, X.-Y.; Ren, D. Effects of salt stress on protective enzyme system, MDA content and membrane permeability of NHC foliar. Grassl. Turf 2008, 2, 30–34. [Google Scholar] [CrossRef]
- Hare, P.D.; Cress, W.A.; Van Staden, J. Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ. 1998, 21, 535–553. [Google Scholar] [CrossRef]
- Munnik, T.; Meijer, H.J.; Ter Riet, B.; Hirt, H.; Frank, W.; Bartels, D.; Musgrave, A. Hyperosmotic stress stimulates phospholipase D activity and elevates the levels of phosphatidic acid and diacylglycerol pyrophosphate. Plant J. 2000, 22, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Zonia, L.; Munnik, T. Osmotically induced cell swelling versus cell shrinking elicits specific changes in phospholipid signals in tobacco pollen tubes. Plant Physiol. 2004, 134, 813–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Wang, X. A novel phospholipase D of Arabidopsis that is activated by oleic acid and associated with the plasma membrane. Plant Physiol. 2001, 127, 1102–1112. [Google Scholar] [CrossRef]
- Zhao, S.-J. Experimental Guidance for Plant Physiology; Agricultural Science and Technology Press: Beijing, China, 2015. [Google Scholar]
- Baker, C.J.; Mock, N.M. An improved method for monitoring cell death in cell suspension and leaf disc assays using Evans blue. Plant Cell Tissue Organ Cult. 1994, 39, 7–12. [Google Scholar] [CrossRef]
- Nayyar, H. Accumulation of osmolytes and osmotic adjustment in water-stressed wheat (Triticum aestivum) and maize (Zea mays) as affected by calcium and its antagonists. Environ. Exp. Bot. 2003, 50, 253–264. [Google Scholar] [CrossRef]
- Liu, N.; Lin, Z.; Guan, L.; Gaughan, G.; Lin, G. Antioxidant enzymes regulate reactive oxygen species during pod elongation in Pisum sativum and Brassica chinensis. PLoS ONE 2014, 9, e87588. [Google Scholar] [CrossRef] [Green Version]
- Tang, K.; Zhan, J.-C.; Yang, H.-R.; Huang, W.-D. Changes of resveratrol and antioxidant enzymes during UV-induced plant defense response in peanut seedlings. J. Plant Physiol. 2010, 167, 95–102. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Prunus persica (L.) Batsch. Stomatal Measurements | |||
|---|---|---|---|
| Length (μm) | Width (μm) | Area (μm2) | |
| Control | 22.8 ± 3.7 a | 15.4 ± 1.5 a | 329.1 ± 0.51 a |
| NaCl | 21.8 ± 2.1 b | 11.3 ± 1.3 d | 213.5 ± 0.21 d |
| NaCl + 100 mg/L PC | 21.3 ± 2.5 b | 12.8 ± 1.2 c | 245.3 ± 0.20 c |
| NaCl + 200 mg/L PC | 21.7 ± 3.5 b | 14.5 ± 1.8 b | 293.3 ± 0.61 b |
| NaCl + 400 mg/L PC | 21.5 ± 2.3 b | 13.1 ± 1.6 c | 267.9 ± 1.01 c |
| Length (cm) | Surface Area (cm2) | Root Volume (cm3) | Tips | Forks | |
|---|---|---|---|---|---|
| Control | 1506.6 ± 92.7 a | 251.6 ± 17.1 a | 3.3 ± 0.2 a | 2927 ± 112 a | 17602 ± 991 a |
| NaCl | 793.7 ± 64.3 e | 131.1 ± 9.9 e | 1.7 ± 0.1 d | 1457 ± 125 e | 5628 ± 481 d |
| NaCl + 100 mg/L PC | 907.7 ± 66.9 d | 160.4 ± 8.7 d | 2.3 ± 0.1 c | 1658 ± 97 d | 7991 ± 698 c |
| NaCl + 200 mg/L PC | 1311.5 ± 97.2 b | 184.9 ± 8.3 b | 2.8 ± 0.2 b | 2433 ± 131 b | 10361 ± 852 b |
| NaCl + 400 mg/L PC | 1029.2 ± 95.5 c | 170.9 ± 7.8 c | 2.3 ± 0.2 c | 2202 ± 148 c | 7952 ± 650 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, M.; Liu, X.; Gao, H.; Zhang, B.; Peng, F.; Xiao, Y. Phosphatidylcholine Enhances Homeostasis in Peach Seedling Cell Membrane and Increases Its Salt Stress Tolerance by Phosphatidic Acid. Int. J. Mol. Sci. 2022, 23, 2585. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052585
Sun M, Liu X, Gao H, Zhang B, Peng F, Xiao Y. Phosphatidylcholine Enhances Homeostasis in Peach Seedling Cell Membrane and Increases Its Salt Stress Tolerance by Phosphatidic Acid. International Journal of Molecular Sciences. 2022; 23(5):2585. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052585
Chicago/Turabian StyleSun, Maoxiang, Xiaolong Liu, Huaifeng Gao, Binbin Zhang, Futian Peng, and Yuansong Xiao. 2022. "Phosphatidylcholine Enhances Homeostasis in Peach Seedling Cell Membrane and Increases Its Salt Stress Tolerance by Phosphatidic Acid" International Journal of Molecular Sciences 23, no. 5: 2585. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052585