
Mitogen-Activated Protein Kinase and Substrate Identification in Plant Growth and Development


Abstract
:1. Introduction
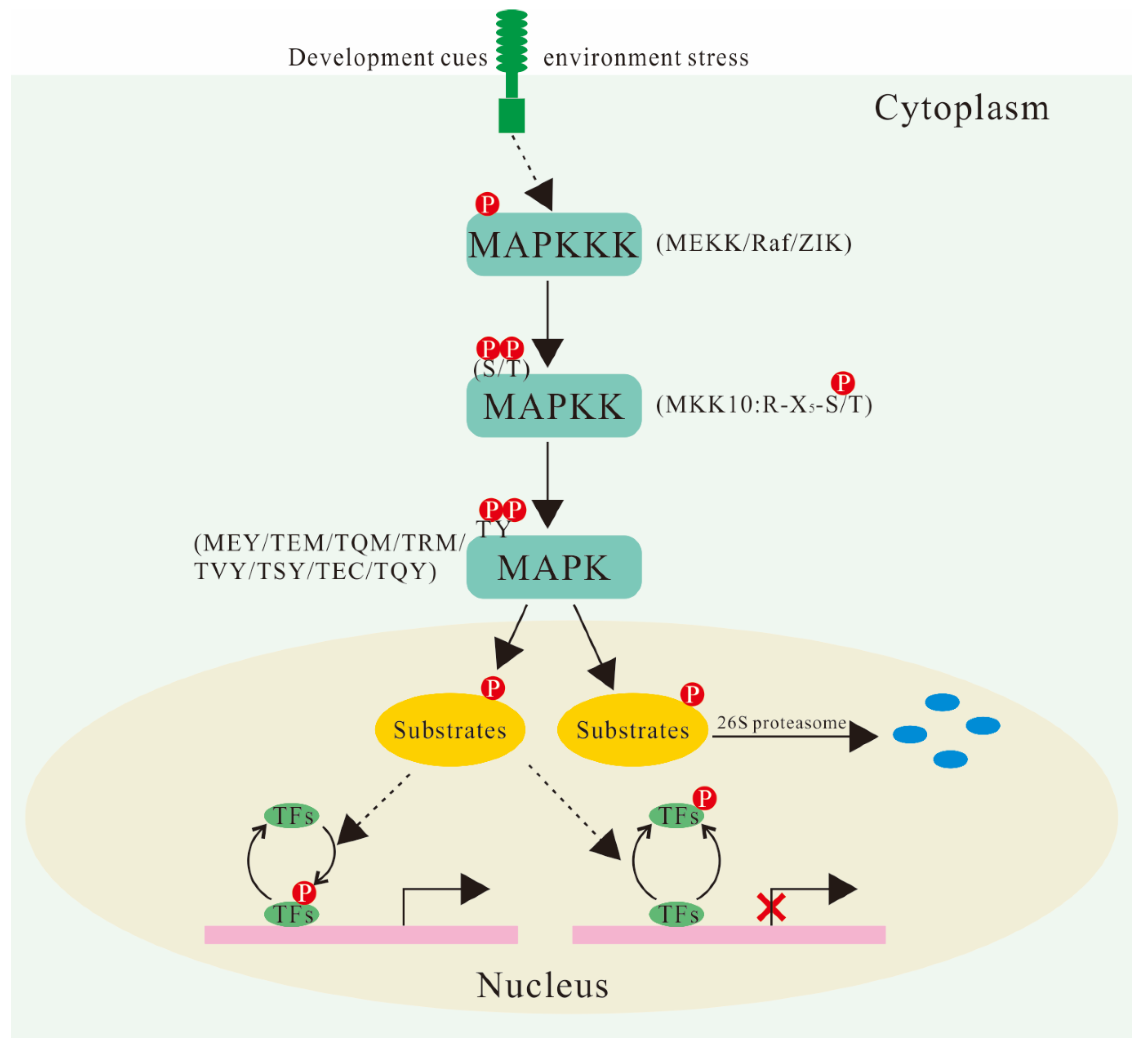
2. Composition and Classification of MAPK Cascades in Plants
3. MAPK Cascades Are Involved in a Variety of Signal Transduction Pathways in Plants
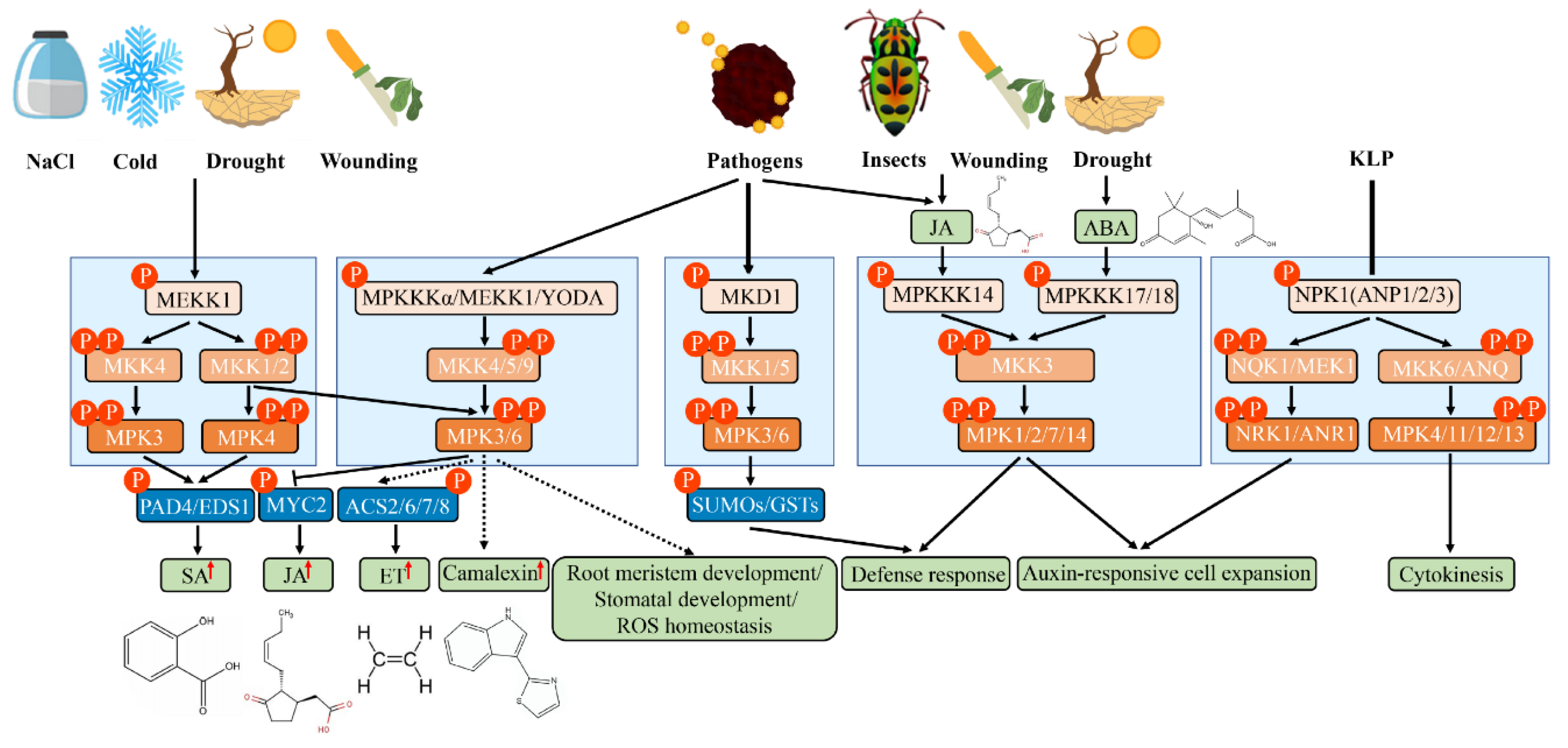
3.1. MAPK Cascades in ROS Signaling
3.2. MAPK Cascade in Hormone Signaling
3.3. MAPK Cascades in Biotic Stress
3.4. MAPK Cascades in Abiotic Stress
3.5. MAPK Cascades in Cell Division and Differentiation
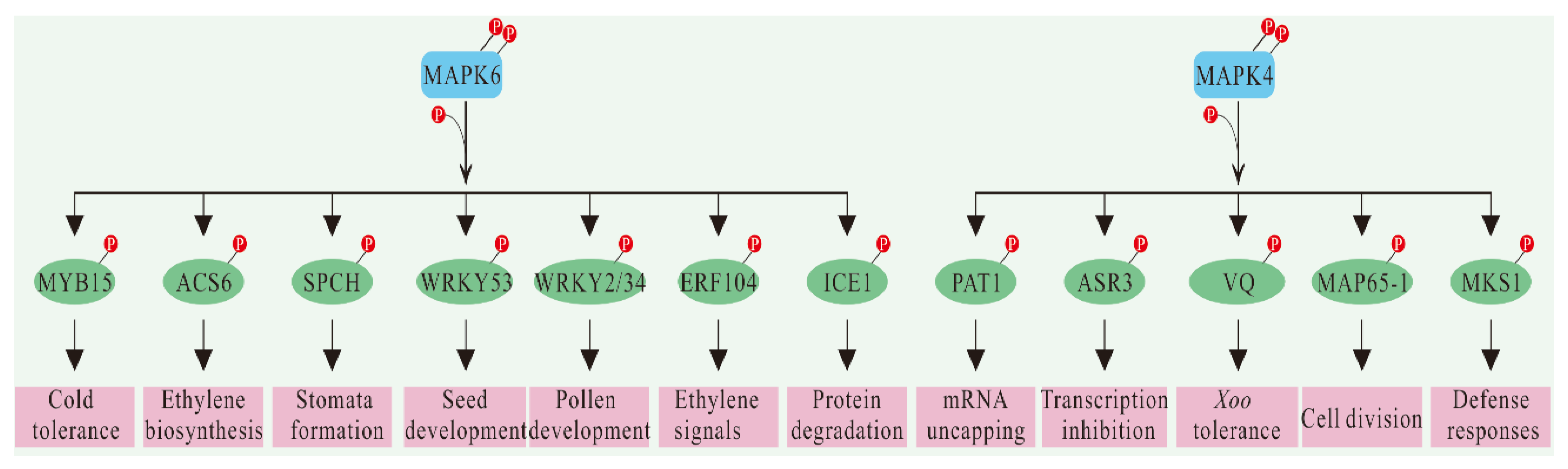
4. Substrates Identification of MAPKs
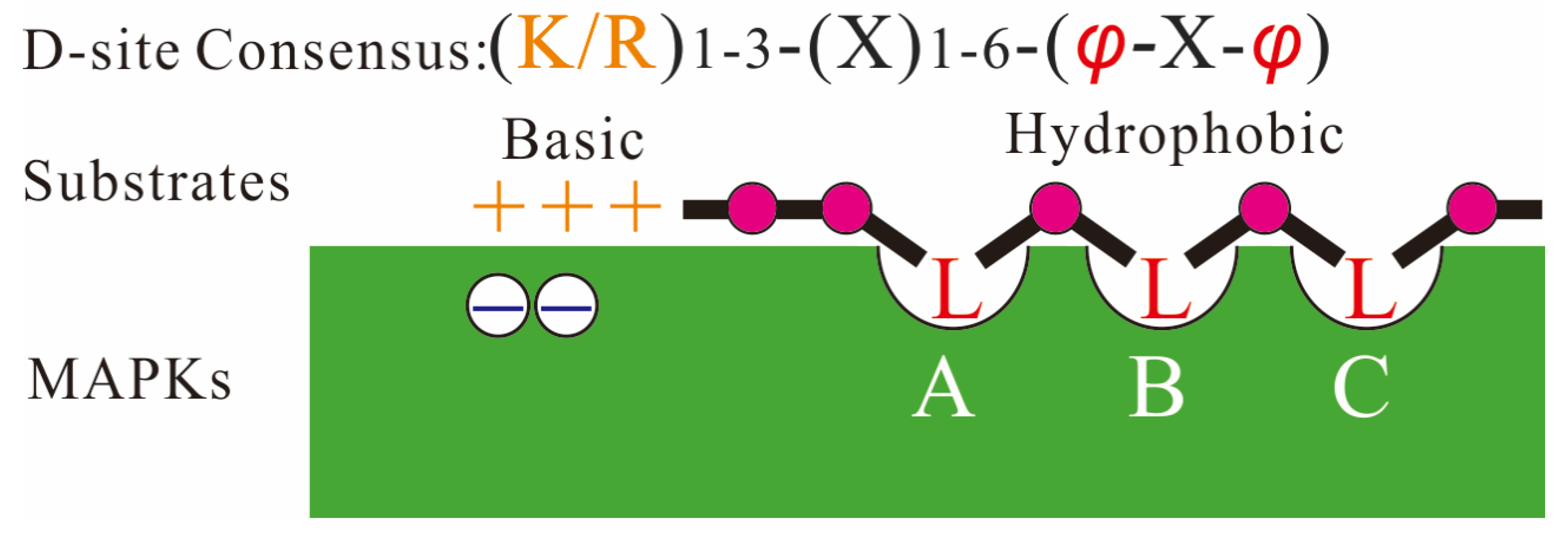
4.1. Interaction Domains of MAPKs and Their Substrates
4.2. General Strategy and Research Status of MAPK Substrate Identification
5. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xu, J.; Zhang, S.Q. Mitogen-activated protein kinase cascades in signaling plant growth and development. Trends Plant Sci. 2015, 20, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Wu, J.; Jiang, M.; Wang, Y. Plant mitogen-activated protein kinase cascades in environmental stresses. Int. J. Mol. Sci. 2021, 22, 1543. [Google Scholar] [CrossRef] [PubMed]
- Jagodzik, P.; Tajdel-Zielinska, M.; Ciesla, A.; Marczak, M.; Ludwikow, A. Mitogen-activated protein kinase cascades in plant hormone signaling. Front. Plant Sci. 2018, 9, 1387. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.Z.; Zhang, S.Q. MAPK cascades in plant disease resistance signaling. Annu. Rev. Phytopathol. 2013, 51, 245–266. [Google Scholar] [CrossRef]
- Hamel, L.P.; Nicole, M.C.; Sritubtim, S.; Morency, M.J.; Ellis, M.; Ehlting, J.; Beaudoin, N.; Barbazuk, B.; Klessig, D.; Lee, J.; et al. Ancient signals: Comparative genomics of plant MAPK and MAPKK gene families. Trends Plant Sci. 2006, 11, 192–198. [Google Scholar] [CrossRef]
- Mohanta, T.K.; Arora, P.K.; Mohanta, N.; Parida, P.; Bae, H. Identification of new members of the MAPK gene family in plants shows diverse conserved domains and novel activation loop variants. BMC Genom. 2015, 16, 58. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Wang, L.; Yuan, M. Update on the roles of rice MAPK cascades. Int. J. Mol. Sci. 2021, 22, 1679. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, S.X.; Harmon, A.C. Protein-protein interactions in plant mitogen-activated protein kinase cascades. J. Exp. Bot. 2016, 67, 607–618. [Google Scholar] [CrossRef] [Green Version]
- Janitza, P.; Ullrich, K.K.; Quint, M. Toward a comprehensive phylogenetic reconstruction of the evolutionary history of mitogen-activated protein kinases in the plant kingdom. Front. Plant Sci. 2012, 3, 271. [Google Scholar] [CrossRef] [Green Version]
- Roux, M.E.; Rasmussen, M.W.; Palma, K.; Lolle, S.; Regue, A.M.; Bethke, G.; Glazebrook, J.; Zhang, W.P.; Sieburth, L.; Larsen, M.R.; et al. The mRNA decay factor PAT1 functions in a pathway including MAP kinase 4 and immune receptor SUMM2. Embo J. 2015, 34, 593–608. [Google Scholar] [CrossRef] [Green Version]
- Andreasson, E.; Jenkins, T.; Brodersen, P.; Thorgrimsen, S.; Petersen, N.H.T.; Zhu, S.J.; Qiu, J.L.; Micheelsen, P.; Rocher, A.; Petersen, M.; et al. The MAP kinase substrate MKS1 is a regulator of plant defense responses. Embo J. 2005, 24, 2579–2589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feilner, T.; Hultschig, C.; Lee, J.; Meyer, S.; Immink, R.G.H.; Koenig, A.; Possling, A.; Seitz, H.; Beveridge, A.; Scheel, D.; et al. High throughput identification of potential Arabidopsis mitogen-activated protein kinases substrates. Mol. Cell. Proteom. 2005, 4, 1558–1568. [Google Scholar] [CrossRef] [Green Version]
- Whisenant, T.C.; Ho, D.T.; Benz, R.W.; Rogers, J.S.; Kaake, R.M.; Gordon, E.A.; Huang, L.; Baldi, P.; Bardwell, L. Computational prediction and experimental verification of new MAP kinase docking sites and substrates including Gli transcription factors. PLoS Comput. Biol. 2010, 6, 1000908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoehenwarter, W.; Thomas, M.; Nukarinen, E.; Egelhofer, V.; Rohrig, H.; Weckwerth, W.; Conrath, U.; Beckers, G.J.M. Identification of novel in vivo MAP kinase substrates in Arabidopsis thaliana through use of tandem metal oxide affinity chromatography. Mol. Cell. Proteom. 2013, 12, 369–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umezawa, T.; Sugiyama, N.; Takahashi, F.; Anderson, J.C.; Ishihama, Y.; Peck, S.C.; Shinozaki, K. Genetics and phosphoproteomics reveal a protein phosphorylation network in the abscisic acid signaling pathway in Arabidopsis thaliana. Sci. Signal. 2013, 6, rs8. [Google Scholar] [CrossRef]
- Wang, P.C.; Xue, L.; Batelli, G.; Lee, S.; Hou, Y.J.; Van Oosten, M.J.; Zhang, H.M.; Tao, W.A.; Zhu, J.K. Quantitative phosphoproteomics identifies SnRK2 protein kinase substrates and reveals the effectors of abscisic acid action. Proc. Natl. Acad. Sci. USA 2013, 110, 11205–11210. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.; Niu, C.; Cao, J.M.; Ni, D.A.; Chu, Z.Q. In silico-prediction of protein-protein interactions network about MAPKs and PP2Cs reveals a novel docking site variants in Brachypodium distachyon. Sci. Rep. 2018, 8, 15083. [Google Scholar] [CrossRef]
- Lee, S.S.; Ayrapetov, M.K.; Kemble, D.J.; Parang, K.; Sun, G.Q. Docking-based substrate recognition by the catalytic domain of a protein tyrosine kinase, C-terminal Src kinase (Csk). J. Biol. Chem. 2006, 281, 8183–8189. [Google Scholar] [CrossRef] [Green Version]
- Ichimura, K.; Shinozaki, K.; Tena, G.; Sheen, J.; Henry, Y.; Champion, A.; Kreis, M.; Zhang, S.Q.; Hirt, H.; Wilson, C.; et al. Mitogen-activated protein kinase cascades in plants: A new nomenclature. Trends Plant Sci. 2002, 7, 301–308. [Google Scholar]
- Jiang, M.; Chu, Z.Q. Comparative analysis of plant MKK gene family reveals novel expansion mechanism of the members and sheds new light on functional conservation. BMC Genom. 2018, 19, 407. [Google Scholar] [CrossRef] [Green Version]
- Jonak, C.; Okresz, L.; Bogre, L.; Hirt, H. Complexity, cross talk and integration of plant MAP kinase signalling. Curr. Opin. Plant Biol. 2002, 5, 415–424. [Google Scholar] [CrossRef]
- Jiang, M.; Wen, F.; Cao, J.M.; Li, P.; She, J.; Chu, Z.Q. Genome-wide exploration of the molecular evolution and regulatory network of mitogen-activated protein kinase cascades upon multiple stresses in Brachypodium distachyon. BMC Genom. 2015, 16, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, S.H.; Krysan, P.J. A double-mutant collection targeting MAP kinase related genes in Arabidopsis for studying genetic interactions. Plant J. 2016, 88, 867–878. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Yang, H.L.; Ren, D.T.; Li, Y. Activation of ZmMKK10, a maize mitogen-activated protein kinase kinase, induces ethylene-dependent cell death. Plant Sci. 2017, 264, 129–137. [Google Scholar] [CrossRef]
- Ma, H.G.; Chen, J.; Zhang, Z.Z.; Ma, L.; Yang, Z.Y.; Zhang, Q.L.; Li, X.H.; Xiao, J.H.; Wang, S.P. MAPK kinase 10.2 promotes disease resistance and drought tolerance by activating different MAPKs in rice. Plant J. 2017, 92, 557–570. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.; Li, P.; Wang, W. Comparative analysis of MAPK and MKK gene families reveals differential evolutionary patterns in Brachypodium distachyon inbred lines. PeerJ 2021, 9, e11238. [Google Scholar] [CrossRef]
- Doczi, R.; Brader, G.; Pettko-Szandtner, A.; Rajh, I.; Djamei, A.; Pitzschke, A.; Teige, M.; Hirt, H. The Arabidopsis mitogen-activated protein kinase kinase MKK3 is upstream of group C mitogen-activated protein kinases and participates in pathogen signaling. Plant Cell 2007, 19, 3266–3279. [Google Scholar] [CrossRef] [Green Version]
- Chardin, C.; Schenk, S.T.; Hirt, H.; Colcombet, J.; Krapp, A. Review: Mitogen-activated protein kinases in nutritional signaling in Arabidopsis. Plant Sci. 2017, 260, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Wang, C.; Yang, B.; Wu, F.F.; Hao, X.Y.; Liang, W.W.; Niu, F.F.; Yan, J.L.; Zhang, H.F.; Wang, B.Y.; et al. Identification and functional analysis of mitogen-activated protein kinase kinase kinase (MAPKKK) genes in canola (Brassica napus L.). J. Exp. Bot. 2014, 65, 2171–2188. [Google Scholar] [CrossRef] [Green Version]
- Pitzschke, A.; Hirt, H. Mitogen-activated protein kinases and reactive oxygen species signaling in plants. Plant Physiol. 2006, 141, 351–356. [Google Scholar] [CrossRef] [Green Version]
- Overmyer, K.; Brosche, M.; Kangasjarvi, J. Reactive oxygen species and hormonal control of cell death. Trends Plant Sci. 2003, 8, 335–342. [Google Scholar] [CrossRef]
- Dat, J.F.; Pellinen, R.; Beeckman, T.; Van de Cotte, B.; Langebartels, C.; Kangasjarvi, J.; Inze, D.; Van Breusegem, F. Changes in hydrogen peroxide homeostasis trigger an active cell death process in tobacco. Plant J. 2003, 33, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Rentel, M.C.; Lecourieux, D.; Ouaked, F.; Usher, S.L.; Petersen, L.; Okamoto, H.; Knight, H.; Peck, S.C.; Grierson, C.S.; Hirt, H.; et al. OXI1 kinase is necessary for oxidative burst-mediated signalling in Arabidopsis. Nature 2004, 427, 858–861. [Google Scholar] [CrossRef] [PubMed]
- Nakagami, H.; Kiegerl, S.; Hirt, H. OMTK1, a novel MAPKKK, channels oxidative stress signaling through direct MAPK interaction. J. Biol. Chem. 2004, 279, 26959–26966. [Google Scholar] [CrossRef] [Green Version]
- Pei, Z.M.; Murata, Y.; Benning, G.; Thomine, S.; Klusener, B.; Allen, G.J.; Grill, E.; Schroeder, J.I. Calcium channels activated by hydrogen peroxide mediate abscisic acid signalling in guard cells. Nature 2000, 406, 731–734. [Google Scholar] [CrossRef]
- Li, K.; Yang, F.B.; Zhang, G.Z.; Song, S.F.; Li, Y.; Ren, D.T.; Miao, Y.C.; Song, C.P. AIK1, A mitogen-activated protein kinase, modulates abscisic acid responses through the MKK5-MPK6 kinase cascade. Plant Physiol. 2017, 173, 1391–1408. [Google Scholar] [CrossRef] [Green Version]
- Bergmann, D.C.; Lukowitz, W.; Somerville, C.R. Stomatal development and pattern controlled by a MAPKK kinase. Science 2004, 304, 1494–1497. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka, H.; Asai, S.; Yoshioka, M.; Kobayashi, M. Molecular mechanisms of generation for nitric oxide and reactive oxygen species, and role of the radical burst in plant immunity. Mol. Cells 2009, 28, 321–329. [Google Scholar] [CrossRef]
- Takahashi, F.; Mizoguchi, T.; Yoshida, R.; Ichimura, K.; Shinozaki, K. Calmodulin-dependent activation of MAP kinase for ROS homeostasis in Arabidopsis. Mol. Cell 2011, 41, 649–660. [Google Scholar] [CrossRef]
- Li, C.H.; Wang, G.; Zhao, J.L.; Zhang, L.Q.; Ai, L.F.; Han, Y.F.; Sun, D.Y.; Zhang, S.W.; Sun, Y. The receptor-like kinase SIT1 mediates salt sensitivity by activating MAPK3/6 and regulating ethylene homeostasis in rice. Plant Cell 2014, 26, 2538–2553. [Google Scholar] [CrossRef] [Green Version]
- Mira, M.M.; Ibrahim, S.; Hill, R.D.; Stasolla, C. Cold stress in maize (Zea mays) is alleviated by the over-expression of Phytoglobin 1 (ZmPgb1.1). Plant Physiol. Biochem. 2021, 167, 901–910. [Google Scholar] [CrossRef]
- Shi, M.; Wang, Z.; Ma, Z.; Song, W.; Lu, W.; Xiao, K. Characterization on TaMPK14, an MAPK family gene of wheat, in modulating N-starvation response through regulating N uptake and ROS homeostasis. Plant Cell Rep. 2020, 39, 1285–1299. [Google Scholar] [CrossRef] [PubMed]
- Joo, S.; Liu, Y.; Lueth, A.; Zhang, S.Q. MAPK phosphorylation-induced stabilization of ACS6 protein is mediated by the non-catalytic C-terminal domain, which also contains the cis-determinant for rapid degradation by the 26S proteasome pathway. Plant J. 2008, 54, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.D.; Zhang, S.Q. Phosphorylation of 1-aminocyclopropane-1-carboxylic acid synthase by MPK6, a stress-responsive mitogen-activated protein kinase, induces ethylene biosynthesis in Arabidopsis. Plant Cell 2004, 16, 3386–3399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwikow, A.; Ciesla, A.; Kasprowicz-Maluski, A.; Mitula, F.; Tajdel, M.; Galganski, L.; Ziolkowski, P.A.; Kubiak, P.; Maleck, A.; Piechalak, A.; et al. Arabidopsis protein phosphatase 2C ABI1 interacts with type I ACC synthases and is involved in the regulation of ozone-induced ethylene biosynthesis. Mol Plant 2014, 7, 960–976. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.D.; Cho, Y.H.; Tena, G.; Xiong, Y.; Sheen, J. Dual control of nuclear EIN3 by bifurcate MAPK cascades in C2H4 signalling. Nature 2008, 451, 789–795. [Google Scholar] [CrossRef]
- Li, S.; Han, X.F.; Yang, L.Y.; Deng, X.X.; Wu, H.J.; Zhang, M.M.; Liu, Y.D.; Zhang, S.Q.; Xu, J. Mitogen-activated protein kinases and calcium-dependent protein kinases are involved in wounding-induced ethylene biosynthesis in Arabidopsis thaliana. Plant Cell Environ. 2018, 41, 134–147. [Google Scholar] [CrossRef]
- Chen, J.W.; Zhang, Q.; Wang, Q.G.; Feng, M.; Li, Y.; Meng, Y.L.; Zhang, Y.; Liu, G.Q.; Ma, Z.M.; Wu, H.Z.; et al. RhMKK9, a rose MAP KINASE KINASE gene, is involved in rehydration-triggered ethylene production in rose gynoecia. BMC Plant Biol. 2017, 17, 51. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, F.; Yoshida, R.; Ichimura, K.; Mizoguchi, T.; Seo, S.; Yonezawa, M.; Maruyama, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. The mitogen-activated protein kinase cascade MKK3-MPK6 is an important part of the jasmonate signal transduction pathway in Arabidopsis. Plant Cell 2007, 19, 805–818. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Masia, D.; Perez-Amador, M.A.; Carbonell, J.; Marcote, M.J. Diverse stress signals activate the C1 subgroup MAP kinases of Arabidopsis. FEBS Lett. 2007, 581, 1834–1840. [Google Scholar] [CrossRef] [Green Version]
- Sozen, C.; Schenk, S.T.; Boudsocq, M.; Chardin, C.; Almeida-Trapp, M.; Krapp, A.; Hirt, H.; Mithofer, A.; Colcombet, J. Wounding and insect feeding trigger two independent MAPK pathways with distinct regulation and kinetics. Plant Cell 2020, 32, 1988–2003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Kim, Y.J.; Kim, M.H.; Kwak, J.M. MAPK cascades in guard cell signal transduction. Front Plant Sci. 2016, 7, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Jwa, N.S. The rice MAPKK-MAPK interactome: The biological significance of MAPK components in hormone signal transduction. Plant Cell Rep. 2013, 32, 923–931. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.T.; Cheng, T.C.; Wang, G.H.; Yan, Y.F.; Xia, Q.Y. Cloning and evolutionary analysis of tobacco MAPK gene family. Mol. Biol. Rep. 2013, 40, 1407–1415. [Google Scholar] [CrossRef]
- Brodersen, P.; Petersen, M.; Nielsen, H.B.; Zhu, S.J.; Newman, M.A.; Shokat, K.M.; Rietz, S.; Parker, J.; Mundy, J. Arabidopsis MAP kinase 4 regulates salicylic acid- and jasmonic acid/ethylene-dependent responses via EDS1 and PAD4. Plant J. 2006, 47, 532–546. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Shao, F.; Cui, H.; Chen, L.J.; Li, H.T.; Zou, Y.; Long, C.Z.; Lan, L.F.; Chai, J.J.; Chen, S.; et al. A Pseudomonas syringae effector inactivates MAPKs to suppress PAMP-Induced immunity in plants. Cell Host Microbe 2007, 1, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Frey, N.F.D.; Garcia, A.V.; Bigeard, J.; Zaag, R.; Bueso, E.; Garmier, M.; Pateyron, S.; de Tauzia-Moreau, M.L.; Brunaud, V.; Balzergue, S.; et al. Functional analysis of Arabidopsis immune-related MAPKs uncovers a role for MPK3 as negative regulator of inducible defences. Genome Biol. 2014, 15, R87. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Chang, Y.; Zhao, C.C.; Yang, H.L.; Ren, D.T. Expression of the inactive ZmMEK1 induces salicylic acid accumulation and salicylic acid-dependent leaf senescence. J. Integr. Plant Biol. 2016, 58, 724–736. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Wang, H.Z.; Li, B.H.; Huang, J.; Liu, X.F.; Zhou, Y.H.; Mou, Z.L.; Li, J.Y. Increased expression of MAP KINASE KINASE7 causes deficiency in polar auxin transport and leads to plant architectural abnormality in Arabidopsis. Plant Cell 2006, 18, 308–320. [Google Scholar] [CrossRef] [Green Version]
- Dory, M.; Hatzimasoura, E.; Kallai, B.M.; Nagy, S.K.; Jager, K.; Darula, Z.; Nadai, T.V.; Meszaros, T.; Lopez-Juez, E.; Barnabas, B.; et al. Coevolving MAPK and PID phosphosites indicate an ancient environmental control of PIN auxin transporters in land plants. Febs Lett. 2018, 592, 89–102. [Google Scholar] [CrossRef]
- Enders, T.A.; Frick, E.M.; Strader, L.C. An Arabidopsis kinase cascade influences auxin-responsive cell expansion. Plant J. 2017, 92, 68–81. [Google Scholar] [CrossRef] [PubMed]
- Danquah, A.; de Zelicourt, A.; Boudsocq, M.; Neubauer, J.; Frey, N.F.D.; Leonhardt, N.; Pateyron, S.; Gwinner, F.; Tamby, J.P.; Ortiz-Masia, D.; et al. Identification and characterization of an ABA-activated MAP kinase cascade in Arabidopsis thaliana. Plant J. 2015, 82, 232–244. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.T.C.; Lee, S.J.; Choi, S.W.; Na, Y.J.; Song, M.R.; Hoang, Q.T.N.; Sim, S.Y.; Kim, M.S.; Kim, J.I.; Soh, M.S.; et al. Arabidopsis Raf-like kinase Raf10 is a regulatory component of core ABA signaling. Mol. Cells 2019, 42, 646–660. [Google Scholar] [PubMed]
- Zhao, L.; Yan, J.; Xiang, Y.; Sun, Y.; Zhang, A. ZmWRKY104 transcription factor phosphorylated by ZmMPK6 functioning in ABA-induced antioxidant defense and enhance drought tolerance in Maize. Biology 2021, 10, 893. [Google Scholar] [CrossRef] [PubMed]
- Asai, T.; Tena, G.; Plotnikova, J.; Willmann, M.R.; Chiu, W.L.; Gomez-Gomez, L.; Boller, T.; Ausubel, F.M.; Sheen, J. MAP kinase signalling cascade in Arabidopsis innate immunity. Nature 2002, 415, 977–983. [Google Scholar] [CrossRef]
- Droillard, M.J.; Boudsocq, M.; Barbier-Brygoo, H.; Lauriere, C. Involvement of MPK4 in osmotic stress response pathways in cell suspensions and plantlets of Arabidopsis thaliana: Activation by hypoosmolarity and negative role in hyperosmolarity tolerance. FEBS Lett. 2004, 574, 42–48. [Google Scholar] [CrossRef]
- Qiu, J.L.; Fiil, B.K.; Petersen, K.; Nielsen, H.B.; Botanga, C.J.; Thorgrimsen, S.; Palma, K.; Suarez-Rodriguez, M.C.; Sandbech-Clausen, S.; Lichota, J.; et al. Arabidopsis MAP kinase 4 regulates gene expression through transcription factor release in the nucleus. EMBO J. 2008, 27, 2214–2221. [Google Scholar] [CrossRef] [Green Version]
- Rufian, J.S.; Rueda-Blanco, J.; Lopez-Marquez, D.; Macho, A.P.; Beuzon, C.R.; Ruiz-Albert, J. The bacterial effector HopZ1a acetylates MKK7 to suppress plant immunity. New Phytol. 2021, 231, 1138–1156. [Google Scholar] [CrossRef]
- Ren, D.; Liu, Y.; Yang, K.Y.; Han, L.; Mao, G.; Glazebrook, J.; Zhang, S. A fungal-responsive MAPK cascade regulates phytoalexin biosynthesis in Arabidopsis. Proc. Natl. Acad. Sci. USA 2008, 105, 5638–5643. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Li, Y.; Wang, Y.; Liu, H.; Lei, L.; Yang, H.; Liu, G.; Ren, D. Activation of MAPK kinase 9 induces ethylene and camalexin biosynthesis and enhances sensitivity to salt stress in Arabidopsis. J. Biol. Chem. 2008, 283, 26996–27006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asano, T.; Nguyen, T.H.; Yasuda, M.; Sidiq, Y.; Nishimura, K.; Nakashita, H.; Nishiuchi, T. Arabidopsis MAPKKK δ-1 is required for full immunity against bacterial and fungal infection. J. Exp. Bot. 2020, 71, 2085–2097. [Google Scholar] [CrossRef] [PubMed]
- Ueno, Y.; Yoshida, R.; Kishi-Kaboshi, M.; Matsushita, A.; Jiang, C.J.; Goto, S.; Takahashi, A.; Hirochika, H.; Takatsuji, H. Abiotic stresses antagonize the rice defence pathway through the tyrosine-dephosphorylation of OsMPK6. PLoS Pathog. 2015, 11, 1005231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Wang, G.; Zhang, C.; Zhu, P.K.; Dai, H.L.; Yu, N.; He, Z.H.; Xu, L.; Wang, E.T. OsCERK1-mediated chitin perception and immune signaling requires receptor-like cytoplasmic kinase 185 to activate an MAPK cascade in rice. Mol. Plant 2017, 10, 619–633. [Google Scholar] [CrossRef] [Green Version]
- Jalmi, S.K.; Sinha, A.K. Functional involvement of a mitogen activated protein kinase module, OsMKK3-OsMPK7-OsWRK30 in mediating resistance against Xanthomonas oryzae in rice. Sci. Rep. 2016, 6, 37974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Wang, L.; Yang, Z.; Liu, H.; Chu, C.; Zhang, Z.; Zhang, Q.; Li, X.; Xiao, J.; Wang, S.; et al. The rice Raf-like MAPKKK OsILA1 confers broad-spectrum resistance to bacterial blight by suppressing the OsMAPKK4-OsMAPK6 cascade. J. Integr. Plant Biol. 2021, 63, 1815–1842. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Yang, Z.; Li, J.; Xie, W.; Qin, X.; Kang, Y.; Zhang, Q.; Li, X.; Xiao, J.; Ma, H.; et al. Two VQ proteins are substrates of the OsMPKK6-OsMPK4 cascade in rice defense against bacterial blight. Rice 2021, 14, 39. [Google Scholar] [CrossRef]
- Hong, Y.; Liu, Q.; Cao, Y.; Zhang, Y.; Chen, D.; Lou, X.; Cheng, S.; Cao, L. The OsMPK15 negatively regulates Magnaporthe oryza and Xoo disease resistance via SA and JA signaling pathway in rice. Front. Plant Sci. 2019, 10, 752. [Google Scholar] [CrossRef]
- Dangol, S.; Nguyen, N.K.; Singh, R.; Chen, Y.; Wang, J.; Lee, H.G.; Hwang, B.K.; Jwa, N.S. Mitogen-activated protein kinase OsMEK2 and OsMPK1 signaling is required for ferroptotic cell death in rice-Magnaporthe oryzae interactions. Front. Plant Sci. 2021, 12, 710794. [Google Scholar] [CrossRef]
- Wang, C.; Guo, H.; He, X.; Zhang, S.; Wang, J.; Wang, L.; Guo, D.; Guo, X. Scaffold protein GhMORG1 enhances the resistance of cotton to Fusarium oxysporum by facilitating the MKK6-MPK4 cascade. Plant Biotechnol. J. 2020, 18, 1421–1433. [Google Scholar] [CrossRef] [Green Version]
- Teige, M.; Scheikl, E.; Eulgem, T.; Doczi, F.; Ichimura, K.; Shinozaki, K.; Dangl, J.L.; Hirt, H. The MKK2 pathway mediates cold and salt stress signaling in Arabidopsis. Mol. Cell 2004, 15, 141–152. [Google Scholar] [CrossRef]
- Ponce-Pineda, I.G.; Carmona-Salazar, L.; Saucedo-Garcia, M.; Cano-Ramirez, D.; Morales-Cedillo, F.; Pena-Moral, A.; Guevara-Garcia, A.A.; Sanchez-Nieto, S.; Gavilanes-Ruiz, M. MPK6 kinase regulates plasma membrane H+-ATPase activity in cold acclimation. Int. J. Mol. Sci. 2021, 22, 6338. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Wang, P.; Si, T.; Hsu, C.C.; Wang, L.; Zayed, O.; Yu, Z.; Zhu, Y.; Dong, J.; Tao, W.A.; et al. MAP kinase cascades regulate the cold response by modulating ICE1 protein stability. Dev. Cell 2017, 43, 618–629.e615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Ding, Y.; Shi, Y.; Zhang, X.; Zhang, S.; Gong, Z.; Yang, S. MPK3- and MPK6-mediated ICE1 phosphorylation negatively regulates ICE1 stability and freezing tolerance in Arabidopsis. Dev. Cell 2017, 43, 630–642.e634. [Google Scholar] [CrossRef] [PubMed]
- Ichimura, K.; Mizoguchi, T.; Yoshida, R.; Yuasa, T.; Shinozaki, K. Various abiotic stresses vapidly activate Arabidopsis MAP kinases ATMPK4 and ATMPK6. Plant J. 2000, 24, 655–665. [Google Scholar] [CrossRef]
- Ning, J.; Li, X.H.; Hicks, L.M.; Xiong, L.Z. A Raf-Like MAPKKK gene DSM1 mediates drought resistance through reactive oxygen species scavenging in rice. Plant Physiol. 2010, 152, 876–890. [Google Scholar] [CrossRef] [Green Version]
- Zong, X.J.; Li, D.P.; Gu, L.K.; Li, D.Q.; Liu, L.X.; Hu, X.L. Abscisic acid and hydrogen peroxide induce a novel maize group C MAP kinase gene, ZmMPK7, which is responsible for the removal of reactive oxygen species. Planta 2009, 229, 485–495. [Google Scholar] [CrossRef]
- Kumar, R.R.; Arora, K.; Goswami, S.; Sakhare, A.; Singh, B.; Chinnusamy, V.; Praveen, S. MAPK enzymes: A ROS activated signaling sensors involved in modulating heat stress response, tolerance and grain stability of wheat under heat stress. 3 Biotech 2020, 10, 380. [Google Scholar] [CrossRef]
- Li, Y.Y.; Cai, H.X.; Liu, P.; Wang, C.Y.; Gao, H.; Wu, C.G.; Yan, K.; Zhang, S.Z.; Huang, J.G.; Zheng, C.C. Arabidopsis MAPKKK18 positively regulates drought stress resistance via downstream MAPKK3. Biochem. Biophys. Res. Commun. 2017, 484, 292–297. [Google Scholar] [CrossRef]
- Weng, C.M.; Lu, J.X.; Wan, H.F.; Wang, S.W.; Wang, Z.; Lu, K.; Liang, Y. Over-expression of BnMAPK1 in Brassica napus enhances tolerance to drought stress. J. Integr. Agr. 2014, 13, 2407–2415. [Google Scholar] [CrossRef]
- Cai, G.H.; Wang, G.D.; Wang, L.; Liu, Y.; Pan, J.W.; Li, D.Q. A maize mitogen-activated protein kinase kinase, ZmMKK1, positively regulated the salt and drought tolerance in transgenic Arabidopsis. J. Plant Physiol. 2014, 171, 1003–1016. [Google Scholar] [CrossRef]
- Jia, H.H.; Hao, L.L.; Guo, X.L.; Liu, S.C.; Yan, Y.; Guo, X.Q. A Raf-like MAPKKK gene, GhRaf19, negatively regulates tolerance to drought and salt and positively regulates resistance to cold stress by modulating reactive oxygen species in cotton. Plant Sci. 2016, 252, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Na, Y.J.; Choi, H.K.; Park, M.Y.; Choi, S.W.; Xuan Vo, K.T.; Jeon, J.S.; Kim, S.Y. OsMAPKKK63 is involved in salt stress response and seed dormancy control. Plant Signal. Behav. 2019, 14, e1578633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Sun, H.; Wang, F.; Yue, D.; Shen, X.; Sun, W.; Zhang, X.; Yang, X. Genome-wide identification of MAPK cascade genes reveals the GhMAP3K14-GhMKK11-GhMPK31 pathway is involved in the drought response in cotton. Plant Mol. Biol. 2020, 103, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.B.; Wang, X.P.; Wang, Y.C.; Chen, Y.H.; Luo, J.W.; Li, D.D.; Li, X.B. Genome-wide identification and functional characterization of cotton (Gossypium hirsutum) MAPKKK gene family in response to drought stress. BMC Plant Biol. 2020, 20, 217. [Google Scholar] [CrossRef]
- Lin, Z.; Li, Y.; Zhang, Z.; Liu, X.; Hsu, C.C.; Du, Y.; Sang, T.; Zhu, C.; Wang, Y.; Satheesh, V.; et al. A RAF-SnRK2 kinase cascade mediates early osmotic stress signaling in higher plants. Nat. Commun. 2020, 11, 613. [Google Scholar] [CrossRef] [Green Version]
- Soma, F.; Takahashi, F.; Suzuki, T.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Plant Raf-like kinases regulate the mRNA population upstream of ABA-unresponsive SnRK2 kinases under drought stress. Nat. Commun. 2020, 11, 1373. [Google Scholar] [CrossRef] [Green Version]
- Xia, C.; Gong, Y.; Chong, K.; Xu, Y. Phosphatase OsPP2C27 directly dephosphorylates OsMAPK3 and OsbHLH002 to negatively regulate cold tolerance in rice. Plant Cell Environ. 2021, 44, 491–505. [Google Scholar] [CrossRef]
- Tak, H.; Negi, S.; Rajpurohit, Y.S.; Misra, H.S.; Ganapathi, T.R. MusaMPK5, a mitogen activated protein kinase is involved in regulation of cold tolerance in banana. Plant Physiol. Biochem. 2020, 146, 112–123. [Google Scholar] [CrossRef]
- Nishihama, R.; Ishikawa, M.; Araki, S.; Soyano, T.; Asada, T.; Machida, Y. The NPK1 mitogen-activated protein kinase kinase kinase is a regulator of cell-plate formation in plant cytokinesis. Gene Dev. 2001, 15, 352–363. [Google Scholar] [CrossRef] [Green Version]
- Soyano, T.; Nishihama, R.; Morikiyo, K.; Ishikawa, M.; Machida, Y. NQK1/NtMEK1 is a MAPKK that acts in the NPK1 MAPKKK-mediated MAPK cascade and is required for plant cytokinesis. Gene Dev. 2003, 17, 1055–1067. [Google Scholar] [CrossRef] [Green Version]
- Nishihama, R.; Soyano, T.; Ishikawa, M.; Araki, S.; Tanaka, H.; Asada, T.; Irie, K.; Ito, M.; Terada, M.; Banno, H.; et al. Expansion of the cell plate in plant cytokinesis requires a kinesin-like protein/MAPKKK complex. Cell 2002, 109, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, Y.; Soyano, T.; Kosetsu, K.; Sasabe, M.; Machida, Y. HINKEL kinesin, ANP MAPKKKs and MKK6/ANQ MAPKK, which phosphorylates and activates MPK4 MAPK, constitute a pathway that is required for cytokinesis in Arabidopsis thaliana. Plant Cell Physiol. 2010, 51, 1766–1776. [Google Scholar] [CrossRef] [Green Version]
- Beck, M.; Komis, G.; Muller, J.; Menzel, D.; Samaj, J. Arabidopsis homologs of nucleus- and phragmoplast-localized kinase 2 and 3 and mitogen-activated protein kinase 4 are essential for microtubule organization. Plant Cell 2010, 22, 755–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vavrdova, T.; Samaj, J.; Komis, G. Phosphorylation of plant microtubule-associated proteins during cell division. Front. Plant Sci. 2019, 10, 238. [Google Scholar] [CrossRef]
- Meng, X.Z.; Wang, H.C.; He, Y.X.; Liu, Y.D.; Walker, J.C.; Torii, K.U.; Zhang, S.Q. A MAPK cascade downstream of ERECTA receptor-like protein kinase regulates Arabidopsis inflorescence architecture by promoting localized cell proliferation. Plant Cell 2012, 24, 4948–4960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smekalova, V.; Luptovciak, I.; Komis, G.; Samajova, O.; Ovecka, M.; Doskocilova, A.; Takac, T.; Vadovic, P.; Novak, O.; Pechan, T.; et al. Involvement of YODA and mitogen activated protein kinase 6 in Arabidopsis post-embryogenic root development through auxin up-regulation and cell division plane orientation. New Phytol. 2014, 203, 1175–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, D.S.; Cha, D.S.; Choi, Y.; Lee, J.W.; Lee, M.H. MPK-1/ERK is required for the full activity of resveratrol in extended lifespan and reproduction. Aging Cell 2019, 18, e12867. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Shi, H.; Ou, Y.; Cui, Y.; Chang, J.; Peng, L.; Gou, X.; He, K.; Li, J. RGF1-RGI1, a peptide-receptor complex, regulates Arabidopsis root meristem development via a MAPK signaling cascade. Mol Plant 2020, 13, 1594–1607. [Google Scholar] [CrossRef]
- Shao, Y.; Yu, X.; Xu, X.; Li, Y.; Yuan, W.; Xu, Y.; Mao, C.; Zhang, S.; Xu, J. The YDA-MKK4/MKK5-MPK3/MPK6 cascade functions downstream of the RGF1-RGI ligand-receptor pair in regulating mitotic activity in root apical meristem. Mol. Plant 2020, 13, 1608–1623. [Google Scholar] [CrossRef]
- Wang, H.C.; Ngwenyama, N.; Liu, Y.D.; Walker, J.C.; Zhang, S.Q. Stomatal development and patterning are regulated by environmentally responsive mitogen-activated protein kinases in Arabidopsis. Plant Cell 2007, 19, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Lampard, G.R.; Wengier, D.L.; Bergmann, D.C. Manipulation of mitogen-activated protein kinase kinase signaling in the Arabidopsis stomatal lineage reveals motifs that contribute to protein localization and signaling specificity. Plant Cell 2014, 26, 3358–3371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walia, A.; Lee, J.S.; Wasteneys, G.; Ellis, B. Arabidopsis mitogen-activated protein kinase MPK18 mediates cortical microtubule functions in plant cells. Plant J. 2009, 59, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Tanoue, T.; Adachi, M.; Moriguchi, T.; Nishida, E. A conserved docking motif in MAP kinases common to substrates, activators and regulators. Nat. Cell Biol. 2000, 2, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Tanoue, T.; Nishida, E. Molecular recognitions in the MAP kinase cascades. Cell Signal. 2003, 15, 455–462. [Google Scholar] [CrossRef]
- Bardwell, A.J.; Frankson, E.; Bardwell, L. Selectivity of docking sites in MAPK kinases. J. Biol. Chem. 2009, 284, 13165–13173. [Google Scholar] [CrossRef] [Green Version]
- Bardwell, L.; Shah, K. Analysis of mitogen-activated protein kinase activation and interactions with regulators and substrates. Methods 2006, 40, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Bardwell, A.J.; Bardwell, L. Two hydrophobic residues can determine the specificity of mitogen-activated protein kinase docking interactions. J. Biol. Chem. 2015, 290, 26661–26674. [Google Scholar] [CrossRef] [Green Version]
- Peti, W.; Page, R. Molecular basis of MAP kinase regulation. Protein Sci. 2013, 22, 1698–1710. [Google Scholar] [CrossRef]
- Alexa, A.; Ember, O.; Szabo, I.; Mo’ath, Y.; Poti, A.L.; Remenyi, A.; Banoczi, Z. Peptide based inhibitors of protein binding to the mitogen-activated protein kinase docking groove. Front. Mol. Biosci. 2021, 8, 690429. [Google Scholar] [CrossRef]
- Lee, T.; Hoofnagle, A.N.; Kabuyama, Y.; Stroud, J.; Min, X.; Goldsmith, E.J.; Chen, L.; Resing, K.A.; Ahn, N.G. Docking motif interactions in MAP kinases revealed by hydrogen exchange mass spectrometry. Mol. Cell 2004, 14, 43–55. [Google Scholar] [CrossRef]
- Palacios, L.; Dickinson, R.J.; Sacristan-Reviriego, A.; Didmon, M.P.; Marin, M.J.; Martin, H.; Keyse, S.M.; Molina, M. Distinct docking mechanisms mediate interactions between the Msg5 phosphatase and mating or cell integrity mitogen-activated protein kinases (MAPKs) in Saccharomyces cerevisiae. J. Biol. Chem. 2011, 286, 42037–42050. [Google Scholar] [CrossRef] [Green Version]
- Kaur, P.; Anand, A.; Bhat, A.; Maras, J.S.; Goyal, N. Comparative phosphoproteomic analysis unravels MAPK1 regulated phosphoproteins in Leishmania donovani. J. Proteom. 2021, 240, 104189. [Google Scholar] [CrossRef] [PubMed]
- Chi, Y.; Clurman, B.E. Mass spectrometry-based identification of protein kinase substrates utilizing engineered kinases and thiophosphate labeling. Curr. Protoc. Chem. Biol. 2010, 2, ch100151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trinh, T.B.; Xiao, Q.; Pei, D.H. Profiling the substrate specificity of protein kinases by on-bead screening of peptide libraries. Biochemistry 2013, 52, 5645–5655. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Jiang, S.; Yu, X.; Cheng, C.; Chen, S.X.; Cheng, Y.B.; Yuan, J.S.; Jiang, D.H.; He, P.; Shan, L.B. Phosphorylation of trihelix transcriptional repressor ASR3 by MAP KINASE4 negatively regulates Arabidopsis immunity. Plant Cell 2015, 27, 839–856. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.I.; Birendra, K.C.; Zhu, W.; Motamedchaboki, K.; Doye, V.; Roux, K.J. Probing nuclear pore complex architecture with proximity-dependent biotinylation. Proc. Natl. Acad. Sci. USA 2014, 111, E2453–E2461. [Google Scholar] [CrossRef] [Green Version]
- Kerbler, S.M.; Natale, R.; Fernie, A.R.; Zhang, Y. From affinity to proximity techniques to investigate protein complexes in plants. Int. J. Mol. Sci. 2021, 22, 7101. [Google Scholar] [CrossRef]
- Struk, S.; Jacobs, A.; Sanchez Martin-Fontecha, E.; Gevaert, K.; Cubas, P.; Goormachtig, S. Exploring the protein-protein interaction landscape in plants. Plant Cell Environ. 2019, 42, 387–409. [Google Scholar] [CrossRef]
- Lampugnani, E.R.; Wink, R.H.; Persson, S.; Somssich, M. The toolbox to study protein-protein interactions in plants. Crit. Rev. Plant Sci. 2018, 37, 308–334. [Google Scholar] [CrossRef]
- Pitzschke, A. Modes of MAPK substrate recognition and control. Trends Plant Sci. 2015, 20, 49–55. [Google Scholar] [CrossRef]
- Samakovli, D.; Ticha, T.; Vavrdova, T.; Ovecka, M.; Luptovciak, I.; Zapletalova, V.; Kucharova, A.; Krenek, P.; Krasylenko, Y.; Margaritopoulou, T.; et al. YODA-HSP90 module regulates phosphorylation-dependent inactivation of SPEECHLESS to control stomatal development under acute heat stress in Arabidopsis. Mol. Plant 2020, 13, 612–633. [Google Scholar] [CrossRef] [PubMed]
- Lampard, G.R.; MacAlister, C.A.; Bergmann, D.C. Arabidopsis stomatal initiation is controlled by MAPK-mediated regulation of the bHLH SPEECHLESS. Science 2008, 322, 1113–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bethke, G.; Unthan, T.; Uhrig, J.F.; Poschl, Y.; Gust, A.A.; Scheel, D.; Lee, J. Flg22 regulates the release of an ethylene response factor substrate from MAP kinase 6 in Arabidopsis thaliana via ethylene signaling. Proc. Natl. Acad. Sci. USA 2009, 106, 8067–8072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, Y.F.; Meng, X.Z.; Khanna, R.; LaMontagne, E.; Liu, Y.D.; Zhang, S.Q. Phosphorylation of a WRKY transcription factor by MAPKs is required for pollen development and function in Arabidopsis. PLoS Genet. 2014, 10, 1004384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Kim, H.S.; Bahk, S.; An, J.; Yoo, Y.; Kim, J.Y.; Chung, W.S. Phosphorylation of the transcriptional repressor MYB15 by mitogen-activated protein kinase 6 is required for freezing tolerance in Arabidopsis. Nucleic Acids Res. 2017, 45, 6613–6627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, X.; He, M.; Mei, E.; Zhang, B.; Tang, J.; Xu, M.; Liu, J.; Li, X.; Wang, Z.; Tang, W.; et al. WRKY53 integrates classic brassinosteroid signaling and the mitogen-activated protein kinase pathway to regulate rice architecture and seed size. Plant Cell 2021, 33, 2753–2775. [Google Scholar] [CrossRef]
- Smertenko, A.P.; Chang, H.Y.; Sonobe, S.; Fenyk, S.I.; Weingartner, M.; Bogre, L.; Hussey, P.J. Control of the AtMAP65-1 interaction with microtubules through the cell cycle. J. Cell Sci. 2006, 119, 3227–3237. [Google Scholar] [CrossRef] [Green Version]
- Sasabe, M.; Kosetsu, K.; Hidaka, M.; Murase, A.; Machida, Y. Arabidopsis thaliana MAP65-1 and MAP65-2 function redundantly with MAP65-3/PLEIADE in cytokinesis downstream of MPK4. Plant Signal. Behav. 2011, 6, 743–747. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Number | Group | Named Members | Active Sites * | References |
|---|---|---|---|---|---|
| MAPK | 20 | A | MPK3/6/10 | TEY, TQY | [5,6] |
| B | MPK4/5/11/12/13 | TEY, MEY, TVY, TEC | |||
| C | MPK1/2/7/14 | TEY | |||
| D | MPK8/9/15/16/17/18/19/20 | TDY | |||
| MAPKK | 10 | A | MKK1/2/6 | S/T-X5-S/T | [5,20] |
| B | MKK3 | S/T-X5-S/T | |||
| C | MKK4/5 | S/T-X5-S/T | |||
| D | MKK7/8/9 | S/T-X5-S/T | |||
| E | MKK10 | R-X5-S/T | |||
| MAPKKK | 80 | MEKK | MEKK1, YDA, ANP1/2/3 | G (T/S) Px (W/F) MAPEV | [21,22] |
| Raf | MKD1, EDR1, CTR1 | GTxx (W/Y) MAPE | |||
| ZIK | ZIK1 | GTPEFMAPE (L/V/M)(Y/F) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, M.; Zhang, Y.; Li, P.; Jian, J.; Zhao, C.; Wen, G. Mitogen-Activated Protein Kinase and Substrate Identification in Plant Growth and Development. Int. J. Mol. Sci. 2022, 23, 2744. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052744
Jiang M, Zhang Y, Li P, Jian J, Zhao C, Wen G. Mitogen-Activated Protein Kinase and Substrate Identification in Plant Growth and Development. International Journal of Molecular Sciences. 2022; 23(5):2744. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052744
Chicago/Turabian StyleJiang, Min, Youze Zhang, Peng Li, Jinjing Jian, Changling Zhao, and Guosong Wen. 2022. "Mitogen-Activated Protein Kinase and Substrate Identification in Plant Growth and Development" International Journal of Molecular Sciences 23, no. 5: 2744. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052744