
Compartmentalized Innate Immune Response of Human Fetal Membranes against Escherichia coli Choriodecidual Infection

 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
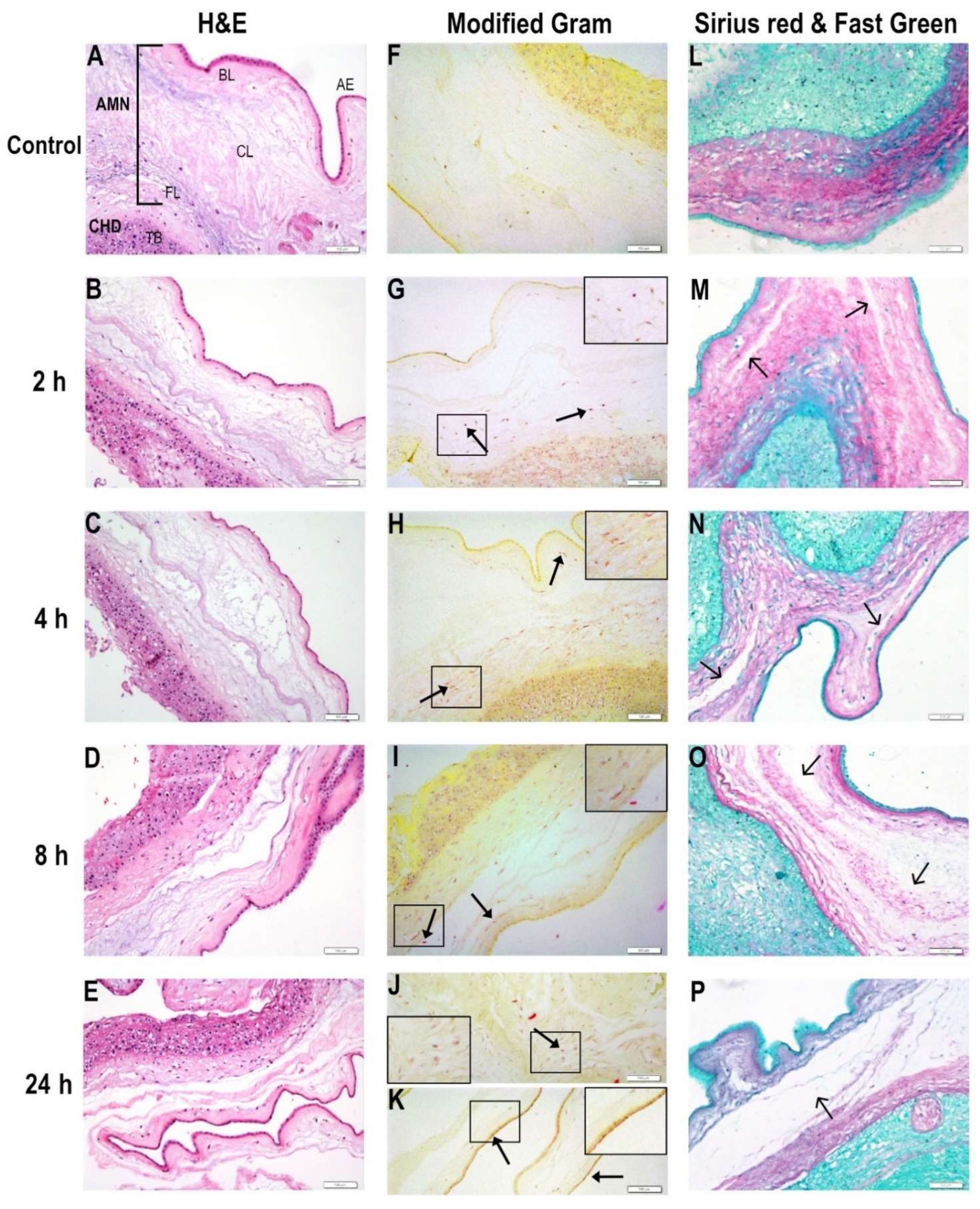
2.1. Bacterial Migration through Chorioamniotic Membranes and Their Associated Structural Damage
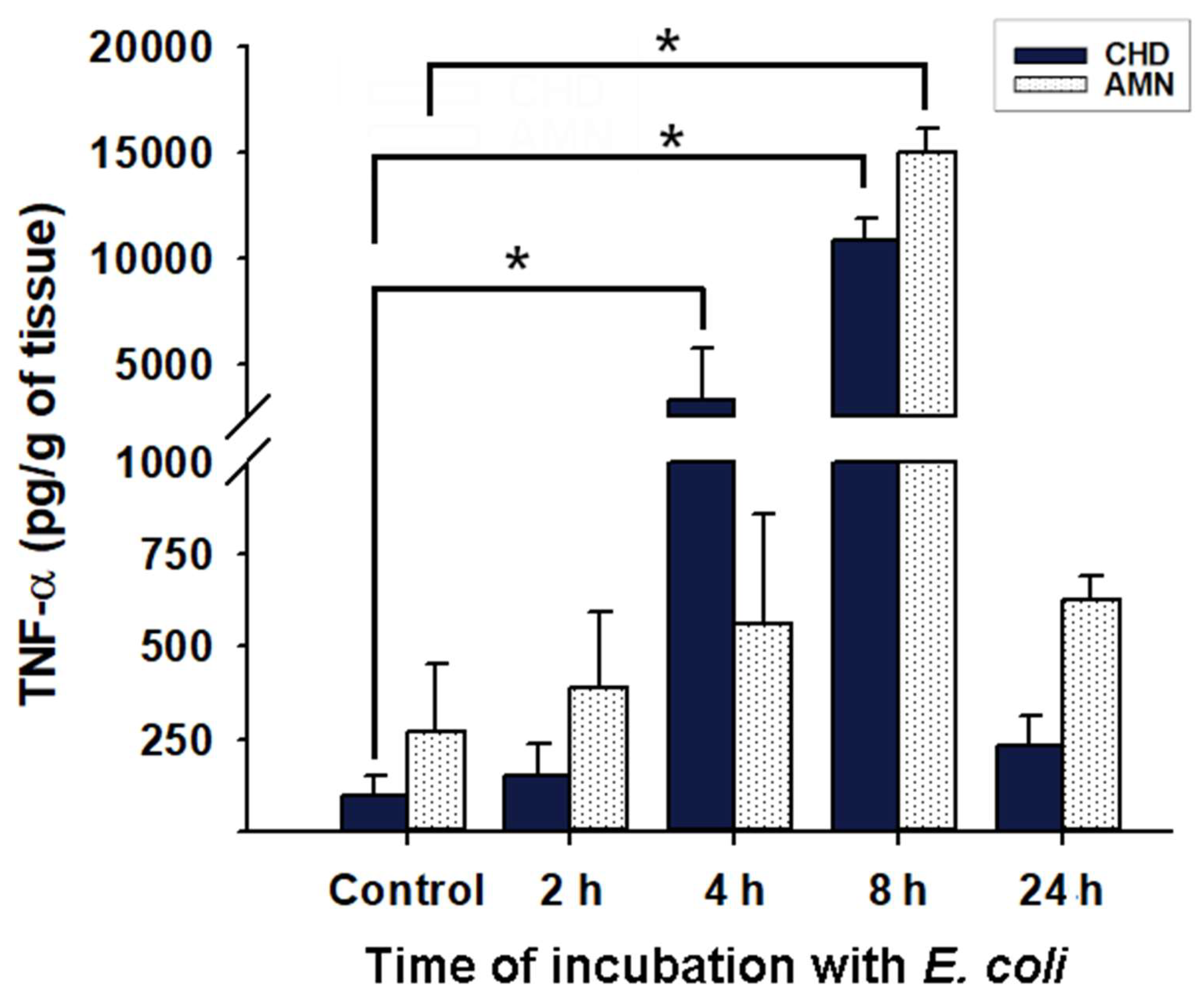
2.2. Inflammatory Milieu Induced by E. coli Infection in Chorioamniotic Membranes
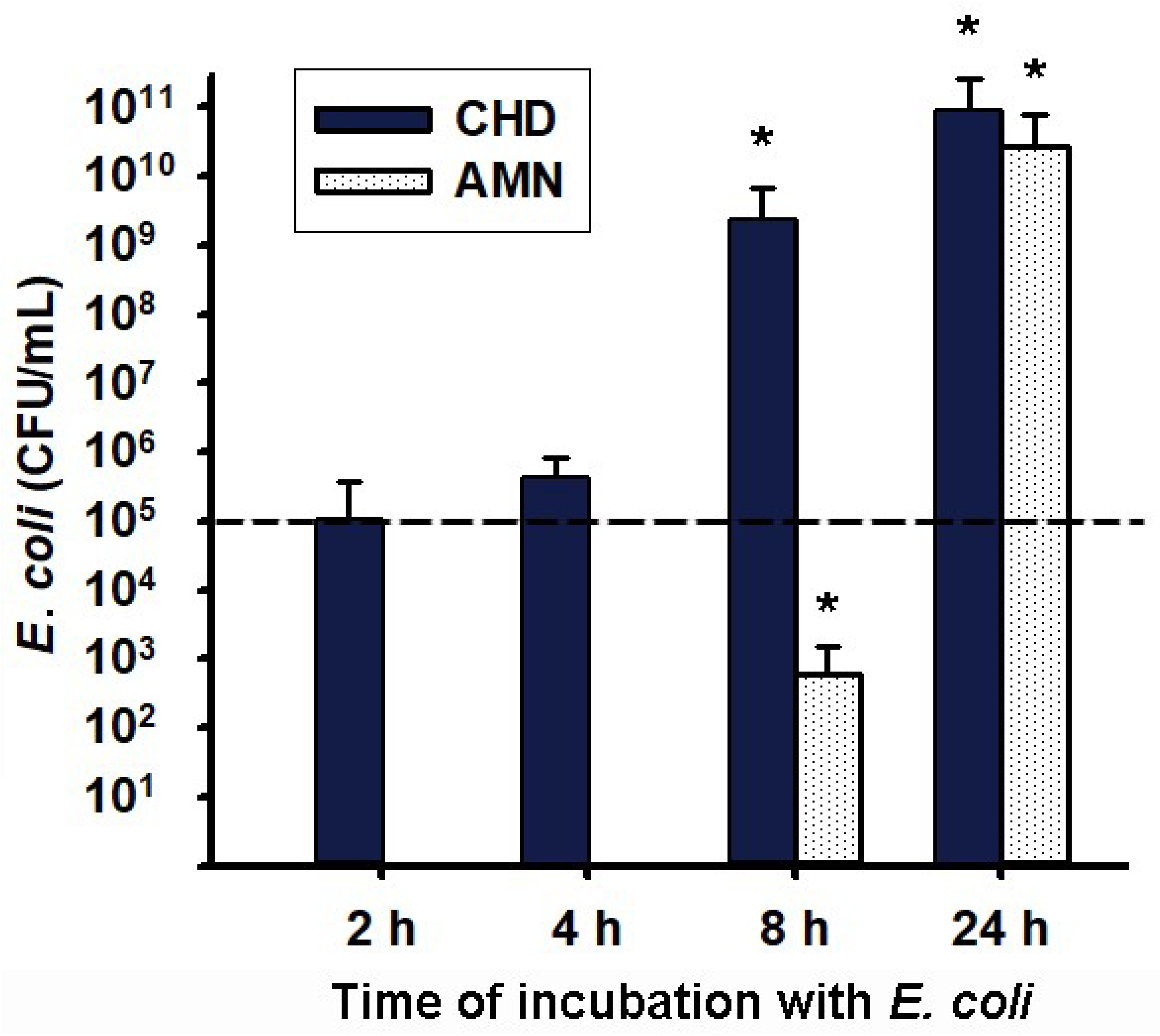
2.3. Transmembrane Migration of Bacteria from Choriodecidua to Amnion Media Culture
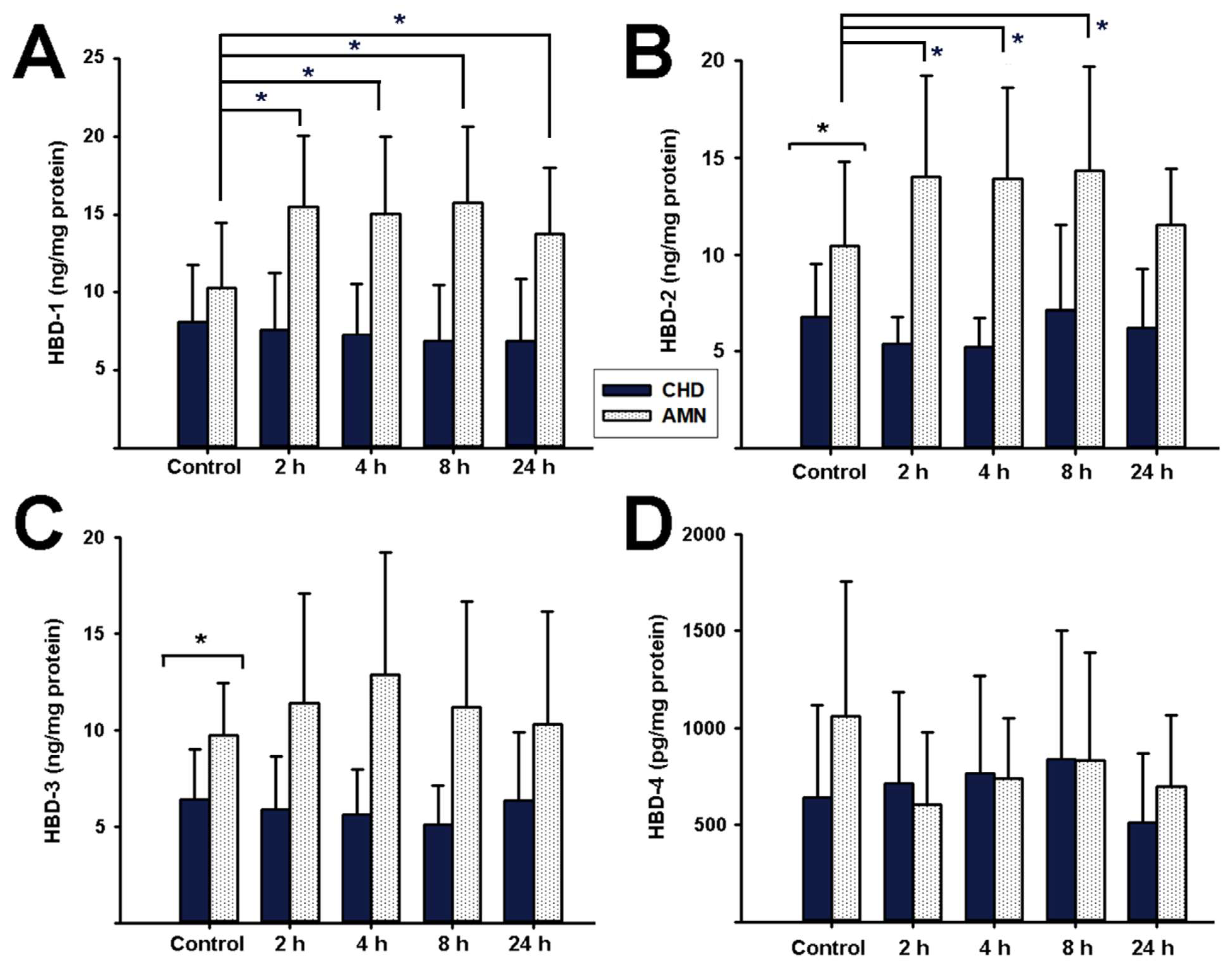
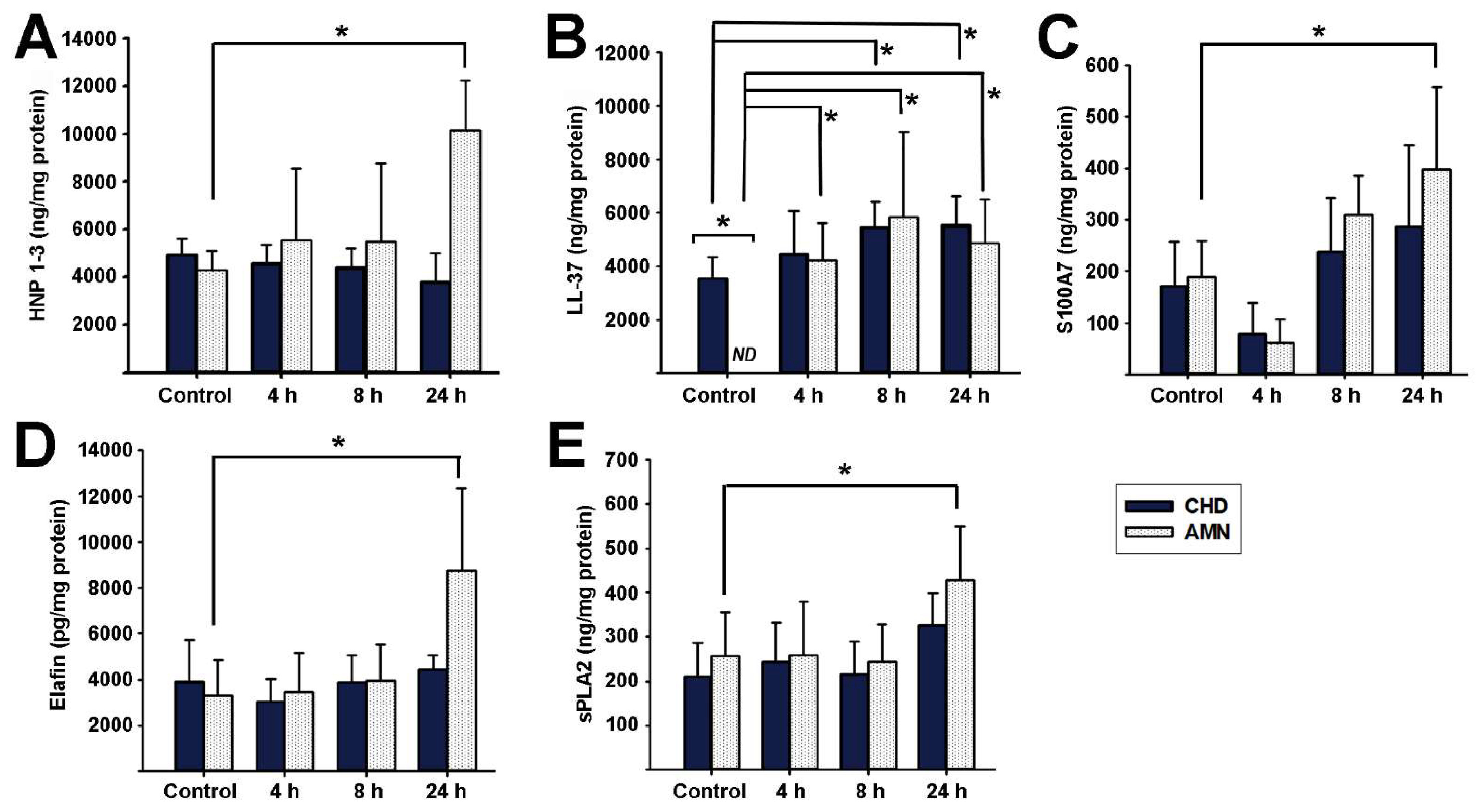
2.4. Antimicrobial Peptides Synthesis Profile in Response to E. coli Choriodecidual Infection
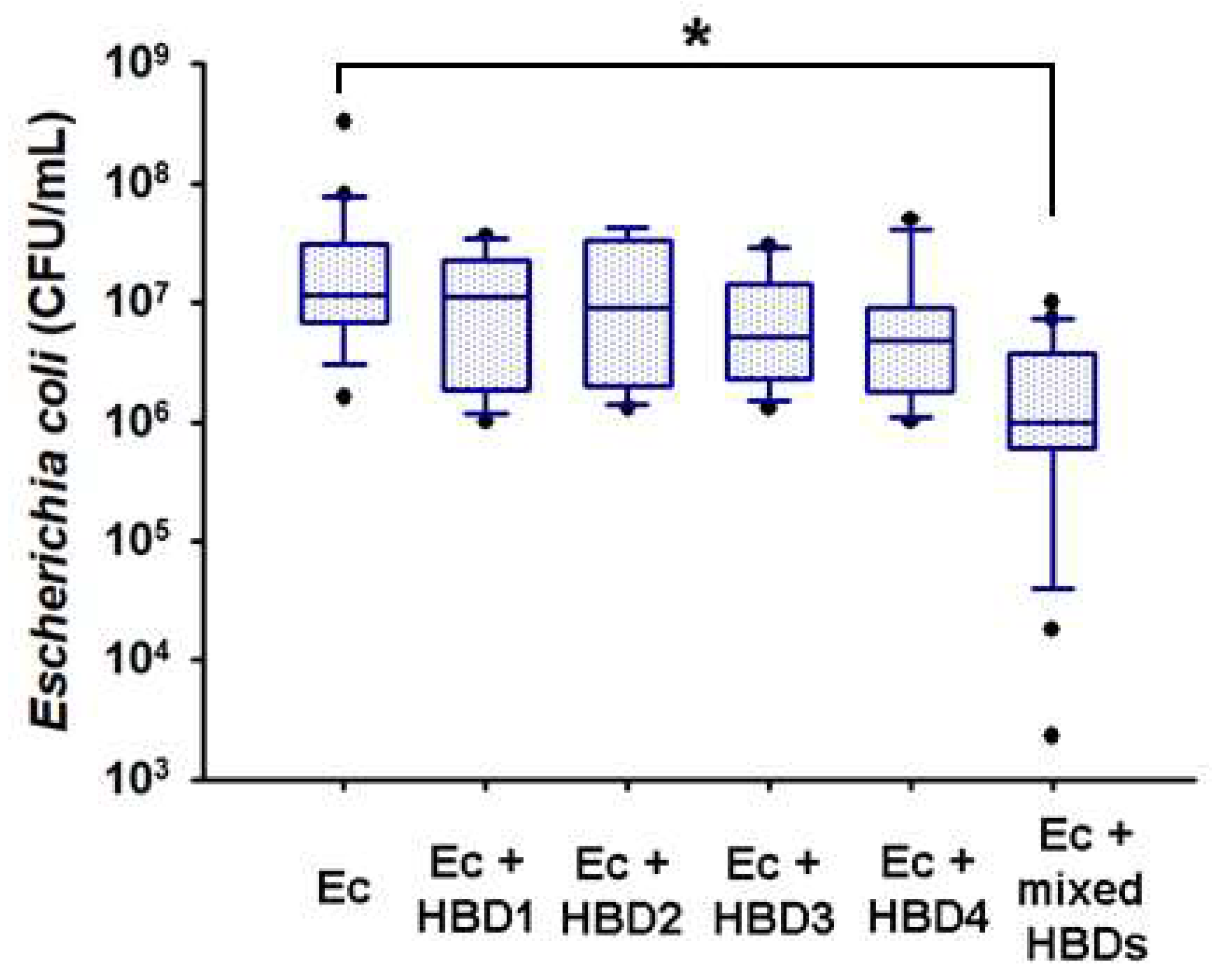
2.5. In Vitro Microbicidal Activity of Recombinant Human Beta Defensins
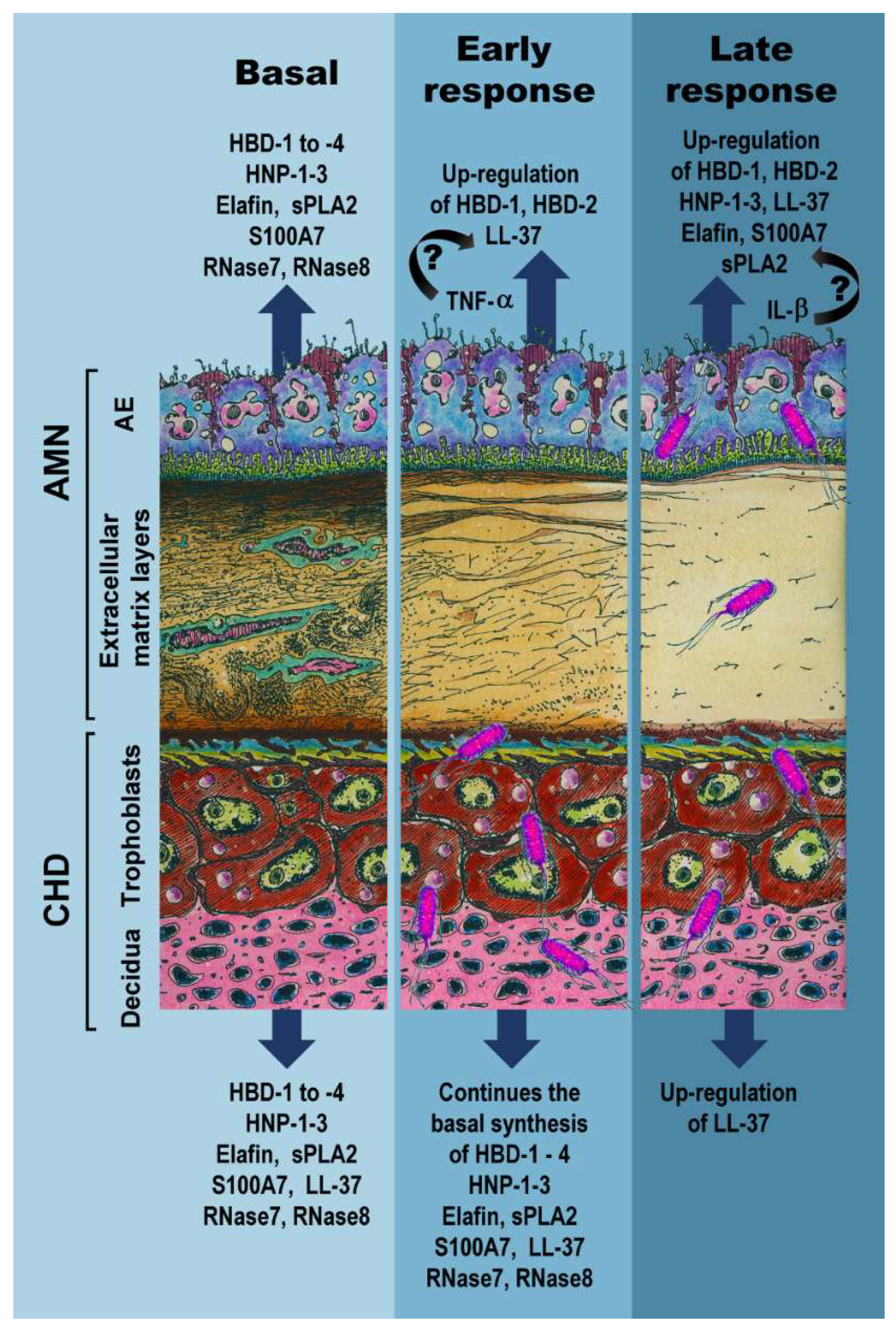
3. Discussion
4. Material and Methods
4.1. Ethics Statement
4.2. Biological Samples
4.3. Chorioamniotic Membrane Culture
4.4. Choriodecidual Infection with Escherichia coli
4.5. CFU Count and Bacterial Transmigration Assay in Cultured Fetal Membranes
4.6. Tissue Protein Extraction and Antimicrobial Peptide Quantification by ELISA
4.7. Cytokine Quantification by ELISA
4.8. Staining Techniques
4.9. Microscopy
4.10. In Vitro Microbicidal Activity of Recombinant HBDs
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chawanpaiboon, S.; Vogel, J.P.; Moller, A.-B.; Lumbiganon, P.; Petzold, M.; Hogan, D.; Landoulsi, S.; Jampathong, N.; Kongwattanakul, K.; Laopaiboon, M.; et al. Global, Regional, and National Estimates of Levels of Preterm Birth in 2014: A Systematic Review and Modelling Analysis. Lancet Glob. Health 2019, 7, e37–e46. [Google Scholar] [CrossRef] [Green Version]
- Crump, C. Preterm Birth and Mortality in Adulthood: A Systematic Review. J. Perinatol. 2020, 40, 833–843. [Google Scholar] [CrossRef] [PubMed]
- McDonald, H.M.; Chambers, H.M. Intrauterine Infection and Spontaneous Midgestation Abortion: Is the Spectrum of Microorganisms Similar to That in Preterm Labor? Infect. Dis. Obstet. Gynecol. 2000, 8, 220–227. [Google Scholar] [CrossRef]
- Sáez-López, E.; Guiral, E.; Fernández-Orth, D.; Villanueva, S.; Goncé, A.; López, M.; Teixidó, I.; Pericot, A.; Figueras, F.; Palacio, M.; et al. Vaginal versus Obstetric Infection Escherichia coli Isolates among Pregnant Women: Antimicrobial Resistance and Genetic Virulence Profile. PLoS ONE 2016, 11, e0146531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dautt-Leyva, J.G.; Canizalez-Román, A.; Acosta-Alfaro, L.F.; Gonzalez-Ibarra, F.; Murillo-Llanes, J. Maternal and Perinatal Complications in Pregnant Women with Urinary Tract Infection Caused by Escherichia coli. J. Obstet. Gynaecol. Res. 2018, 44, 1384–1390. [Google Scholar] [CrossRef]
- Bauer, M.E.; Housey, M.; Bauer, S.T.; Behrmann, S.; Chau, A.; Clancy, C.; Clark, E.A.S.; Einav, S.; Langen, E.; Leffert, L.; et al. Risk Factors, Etiologies, and Screening Tools for Sepsis in Pregnant Women: A Multicenter Case-Control Study. Anesth. Analg. 2019, 129, 1613–1620. [Google Scholar] [CrossRef]
- Li, Y.Y.; Kong, C.W.; To, W.W.K. Pathogens in Preterm Prelabour Rupture of Membranes and Erythromycin for Antibiotic Prophylaxis: A Retrospective Analysis. Hong Kong Med. J. 2019, 25, 287–294. [Google Scholar] [CrossRef] [Green Version]
- Page, J.M.; Bardsley, T.; Thorsten, V.; Allshouse, A.A.; Varner, M.W.; Debbink, M.P.; Dudley, D.J.; Saade, G.R.; Goldenberg, R.L.; Stoll, B.; et al. Stillbirth Associated with Infection in a Diverse, U.S. Cohort. Obstet. Gynecol. 2019, 134, 1187–1196. [Google Scholar] [CrossRef]
- Cools, P. The Role of Escherichia coli in Reproductive Health: State of the Art. Res. Microbiol. 2017, 168, 892–901. [Google Scholar] [CrossRef]
- Sperling, R.S.; Newton, E.; Gibbs, R.S. Intraamniotic Infection in Low-Birth-Weight Infants. J. Infect. Dis. 1988, 157, 113–117. [Google Scholar] [CrossRef]
- Gibbs, R.S.; Blanco, J.D.; St. Clair, P.J.; Castaneda, Y.S. Quantitative Bacteriology of Amniotic Fluid from Women with Clinical Intraamniotic Infection at Term. J. Infect. Dis. 1982, 145, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Thakur, M.; Lata, S.; Pal, A.; Sharma, H.; Dhiman, B. Relationship between Histologic Chorioamnionitis and Genital Tract Cultures in Pre Term Labour. J. Obstet. Gynaecol. 2021, 41, 721–725. [Google Scholar] [CrossRef] [PubMed]
- Romero, R.; Gomez-Lopez, N.; Winters, A.D.; Jung, E.; Shaman, M.; Bieda, J.; Panaitescu, B.; Pacora, P.; Erez, O.; Greenberg, J.M.; et al. Evidence That Intra-Amniotic Infections Are Often the Result of an Ascending Invasion—A Molecular Microbiological Study. J. Perinat. Med. 2019, 47, 915–931. [Google Scholar] [CrossRef]
- Frew, L.; Stock, S.J. Antimicrobial Peptides and Pregnancy. Reproduction 2011, 141, 725–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kai-Larsen, Y.; Gudmundsson, G.H.; Agerberth, B. A Review of the Innate Immune Defence of the Human Foetus and Newborn, with the Emphasis on Antimicrobial Peptides. Acta Paediatr. 2014, 103, 1000–1008. [Google Scholar] [CrossRef]
- Ramuta, T.Ž.; Šket, T.; Erjavec, M.S.; Kreft, M.E. Antimicrobial Activity of Human Fetal Membranes: From Biological Function to Clinical Use. Front. Bioeng. Biotechnol. 2021, 9, 691522. [Google Scholar] [CrossRef]
- Olmos-Ortiz, A.; Flores-Espinosa, P.; Mancilla-Herrera, I.; Vega-Sánchez, R.; Díaz, L.; Zaga-Clavellina, V. Innate Immune Cells and Toll-like Receptor–Dependent Responses at the Maternal–Fetal Interface. Int. J. Mol. Sci. 2019, 20, 3654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yarbrough, V.L.; Winkle, S.; Herbst-Kralovetz, M.M. Antimicrobial Peptides in the Female Reproductive Tract: A Critical Component of the Mucosal Immune Barrier with Physiological and Clinical Implications. Hum. Reprod. Update 2015, 21, 353–377. [Google Scholar] [CrossRef] [Green Version]
- Son, G.H.; Lee, J.J.; Kim, Y.; Lee, K.Y. The Role of Antimicrobial Peptides in Preterm Birth. Int. J. Mol. Sci. 2021, 22, 8905. [Google Scholar] [CrossRef]
- Kjaergaard, N.; Hein, M.; Hyttel, L.; Helmig, R.B.; Schonheyder, H.C.; Uldbjerg, N.; Madsen, H. Antibacterial Properties of Human Amnion and Chorion in Vitro. Eur. J. Obstet. Gynecol. Reprod. Biol. 2001, 94, 224–229. [Google Scholar] [CrossRef] [Green Version]
- King, A.E.; Paltoo, A.; Kelly, R.W.; Sallenave, J.M.; Bocking, A.D.; Challis, J.R.G. Expression of Natural Antimicrobials by Human Placenta and Fetal Membranes. Placenta 2007, 28, 161–169. [Google Scholar] [CrossRef]
- Stock, S.J.; Kelly, R.W.; Riley, S.C.; Calder, A.A. Natural Antimicrobial Production by the Amnion. Am. J. Obstet. Gynecol. 2007, 196, 255.e1–255.e6. [Google Scholar] [CrossRef]
- Hein, M.; Valore, E.V.; Helmig, R.B.; Uldbjerg, N.; Ganz, T. Antimicrobial Factors in the Cervical Mucus Plug. Am. J. Obstet. Gynecol. 2002, 187, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Svinarich, D.M.; Gomez, R.; Romero, R. Detection of Human Defensins in the Placenta. Am. J. Reprod. Immunol. 1997, 38, 252–255. [Google Scholar] [CrossRef] [PubMed]
- Erez, O.; Romero, R.; Tarca, A.L.; Chaiworapongsa, T.; Kim, Y.M.; Than, N.G.; Vaisbuch, E.; Raghici, S.; Tromp, G. Differential Expression Pattern of Genes Encoding for Anti-Microbial Peptides in the Fetal Membranes of Patients with Spontaneous Preterm Labor and Intact Membranes and Those with Preterm Prelabor Rupture of the Membranes. J. Matern.-Fetal Neonatal Med. 2009, 22, 1103–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshio, H.; Tollin, M.; Gudmundsson, G.H.; Lagercrantz, H.; Jörnvall, H.; Marchini, G.; Agerberth, B. Antimicrobial Polypeptides of Human Vernix Caseosa and Amniotic Fluid: Implications for Newborn Innate Defense. Pediatr. Res. 2003, 53, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Ericksen, B.; Wu, Z.; Lu, W.; Lehrer, R.I. Antibacterial Activity and Specificity of Six Human α-Defensins. Antimicrob. Agents Chemother. 2005, 49, 269–275. [Google Scholar] [CrossRef] [Green Version]
- Stock, S.J.; Duthie, L.; Tremaine, T.; Calder, A.A.; Kelly, R.W.; Riley, S.C. Elafin (SKALP/Trappin-2/Proteinase Inhibitor-3) Is Produced by the Cervix in Pregnancy and Cervicovaginal Levels Are Diminished in Bacterial Vaginosis. Reprod. Sci. 2009, 16, 1125–1134. [Google Scholar] [CrossRef]
- Rademacher, F.; Dreyer, S.; Kopfnagel, V.; Gläser, R.; Werfel, T.; Harder, J. The Antimicrobial and Immunomodulatory Function of RNase 7 in Skin. Front. Immunol. 2019, 10, 2553. [Google Scholar] [CrossRef] [Green Version]
- Rudolph, B.; Podschun, R.; Sahly, H.; Schubert, S.; Schröder, J.M.; Harder, J. Identification of RNase 8 as a Novel Human Antimicrobial Protein. Antimicrob. Agents Chemother. 2006, 50, 3194–3196. [Google Scholar] [CrossRef] [Green Version]
- Touqui, L.; Alaoui-El-Azher, M. Mammalian Secreted Phospholipases A2 and Their Pathophysiolo-Gical Significance in Inflammatory Diseases. Curr. Mol. Med. 2005, 1, 739–754. [Google Scholar] [CrossRef]
- D’Amico, F.; Skarmoutsou, E.; Granata, M.; Trovato, C.; Rossi, G.A.; Mazzarino, M.C. S100A7: A RAMPing up AMP Molecule in Psoriasis. Cytokine Growth Factor Rev. 2016, 32, 97–104. [Google Scholar] [CrossRef]
- Bandurska, K.; Berdowska, A.; Barczyńska-Felusiak, R.; Krupa, P. Unique Features of Human Cathelicidin LL-37. BioFactors 2015, 41, 289–300. [Google Scholar] [CrossRef] [PubMed]
- King, A.E.; Critchley, H.O.D.; Kelly, R.W. Innate Immune Defences in the Human Endometrium. Reprod. Biol. Endocrinol. 2003, 1, 116. [Google Scholar] [CrossRef] [Green Version]
- Olmos-Ortiz, A.; García-Quiroz, J.; López-Marure, R.; González-Curiel, I.; Rivas-Santiago, B.; Olivares, A.; Avila, E.; Barrera, D.; Halhali, A.; Caldiño, F.; et al. Evidence of Sexual Dimorphism in Placental Vitamin D Metabolism: Testosterone Inhibits Calcitriol-Dependent Cathelicidin Expression. J. Steroid Biochem. Mol. Biol. 2016, 163, 173–182. [Google Scholar] [CrossRef]
- Garcia-Lopez, G.; Flores-Espinosa, P.; Zaga-Clavellina, V. Tissue-Specific Human Beta-Defensins (HBD)1, HBD2, and HBD3 Secretion from Human Extra-Placental Membranes Stimulated with Escherichia coli. Reprod. Biol. Endocrinol. 2010, 8, 146. [Google Scholar] [CrossRef] [Green Version]
- Šket, T.; Ramuta, T.Ž.; Erjavec, M.S.; Kreft, M.E. The Role of Innate Immune System in the Human Amniotic Membrane and Human Amniotic Fluid in Protection Against Intra-Amniotic Infections and Inflammation. Front. Immunol. 2021, 12, 4241. [Google Scholar] [CrossRef]
- Galask, R.P.; Varner, M.W.; Petzold, C.R.; Wilbur, S.L. Bacterial Attachment to the Chorioamniotic Membranes. Am. J. Obstet. Gynecol. 1984, 148, 915–928. [Google Scholar] [CrossRef]
- Gyr, T.N.; Malek, A.; Mathez-Loic, F.; Altermatt, H.J.; Bodmer, T.; Nicolaides, K.; Schneider, H. Permeation of Human Chorioamniotic Membranes by Escherichia coli in Vitro. Am. J. Obstet. Gynecol. 1994, 170, 223–227. [Google Scholar] [CrossRef]
- Klaffenbach, D.; Friedrich, D.; Strick, R.; Strissel, P.L.; Beckmann, M.W.; Rascher, W.; Gessner, A.; Dötsch, J.; Meißner, U.; Schnare, M. Contribution of Different Placental Cells to the Expression and Stimulation of Antimicrobial Proteins (AMPs). Placenta 2011, 32, 830–837. [Google Scholar] [CrossRef]
- Tehrani, F.A.; Modaresifar, K.; Azizian, S.; Niknejad, H. Induction of Antimicrobial Peptides Secretion by IL-1β Enhances Human Amniotic Membrane for Regenerative Medicine. Sci. Rep. 2017, 7, 17022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores-Espinosa, P.; Preciado-Martínez, E.; Mejía-Salvador, A.; Sedano-González, G.; Bermejo-Martínez, L.; Parra-Covarruvias, A.; Estrada-Gutiérrez, G.; Vega-Sánchez, R.; Méndez, I.; Quesada-Reyna, B.; et al. Selective Immuno-Modulatory Effect of Prolactin upon pro-Inflammatory Response in Human Fetal Membranes. J. Reprod. Immunol. 2017, 123, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Grigsby, P.L.; Novy, M.J.; Waldorf, K.M.A.; Sadowsky, D.W.; Gravett, M.G. Choriodecidual Inflammation: A Harbinger of the Preterm Labor Syndrome. Reprod. Sci. 2010, 17, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Adams-Waldorf, K.M.; Persing, D.; Novy, M.J.; Sadowsky, D.W.; Gravett, M.G. Pretreatment with Toll-like Receptor 4 Antagonist Inhibits Lipopolysaccharide-Induced Preterm Uterine Contractility, Cytokines, and Prostaglandins in Rhesus Monkeys. Reprod. Sci. 2008, 15, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Flores-Espinosa, P.; Pineda-Torres, M.; Vega-Sánchez, R.; Estrada-Gutiérrez, G.; Espejel-Nuñez, A.; Flores-Pliego, A.; Maida-Claros, R.; Paredes-Vivas, Y.; Morales-Méndez, I.; Sosa-González, I.; et al. Progesterone Elicits an Inhibitory Effect upon LPS-Induced Innate Immune Response in Pre-Labor Human Amniotic Epithelium. Am. J. Reprod. Immunol. 2014, 71, 61–72. [Google Scholar] [CrossRef]
- Thiex, N.W.; Chames, M.C.; Loch-Caruso, R.K. Tissue-Specific Cytokine Release from Human Extra-Placental Membranes Stimulated by Lipopolysaccharide in a Two-Compartment Tissue Culture System. Reprod. Biol. Endocrinol. 2009, 7, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaga, V.; Estrada-Gutierrez, G.; Beltran-Montoya, J.; Maida-Claros, R.; Lopez-Vancell, R.; Vadillo-Ortega, F. Secretions of Interleukin-1β and Tumor Necrosis Factor α by Whole Fetal Membranes Depend on Initial Interactions of Amnion or Choriodecidua with Lipopolysaccharides or Group B Streptococci. Biol. Reprod. 2004, 71, 1296–1302. [Google Scholar] [CrossRef]
- Iavazzo, C.; Tassis, K.; Gourgiotis, D.; Boutsikou, M.; Baka, S.; Hassiakos, D.; Hadjithomas, A.; Botsis, D.; Malamitsi-Puchner, A. The Role of Human Beta Defensins 2 and 3 in the Second Trimester Amniotic Fluid in Predicting Preterm Labor and Premature Rupture of Membranes. Arch. Gynecol. Obstet. 2010, 281, 793–799. [Google Scholar] [CrossRef]
- Varrey, A.; Romero, R.; Panaitescu, B.; Miller, D.; Chaiworapongsa, T.; Patwardhan, M.; Faro, J.; Pacora, P.; Hassan, S.S.; Hsu, C.D.; et al. Human β-Defensin-1: A Natural Antimicrobial Peptide Present in Amniotic Fluid That Is Increased in Spontaneous Preterm Labor with Intra-Amniotic Infection. Am. J. Reprod. Immunol. 2018, 80, e13031. [Google Scholar] [CrossRef]
- Espinoza, J.; Chaiworapongsa, T.; Romero, R.; Edwin, S.; Rathnasabapathy, C.; Gomez, R.; Bujold, E.; Camacho, N.; Kim, Y.M.; Hassan, S.; et al. Antimicrobial Peptides in Amniotic Fluid: Defensins, Calprotectin and Bacterial/Permeability-Increasing Protein in Patients with Microbial Invasion of the Amniotic Cavity, Intra-Amniotic Inflammation, Preterm Labor and Premature Rupture of Membranes. J. Matern.-Fetal Neonatal Med. 2003, 13, 2–21. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Y.; Du, C.; Zhang, R.; Feng, Z.; Zhang, J. Diagnostic Value of Amniotic Fluid Inflammatory Biomarkers for Subclinical Chorioamnionitis. Int. J. Gynecol. Obstet. 2016, 134, 160–164. [Google Scholar] [CrossRef]
- Buhimschi, I.A.; Jabr, M.; Buhimschi, C.S.; Petkova, A.P.; Weiner, C.P.; Saed, G.M. The Novel Antimicrobial Peptide Β3-Defensin Is Produced by the Amnion: A Possible Role of the Fetal Membranes in Innate Immunity of the Amniotic Cavity. Am. J. Obstet. Gynecol. 2004, 191, 1678–1687. [Google Scholar] [CrossRef] [PubMed]
- Szukiewicz, D.; Szewczyk, G.; Pyzlak, M.; Klimkiewicz, J.; Maslinska, D. Increased Production of β-Defensin 3 (HBD-3) by Human Amniotic Epithelial Cells (HAEC) after Activation of Toll-like Receptor 4 in Chorioamnionitis. Inflamm. Res. 2008, 57, 67. [Google Scholar] [CrossRef] [PubMed]
- Zaga-Clavellina, V.; Garcia-Lopez, G.; Flores-Espinosa, P. Evidence of in Vitro Differential Secretion of Human Beta-Defensins-1, -2, and -3 after Selective Exposure to Streptococcus agalactiae in Human Fetal Membranes. J. Matern.-Fetal Neonatal Med. 2012, 25, 358–363. [Google Scholar] [CrossRef]
- Zaga-Clavellina, V.; Martha, R.V.M.; Flores-Espinosa, P. In Vitro Secretion Profile of Pro-Inflammatory Cytokines IL-1β, TNF-α, IL-6, and of Human Beta-Defensins (HBD)-1, HBD-2, and HBD-3 from Human Chorioamniotic Membranes after Selective Stimulation with Gardnerella vaginalis. Am. J. Reprod. Immunol. 2012, 67, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Olmos-Ortiz, A.; Noyola-Martínez, N.; Barrera, D.; Zaga-Clavellina, V.; Avila, E.; Halhali, A.; Biruete, B.; Larrea, F.; Díaz, L. IL-10 Inhibits While Calcitriol Reestablishes Placental Antimicrobial Peptides Gene Expression. J. Steroid Biochem. Mol. Biol. 2015, 148, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Olmos-Ortiz, A.; García-Quiroz, J.; Avila, E.; Caldiño-Soto, F.; Halhali, A.; Larrea, F.; Díaz, L. Lipopolysaccharide and cAMP Modify Placental Calcitriol Biosynthesis Reducing Antimicrobial Peptides Gene Expression. Am. J. Reprod. Immunol. 2018, 79, e12841. [Google Scholar] [CrossRef]
- Chen, X.; Niyonsaba, F.; Ushio, H.; Okuda, D.; Nagaoka, I.; Ikeda, S.; Okumura, K.; Ogawa, H. Synergistic Effect of Antibacterial Agents Human β-Defensins, Cathelicidin LL-37 and Lysozyme against Staphylococcus aureus and Escherichia coli. J. Dermatol. Sci. 2005, 40, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Vanzolini, T.; Bruschi, M.; Rinaldi, A.C.; Magnani, M.; Fraternale, A. Multitalented Synthetic Antimicrobial Peptides and Their Antibacterial, Antifungal and Antiviral Mechanisms. Int. J. Mol. Sci. 2022, 23, 545. [Google Scholar] [CrossRef]
- Luo, Y.; Song, Y. Mechanism of Antimicrobial Peptides: Antimicrobial, Anti-Inflammatory and Antibiofilm Activities. Int. J. Mol. Sci. 2021, 22, 11401. [Google Scholar] [CrossRef]
- Oppenheim, J.J.; Biragyn, A.; Kwak, L.W.; Yang, D. Roles of Antimicrobial Peptides Such as Defensins in Innate and Adaptive Immunity. Ann. Rheum. Dis. 2003, 62 (Suppl. S2), ii17–ii21. [Google Scholar] [CrossRef] [Green Version]
- Wang, G. Antimicrobial Peptides: Discovery, Design, and Novel Therapeutic Strategies; CABI: Wallingford, UK, 2010; ISBN 9781845936570. [Google Scholar]
- Duong, L.; Gross, S.P.; Siryaporn, A. Developing Antimicrobial Synergy With AMPs. Front. Med. Technol. 2021, 3, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Hao, W.; Wang, X.; Ouyang, J.; Deng, X.; Yu, H.; Wang, Y. Antimicrobial Peptides, Conventional Antibiotics, and Their Synergistic Utility for the Treatment of Drug-Resistant Infections. Med. Res. Rev. 2022. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.; Nagaraj, R. Antimicrobial Activity of Human β-Defensin 4 Analogs: Insights into the Role of Disulfide Linkages in Modulating Activity. Peptides 2012, 38, 255–265. [Google Scholar] [CrossRef]
- Krishnakumari, V.; Packiyanathan, K.K.; Nagaraj, R. Human-β-Defensins-1-3 and Analogs Do Not Require Proton Motive Force for Antibacterial Activity against Escherichia coli. FEMS Microbiol. Lett. 2013, 348, 52–57. [Google Scholar] [CrossRef] [Green Version]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and Simple Determination of the Escherichia coli Phylogenetic Group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R.; Stell, A.L. Extended Virulence Genotypes of Escherichia coli Strains from Patients with Urosepsis in Relation to Phylogeny and Host Compromise. J. Infect. Dis. 2000, 181, 261–272. [Google Scholar] [CrossRef] [Green Version]
- Scottish Intercollegiate Guidelines Network (SIGN). Management of Suspected Bacterial Urinary Tract Infection in Adults; Scottish Intercollegiate Guidelines Network (SIGN): Edinburgh, UK, 2012. [Google Scholar]
- Kass, E.H. Bacteriuria and the Diagnosis of Infections of the Urinary Tract: With Observations on the Use of Methionine as a Urinary Antiseptic. AMA Arch. Intern. Med. 1957, 100, 709–714. [Google Scholar] [CrossRef] [PubMed]
- Becerra, S.C.; Roy, D.C.; Sanchez, C.J.; Christy, R.J.; Burmeister, D.M. An Optimized Staining Technique for the Detection of Gram Positive and Gram Negative Bacteria within Tissue. BMC Res. Notes 2016, 9, 216. [Google Scholar] [CrossRef] [Green Version]
- Segnani, C.; Ippolito, C.; Antonioli, L.; Pellegrini, C.; Blandizzi, C.; Dolfi, A.; Bernardini, N. Histochemical Detection of Collagen Fibers by Sirius Red/Fast Green Is More Sensitive than van Gieson or Sirius Red Alone in Normal and Inflamed Rat Colon. PLoS ONE 2015, 10, e0144630. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Parameter | Mean ± SD | Range |
|---|---|---|
| (n = 13) | (Min–Max) | |
| Maternal age (years) | 30.9 ± 5.8 | (22–39) |
| Gestational age (weeks) | 38.6 ± 0.6 | (38–40) |
| Number of pregnancies | 2.5 ± 0.7 | (2–4) |
| Newborn cephalic perimeter (cm) | 34.8 ± 0.9 | (33–36) |
| Newborn length (cm) | 48.6 ± 2.5 | (46–52) |
| Newborn weight (g) | 3184.1 ± 453.7 | (2760–4040) |
| Apgar 1 min | 8.6 ± 0.4 | (8–9) |
| Apgar 5 min | 8.9 ± 0.2 | (8–9) |
| Newborn sex (male/female) (%/%) | 5/8 | 38%/62% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olmos-Ortiz, A.; Hernández-Pérez, M.; Flores-Espinosa, P.; Sedano, G.; Helguera-Repetto, A.C.; Villavicencio-Carrisoza, Ó.; Valdespino-Vazquez, M.Y.; Flores-Pliego, A.; Irles, C.; Rivas-Santiago, B.; et al. Compartmentalized Innate Immune Response of Human Fetal Membranes against Escherichia coli Choriodecidual Infection. Int. J. Mol. Sci. 2022, 23, 2994. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23062994
Olmos-Ortiz A, Hernández-Pérez M, Flores-Espinosa P, Sedano G, Helguera-Repetto AC, Villavicencio-Carrisoza Ó, Valdespino-Vazquez MY, Flores-Pliego A, Irles C, Rivas-Santiago B, et al. Compartmentalized Innate Immune Response of Human Fetal Membranes against Escherichia coli Choriodecidual Infection. International Journal of Molecular Sciences. 2022; 23(6):2994. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23062994
Chicago/Turabian StyleOlmos-Ortiz, Andrea, Mayra Hernández-Pérez, Pilar Flores-Espinosa, Gabriela Sedano, Addy Cecilia Helguera-Repetto, Óscar Villavicencio-Carrisoza, María Yolotzin Valdespino-Vazquez, Arturo Flores-Pliego, Claudine Irles, Bruno Rivas-Santiago, and et al. 2022. "Compartmentalized Innate Immune Response of Human Fetal Membranes against Escherichia coli Choriodecidual Infection" International Journal of Molecular Sciences 23, no. 6: 2994. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23062994