
Characterization of a Novel Aspect of Tissue Scarring Following Experimental Spinal Cord Injury and the Implantation of Bioengineered Type-I Collagen Scaffolds in the Adult Rat: Involvement of Perineurial-like Cells?

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
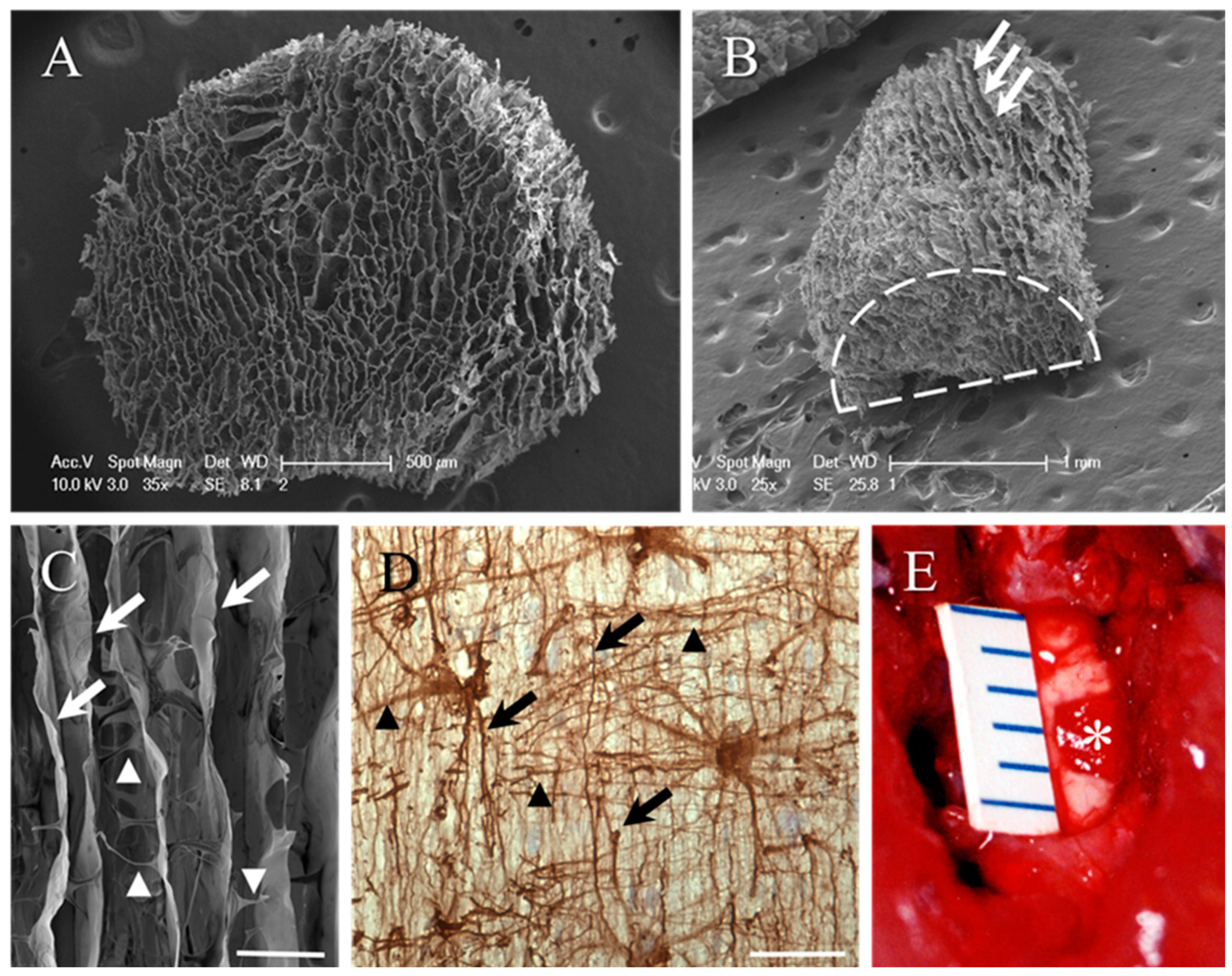
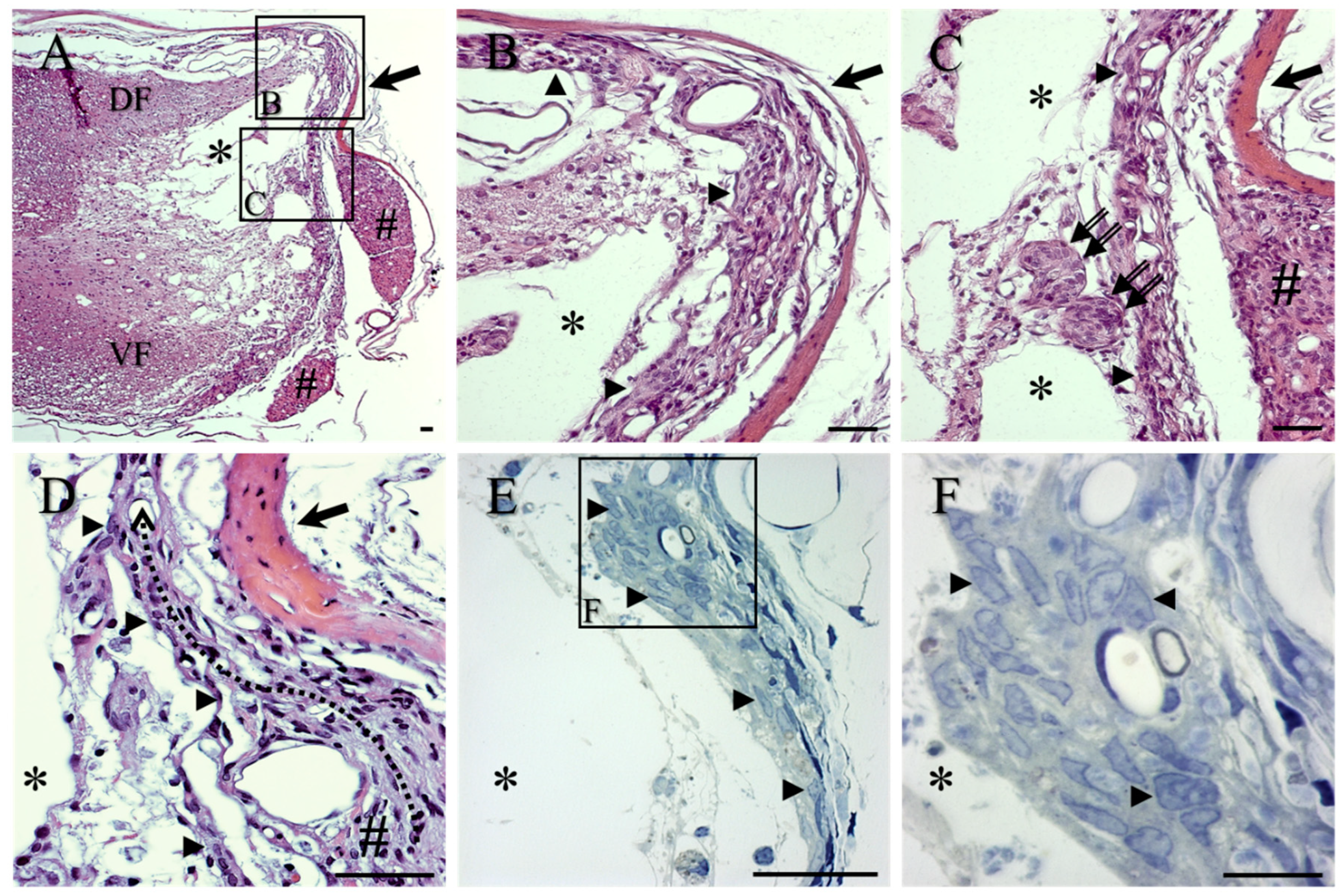
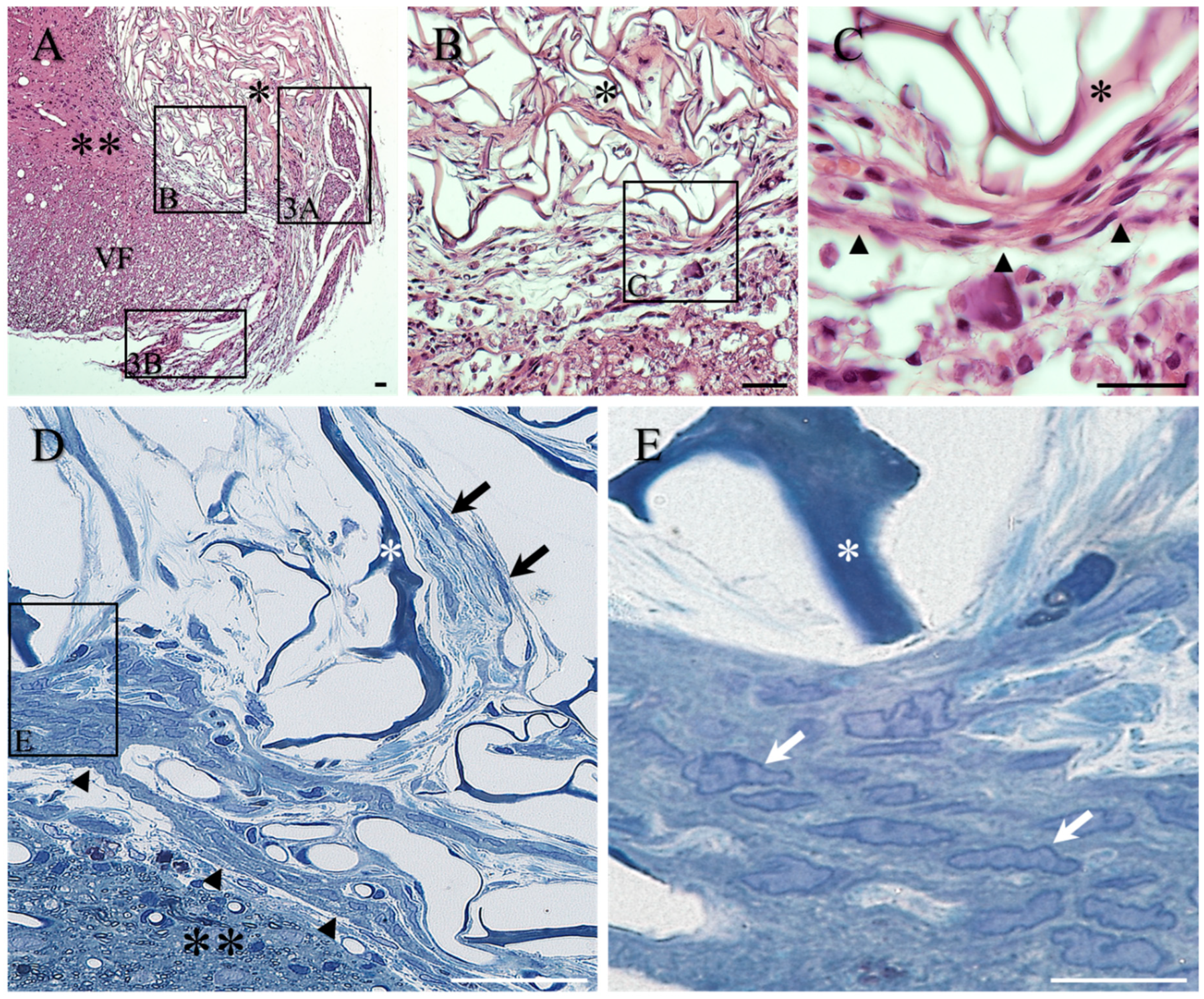
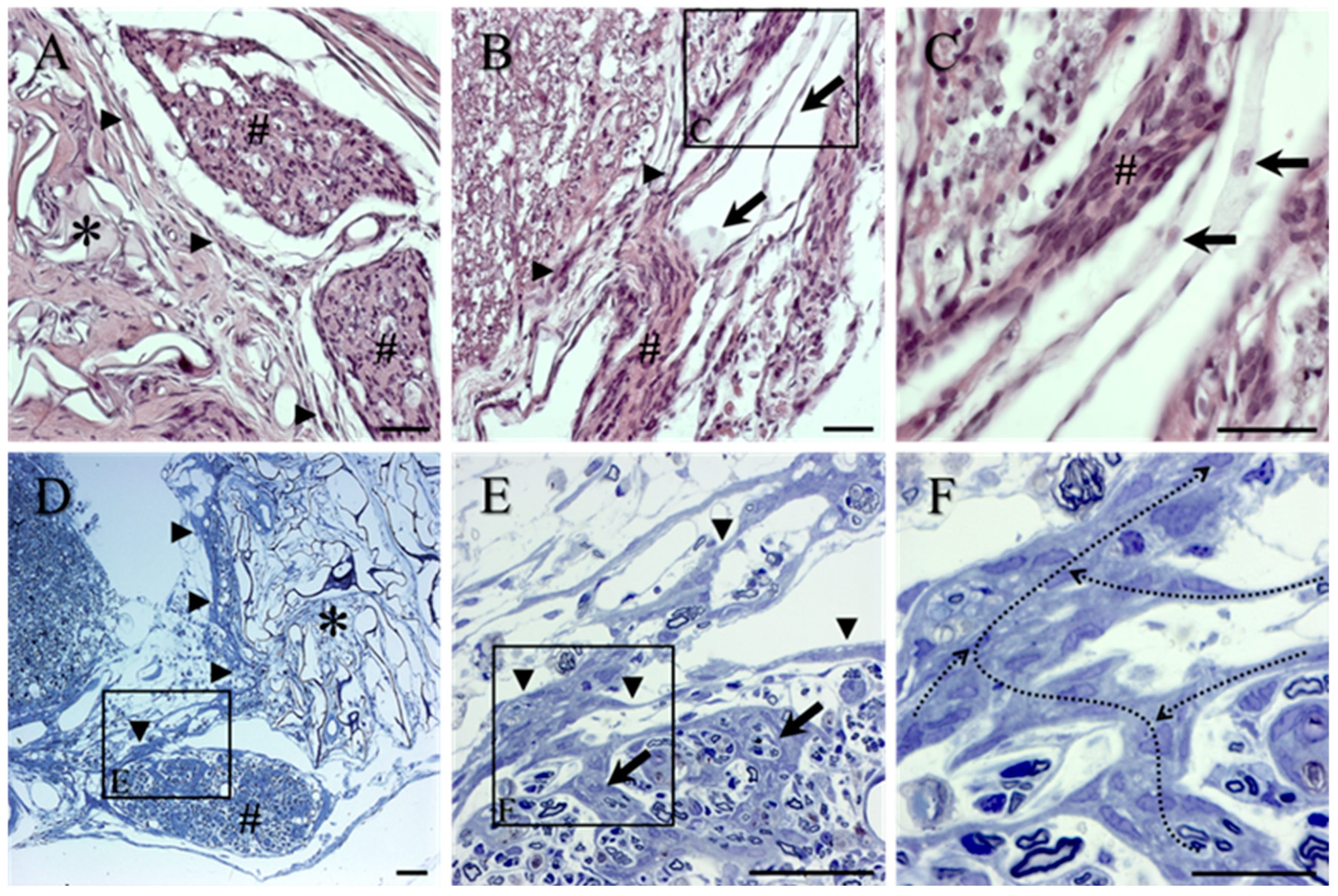
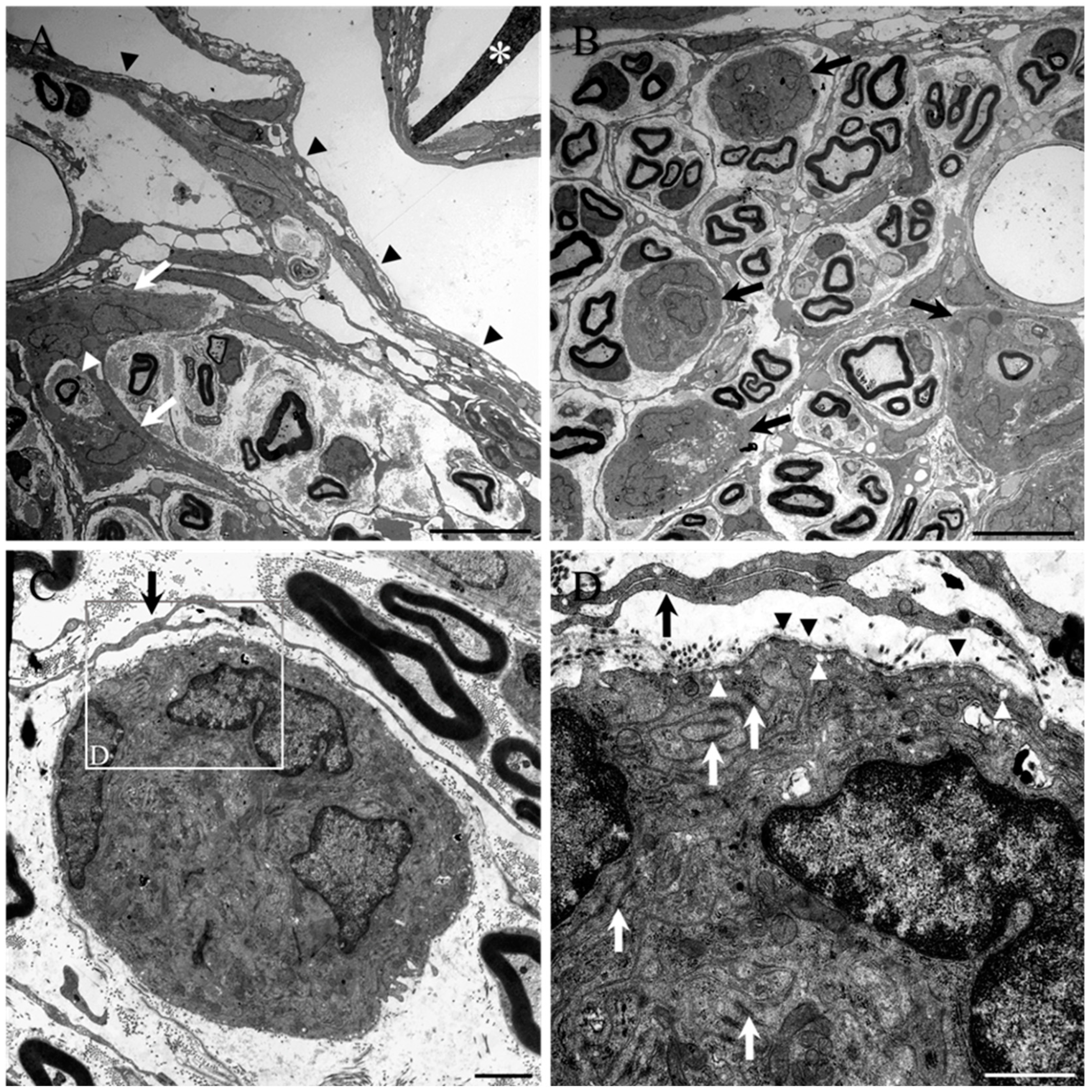
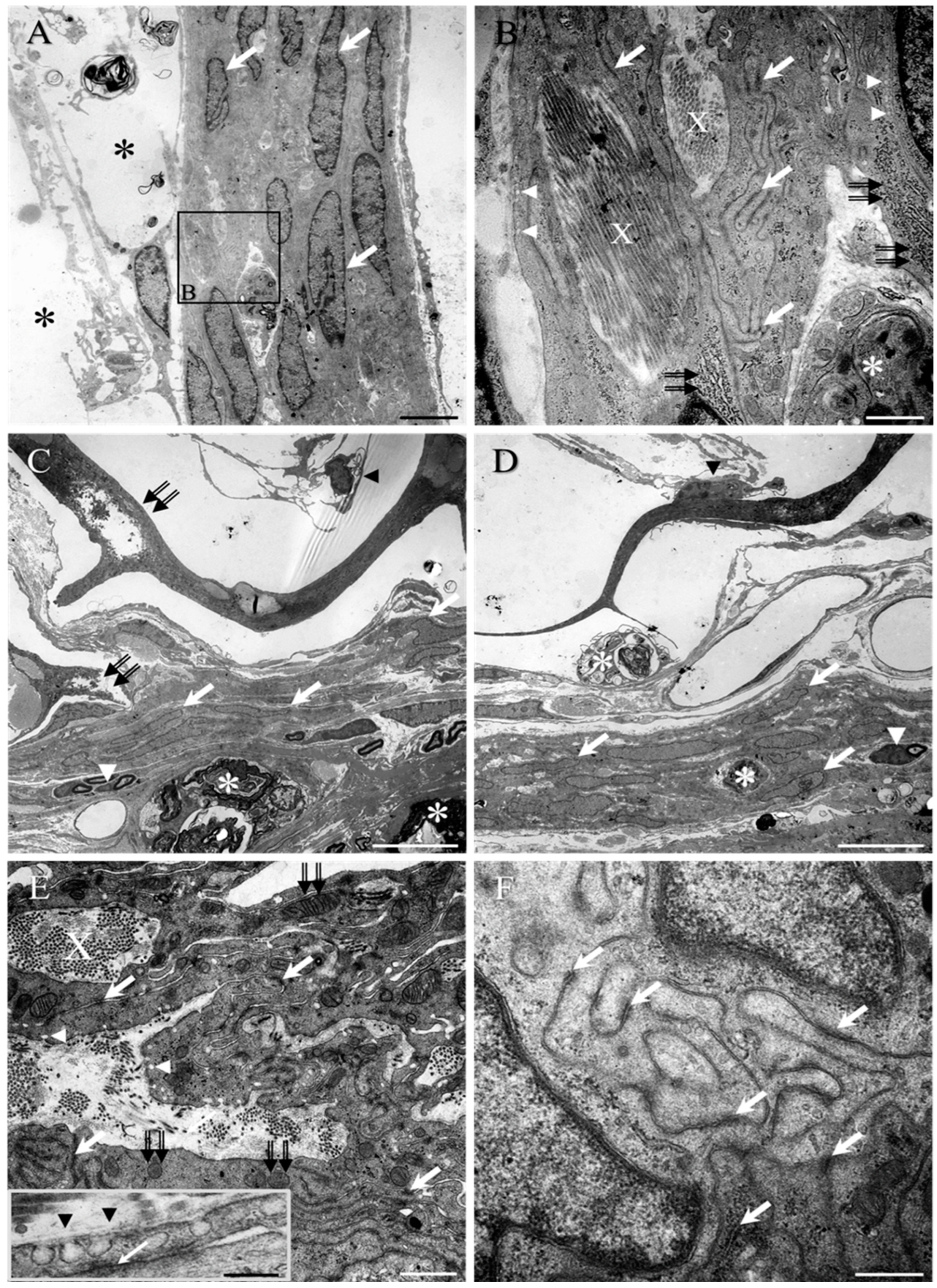
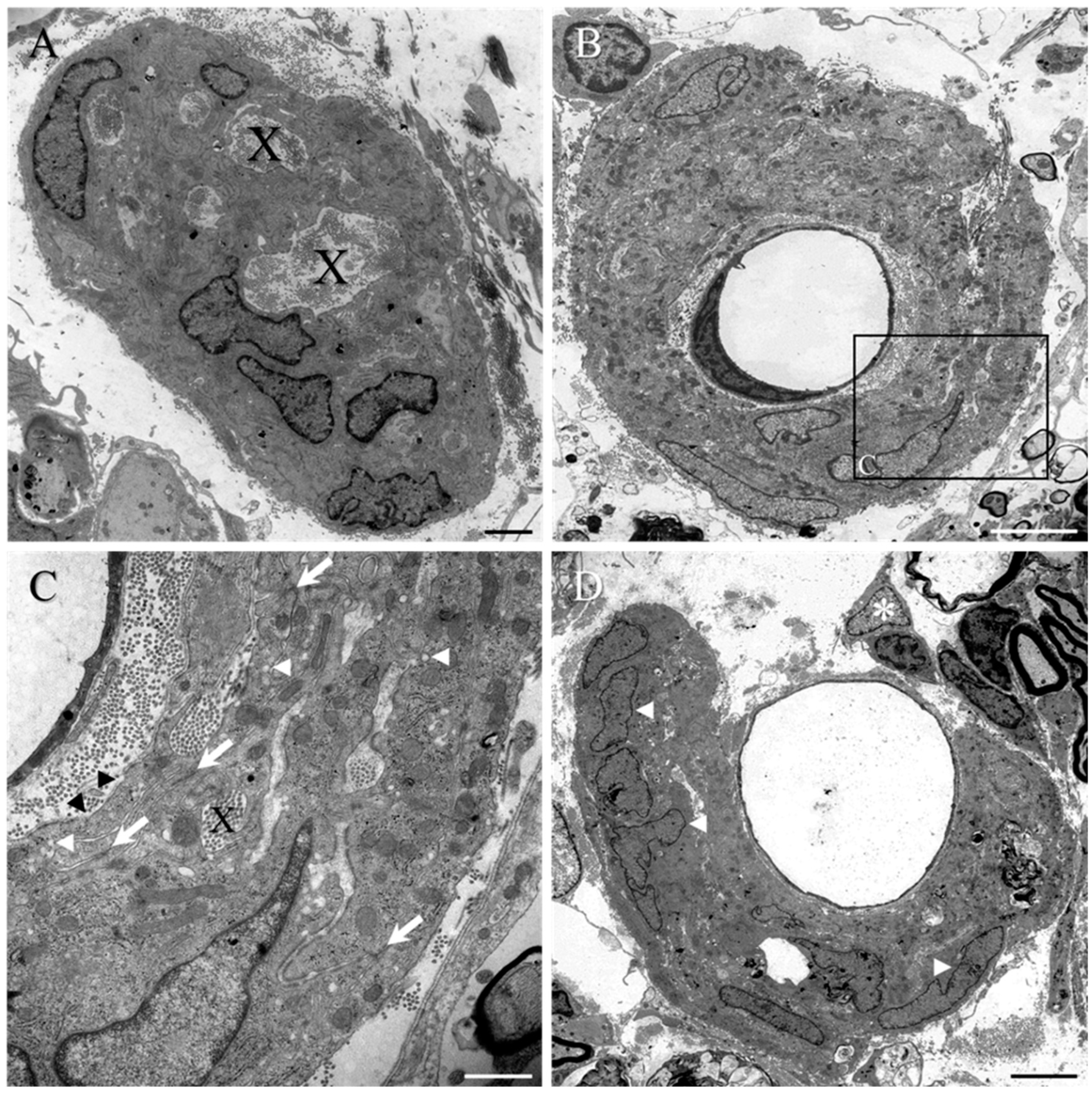
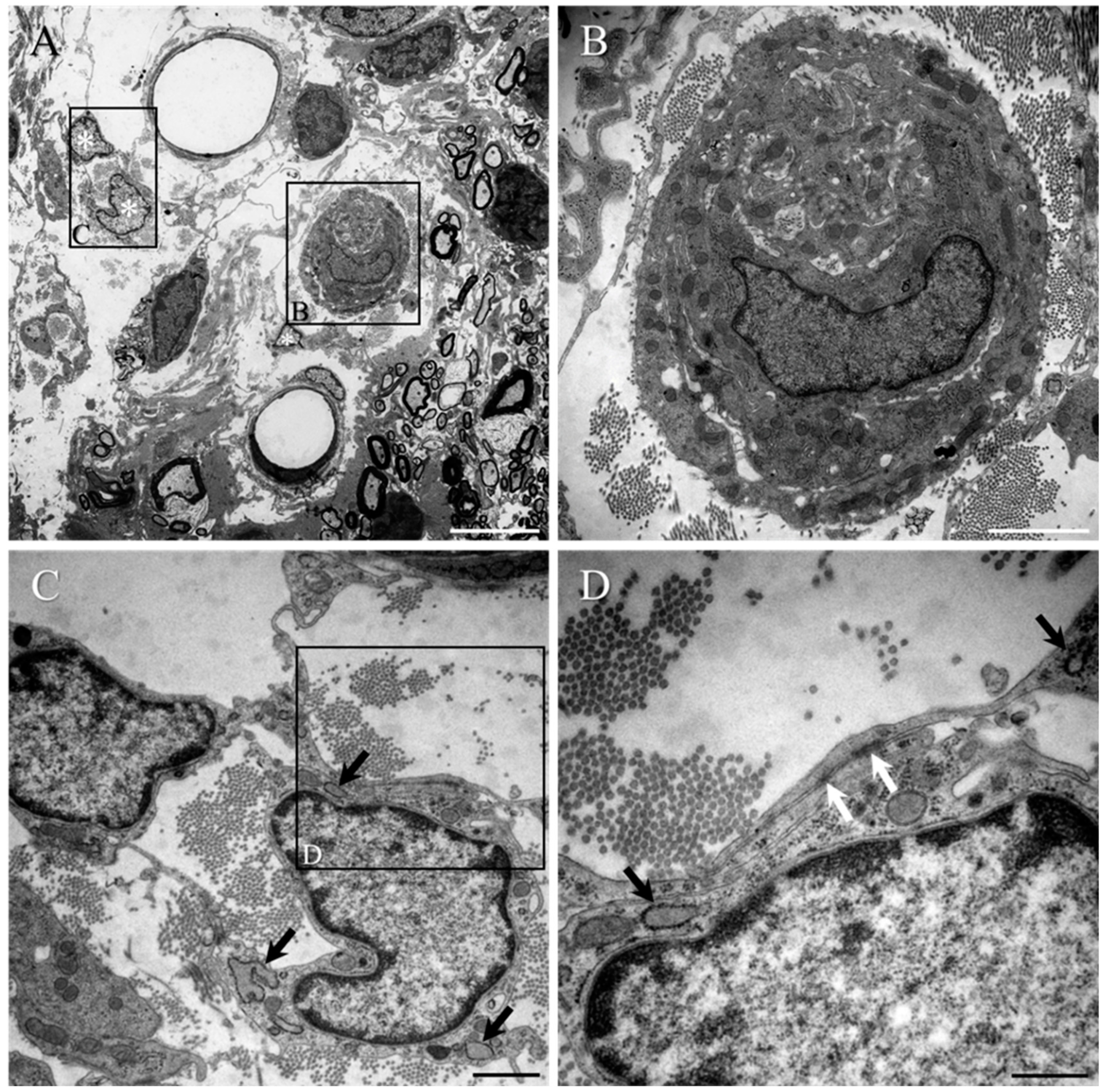
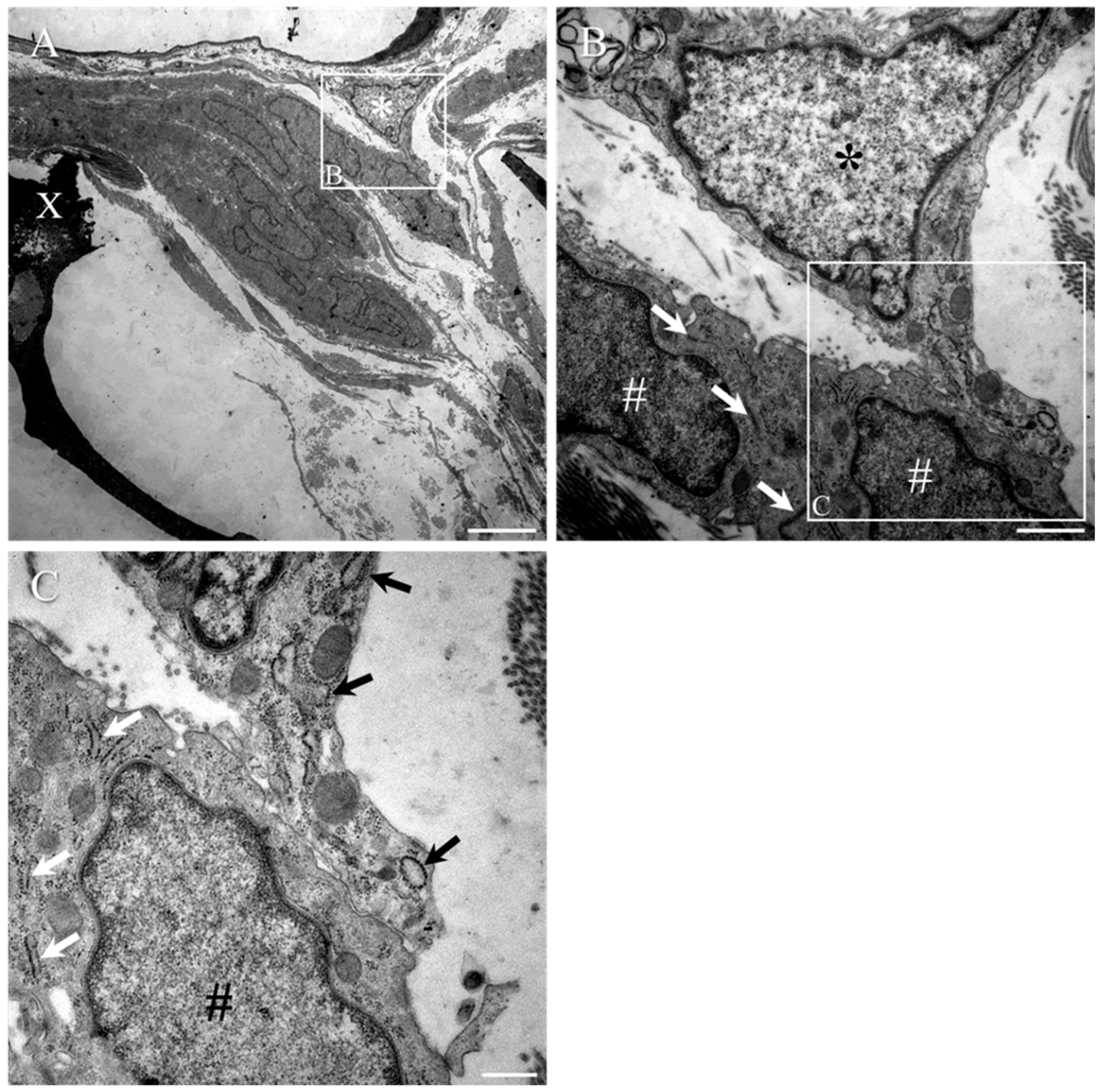
2. Results
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Surgical Procedure
4.3. Tissue Processing and Staining, Light Microscopy, and Transmission Electron Microscopy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bunge, R.P.; Puckett, W.R.; Becerra, J.L.; Marcillo, A.; Quencer, R.M. Observations on the pathology of human spinal cord injury. A review and classification of 22 new cases with details from a case of chronic cord compression with extensive focal demyelination. Adv. Neurol. 1993, 59, 75–89. [Google Scholar] [PubMed]
- Kakulas, B.; Taylor, J. Pathology of injuries of the vertebral column and spinal cord. Handb. Clin. Neurol. 1992, 17, 21–51. [Google Scholar]
- Silver, J.; Miller, J.H. Regeneration beyond the glial scar. Nat. Rev. Neurosci. 2004, 5, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Fawcett, J.W.; Asher, R.A. The glial scar and central nervous system repair. Brain Res. Bull. 1999, 49, 377–391. [Google Scholar] [CrossRef]
- Stichel, C.C.; Muller, H.W. The CNS lesion scar: New vistas on an old regeneration barrier. Cell Tissue Res. 1998, 294, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pettigrew, D.B.; Shockley, K.P.; Crutcher, K.A. Disruption of spinal cord white matter and sciatic nerve geometry inhibits axonal growth in vitro in the absence of glial scarring. BMC Neurosci. 2001, 2, 8. [Google Scholar]
- Filli, L.; Schwab, M.E. The rocky road to translation in spinal cord repair. Ann. Neurol. 2012, 72, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tan, H.; Hui, X. Biomaterial Scaffolds in Regenerative Therapy of the Central Nervous System. BioMed Res. Int. 2018, 2018, 7848901. [Google Scholar] [CrossRef] [Green Version]
- Fuhrmann, T.; Anandakumaran, P.N.; Shoichet, M.S. Combinatorial Therapies After Spinal Cord Injury: How Can Biomaterials Help. Adv. Healthcare Mater. 2017, 6, 1601130. [Google Scholar] [CrossRef]
- Oudega, M.; Bradbury, E.J.; Ramer, M.S. Combination therapies. Handb. Clin. Neurol. 2012, 109, 617–636. [Google Scholar] [CrossRef]
- Zhang, Q.; Shi, B.; Ding, J.; Yan, L.; Thawani, J.P.; Fu, C.; Chen, X. Polymer scaffolds facilitate spinal cord injury repair. Acta Biomater. 2019, 88, 57–77. [Google Scholar] [CrossRef]
- Bozkurt, A.; Boecker, A.; Tank, J.; Altinova, H.; Deumens, R.; Dabhi, C.; Tolba, R.; Weis, J.; Brook, G.A.; Pallua, N.; et al. Efficient bridging of 20 mm rat sciatic nerve lesions with a longitudinally micro-structured collagen scaffold. Biomaterials 2016, 75, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, A.; Brook, G.A.; Moellers, S.; Lassner, F.; Sellhaus, B.; Weis, J.; Woeltje, M.; Tank, J.; Beckmann, C.; Fuchs, P. In vitro assessment of axonal growth using dorsal root ganglia explants in a novel three-dimensional collagen matrix. Tissue Eng. 2007, 13, 2971–2979. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, A.; Deumens, R.; Beckmann, C.; Olde Damink, L.; Schugner, F.; Heschel, I.; Sellhaus, B.; Weis, J.; Jahnen-Dechent, W.; Brook, G.A.; et al. In vitro cell alignment obtained with a Schwann cell enriched microstructured nerve guide with longitudinal guidance channels. Biomaterials 2009, 30, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Van Neerven, S.G.; Krings, L.; Haastert-Talini, K.; Vogt, M.; Tolba, R.H.; Brook, G.; Pallua, N.; Bozkurt, A. Human Schwann cells seeded on a novel collagen-based microstructured nerve guide survive, proliferate, and modify neurite outgrowth. BioMed Res. Int. 2014, 2014, 493823. [Google Scholar] [CrossRef] [PubMed]
- Deumens, R.; Bozkurt, A.; Meek, M.F.; Marcus, M.A.; Joosten, E.A.; Weis, J.; Brook, G.A. Repairing injured peripheral nerves: Bridging the gap. Prog. Neurobiol. 2010, 92, 245–276. [Google Scholar] [CrossRef]
- Mollers, S.; Heschel, I.; Damink, L.H.; Schugner, F.; Deumens, R.; Muller, B.; Bozkurt, A.; Nava, J.G.; Noth, J.; Brook, G.A. Cytocompatibility of a novel, longitudinally microstructured collagen scaffold intended for nerve tissue repair. Tissue Eng. A 2009, 15, 461–472. [Google Scholar] [CrossRef]
- Van Neerven, S.G.A.; Haastert-Talini, K.; Boecker, A.; Schriever, T.; Dabhi, C.; Claeys, K.; Deumens, R.; Brook, G.A.; Weis, J.; Pallua, N.; et al. Two-component collagen nerve guides support axonal regeneration in the rat peripheral nerve injury model. J. Tissue Eng. Regen. Med. 2017, 11, 3349–3361. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, A.; Claeys, K.G.; Schrading, S.; Rodler, J.V.; Altinova, H.; Schulz, J.B.; Weis, J.; Pallua, N.; van Neerven, S.G.A. Clinical and biometrical 12-month follow-up in patients after reconstruction of the sural nerve biopsy defect by the collagen-based nerve guide Neuromaix. Eur. J. Med. Res. 2017, 22, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozkurt, A.; van Neerven, S.G.; Claeys, K.G.; O’Dey, D.M.; Sudhoff, A.; Brook, G.A.; Sellhaus, B.; Schulz, J.B.; Weis, J.; Pallua, N. The proximal medial sural nerve biopsy model: A standardised and reproducible baseline clinical model for the translational evaluation of bioengineered nerve guides. BioMed Res. Int. 2014, 2014, 121452. [Google Scholar] [CrossRef] [PubMed]
- Cholas, R.; Hsu, H.P.; Spector, M. Collagen scaffolds incorporating select therapeutic agents to facilitate a reparative response in a standardized hemiresection defect in the rat spinal cord. Tissue Eng. A 2012, 18, 2158–2172. [Google Scholar] [CrossRef] [PubMed]
- Cholas, R.H.; Hsu, H.P.; Spector, M. The reparative response to cross-linked collagen-based scaffolds in a rat spinal cord gap model. Biomaterials 2012, 33, 2050–2059. [Google Scholar] [CrossRef] [PubMed]
- Guth, L.; Zhang, Z.; DiProspero, N.A.; Joubin, K.; Fitch, M.T. Spinal cord injury in the rat: Treatment with bacterial lipopolysaccharide and indomethacin enhances cellular repair and locomotor function. Exp. Neurol. 1994, 126, 76–87. [Google Scholar] [CrossRef]
- Hejcl, A.; Sedy, J.; Kapcalova, M.; Toro, D.A.; Amemori, T.; Lesny, P.; Likavcanova-Masinova, K.; Krumbholcova, E.; Pradny, M.; Michalek, J.; et al. HPMA-RGD hydrogels seeded with mesenchymal stem cells improve functional outcome in chronic spinal cord injury. Stem Cells Dev. 2010, 19, 1535–1546. [Google Scholar] [CrossRef]
- Pasterkamp, R.J.; Giger, R.J.; Ruitenberg, M.J.; Holtmaat, A.J.; De Wit, J.; De Winter, F.; Verhaagen, J. Expression of the gene encoding the chemorepellent semaphorin III is induced in the fibroblast component of neural scar tissue formed following injuries of adult but not neonatal CNS. Mol. Cell. Neurosci. 1999, 13, 143–166. [Google Scholar] [CrossRef]
- Brook, G.A.; Houweling, D.A.; Gieling, R.G.; Hermanns, T.; Joosten, E.A.; Bar, D.P.; Gispen, W.H.; Schmitt, A.B.; Leprince, P.; Noth, J.; et al. Attempted endogenous tissue repair following experimental spinal cord injury in the rat: Involvement of cell adhesion molecules L1 and NCAM. Eur. J. Neurosci. 2000, 12, 3224–3238. [Google Scholar] [CrossRef]
- Brook, G.A.; Plate, D.; Franzen, R.; Martin, D.; Moonen, G.; Schoenen, J.; Schmitt, A.B.; Noth, J.; Nacimiento, W. Spontaneous longitudinally orientated axonal regeneration is associated with the Schwann cell framework within the lesion site following spinal cord compression injury of the rat. J. Neurosci. Res. 1998, 53, 51–65. [Google Scholar] [CrossRef]
- Goritz, C.; Dias, D.O.; Tomilin, N.; Barbacid, M.; Shupliakov, O.; Frisen, J. A pericyte origin of spinal cord scar tissue. Science 2011, 333, 238–242. [Google Scholar] [CrossRef]
- Soderblom, C.; Luo, X.; Blumenthal, E.; Bray, E.; Lyapichev, K.; Ramos, J.; Krishnan, V.; Lai-Hsu, C.; Park, K.K.; Tsoulfas, P.; et al. Perivascular fibroblasts form the fibrotic scar after contusive spinal cord injury. J. Neurosci. 2013, 33, 13882–13887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altinova, H.; Mollers, S.; Fuhrmann, T.; Deumens, R.; Bozkurt, A.; Heschel, I.; Damink, L.H.; Schugner, F.; Weis, J.; Brook, G.A. Functional improvement following implantation of a microstructured, type-I collagen scaffold into experimental injuries of the adult rat spinal cord. Brain Res. 2014, 1585, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Altinova, H.; Möllers, S.; Deumens, R.; Gerardo-Nava, J.; Führmann, T.; van Neerven, S.G.A.; Bozkurt, A.; Mueller, C.A.; Hoff, H.J.; Heschel, I.; et al. Functional recovery not correlated with axon regeneration through olfactory ensheathing cell-seeded scaffolds in a model of acute spinal cord injury. Tissue Eng. Regen. Med. 2016, 13, 585–600. [Google Scholar] [CrossRef] [PubMed]
- Altinova, H.; Hammes, S.; Palm, M.; Achenbach, P.; Gerardo-Nava, J.; Deumens, R.; Fuhrmann, T.; van Neerven, S.G.A.; Hermans, E.; Weis, J.; et al. Dense fibroadhesive scarring and poor blood vessel-maturation hamper the integration of implanted collagen scaffolds in an experimental model of spinal cord injury. Biomed. Mater. 2020, 15, 015012. [Google Scholar] [CrossRef] [PubMed]
- Altinova, H.; Hammes, S.; Palm, M.; Gerardo-Nava, J.; Achenbach, P.; Deumens, R.; Hermans, E.; Fuhrmann, T.; Boecker, A.; van Neerven, S.G.A.; et al. Fibroadhesive scarring of grafted collagen scaffolds interferes with implant-host neural tissue integration and bridging in experimental spinal cord injury. Regen. Biomater. 2019, 6, 75–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradbury, E.J.; Burnside, E.R. Moving beyond the glial scar for spinal cord repair. Nat. Commun. 2019, 10, 3879. [Google Scholar] [CrossRef] [PubMed]
- Kawano, H.; Kimura-Kuroda, J.; Komuta, Y.; Yoshioka, N.; Li, H.P.; Kawamura, K.; Li, Y.; Raisman, G. Role of the lesion scar in the response to damage and repair of the central nervous system. Cell Tissue Res. 2012, 349, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Profyris, C.; Cheema, S.S.; Zang, D.; Azari, M.F.; Boyle, K.; Petratos, S. Degenerative and regenerative mechanisms governing spinal cord injury. Neurobiol. Dis. 2004, 15, 415–436. [Google Scholar] [CrossRef] [PubMed]
- Schwab, M.E.; Bartholdi, D. Degeneration and regeneration of axons in the lesioned spinal cord. Physiol. Rev. 1996, 76, 319–370. [Google Scholar] [CrossRef]
- Lu, P.; Tuszynski, M.H. Growth factors and combinatorial therapies for CNS regeneration. Exp. Neurol. 2008, 209, 313–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Xie, Y.Y.; Wang, B. Role and prospects of regenerative biomaterials in the repair of spinal cord injury. Neural Regen. Res. 2019, 14, 1352–1363. [Google Scholar] [CrossRef]
- Bozkurt, A.; Lassner, F.; O’Dey, D.; Deumens, R.; Bocker, A.; Schwendt, T.; Janzen, C.; Suschek, C.V.; Tolba, R.; Kobayashi, E.; et al. The role of microstructured and interconnected pore channels in a collagen-based nerve guide on axonal regeneration in peripheral nerves. Biomaterials 2012, 33, 1363–1375. [Google Scholar] [CrossRef]
- Van Neerven, S.G.; Bozkurt, A.; O’Dey, D.M.; Scheffel, J.; Boecker, A.H.; Stromps, J.P.; Dunda, S.; Brook, G.A.; Pallua, N. Retrograde tracing and toe spreading after experimental autologous nerve transplantation and crush injury of the sciatic nerve: A descriptive methodological study. J. Brachial Plex. Peripher. Nerve Inj. 2012, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Schoof, H.; Apel, J.; Heschel, I.; Rau, G. Control of pore structure and size in freeze-dried collagen sponges. J. Biomed. Mater. Res. 2001, 58, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Raisman, G. The glial framework of central white matter tracts: Segmented rows of contiguous interfascicular oligodendrocytes and solitary astrocytes give rise to a continuous meshwork of transverse and longitudinal processes in the adult rat fimbria. Glia 1992, 6, 222–235. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, H.; Matsumoto, T.; Ohashi, T.; Sato, M.; Kokubun, S. Comparison of spinal cord gray matter and white matter softness: Measurement by pipette aspiration method. J. Neurosurg. 2001, 95, 221–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuhrmann, T.; Hillen, L.M.; Montzka, K.; Woltje, M.; Brook, G.A. Cell-cell interactions of human neural progenitor-derived astrocytes within a microstructured 3D-scaffold. Biomaterials 2010, 31, 7705–7715. [Google Scholar] [CrossRef] [PubMed]
- Gerardo-Nava, J.; Hodde, D.; Katona, I.; Bozkurt, A.; Grehl, T.; Steinbusch, H.W.; Weis, J.; Brook, G.A. Spinal cord organotypic slice cultures for the study of regenerating motor axon interactions with 3D scaffolds. Biomaterials 2014, 35, 4288–4296. [Google Scholar] [CrossRef] [PubMed]
- Joosten, E.A.; Bar, P.R.; Gispen, W.H. Collagen implants and cortico-spinal axonal growth after mid-thoracic spinal cord lesion in the adult rat. J. Neurosci. Res. 1995, 41, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Snider, S.; Cavalli, A.; Colombo, F.; Gallotti, A.L.; Quattrini, A.; Salvatore, L.; Madaghiele, M.; Terreni, M.R.; Sannino, A.; Mortini, P. A novel composite type I collagen scaffold with micropatterned porosity regulates the entrance of phagocytes in a severe model of spinal cord injury. J. Biomed. Mater. Res. B Appl. Biomater. 2016, 105, 1040–1053. [Google Scholar] [CrossRef] [PubMed]
- Spilker, M.H.; Yannas, I.V.; Kostyk, S.K.; Norregaard, T.V.; Hsu, H.P.; Spector, M. The effects of tubulation on healing and scar formation after transection of the adult rat spinal cord. Restor. Neurol. Neurosci. 2001, 18, 23–38. [Google Scholar] [PubMed]
- Lazarus, S.S.; Trombetta, L.D. Ultrastructural identification of a benign perineurial cell tumor. Cancer 1978, 41, 1823–1829. [Google Scholar] [CrossRef]
- Macarenco, R.S.; Ellinger, F.; Oliveira, A.M. Perineurioma: A distinctive and underrecognized peripheral nerve sheath neoplasm. Arch. Pathol. Lab. Med. 2007, 131, 625–636. [Google Scholar] [CrossRef] [PubMed]
- Nolte, K.W.; Hans, V.J.; Schattenfroh, C.; Weis, J.; Schroder, J.M. Perineurial cells filled with collagen in ‘atypical’ Cogan’s syndrome. Acta Neuropathol. 2008, 115, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Katenkamp, D.; Stiller, D. Ultrastructure of perineurial cells during peripheral nerve regeneration. Electron microscopical investigations on the so-called amputation neuroma. Exp. Pathol. 1978, 16, 5–15. [Google Scholar] [CrossRef]
- Peltonen, J.; Jaakkola, S.; Virtanen, I.; Pelliniemi, L. Perineurial cells in culture. An immunocytochemical and electron microscopic study. Lab. Investig. A J. Tech. Methods Pathol. 1987, 57, 480–488. [Google Scholar]
- Weis, J.; May, R.; Schroder, J.M. Fine structural and immunohistochemical identification of perineurial cells connecting proximal and distal stumps of transected peripheral nerves at early stages of regeneration in silicone tubes. Acta Neuropathol. 1994, 88, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Pina-Oviedo, S.; Ortiz-Hidalgo, C. The normal and neoplastic perineurium: A review. Adv. Anat. Pathol. 2008, 15, 147–164. [Google Scholar] [CrossRef] [PubMed]
- Richard, L.; Topilko, P.; Magy, L.; Decouvelaere, A.V.; Charnay, P.; Funalot, B.; Vallat, J.M. Endoneurial fibroblast-like cells. J. Neuropathol. Exp. Neurol. 2012, 71, 938–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroder, J.M.; May, R.; Weis, J. Perineurial cells are the first to traverse gaps of peripheral nerves in silicone tubes. Clin. Neurol. Neurosurg. 1993, 95, S78–S83. [Google Scholar] [CrossRef]
- Parrinello, S.; Napoli, I.; Ribeiro, S.; Wingfield Digby, P.; Fedorova, M.; Parkinson, D.B.; Doddrell, R.D.; Nakayama, M.; Adams, R.H.; Lloyd, A.C. EphB signaling directs peripheral nerve regeneration through Sox2-dependent Schwann cell sorting. Cell 2010, 143, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Buss, A.; Pech, K.; Kakulas, B.A.; Martin, D.; Schoenen, J.; Noth, J.; Brook, G.A. Growth-modulating molecules are associated with invading Schwann cells and not astrocytes in human traumatic spinal cord injury. Brain 2007, 130, 940–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakulas, B.A. Pathology of spinal injuries. Cent. Nerv. Syst. Trauma 1984, 1, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Bruce, J.H.; Norenberg, M.D.; Kraydieh, S.; Puckett, W.; Marcillo, A.; Dietrich, D. Schwannosis: Role of gliosis and proteoglycan in human spinal cord injury. J. Neurotrauma 2000, 17, 781–788. [Google Scholar] [CrossRef] [PubMed]
- Guest, J.D.; Hiester, E.D.; Bunge, R.P. Demyelination and Schwann cell responses adjacent to injury epicenter cavities following chronic human spinal cord injury. Exp. Neurol. 2005, 192, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Bresnahan, J.C. An electron-microscopic analysis of axonal alterations following blunt contusion of the spinal cord of the rhesus monkey (Macaca mulatta). J. Neurol. Sci. 1978, 37, 59–82. [Google Scholar] [CrossRef]
- Bunge, M.B.; Holets, V.R.; Bates, M.L.; Clarke, T.S.; Watson, B.D. Characterization of photochemically induced spinal cord injury in the rat by light and electron microscopy. Exp. Neurol. 1994, 127, 76–93. [Google Scholar] [CrossRef] [PubMed]
- Beattie, M.S.; Bresnahan, J.C.; Komon, J.; Tovar, C.A.; Van Meter, M.; Anderson, D.K.; Faden, A.I.; Hsu, C.Y.; Noble, L.J.; Salzman, S.; et al. Endogenous repair after spinal cord contusion injuries in the rat. Exp. Neurol. 1997, 148, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Achenbach, P.; Hambeukers, I.; Pierling, A.L.; Gerardo-Nava, J.L.; Hillerbrand, L.; Sechi, A.S.; Glucks, K.J.; Dalton, P.D.; Pich, A.; Dievernich, A.; et al. A novel in vitro assay for peripheral nerve-related cell migration that preserves both extracellular matrix-derived molecular cues and nanofiber-derived topography. J. Neurosci. Methods 2021, 361, 109289. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, L.L.; Peltonen, J.; Jaakkola, S.; Gralnick, H.; Uitto, J. Plasticity of integrin expression by nerve-derived connective tissue cells. Human Schwann cells, perineurial cells, and fibroblasts express markedly different patterns of beta 1 integrins during nerve development, neoplasia, and in vitro. J. Clin. Investig. 1991, 87, 811–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Postlethwaite, A.E.; Seyer, J.M.; Kang, A.H. Chemotactic attraction of human fibroblasts to type I, II, and III collagens and collagen-derived peptides. Proc. Natl. Acad. Sci. USA 1978, 75, 871–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, R.J.; Crang, A.J.; Blakemore, W.F. The behaviour of meningeal cells following glial cell transplantation into chemically-induced areas of demyelination in the CNS. Neuropathol. Appl. Neurobiol. 1992, 18, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Klett, F.; Priller, J. The fibrotic scar in neurological disorders. Brain Pathol. 2014, 24, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Key, A.; Retzius, G. Studien in der Anatomie des Nervensystemes. Arch. Mikrosk. Anat. 1873, 9, 308–386. [Google Scholar] [CrossRef]
- Shanthaveerappa, T.R.; Bourne, G.H. New observations on the structure of the Pacinian corpuscle and its relation to the perineural epithelium of peripheral nerves. Am. J. Anat. 1963, 112, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Achtstatter, T.; Fouquet, B.; Rungger-Brandle, E.; Franke, W.W. Cytokeratin filaments and desmosomes in the epithelioid cells of the perineurial and arachnoidal sheaths of some vertebrate species. Differentiation 1989, 40, 129–149. [Google Scholar] [CrossRef] [PubMed]
- Hornick, J.L.; Fletcher, C.D. Soft tissue perineurioma: Clinicopathologic analysis of 81 cases including those with atypical histologic features. Am. J. Surg. Pathol. 2005, 29, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Perry, A.; Lusis, E.A.; Gutmann, D.H. Meningothelial hyperplasia: A detailed clinicopathologic, immunohistochemical and genetic study of 11 cases. Brain Pathol. 2005, 15, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Reina, M.; Sala-Blanch, X.; Arriazu, R.; Machés, F. Microscopic Morphology and Ultrastructure of Human Peripheral Nerves. In Nerves and Nerve Injuries; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- De Winter, F.; Oudega, M.; Lankhorst, A.J.; Hamers, F.P.; Blits, B.; Ruitenberg, M.J.; Pasterkamp, R.J.; Gispen, W.H.; Verhaagen, J. Injury-induced class 3 semaphorin expression in the rat spinal cord. Exp. Neurol. 2002, 175, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Scarlato, M.; Ara, J.; Bannerman, P.; Scherer, S.; Pleasure, D. Induction of neuropilins-1 and -2 and their ligands, Sema3A, Sema3F, and VEGF, during Wallerian degeneration in the peripheral nervous system. Exp. Neurol. 2003, 183, 489–498. [Google Scholar] [CrossRef]
- Ara, J.; Bannerman, P.; Hahn, A.; Ramirez, S.; Pleasure, D. Modulation of sciatic nerve expression of class 3 semaphorins by nerve injury. Neurochem. Res. 2004, 29, 1153–1159. [Google Scholar] [CrossRef]
- Tannemaat, M.R.; Korecka, J.; Ehlert, E.M.; Mason, M.R.; van Duinen, S.G.; Boer, G.J.; Malessy, M.J.; Verhaagen, J. Human neuroma contains increased levels of semaphorin 3A, which surrounds nerve fibers and reduces neurite extension in vitro. J. Neurosci. 2007, 27, 14260–14264. [Google Scholar] [CrossRef] [Green Version]
- Riew, T.R.; Choi, J.H.; Kim, H.L.; Jin, X.; Lee, M.Y. PDGFR-beta-Positive Perivascular Adventitial Cells Expressing Nestin Contribute to Fibrotic Scar Formation in the Striatum of 3-NP Intoxicated Rats. Front. Mol. Neurosci. 2018, 11, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaakkola, S.; Peltonen, J.; Uitto, J.J. Perineurial cells coexpress genes encoding interstitial collagens and basement membrane zone components. J. Cell Biol. 1989, 108, 1157–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghadially, F.N. Ultrastructural Pathology of the Cell and Matrix: A Text and Atlas of Physiological and Pathological Alterations in the Fine Structure of Cellular and Extracellular Components, 3rd ed.; Butterworth-Heinemann: Oxford, UK, 1988. [Google Scholar]
- Novikova, L.N.; Pettersson, J.; Brohlin, M.; Wiberg, M.; Novikov, L.N. Biodegradable poly-beta-hydroxybutyrate scaffold seeded with Schwann cells to promote spinal cord repair. Biomaterials 2008, 29, 1198–1206. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.A.; Chen, F.J.; Cui, H.L.; Lin, T.; Guo, N.; Wu, H.G. Efficacy of chitosan and sodium alginate scaffolds for repair of spinal cord injury in rats. Neural Regen. Res. 2018, 13, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Brook, G.A.; Lawrence, J.M.; Raisman, G. Columns of Schwann cells extruded into the CNS induce in-growth of astrocytes to form organized new glial pathways. Glia 2001, 33, 118–130. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Altinova, H.; Achenbach, P.; Palm, M.; Katona, I.; Hermans, E.; Clusmann, H.; Weis, J.; Brook, G.A. Characterization of a Novel Aspect of Tissue Scarring Following Experimental Spinal Cord Injury and the Implantation of Bioengineered Type-I Collagen Scaffolds in the Adult Rat: Involvement of Perineurial-like Cells? Int. J. Mol. Sci. 2022, 23, 3221. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23063221
Altinova H, Achenbach P, Palm M, Katona I, Hermans E, Clusmann H, Weis J, Brook GA. Characterization of a Novel Aspect of Tissue Scarring Following Experimental Spinal Cord Injury and the Implantation of Bioengineered Type-I Collagen Scaffolds in the Adult Rat: Involvement of Perineurial-like Cells? International Journal of Molecular Sciences. 2022; 23(6):3221. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23063221
Chicago/Turabian StyleAltinova, Haktan, Pascal Achenbach, Moniek Palm, Istvan Katona, Emmanuel Hermans, Hans Clusmann, Joachim Weis, and Gary Anthony Brook. 2022. "Characterization of a Novel Aspect of Tissue Scarring Following Experimental Spinal Cord Injury and the Implantation of Bioengineered Type-I Collagen Scaffolds in the Adult Rat: Involvement of Perineurial-like Cells?" International Journal of Molecular Sciences 23, no. 6: 3221. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23063221