
Scent of a Symbiont: The Personalized Genetic Relationships of Rhizobium—Plant Interaction



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
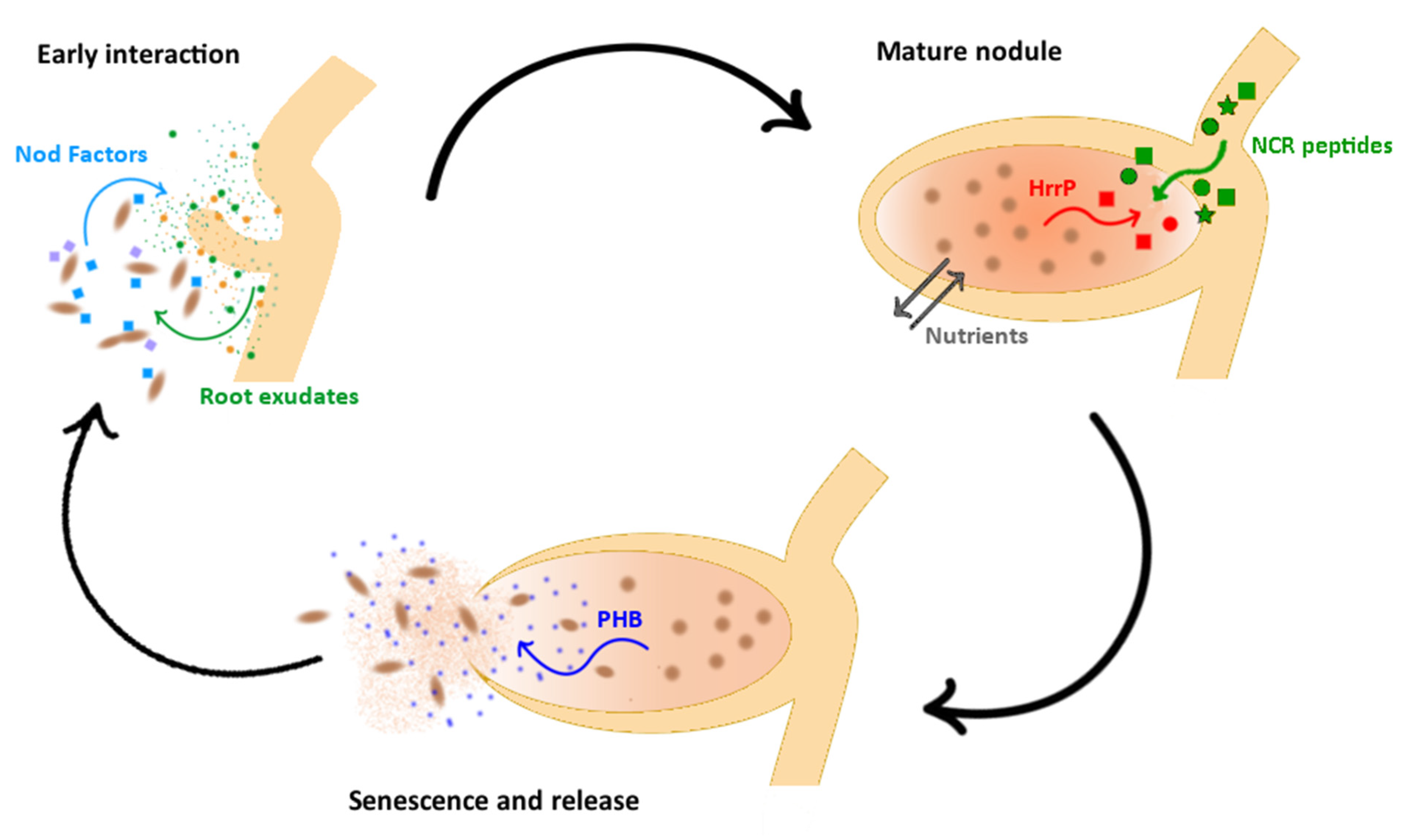
2. Evidence of Genotypic Signature in the Rhizobium—Plant Partner Choice
3. Fitness Alignment between Rhizobium and Host Plant
4. Detecting Genotype × Genotype Interaction in Nodule Colonization
5. Molecular Tasting in the Rhizosphere
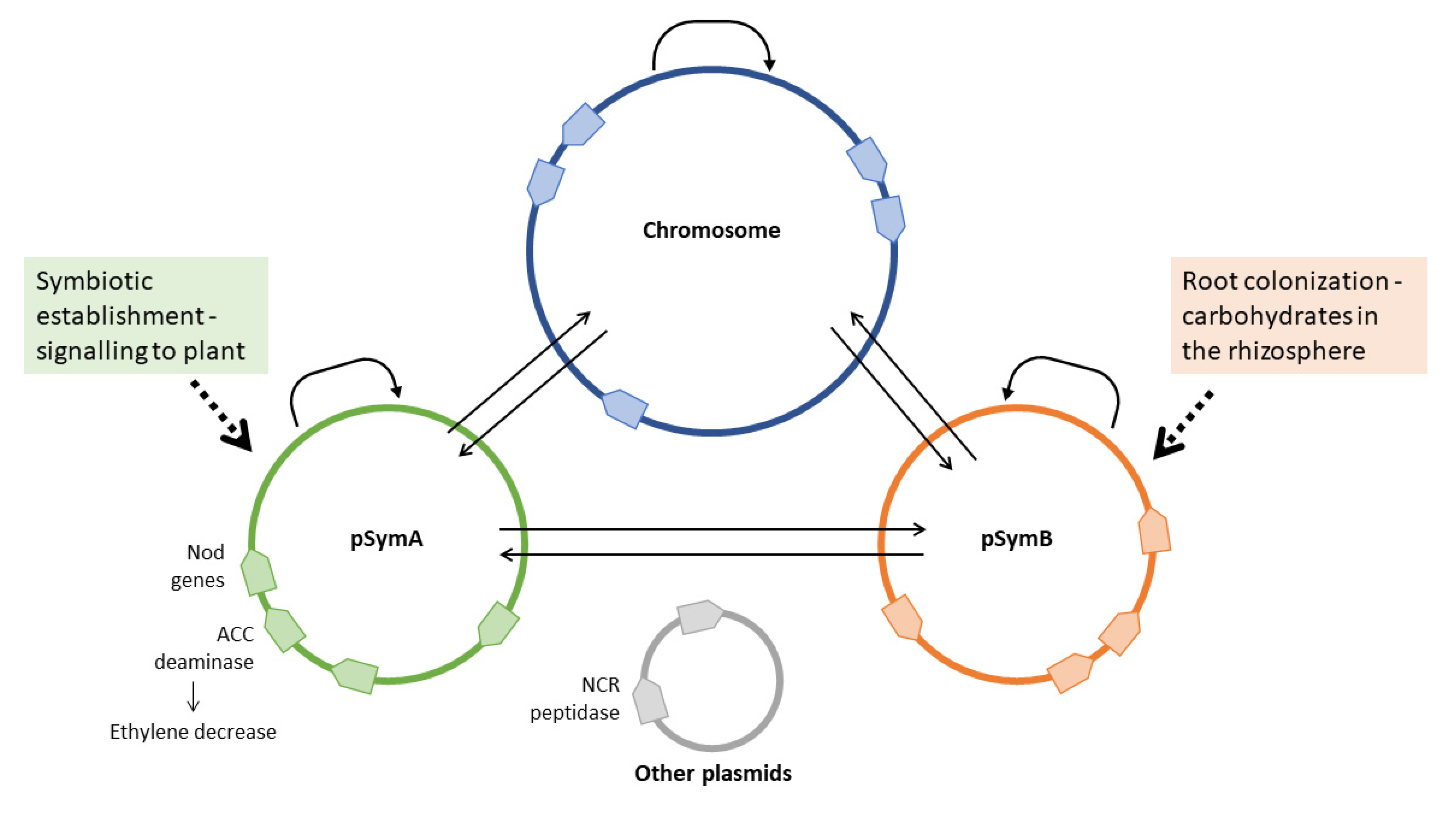
6. Role of the Rhizobial Mobile Genome in the Perception of the Plant
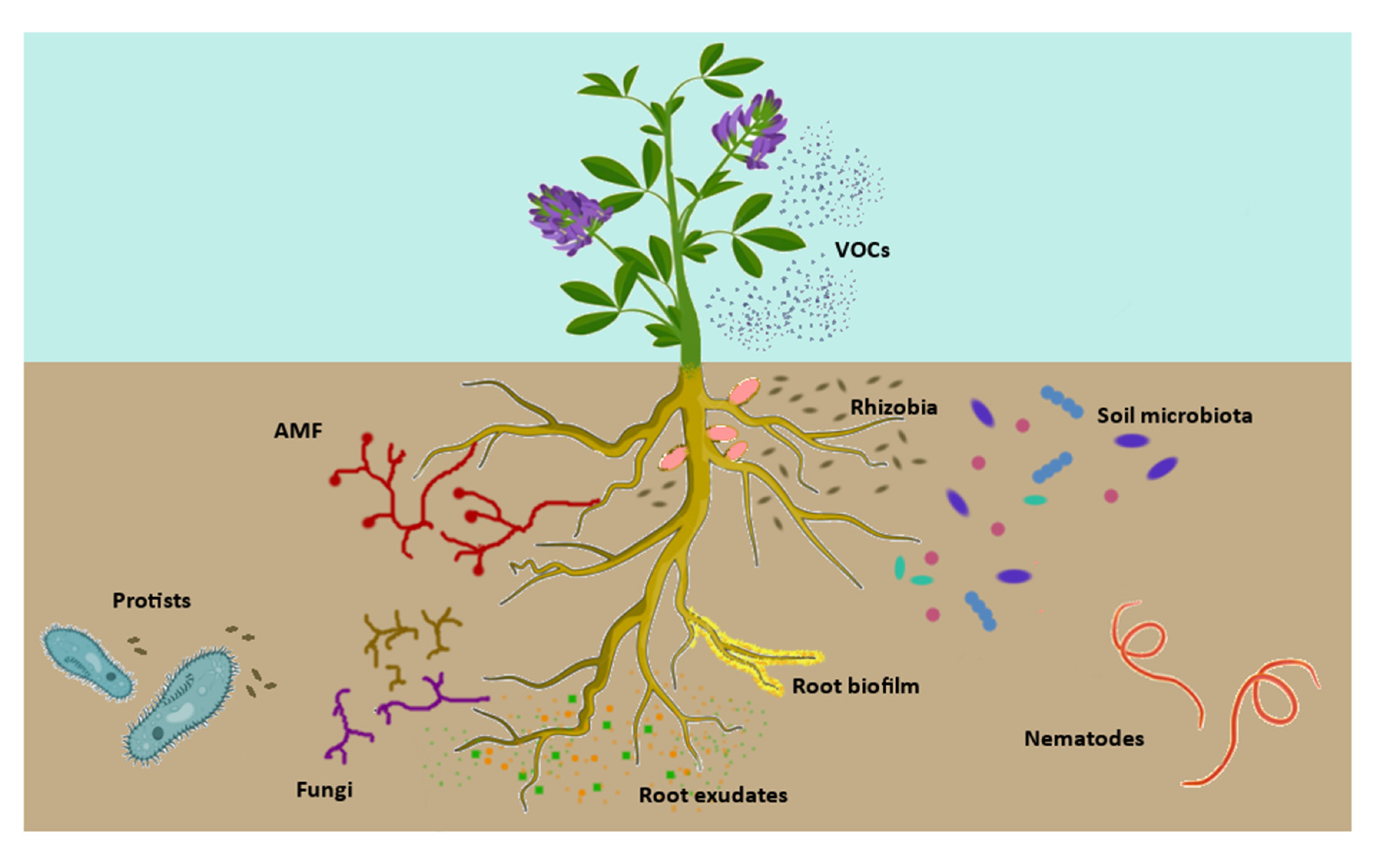
7. A Cross-Talk in the Rhizomicrobiota
8. Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- O’Brien, A.M.; Jack, C.N.; Friesen, M.L.; Frederickson, M.E. Whose trait is it anyways? Coevolution of joint phenotypes and genetic architecture in mutualisms. Proc. R. Soc. B Biol. Sci. 2021, 288, 20202483. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.R. Prioritizing host phenotype to understand microbiome heritability in plants. New Phytol. 2021, 232, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Friesen, M.L.; Porter, S.S.; Stark, S.C.; Von Wettberg, E.J.; Sachs, J.L.; Martinez-Romero, E. Microbially mediated plant functional traits. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 23–46. [Google Scholar] [CrossRef] [Green Version]
- Batstone, R.T. Genomes within genomes: Nested symbiosis and its implications for plant evolution. New Phytol. 2022, 234, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Bacci, G.; Cerri, M.; Lastrucci, L.; Ferranti, F.; Ferri, V.; Foggi, B.; Gigante, D.; Venanzoni, R.; Viciani, D.; Mengoni, A.; et al. Applying predictive models to decipher rhizobacterial modifications in common reed die-back affected populations. Sci. Total Environ. 2018, 642, 708–722. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Thiergart, T.; Vannier, N.; Mesny, F.; Ziegler, J.; Pickel, B.; Hacquard, S. A microbiota–root–shoot circuit favours Arabidopsis growth over defence under suboptimal light. Nat. Plants 2021, 7, 1078–1092. [Google Scholar] [CrossRef] [PubMed]
- do Amaral, F.P.; Tuleski, T.R.; Pankievicz, V.C.S.; Melnyk, R.A.; Arkin, A.P.; Griffitts, J.; Tadra-Sfeir, M.Z.; Maltempi de Souza, E.; Deutschbauer, A.; Monteiro, R.A.; et al. Diverse Bacterial Genes Modulate Plant Root Association by Beneficial Bacteria. mBio 2020, 11, e03078-20. [Google Scholar] [CrossRef] [PubMed]
- Levy, A.; Gonzalez, I.S.; Mittelviefhaus, M.; Clingenpeel, S.; Paredes, S.H.; Miao, J.; Wang, K.; Devescovi, G.; Stillman, K.; Monteiro, F.; et al. Genomic features of bacterial adaptation to plants. Nat. Genet. 2018, 50, 138–150. [Google Scholar] [CrossRef] [Green Version]
- Levy, A.; Conway, J.M.; Dangl, J.L.; Woyke, T. Elucidating Bacterial Gene Functions in the Plant Microbiome. Cell Host Microbe 2018, 24, 475–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulgarelli, D.; Garrido-Oter, R.; Münch, P.C.; Weiman, A.; Dröge, J.; Pan, Y.; McHardy, A.C.; Schulze-Lefert, P. Structure and function of the bacterial root microbiota in wild and domesticated barley. Cell Host Microbe 2015, 17, 392–403. [Google Scholar] [CrossRef] [Green Version]
- Poole, P.; Ramachandran, V.; Terpolilli, J. Rhizobia: From saprophytes to endosymbionts. Nat. Rev. Microbiol. 2018, 16, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Oldroyd, G.E.D.; Murray, J.D.; Poole, P.S.; Downie, J.A. The rules of engagement in the legume-rhizobial symbiosis. Annu. Rev. Genet. 2011, 45, 119–144. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, J.P.; Oresnik, I.J. The Rhizobium-Legume Symbiosis: Co-opting Successful Stress Management. Front. Plant Sci. 2022, 12, 796045. [Google Scholar] [CrossRef]
- Ledermann, R.; Schulte, C.C.M.; Poole, P.S. How Rhizobia Adapt to the Nodule Environment. J. Bacteriol. 2021, 203, e00539-20. [Google Scholar] [CrossRef] [PubMed]
- Nicoud, Q.; Barrière, Q.; Busset, N.; Bourge, M.; Le Bars, R.; Mergaert, P. Sinorhizobium meliloti functions required for resistance to antimicrobial NCR peptides and bacteroid differentiation. mbio 2021, 12, e00895-21. [Google Scholar] [CrossRef] [PubMed]
- Haag, A.F.; Arnold, M.F.F.; Myka, K.K.; Kerscher, B.; Dall’Angelo, S.; Zanda, M.; Mergaert, P.; Ferguson, G.P. Molecular insights into bacteroid development during Rhizobium–legume symbiosis. FEMS Microbiol. Rev. 2013, 37, 364–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kereszt, A.; Mergaert, P.; Kondorosi, E. Bacteroid development in legume nodules: Evolution of mutual benefit or of sacrificial victims? Mol. Plant-Microbe Interact. 2011, 24, 1300–1309. [Google Scholar] [CrossRef] [Green Version]
- Crook, M.B.; Lindsay, D.P.; Biggs, M.B.; Bentley, J.S.; Price, J.C.; Clement, S.C.; Clement, M.J.; Long, S.R.; Griffitts, J.S. Rhizobial Plasmids That Cause Impaired Symbiotic Nitrogen Fixation and Enhanced Host Invasion. Mol. Plant-Microbe Interact. 2012, 25, 1026–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazmierczak, T.; Nagymihaly, M.; Lamouche, F.; Barriére, Q.; Guefrachi, I.; Alunni, B.; Ouadghiri, M.; Ibijbijen, J.; Kondorosi, É.; Mergaert, P.; et al. Specific host-responsive associations between Medicago truncatula accessions and Sinorhizobium strains. Mol. Plant-Microbe Interact. 2017, 30, 399–409. [Google Scholar] [CrossRef] [Green Version]
- Dicenzo, G.C.; Zamani, M.; Checcucci, A.; Fondi, M.; Griffitts, J.S.; Finan, T.M.; Mengoni, A. Multidisciplinary approaches for studying rhizobium–legume symbioses. Can. J. Microbiol. 2019, 65, 1–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, S.; Liu, W.; Nandety, R.S.; Crook, A.D.; Mysore, K.S.; Pislariu, C.I.; Frugoli, J.A.; Dickstein, R.; Udvardi, M.K. Celebrating 20 years of genetic discoveries in legume nodulation and symbiotic nitrogen fixation. Plant Cell 2020, 32, 15–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carelli, M.; Gnocchi, S.; Fancelli, S.; Mengoni, A.; Paffetti, D.; Scotti, C.; Bazzicalupo, M. Genetic Diversity and Dynamics of Sinorhizobium meliloti Populations Nodulating Different Alfalfa Cultivars in Italian Soils. Appl. Environ. Microbiol. 2000, 66, 4785–4789. [Google Scholar] [CrossRef] [Green Version]
- Paffetti, D.; Scotti, C.; Gnocchi, S.; Fancelli, S. Genetic Diversity of an Italian Rhizobium meliloti Population from Different Medicago sativa Varieties. Appl. Environ. Microbiol. 1996, 62, 2279–2285. [Google Scholar] [CrossRef] [Green Version]
- Kumar, N.; Lad, G.; Giuntini, E.; Kaye, M.E.; Udomwong, P.; Shamsani, N.J.; Young, J.P.W.; Bailly, X. Bacterial genospecies that are not ecologically coherent: Population genomics of Rhizobium leguminosarum. Open Biol. 2015, 5, 140133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailly, X.; Giuntini, E.; Sexton, M.C.; Lower, R.P.J.; Harrison, P.W.; Kumar, N.; Young, J.P.W. Population genomics of Sinorhizobium medicae based on low-coverage sequencing of sympatric isolates. ISME J. 2011, 5, 1722–1734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, W.-J.; Zhang, B.; Zhao, R.; Liu, L.-X.; Jiao, J.; Zhang, Z.; Tian, C.-F. Lineage-Specific Rewiring of Core Pathways Predating Innovation of Legume Nodules Shapes Symbiotic Efficiency. mSystems 2021, 6, e01299-20. [Google Scholar] [CrossRef] [PubMed]
- Epstein, B.; Branca, A.; Mudge, J.; Bharti, A.K.; Briskine, R.; Farmer, A.D.; Sugawara, M.; Young, N.D.; Sadowsky, M.J.; Tiffin, P. Population Genomics of the Facultatively Mutualistic Bacteria Sinorhizobium meliloti and S. medicae. Genomics 2012, 8, e1002868. [Google Scholar] [CrossRef] [PubMed]
- Jorrin, B.; Imperial, J. Population genomics analysis of legume host preference for specific rhizobial genotypes in the Rhizobium leguminosarum bv. viciae symbioses. Mol. Plant-Microbe Interact. 2015, 28, 310–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailly, X.; Olivier, I.; De Mita, S.; Cleyet-Marel, J.-C.; Béna, G. Recombination and selection shape the molecular diversity pattern of nitrogen-fixing Sinorhizobium sp. associated to Medicago. Mol. Ecol. 2006, 15, 2719–2734. [Google Scholar] [CrossRef] [PubMed]
- Burghardt, L.T.; Trujillo, D.I.; Epstein, B.; Tiffin, P.; Young, N.D. A Select and Resequence Approach Reveals Strain-Specific Effects of Medicago Nodule-Specific PLAT-Domain Genes. Plant Physiol. 2020, 182, 463–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mengoni, A.; Gori, A.; Bazzicalupo, M. Use of RAPD and microsatellite (SSR) variation to assess genetic relationships among populations of tetraploid alfalfa, Medicago sativa. Plant Breed. 2000, 119, 311–317. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, J.; Li, H.; Yang, S.; Kereszt, A.; Zhu, H. Nodule-Specific Cysteine-Rich Peptides Negatively Regulate Nitrogen-Fixing Symbiosis in a Strain-Specific Manner in Medicago truncatula. Mol. Plant-Microbe Interact. 2018, 31, 240–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haag, A.F.; Baloban, M.; Sani, M.; Kerscher, B.; Pierre, O.; Angelo, S.D.; Kondorosi, E.; Longhi, R.; Boncompagni, E.; He, D.; et al. Protection of Sinorhizobium against Host Cysteine-Rich Antimicrobial Peptides Is Critical for Symbiosis. PLoS Biol. 2011, 9, e1001169. [Google Scholar] [CrossRef] [PubMed]
- DiCenzo, G.C.; Zamani, M.; Ludwig, H.N.; Finan, T.M. Heterologous Complementation Reveals a Specialized Activity for BacA in the Medicago—Sinorhizobium meliloti Symbiosis. Mol. Plant Microbe Interact. 2017, 30, 312–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denison, R.F.; Kiers, E.T. Life histories of symbiotic rhizobia and mycorrhizal fungi. Curr. Biol. 2011, 21, R775–R785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friesen, M.L. Widespread fitness alignment in the legume-rhizobium symbiosis. New Phytol. 2012, 194, 1096–1111. [Google Scholar] [CrossRef] [PubMed]
- diCenzo, G.C.; Tesi, M.; Pfau, T.; Mengoni, A.; Fondi, M. Genome-scale metabolic reconstruction of the symbiosis between a leguminous plant and a nitrogen-fixing bacterium. Nat. Commun. 2020, 11, 2574. [Google Scholar] [CrossRef] [PubMed]
- Ratcliff, W.C.; Underbakke, K.; Denison, R.F. Measuring the fitness of symbiotic rhizobia. Symbiosis 2012, 55, 85–90. [Google Scholar] [CrossRef]
- Denison, R.F.; Kiers, E.T. Lifestyle alternatives for rhizobia: Mutualism, parasitism, and forgoing symbiosis. FEMS Microbiol. Lett. 2004, 237, 187–193. [Google Scholar] [CrossRef]
- Checcucci, A.; Azzarello, E.; Bazzicalupo, M.; Galardini, M.; Lagomarsino, A.; Mancuso, S.; Marti, L.; Marzano, M.C.; Mocali, S.; Squartini, A.; et al. Mixed nodule infection in Sinorhizobium meliloti-medicago sativa symbiosis suggest the presence of cheating behavior. Front. Plant Sci. 2016, 7, 835. [Google Scholar] [CrossRef] [Green Version]
- Bellabarba, A.; Bacci, G.; Decorosi, F.; Aun, E.; Azzarello, E.; Remm, M.; Giovannetti, L. Competitiveness for Nodule Colonization in Sinorhizobium meliloti: Combined In Vitro-Tagged Strain Competition and Genome-Wide Association Analysis. mSystems 2021, 6, e00550-21. [Google Scholar] [CrossRef]
- Gano-Cohen, K.A.; Wendlandt, C.E.; Stokes, P.J.; Blanton, M.A.; Quides, K.W.; Zomorrodian, A.; Adinata, E.S.; Sachs, J.L. Interspecific conflict and the evolution of ineffective rhizobia. Ecol. Lett. 2019, 22, 914–924. [Google Scholar] [CrossRef]
- Sachs, J.L.; Simms, E.L. The origins of uncooperative rhizobia. Oikos 2008, 117, 961–966. [Google Scholar] [CrossRef]
- Batstone, R.T.; Dutton, E.M.; Wang, D.; Yang, M.; Frederickson, M.E. The evolution of symbiont preference traits in the model legume Medicago truncatula. New Phytol. 2017, 213, 1850–1861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heath, K.D.; Tiffin, P. Stabilizing mechanisms in a legume-rhizobium mutualism. Evolution 2009, 63, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Kiers, E.T.; Rousseau, R.A.; Denison, R.F. Measured sanctions: Legume hosts detect quantitative variation in rhizobium cooperation and punish accordingly. Evol. Ecol. 2006, 1077–1086. [Google Scholar]
- Burghardt, L.T.; Epstein, B.; Guhlin, J.; Nelson, M.S.; Taylor, M.R.; Young, N.D.; Sadowsky, M.J.; Tiffin, P. Select and resequence reveals relative fitness of bacteria in symbiotic and free-living environments. Proc. Natl. Acad. Sci. USA 2018, 115, 2425–2430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pobigaylo, N.; Szymczak, S.; Nattkemper, T.W.; Becker, A. Identification of genes relevant to symbiosis and competitiveness in Sinorhizobium meliloti using signature-tagged mutants. Mol. Plant-Microbe Interact. 2008, 21, 219–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burghardt, L.T. Evolving together, evolving apart: Measuring the fitness of rhizobial bacteria in and out of symbiosis with leguminous plants. New Phytol. 2020, 228, 28–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- di Cenzo, G.C.; Zamani, M.; Milunovic, B.; Finan, T.M. Genomic resources for identification of the minimal N 2 -fixing symbiotic genome. Environ. Microbiol. 2016, 18, 2534–2547. [Google Scholar] [CrossRef] [PubMed]
- Geddes, B.A.; Kearsley, J.V.S.; Huang, J.; Zamani, M.; Muhammed, Z.; Sather, L. Minimal gene set from Sinorhizobium (Ensifer) meliloti pSymA required for efficient symbiosis with Medicago. Proc. Natl. Acad. Sci. USA 2021, 118, e2018015118. [Google Scholar] [CrossRef] [PubMed]
- Checcucci, A.; Dicenzo, G.C.; Ghini, V.; Bazzicalupo, M.; Becker, A.; Decorosi, F.; Döhlemann, J.; Fagorzi, C.; Finan, T.M.; Fondi, M.; et al. Creation and Characterization of a Genomically Hybrid Strain in the Nitrogen-Fixing Symbiotic Bacterium Sinorhizobium meliloti. ACS Synth. Biol. 2018, 7, 2365–2378. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.M.; Kobayashi, H.; Davies, B.W.; Taga, M.E.; Walker, G.C. How rhizobial symbionts invade plants: The Sinorhizobium—Medicago model. Nature 2007, 5, 619–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burghardt, L.T.; Guhlin, J.; Chun, C.L.; Liu, J.; Sadowsky, M.J.; Stupar, R.M.; Young, N.D.; Tiffin, P. Transcriptomic basis of genome by genome variation in a legume-rhizobia mutualism. Mol. Ecol. 2017, 26, 6122–6135. [Google Scholar] [CrossRef]
- Fagorzi, C.; Bacci, G.; Huang, R.; Cangioli, L.; Checcucci, A.; Fini, M.; Perrin, E.; Natali, C.; diCenzo, G.C.; Mengoni, A. Nonadditive transcriptomic signatures of genotype-by-genotype interactions during the initiation of plant-rhizobium symbiosis. mSystems 2021, 6, e00974-20. [Google Scholar] [CrossRef] [PubMed]
- West, S.A.; Kiers, E.T.; Simms, E.L.; Denison, R.F. Sanctions and mutualism stability: Why do rhizobia fix nitrogen? Proc. Biol. Sci. 2002, 269, 685–694. [Google Scholar] [CrossRef] [Green Version]
- Kiers, E.T.; Rousseau, R.A.; West, S.A.; Denison, R.F. Host sanctions and the legume–rhizobium mutualism. Nature 2003, 425, 78–81. [Google Scholar] [CrossRef]
- Porter, S.S.; Simms, E.L. Selection for cheating across disparate environments in the legume-rhizobium mutualism. Ecol. Lett. 2014, 17, 1121–1129. [Google Scholar] [CrossRef] [PubMed]
- Gubry-Rangin, C.; Garcia, M.; Béna, G. Partner choice in Medicago truncatula-Sinorhizobium symbiosis. Proc. Biol. Sci. 2010, 277, 1947–1951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogel, M.A.; Ormeno-Orrillo, E.; Martinez-Romero, E. Symbiovars in rhizobia reflect bacterial adaptation to legumes. Appl. Microbiol. 2011, 34, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Mergaert, P.; Van Montagu, M.; Holsters, M. Molecular mechanisms of Nod factor diversity. Mol. Microbiol. 1997, 25, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.T.; Young, J.P.W. Symbiosis Genes: Organisation and Diversity. In Ecology and Evolution of Rhizobia; Springer: Berlin/Heidelberg, Germany, 2019; pp. 123–144. [Google Scholar]
- Andrews, M.; Andrews, M.E. Specificity in Legume-Rhizobia Symbioses. Int. J. Mol. Sci. 2017, 18, 705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Souza, R.S.C.; Armanhi, J.S.L.; Damasceno, N.d.B.; Imperial, J.; Arruda, P. Genome Sequences of a Plant Beneficial Synthetic Bacterial Community Reveal Genetic Features for Successful Plant Colonization. Front. Microbiol. 2019, 10, 1779. [Google Scholar] [CrossRef] [Green Version]
- Pini, F.; Galardini, M.; Bazzicalupo, M.; Mengoni, A. Plant-Bacteria Association and Symbiosis: Are There Common Genomic Traits in Alphaproteobacteria? Genes 2011, 2, 1017–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.; Halane, M.K.; Gassmann, W.; Stacey, G. The Role of Plant Innate Immunity in the Legume-Rhizobium Symbiosis. Annu. Rev. Plant Biol 2017, 6812, 535–561. [Google Scholar] [CrossRef] [PubMed]
- Gourion, B.; Berrabah, F.; Ratet, P.; Stacey, G. Rhizobium-legume symbioses: The crucial role of plant immunity. Trends Plant Sci. 2015, 20, 186–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katalin, T.; Stacey, G. Does plant immunity play a critical role during initiation of the legume-rhizobium symbiosis? Front. Plant Sci. 2015, 6, 401. [Google Scholar] [CrossRef]
- Nelson, M.S.; Sadowsky, M.J. Secretion systems and signal exchange between nitrogen-fixing rhizobia and legumes. Front. Plant Sci. 2015, 6, 491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marczak, M.; Mazur, A.; Koper, P.; Żebracki, K.; Skorupska, A. Synthesis of rhizobial exopolysaccharides and their importance for symbiosis with legume plants. Genes 2017, 8, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niehaus, K.; Kapp, D.; Pühler, A. Plant defence and delayed infection of alfalfa pseudonodules induced by an exopolysaccharide (EPS I)-deficient Rhizobium meliloti mutant. Planta 1993, 190, 415–425. [Google Scholar] [CrossRef]
- Liu, Y.H.; Wang, E.T.; Jiao, Y.S.; Tian, C.F.; Wang, L.; Wang, Z.J.; Guan, J.J.; Singh, R.P.; Chen, W.X.; Chen, W.F. Symbiotic characteristics of Bradyrhizobium diazoefficiens USDA 110 mutants associated with shrubby sophora (Sophora flavescens) and soybean (Glycine max). Microbiol. Res. 2018, 214, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Salas, M.E.; Lozano, M.J.; López, J.L.; Draghi, W.O.; Serrania, J.; Torres Tejerizo, G.A.; Albicoro, F.J.; Nilsson, J.F.; Pistorio, M.; Del Papa, M.F.; et al. Specificity traits consistent with legume-rhizobia coevolution displayed by Ensifer meliloti rhizosphere colonization. Environ. Microbiol. 2017, 19, 3423–3438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagorzi, C.; Bacci, G.; Huang, R.; Cangioli, L.; Checcucci, A.; Fini, M.; Perrin, E.; Natali, C.; DiCenzo, G.C.; Mengoni, A. Dissecting transcriptomic signatures of genotype x genotype interactions during the initiation of plant-rhizobium symbiosis. bioRxiv 2020. [Google Scholar] [CrossRef]
- Benedict, A.B.; Ghosh, P.; Scott, S.M.; Griffits, J.S. A conserved rhizobial peptidase that interacts with host-derived symbiotic peptides. Sci. Rep. 2021, 11, 11779. [Google Scholar] [CrossRef] [PubMed]
- Perret, X.; Staehelin, C. Molecular Basis of Symbiotic Promiscuity. Microbiol. Mol. Biol. Rev. 2000, 64, 180–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cangioli, L.; Checcucci, A.; Mengoni, A.; Fagorzi, C. Legume tasters: Symbiotic rhizobia host preference and smart inoculant formulations. Biol. Commun. 2021, 66, 47–54. [Google Scholar] [CrossRef]
- diCenzo, G.C.; MacLean, A.M.; Milunovic, B.; Golding, G.B.; Finan, T.M. Examination of Prokaryotic Multipartite Genome Evolution through Experimental Genome Reduction. PLoS Genet. 2014, 10, e1004742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galardini, M.; Pini, F.; Bazzicalupo, M.; Biondi, E.G.; Mengoni, A. Replicon-dependent bacterial genome evolution: The case of Sinorhizobium meliloti. Genome Biol. Evol. 2013, 5, 542–558. [Google Scholar] [CrossRef] [PubMed]
- DiCenzo, G.C.; Checcucci, A.; Bazzicalupo, M.; Mengoni, A.; Viti, C.; Dziewit, L.; Finan, T.M.; Galardini, M.; Fondi, M. Metabolic modelling reveals the specialization of secondary replicons for niche adaptation in Sinorhizobium meliloti. Nat. Commun. 2016, 7, 12219. [Google Scholar] [CrossRef]
- Oono, R.; Anderson, C.G.; Denison, R.F. Failure to fix nitrogen by non-reproductive symbiotic rhizobia triggers host sanctions that reduce fitness of their reproductive clonemates. Proc. R. Soc. B Biol. Sci. 2011, 278, 2698–2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mergaert, P.; Nikovics, K.; Kelemen, Z.; Maunoury, N.; Vaubert, D.; Kondorosi, A.; Kondorosi, E. A novel family in Medicago truncatula consisting of more than 300 nodule-specific genes coding for small, secreted polypeptides with conserved cysteine motifs. Plant Physiol. 2003, 132, 161–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Velde, W.; Zehirov, G.; Szatmari, A.; Debreczeny, M.; Ishihara, H.; Kevei, Z.; Farkas, A.; Mikulass, K.; Nagy, A.; Tiricz, H. Plant peptides govern terminal differentiation of bacteria in symbiosis. Science 2010, 327, 1122–1126. [Google Scholar] [CrossRef] [PubMed]
- Horváth, B.; Domonkos, Á.; Kereszt, A.; Szűcs, A.; Ábrahám, E.; Ayaydin, F.; Bóka, K.; Chen, Y.; Chen, R.; Murray, J.D. Loss of the nodule-specific cysteine rich peptide, NCR169, abolishes symbiotic nitrogen fixation in the Medicago truncatula dnf7 mutant. Proc. Natl. Acad. Sci. USA 2015, 112, 15232–15237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Chen, Y.; Xi, J.; Waters, C.; Chen, R.; Wang, D. An antimicrobial peptide essential for bacterial survival in the nitrogen-fixing symbiosis. Proc. Natl. Acad. Sci. USA 2015, 112, 201500123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Downie, J.A.; Kondorosi, E. Why Should Nodule Cysteine-Rich (NCR) Peptides Be Absent From Nodules of Some Groups of Legumes but Essential for Symbiotic N-Fixation in Others? Front. Agron. 2021, 3, 654576. [Google Scholar] [CrossRef]
- Nallu, S.; Silverstein, K.A.T.; Zhou, P.; Young, N.D.; VandenBosch, K.A. Patterns of divergence of a large family of nodule cysteine-rich peptides in accessions of Medicago truncatula. Plant J. 2014, 78, 697–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clay, K.; Kover, P.X. The Red Queen Hypothesis and Plant/Pathogen Interactions. Annu. Rev. Phytopathol. 1996, 34, 29–50. [Google Scholar] [CrossRef] [PubMed]
- Price, P.A.; Tanner, H.R.; Dillon, B.A.; Shabab, M.; Walker, G.C.; Griffitts, J.S. Rhizobial peptidase HrrP cleaves host-encoded signaling peptides and mediates symbiotic compatibility. Proc. Natl. Acad. Sci. USA 2015, 112, 201417797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Checcucci, A.; Azzarello, E.; Bazzicalupo, M.; De Carlo, A.; Emiliani, G.; Mancuso, S.; Spini, G.; Viti, C.; Mengoni, A. Role and regulation of ACC deaminase gene in Sinorhizobium meliloti: Is it a symbiotic, rhizospheric or endophytic gene? Front. Genet. 2017, 8, 6. [Google Scholar] [CrossRef]
- Okazaki, S.; Nukui, N.; Sugawara, M.; Minamisawa, K. Rhizobial Strategies to Enhance Symbiotic Interactions: Rhizobitoxine and 1-Aminocyclopropane-1-Carboxylate Deaminase. Microbes Environ. 2004, 19, 99–111. [Google Scholar] [CrossRef] [Green Version]
- Ratcliff, W.C.; Denison, R.F. Rhizobitoxine producers gain more poly-3-hydroxybutyrate in symbiosis than do competing rhizobia, but reduce plant growth. ISME J. 2009, 3, 870–872. [Google Scholar] [CrossRef]
- Ratcliff, W.C.; Kadam, S.V.; Denison, R.F. Poly-3-hydroxybutyrate (PHB) supports survival and reproduction in starving rhizobia. FEMS Microbiol. Ecol. 2008, 65, 391–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Saldanha, M.; Sheng, X.; Shelswell, K.J.; Walsh, K.T.; Sobral, B.W.S.; Charles, T.C. Roles of poly-3-hydroxybutyrate (PHB) and glycogen in symbiosis of Sinorhizobium meliloti with Medicago sp. Microbiology 2007, 153, 388–398. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.S.; Liu, Y.H.; Yan, H.; Wang, E.T.; Tian, C.F.; Chen, W.X.; Guo, B.L.; Chen, W.F. Rhizobial diversity and nodulation characteristics of the extremely promiscuous legume Sophora flavescens. Mol. Plant-Microbe Interact. 2015, 28, 1338–1352. [Google Scholar] [CrossRef] [Green Version]
- Ratcliff, W.C.; Denison, R.F. Report Individual-Level Bet Hedging in the Bacterium Sinorhizobium meliloti. CURBIO 2010, 20, 1740–1744. [Google Scholar] [CrossRef] [Green Version]
- Rolli, E.; Vergani, L.; Ghitti, E.; Patania, G.; Mapelli, F.; Borin, S. ‘Cry-for-help’ in contaminated soil: A dialogue among plants and soil microbiome to survive in hostile conditions. Environ. Microbiol. 2021, 23, 5690–5703. [Google Scholar] [CrossRef] [PubMed]
- Schulz-Bohm, K.; Gerards, S.; Hundscheid, M.; Melenhorst, J.; De Boer, W.; Garbeva, P. Calling from distance: Attraction of soil bacteria by plant root volatiles. ISME J. 2018, 12, 1252–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de la Porte, A.; Schmidt, R.; Yergeau, É.; Constant, P. A Gaseous Milieu: Extending the Boundaries of the Rhizosphere. Trends Microbiol. 2020, 28, 536–542. [Google Scholar] [CrossRef]
- Liu, H.; Brettell, L.E. Plant Defense by VOC-Induced Microbial Priming. Trends Plant Sci. 2019, 24, 187–189. [Google Scholar] [CrossRef] [PubMed]
- Soto, M.J.; López-Lara, I.M.; Geiger, O.; Romero-Puertas, M.C.; van Dillewijn, P. Rhizobial Volatiles: Potential New Players in the Complex Interkingdom Signaling With Legumes. Front. Plant Sci. 2021, 12, 1219. [Google Scholar] [CrossRef] [PubMed]
- Venturi, V.; Keel, C. Signaling in the Rhizosphere. Trends Plant Sci. 2016, 21, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Chen, H.; Eberhard, A.; Gronquist, M.R.; Robinson, J.B.; Rolfe, B.G.; Bauer, W.D. sinI- and expR-Dependent Quorum Sensing in Sinorhizobium meliloti. J. Bacteriol. 2005, 187, 7931–7944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Contreras, M.; Bauer, W.D.; Gao, M.; Robinson, J.B.; Downie, J.A. Quorum-sensing regulation in rhizobia and its role in symbiotic interactions with legumes. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 1149–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, M.; Teplitski, M.; Robinson, J.B.; Bauer, W.D. Production of substances by Medicago truncatula that affect bacterial quorum sensing. Mol. Plant-Microbe Interact. 2003, 16, 827–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Romero, E.; Aguirre-Noyola, J.L.; Taco-Taype, N.; Martínez-Romero, J.; Zuñiga-Dávila, D. Plant microbiota modified by plant domestication. Syst. Appl. Microbiol. 2020, 43, 126106. [Google Scholar] [CrossRef]
- Chaparro, J.M.; Badri, D.V.; Vivanco, J.M. Rhizosphere microbiome assemblage is affected by plant development. ISME J. 2014, 8, 790–803. [Google Scholar] [CrossRef] [Green Version]
- Schütz, V.; Frindte, K.; Cui, J.; Zhang, P.; Hacquard, S.; Schulze-Lefert, P.; Knief, C.; Schulz, M.; Dörmann, P. Differential Impact of Plant Secondary Metabolites on the Soil Microbiota. Front. Microbiol. 2021, 12, 1267. [Google Scholar] [CrossRef] [PubMed]
- Knee, E.M.; Gong, F.-C.; Gao, M.; Teplitski, M.; Jones, A.R.; Foxworthy, A.; Mort, A.J.; Bauer, W.D. Root mucilage from pea and its utilization by rhizosphere bacteria as a sole carbon source. Mol. Plant-Microbe Interact. 2001, 14, 775–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moody, S.F.; Clarke, A.E.; Bacic, A. Structural analysis of secreted slime from wheat and cowpea roots. Phytochemistry 1988, 27, 2857–2861. [Google Scholar] [CrossRef]
- Bringhurst, R.M.; Cardon, Z.G.; Gage, D.J. Galactosides in the rhizosphere: Utilization by Sinorhizobium meliloti and development of a biosensor. Proc. Natl. Acad. Sci. USA 2001, 98, 4540–4545. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Jiang, X.; Guan, D.; Zhou, W.; Ma, M.; Zhao, B.; Cao, F.; Li, L.; Li, J. Transcriptional analysis of genes involved in competitive nodulation in Bradyrhizobium diazoefficiens at the presence of soybean root exudates. Sci. Rep. 2017, 7, 10946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klonowska, A.; Melkonian, R.; Miché, L.; Tisseyre, P.; Moulin, L. Transcriptomic profiling of Burkholderia phymatum STM815, Cupriavidus taiwanensis LMG19424 and Rhizobium mesoamericanum STM3625 in response to Mimosa pudica root exudates illuminates the molecular basis of their nodulation competitiveness and symbiotic ev. BMC Genomics 2018, 19, 105. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Li, Y.-Y.; Wu, H.-M.; Zhang, F.-F.; Li, C.-J.; Li, X.-X.; Lambers, H.; Li, L. Root exudates drive interspecific facilitation by enhancing nodulation and N2 fixation. Proc. Natl. Acad. Sci. USA 2016, 113, 6496–6501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.W.; Murray, J.D. The role of flavonoids in nodulation host-range specificity: An update. Plants 2016, 5, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peck, M.C.; Fisher, R.F.; Long, S.R. Diverse Flavonoids Stimulate NodD1 Binding to nod Gene Promoters in Sinorhizobium meliloti. J. Bacteriol. 2006, 188, 5417–5427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spini, G.; Decorosi, F.; Cerboneschi, M.; Tegli, S.; Mengoni, A.; Viti, C.; Giovannetti, L. Effect of the plant flavonoid luteolin on Ensifer meliloti 3001 phenotypic responses. Plant Soil 2016, 399, 159–178. [Google Scholar] [CrossRef] [Green Version]
- Compton, K.K.; Hildreth, S.B.; Helm, R.F.; Scharf, B.E. An Updated Perspective on Sinorhizobium meliloti Chemotaxis to Alfalfa Flavonoids. Front. Microbiol. 2020, 11, 581482. [Google Scholar] [CrossRef]
- Janczarek, M.; Rachwał, K.; Marzec, A.; Grzadziel, J.; Palusińska-Szysz, M. Signal molecules and cell-surface components involved in early stages of the legume-rhizobium interactions. Appl. Soil Ecol. 2014, 85, 94–113. [Google Scholar] [CrossRef]
- Di Cenzo, G.C.; Finan, T.M. The Divided Bacterial Genome: Structure, Function, and Evolution. Microbiol. Mol. Biol. Rev. 2017, 81, e00019-17. [Google Scholar]
- Galibert, F.; Finan, T.M.; Long, S.R.; Puhler, A.; Abola, P.; Ampe, F.; Barloy-Hubler, F.; Barnett, M.J.; Becker, A.; Boistard, P.; et al. The composite genome of the legume symbiont Sinorhizobium meliloti. Science 2001, 293, 668–672. [Google Scholar] [CrossRef] [Green Version]
- Santamarı, R.I.; Medrano-soto, A.; Bustos, P.; Herna, I.; Moreno-hagelsieb, G.; Janga, S.C.; Ramı, M.A.; Collado-vides, J.; Da, G. The partitioned Rhizobium etli genome: Genetic and metabolic redundancy in seven interacting replicons. Proc. Natl. Acad. Sci. USA 2006, 103, 3834–3839. [Google Scholar]
- Young, J.P.W.; Crossman, L.C.; Johnston, A.W.B.; Thomson, N.R.; Ghazoui, Z.F.; Hull, K.H.; Wexler, M.; Curson, A.R.J.; Todd, J.D.; Poole, P.S.; et al. The genome of Rhizobium leguminosarum has recognizable core and accessory components. Young 2006, 7, R34. [Google Scholar] [CrossRef] [Green Version]
- Fagorzi, C.; Ilie, A.; Decorosi, F.; Cangioli, L.; Viti, C.; Mengoni, A.; diCenzo, G.C. Symbiotic and non-symbiotic members of the genus Ensifer (syn. Sinorhizobium) are separated into two clades based on comparative genomics and high-throughput phenotyping. Genome Biol. Evol. 2020, 12, 2521–2534. [Google Scholar] [CrossRef] [PubMed]
- Ochman, H.; Moran, N.A. Genes lost and genes found: Evolution of bacterial pathogenesis and symbiosis. Science 2001, 292, 1096–1099. [Google Scholar] [CrossRef] [Green Version]
- Barnett, M.J.; Fisher, R.F.; Jones, T.; Komp, C.; Abola, A.P.; Gurjal, M.; Hong, A.; Huizar, L.; Bowser, L.; Capela, D.; et al. Nucleotide sequence and predicted functions of the entire Sinorhizobium meliloti pSymA megaplasmid. Proc. Natl. Acad. Sci. USA 2001, 98, 9883–9888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oresnik, I.J.; Liu, S.-L.; Yost, C.K.; Hynes, M.F. Megaplasmid pRme2011a of Sinorhizobium meliloti is not required for viability. J. Bacteriol. 2000, 182, 3582–3586. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Charles, T.C.; Glick, B.R. Expression of an exogenous 1-aminocyclopropane-1-carboxylate deaminase gene in Sinorhizobium meliloti increases its ability to nodulate alfalfa. Appl. Environ. Microbiol. 2004, 70, 5891–5897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pini, F.; Spini, G.; Galardini, M.; Bazzicalupo, M.; Benedetti, A.; Chiancianesi, M.; Florio, A.; Lagomarsino, A.; Migliore, M.; Mocali, S.; et al. Molecular phylogeny of the nickel-resistance gene nreB and functional role in the nickel sensitive symbiotic nitrogen fixing bacterium Sinorhizobium meliloti. Plant Soil 2013, 377, 189–201. [Google Scholar] [CrossRef]
- Galardini, M.; Brilli, M.; Spini, G.; Rossi, M.; Roncaglia, B.; Bani, A.; Chiancianesi, M.; Moretto, M.; Engelen, K.; Bacci, G.; et al. Evolution of Intra-specific Regulatory Networks in a Multipartite Bacterial Genome. PLoS Comput. Biol. 2015, 11, e1004478. [Google Scholar] [CrossRef] [PubMed]
- diCenzo, G.C.; Benedict, A.B.; Fondi, M.; Walker, G.C.; Finan, T.M.; Mengoni, A.; Griffitts, J.S. Robustness encoded across essential and accessory replicons of the ecologically versatile bacterium Sinorhizobium meliloti. PLoS Genet. 2018, 14, e1007357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.; Mao, Y.; Zhu, Q.; Ling, J.; Zhang, N.; Naseer, N.; Zhong, Z.; Zhu, J. The quorum sensing regulator CinR hierarchically regulates two other quorum sensing pathways in ligand-dependent and-independent fashions in Rhizobium etli. J. Bacteriol. 2015, 197, 1573–1581. [Google Scholar] [CrossRef] [Green Version]
- Danino, V.E.; Wilkinson, A.; Edwards, A.; Downie, J.A. Recipient-induced transfer of the symbiotic plasmid pRL1JI in Rhizobium leguminosarum bv. viciae is regulated by a quorum-sensing relay. Mol. Microbiol. 2003, 50, 511–525. [Google Scholar] [PubMed] [Green Version]
- Zhang, L.H.; Kerr, A. A diffusible compound can enhance conjugal transfer of the Ti plasmid in Agrobacterium tumefaciens. J. Bacteriol. 1991, 173, 1867–1872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanca-Ordóñez, H.; Oliva-García, J.J.; Pérez-Mendoza, D.; Soto, M.J.; Olivares, J.; Sanjuán, J.; Nogales, J. pSymA-dependent mobilization of the Sinorhizobium meliloti pSymB megaplasmid. J. Bacteriol. 2010, 192, 6309–6312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pistorio, M.; Giusti, A.; Papa, F.D.; Draghi, W.O.; Lozano, M.J.; Tejerizo, T.; Lagares, A. Conjugal properties of the Sinorhizobium meliloti plasmid mobilome. FEMS Microbiol. Ecol. 2008, 65, 372–382. [Google Scholar] [CrossRef] [PubMed]
- Nogales, J.; Blanca-Ordóñez, H.; Olivares, J.; Sanjuán, J. Conjugal transfer of the Sinorhizobium meliloti 1021 symbiotic plasmid is governed through the concerted action of one- and two-component signal transduction regulators. Environ. Microbiol. 2013, 15, 811–821. [Google Scholar] [CrossRef]
- Pérez-Mendoza, D.; Sepúlveda, E.; Pando, V.; Munoz, S.; Nogales, J.; Olivares, J.; Soto, M.J.; Herrera-Cervera, J.A.; Romero, D.; Brom, S. Identification of the rctA gene, which is required for repression of conjugative transfer of rhizobial symbiotic megaplasmids. J. Bacteriol. 2005, 187, 7341–7350. [Google Scholar] [CrossRef] [Green Version]
- Wozniak, R.A.F.; Waldor, M.K. Integrative and conjugative elements: Mosaic mobile genetic elements enabling dynamic lateral gene flow. Nat. Rev. Microbiol. 2010, 8, 552–563. [Google Scholar] [CrossRef] [PubMed]
- Ling, J.; Wang, H.; Wu, P.; Li, T.; Tang, Y.; Naseer, N.; Zheng, H.; Masson-Boivin, C. Plant nodulation inducers enhance horizontal gene transfer of Azorhizobium caulinodans symbiosis island. Proc. Natl. Acad. Sci. USA 2016, 113, 13875–13880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassani, M.A.; Durán, P.; Hacquard, S. Microbial interactions within the plant holobiont. Microbiome 2018, 6, 58. [Google Scholar] [CrossRef]
- Duran, P.; Thiergart, T.; Garrido-Oter, R.; Agler, M.; Kemen, E.; Schulze-Lefert, P.; Hacquard, S. Microbial interkingdom interactions in roots promote Arabidopsis survival. Cell 2018, 175, 973–983. [Google Scholar] [CrossRef] [PubMed]
- Checcucci, A.; Marchetti, M. The Rhizosphere Talk Show: The Rhizobia on Stage. Front. Agron. 2020, 2, 591494. [Google Scholar] [CrossRef]
- Bonfante, P.; Genre, A. Mechanisms underlying beneficial plant—Fungus interactions in mycorrhizal symbiosis. Nat. Commun. 2010, 1, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozsoki, Z.; Cheng, J.; Feng, F.; Gysel, K.; Vinther, M.; Andersen, K.R.; Oldroyd, G.; Blaise, M.; Radutoiu, S.; Stougaard, J. Receptor-mediated chitin perception in legume roots is functionally separable from Nod factor perception. Proc. Natl. Acad. Sci. USA 2017, 114, E8118–E8127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antunes, P.M.; Rajcan, I.; Goss, M.J. Specific flavonoids as interconnecting signals in the tripartite symbiosis formed by arbuscular mycorrhizal fungi, Bradyrhizobium japonicum (Kirchner) Jordan and soybean (Glycine max (L.) Merr.). Soil Biol. Biochem. 2006, 38, 533–543. [Google Scholar] [CrossRef] [Green Version]
- Afkhami, M.E.; Friesen, M.L.; Stinchcombe, J.R. Multiple Mutualism Effects generate synergistic selection and strengthen fitness alignment in the interaction between legumes, rhizobia and mycorrhizal fungi. Ecol. Lett. 2021, 24, 1824–1834. [Google Scholar] [CrossRef]
- Kaschuk, G.; Leffelaar, P.A.; Giller, K.E.; Alberton, O.; Hungria, M.; Kuyper, T.W. Responses of legumes to rhizobia and arbuscular mycorrhizal fungi: A meta-analysis of potential photosynthate limitation of symbioses. Soil Biol. Biochem. 2010, 42, 125–127. [Google Scholar] [CrossRef]
- Horiuchi, J.I.; Prithiviraj, B.; Bais, H.P.; Kimball, B.A.; Vivanco, J.M. Soil nematodes mediate positive interactions between legume plants and rhizobium bacteria. Planta 2005, 222, 848–857. [Google Scholar] [CrossRef] [PubMed]
- Bellabarba, A.; Fagorzi, C.; George, C.; Pini, F.; Viti, C.; Checcucci, A. Deciphering the Symbiotic Plant Microbiome: Translating the Most Recent Discoveries on Rhizobia for the Improvement of Agricultural Practices in Metal-Contaminated and High Saline Lands. Agronomy 2019, 9, 529. [Google Scholar] [CrossRef] [Green Version]
- Schulte, C.C.M.; Ramachandran, V.K.; Papachristodoulou, A.; Poole, P.S. Genome-scale metabolic modelling of lifestyle changes in Rhizobium leguminosarum. bioRxiv 2021, 85, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Schulte, C.C.M.; Borah, K.; Wheatley, R.M.; Terpolilli, J.J.; Saalbach, G.; Crang, N.; Groot, D.H.D.; Ratcliffe, R.G.; Kruger, N.J.; Papachristodoulou, A.; et al. Metabolic control of nitrogen fixation in rhizobium-legume symbioses. Sci. Adv. 2021, 7, eabh2433. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.; Stewart, J.D.; Porter, S.S.; Weedon, J.T.; Kiers, E.T. The evolution of manipulative microbial behaviors in the rhizosphere. Evol. Appl. 2021, 1–16. [Google Scholar] [CrossRef]
- Bevivino, A.; Bacci, G.; Drevinek, P.; Nelson, M.T.; Hoffman, L.; Mengoni, A. Deciphering the Ecology of Cystic Fibrosis Bacterial Communities: Towards Systems-Level Integration. Trends Mol. Med. 2019, 25, 1110–1122. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cangioli, L.; Vaccaro, F.; Fini, M.; Mengoni, A.; Fagorzi, C. Scent of a Symbiont: The Personalized Genetic Relationships of Rhizobium—Plant Interaction. Int. J. Mol. Sci. 2022, 23, 3358. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23063358
Cangioli L, Vaccaro F, Fini M, Mengoni A, Fagorzi C. Scent of a Symbiont: The Personalized Genetic Relationships of Rhizobium—Plant Interaction. International Journal of Molecular Sciences. 2022; 23(6):3358. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23063358
Chicago/Turabian StyleCangioli, Lisa, Francesca Vaccaro, Margherita Fini, Alessio Mengoni, and Camilla Fagorzi. 2022. "Scent of a Symbiont: The Personalized Genetic Relationships of Rhizobium—Plant Interaction" International Journal of Molecular Sciences 23, no. 6: 3358. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23063358