
Treatment with Angiotensin-(1-7) Prevents Development of Oral Papilloma Induced in K-ras Transgenic Mice

 , ,
, , 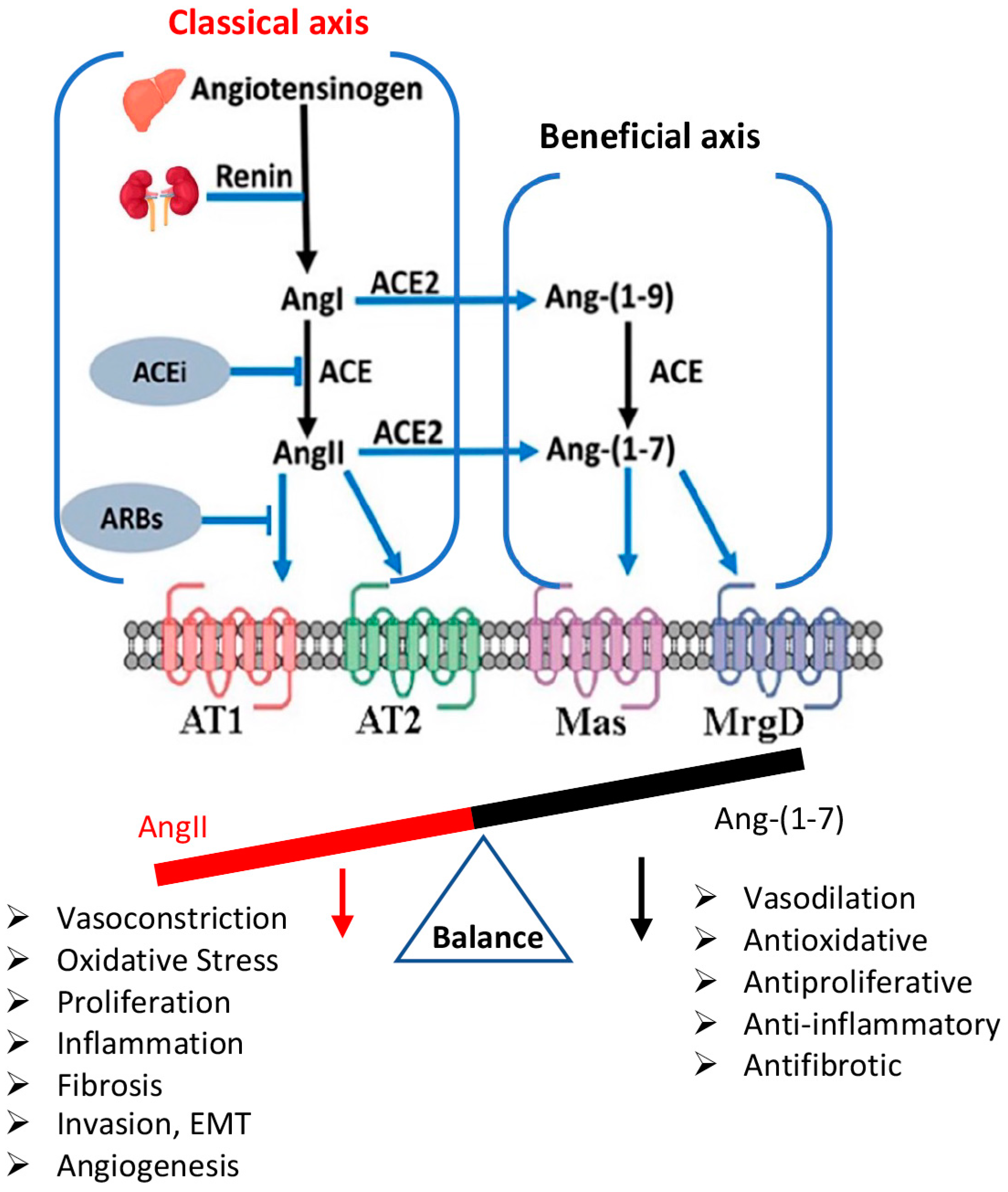{kind=link}
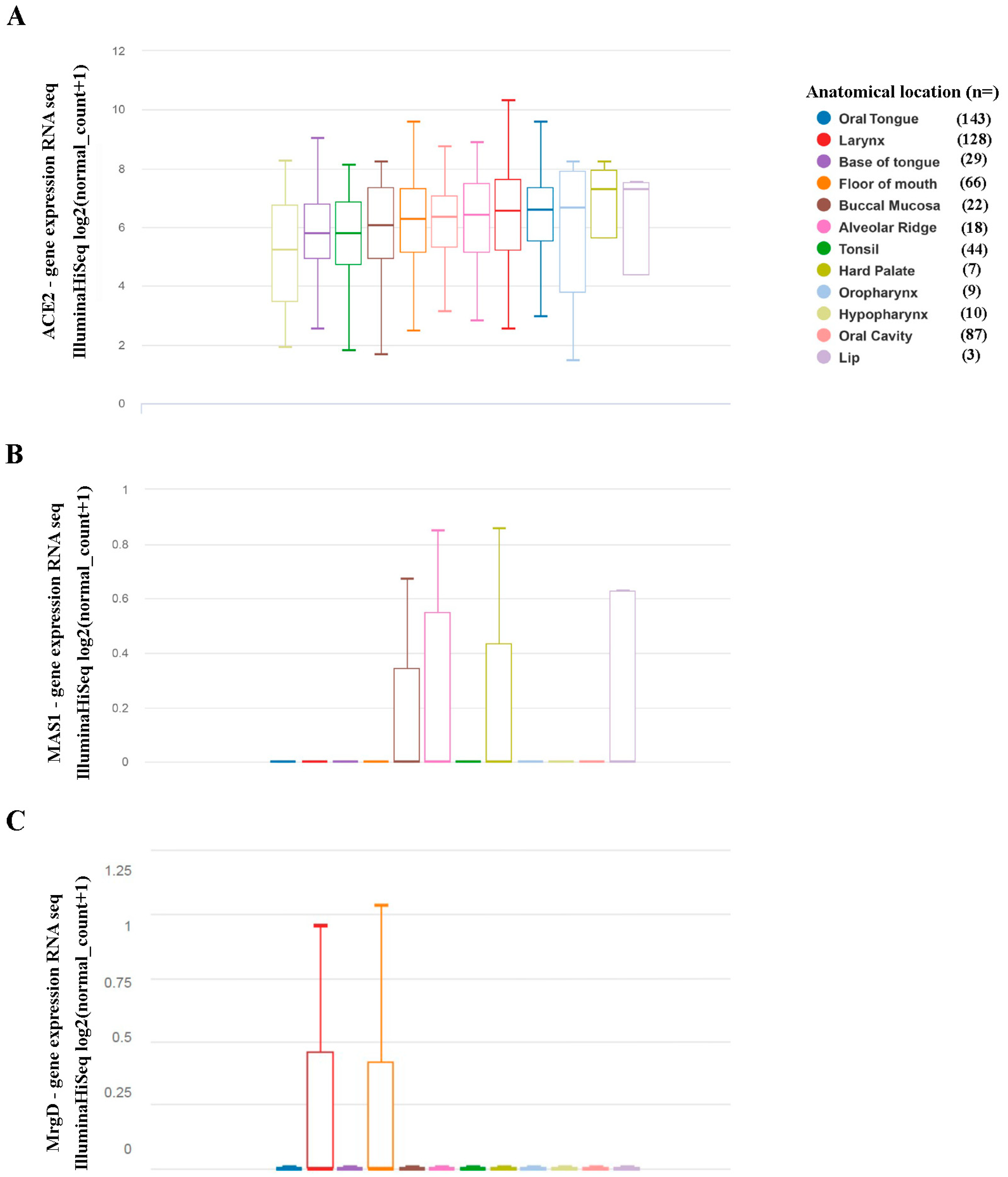{kind=link}
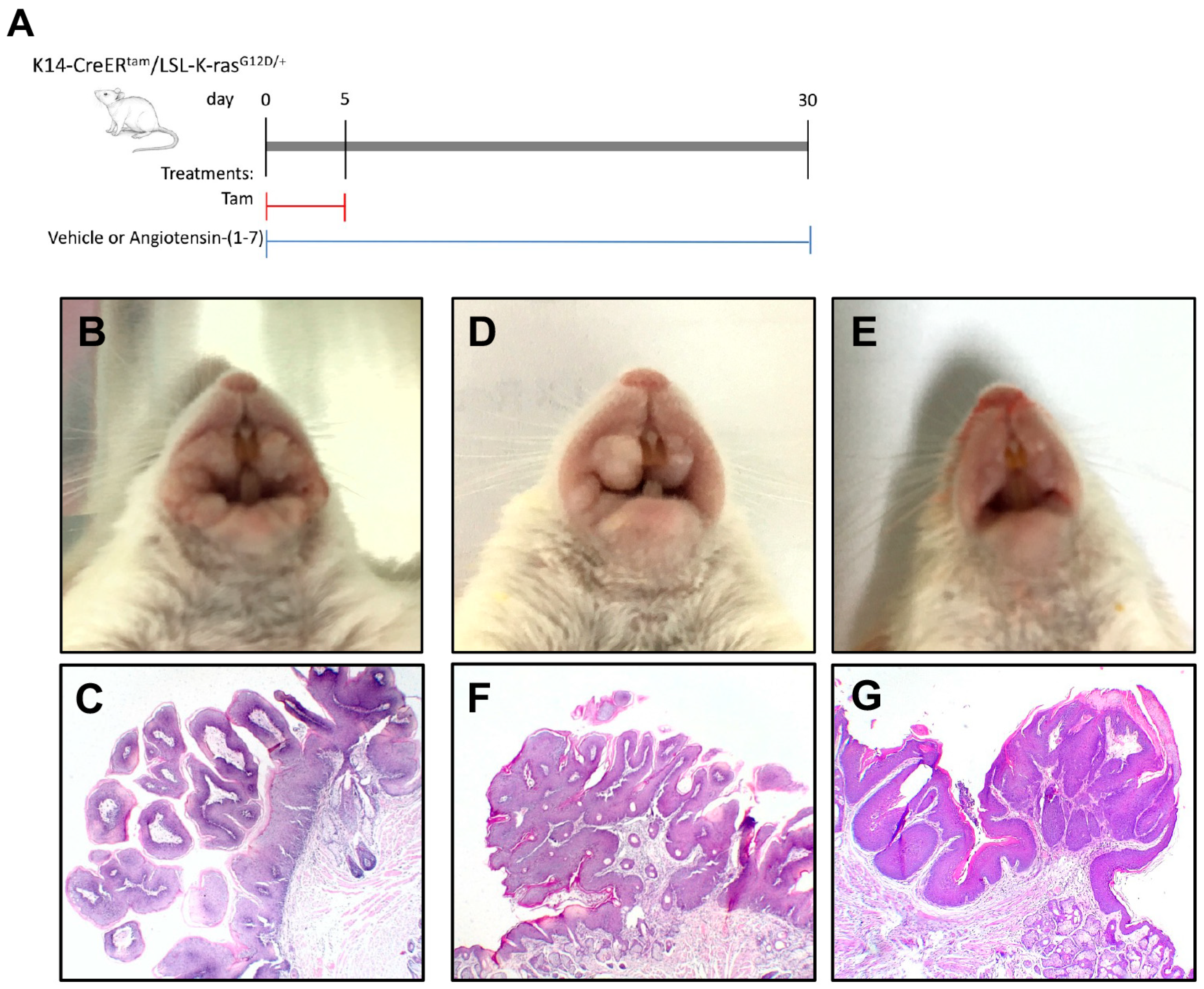{kind=link}
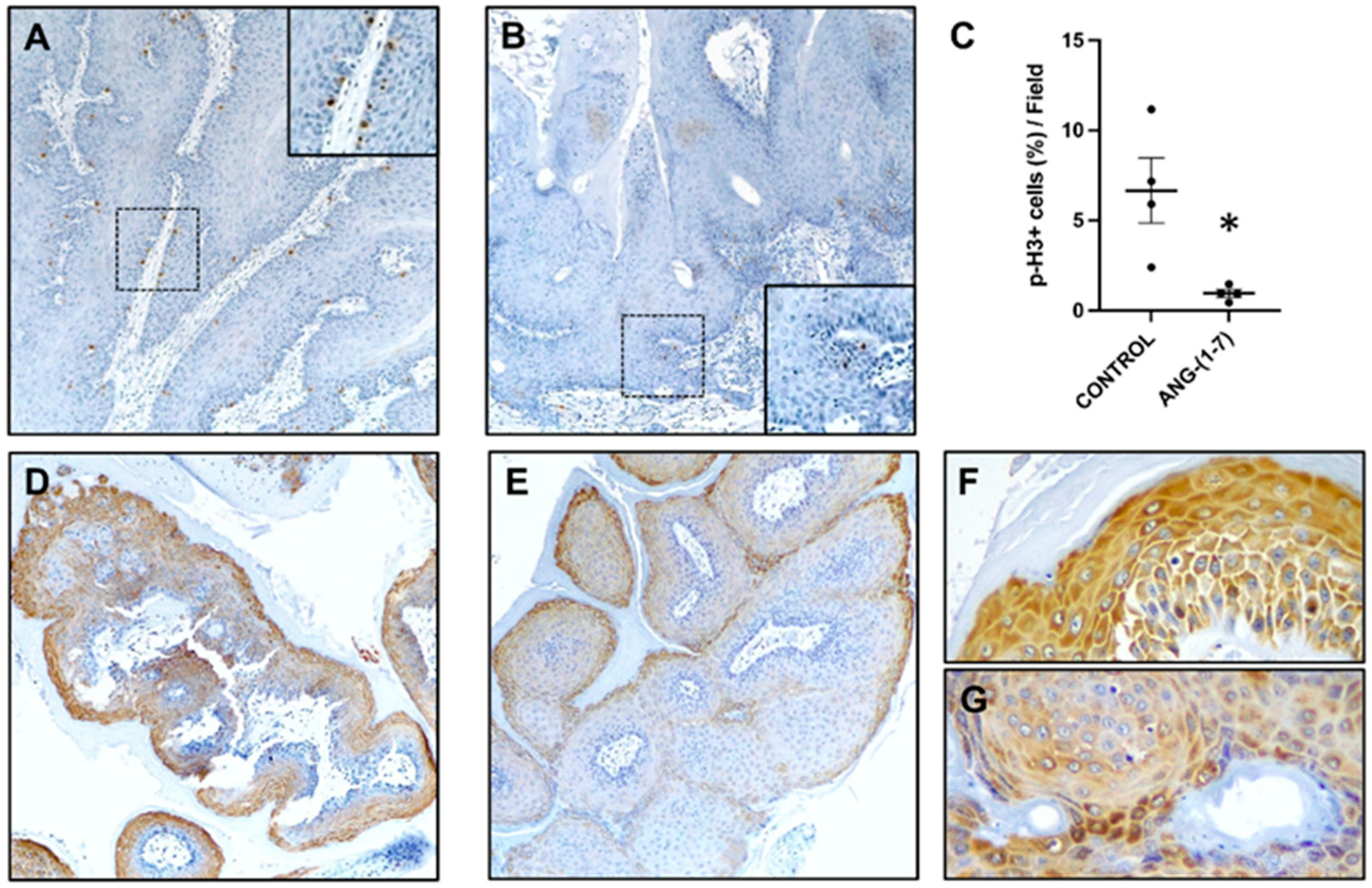{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Expression of Components of The Renin-Angiotensin System in Head and Neck Squamous Cell Carcinoma Originated in Distinct Head and Neck Sites
2.2. Angiotensin-(1-7) Prevents Growth of Oral Lesions Induced by K-ras Activation in K14-CreERtam/LSL-K-rasG12D/+ Mice
2.3. Angiotensin-(1-7) Prevents Cell Proliferation and Activation of the mTOR Pathway
3. Discussion
4. Materials and Methods
4.1. UCSC Xena Cancer Genomics Browser Analysis
4.2. Transgenic Mice
4.3. Immunohistochemistry (IHC) Staining
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- George, A.J.; Thomas, W.G.; Hannan, R.D. The renin-angiotensin system and cancer: Old dog, new tricks. Nat. Rev. Cancer 2010, 10, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Greco, S.; Muscella, A.; Elia, M.; Salvatore, P.; Storelli, C.; Mazzotta, A.; Manca, C.; Marsigliante, S. Angiotensin II activates extracellular signal regulated kinases via protein kinase C and epidermal growth factor receptor in breast cancer cells. J. Cell Physiol. 2003, 196, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Walther, T.; Menrad, A.; Orzechowski, H.-D.; Siemeister, G.; Paul, M.; Schirner, M. Differential regulation of in vivo angiogenesis by angiotensin II receptors. FASEB J. 2003, 17, 2061–2067. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Tai, Z.; Chen, Z.; Zhu, Q.; Bao, L. Clinical applicability of renin-angiotensin system inhibitors in cancer treatment. Am. J. Cancer Res. 2021, 11, 318–336. [Google Scholar]
- Paul, M.; Poyan Mehr, A.; Kreutz, R. Physiology of local renin-angiotensin systems. Physiol. Rev. 2006, 86, 747–803. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef]
- Santos, R.A.S.; e Silva, A.C.S.; Maric, C.; Silva, D.M.R.; Machado, R.P.; de Buhr, I.; Heringer-Walther, S.; Pinheiro, S.V.B.; Lopes, M.T.; Bader, M.; et al. Angiotensin-(1-7) is an endogenous ligand for the G protein-coupled receptor Mas. Proc. Natl. Acad. Sci. USA 2003, 100, 8258–8263. [Google Scholar] [CrossRef] [Green Version]
- Mercure, C.; Yogi, A.; Callera, G.E.; Aranha, A.B.; Bader, M.; Ferreira, A.J.; Santos, A.S.; Walther, T.; Touyz, R.M.; Reudelhuber, T.L. Angiotensin(1–7) blunts hypertensive cardiac remodeling by a direct effect on the heart. Circ. Res. 2008, 103, 1319–1326. [Google Scholar] [CrossRef] [Green Version]
- Cambados, N.; Walther, T.; Nahmod, K.; Tocci, J.M.; Rubinstein, N.; Böhme, I.; Simian, M.; Sampayo, R.; Suberbordes, M.D.V.; Kordon, E.C.; et al. Angiotensin-(1-7) counteracts the transforming effects triggered by angiotensin II in breast cancer cells. Oncotarget 2017, 8, 88475–88487. [Google Scholar] [CrossRef] [Green Version]
- Tetzner, A.; Gebolys, K.; Meinert, C.; Klein, S.; Uhlich, A.; Trebicka, J.; Villacañas, O.; Walther, T. G-Protein-Coupled Receptor MrgD Is a Receptor for Angiotensin-(1-7) Involving Adenylyl Cyclase, cAMP, and Phosphokinase, A. Hypertension 2016, 68, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Menon, J.; Soto-Pantoja, D.R.; Callahan, M.F.; Cline, J.M.; Ferrario, C.M.; Tallant, E.A. Angiotensin-(1-7) inhibits growth of human lung adenocarcinoma xenografts in nude mice through a reduction in cyclooxygenase-2. Cancer Res. 2007, 67, 2809–2815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.T.; Wang, H.C.; Chuang, H.C.; Hsu, Y.C.; Yang, M.Y.; Chien, C.Y. Pre-treatment with Angiotensin-(1-7) inhibits tumor growth via autophagy by downregulating PI3K/Akt/mTOR signaling in human nasopharyngeal carcinoma xenografts. J. Mol. Med. 2018, 96, 1407–1418. [Google Scholar] [CrossRef] [PubMed]
- Khanna, P.; Soh, H.J.; Chen, C.H.; Saxena, R.; Amin, S.; Naughton, M.; Joslin, P.N.; Moore, A.; Bakouny, Z.; Callaghan, C.O.; et al. ACE2 abrogates tumor resistance to VEGFR inhibitors suggesting Angiotensin-(1-7) as a therapy for clear cell renal cell carcinoma. Sci. Transl. Med. 2021, 20, 13. [Google Scholar] [CrossRef] [PubMed]
- Walther, T.; Khanna, P.; Bhatt, R.S. Combining VEGF receptor inhibitors and Angiotensin-(1-7) to target renal cell carcinoma. Mol. Cell Oncol. 2021, 8, 1918529. [Google Scholar] [CrossRef]
- Chai, A.W.Y.; Lim, K.P.; Cheong, S.C. Translational genomics and recent advances in oral squamous cell carcinoma. Semin. Cancer Biol. 2020, 61, 71–83. [Google Scholar] [CrossRef]
- Sasahira, T.; Kirita, T. Hallmarks of cancer related newly prognostic factors of oral squamous cell carcinoma. Int. J. Mol. Sci. 2018, 19, 2413. [Google Scholar] [CrossRef] [Green Version]
- Gutkind, J.S.; Molinolo, A.A.; Wu, X.; Wang, Z.; Nachmanson, D.; Harismendy, O.; Alexandrov, L.B.; Wuertz, B.R.; Ondrey, F.G.; Laronde, D.; et al. Inhibition of mTOR signaling and clinical activity of metformin in oral premalignant lesions. JCI Insight. 2021, 6, e147096. [Google Scholar] [CrossRef]
- Ni, L.; Feng, Y.; Wan, H.; Ma, Q.; Fan, L.; Qian, Y.; Li, Q.; Xiang, Y.; Gao, B. Angiotensin-(1-7) inhibits the migration and invasion of A549 human lung adenocarcinoma cells through inactivation of the PI3K/Akt and MAPK signaling pathways. Oncol. Rep. 2012, 27, 783–790. [Google Scholar]
- Pei, N.; Wan, R.; Chen, X.; Li, A.; Zhang, Y.; Li, J.; Du, H.; Chen, B.; Wei, W.; Qi, Y.; et al. Angiotensin-(1-7) Decreases Cell Growth and Angiogenesis of Human Nasopharyngeal Carcinoma Xenografts. Mol. Cancer Ther. 2016, 15, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Matsushima-Otsuka, S.; Fujiwara-Tani, R.; Sasaki, T.; Ohmori, H.; Nakashima, C.; Kishi, S.; Nishiguchi, Y.; Fujii, K.; Luo, Y.; Kuniyasu, H. Significance of intranuclear angiotensin-II type 2 receptor in oral squamous cell carcinoma. Oncotarget 2018, 9, 36561–36574. [Google Scholar] [CrossRef] [Green Version]
- Hinsley, E.E.; De Oliveira, C.E.; Hunt, S.; Coletta, R.D.; Lambert, D.W. Angiotensin 1–7 inhibits angiotensin II-stimulated head and neck cáncer progression. Eur. J. Oral. Sci. 2017, 125, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Raimondi, A.R.; Molinolo, A.; Gutkind, J.S. Rapamycin prevents early onset of tumorigenesis in an oral-specific K-ras and p53 two-hit carcinogenesis model. Cancer Res. 2009, 69, 4159–4166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itinteang, T.; Dunne, J.C.; Chibnall, A.M.; Brasch, H.D.; Davis, P.F.; Tan, S.T. Cancer stem cells in moderately differentiated oral tongue squamous cell carcinoma express components of the renin-angiotensin system. J. Clin. Pathol. 2016, 69, 942–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrigo, M.; Alvarez, R.; Paparella, M.L.; Calb, D.E.; Bal de Kier Joffe, E.; Gutkind, S.; Raimondi, A.R. Impairing squamous differentiation by Klf4 deletion is sufficient to initiate tongue carcinoma development upon K-Ras activation in mice. Carcinogenesis 2014, 35, 662–669. [Google Scholar] [CrossRef] [Green Version]
- Siljee, S.; Pilkington, T.; Brasch, H.D.; Bockett, N.; Patel, J.; Paterson, E.; Davis, P.F.; Tan, S.T. Cancer Stem Cells in Head and Neck Metastatic Malignant Melanoma Express Components of the Renin-Angiotensin System. Life 2020, 10, 268. [Google Scholar] [CrossRef]
- Alenina, N.; Bader, M.; Walther, T. Imprinting of the murine MAS protooncogene is restricted to its antisense RNA. Biochem. Biophys. Res. Commun. 2002, 290, 1072–1078. [Google Scholar] [CrossRef]
- Supé, S.; Kohse, F.; Gembardt, F.; Kuebler, W.M.; Walther, T. Therapeutic time window for Angiotensin-(1-7) in acute lung injury. Br. J. Pharmacol. 2016, 173, 1618–1628. [Google Scholar] [CrossRef]
- Krishnan, B.; Torti, F.M.; Gallagher, P.E.; Tallant, E.A. Angiotensin-(1-7) reduces proliferation and angiogenesis of human prostate cancer xenografts with a decrease in angiogenic factors and an increase in sFlt-1. Prostate 2013, 73, 60–70. [Google Scholar] [CrossRef]
- Cook, K.L.; Metheny-Barlow, L.J.; Tallant, E.A.; Gallagher, P.E. Angiotensin-(1-7) reduces fibrosis in orthotopic breast tumors. Cancer Res. 2010, 70, 8319–8328. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Tanabe, E.; Kitayoshi, M.; Nishiguchi, Y.; Fujiwara, R.; Matsushima, S.; Sasaki, T.; Sasahira, T.; Chihara, Y.; Nakae, D.; et al. Expression of MAS1 in breast cancer. Cancer Sci. 2015, 106, 1240–1248. [Google Scholar] [CrossRef]
- Liu, Y.; Li, B.; Wang, X.; Li, G.; Shang, R.; Yang, J.; Wang, J.; Zhang, M.; Chen, Y.; Zhang, Y.; et al. Angiotensin-(1-7) suppresses hepatocellular carcinoma growth and angiogenesis via complex interactions of angiotensin II type 1 receptor, angiotensin II type 2 receptor and Mas receptor. Mol. Med. 2015, 21, 626–636. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Wu, S.; Chen, W.; Yang, Y.; Chien, C.; Fang, F.; Li, S.; Luo, S.; Chiu, T. Angiotensin II receptor blockers and oral squamous cell carcinoma survival: A propensity-score-matched cohort study. PLoS ONE 2021, 16, e0260772. [Google Scholar] [CrossRef] [PubMed]
- Day, T.A.; Shirai, K.; O’Brien, P.E.; Matheus, M.G.; Godwin, K.; Sood, A.J.; Kompelli, A.; Vick, J.A.; Martin, D.; Vitale-Cross, L.; et al. Inhibition of mTOR signaling and clinical activity of Rapamycin in head and neck cancer in a window of opportunity trial. Clin. Cancer Res. 2019, 25, 1156–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, L.K.; Totou, N.; Moura, S.; Kangussu, L.; Millán, R.D.S.; Campagnole-Santos, M.J.; Santos, R.A.S. Eccentric Overload Muscle Damage is Attenuated By a Novel Angiotensin-(1-7) Treatment. Int. J. Sports Med. 2018, 39, 743–748. [Google Scholar] [CrossRef]
- Hemnes, A.R.; Rathinasabapathy, A.; Austin, E.A.; Brittain, E.L.; Carrier, E.J.; Chen, X.; Fessel, J.P.; Fike, C.D.; Fong, P.; Fortune, N.; et al. A potential therapeutic role for angiotensin-converting enzyme 2 in human pulmonary arterial hypertension. Eur. Respir. J. 2018, 51, 1702638. [Google Scholar] [CrossRef]
- Raimondi, A.R.; Vitale-Cross, L.; Amornphimoltham, P.; Gutkind, J.S.; Molinolo, A. Rapid development of salivary gland carcinomas upon conditional expression of K-ras driven by the cytokeratin 5 promoter. Am. J. Pathol. 2006, 168, 1654–1665. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schere-Levy, C.; Suberbordes, M.; Ferri, D.M.; Ayre, M.; Gattelli, A.; Kordon, E.C.; Raimondi, A.R.; Walther, T. Treatment with Angiotensin-(1-7) Prevents Development of Oral Papilloma Induced in K-ras Transgenic Mice. Int. J. Mol. Sci. 2022, 23, 3642. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23073642
Schere-Levy C, Suberbordes M, Ferri DM, Ayre M, Gattelli A, Kordon EC, Raimondi AR, Walther T. Treatment with Angiotensin-(1-7) Prevents Development of Oral Papilloma Induced in K-ras Transgenic Mice. International Journal of Molecular Sciences. 2022; 23(7):3642. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23073642
Chicago/Turabian StyleSchere-Levy, Carolina, Melisa Suberbordes, Darío M. Ferri, Marina Ayre, Albana Gattelli, Edith C. Kordon, Ana R. Raimondi, and Thomas Walther. 2022. "Treatment with Angiotensin-(1-7) Prevents Development of Oral Papilloma Induced in K-ras Transgenic Mice" International Journal of Molecular Sciences 23, no. 7: 3642. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23073642