
The Microbial Mechanisms of a Novel Photosensitive Material (Treated Rape Pollen) in Anti-Biofilm Process under Marine Environment


Abstract
:1. Introduction
2. Results
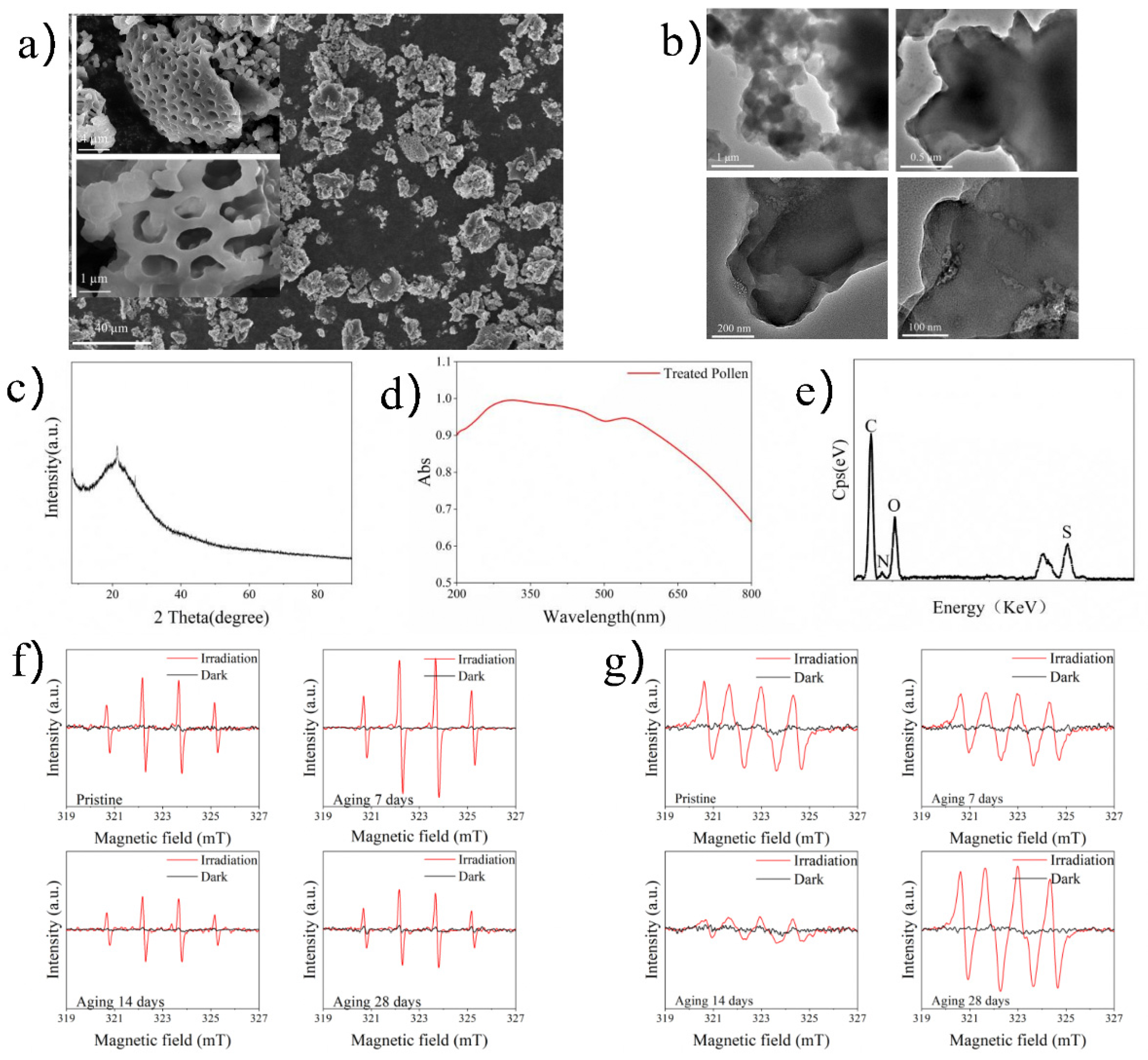
2.1. Characterization of TRP
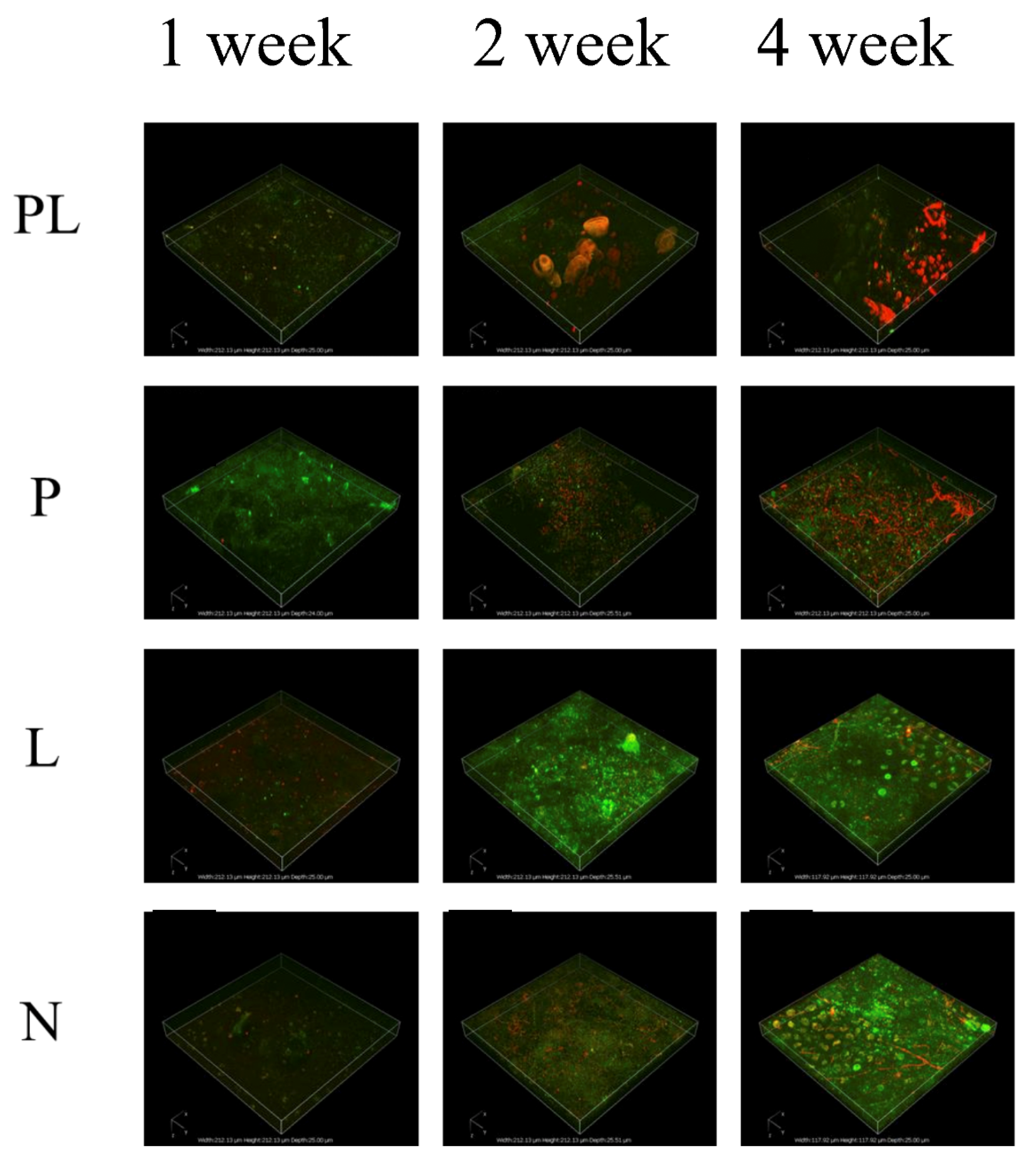
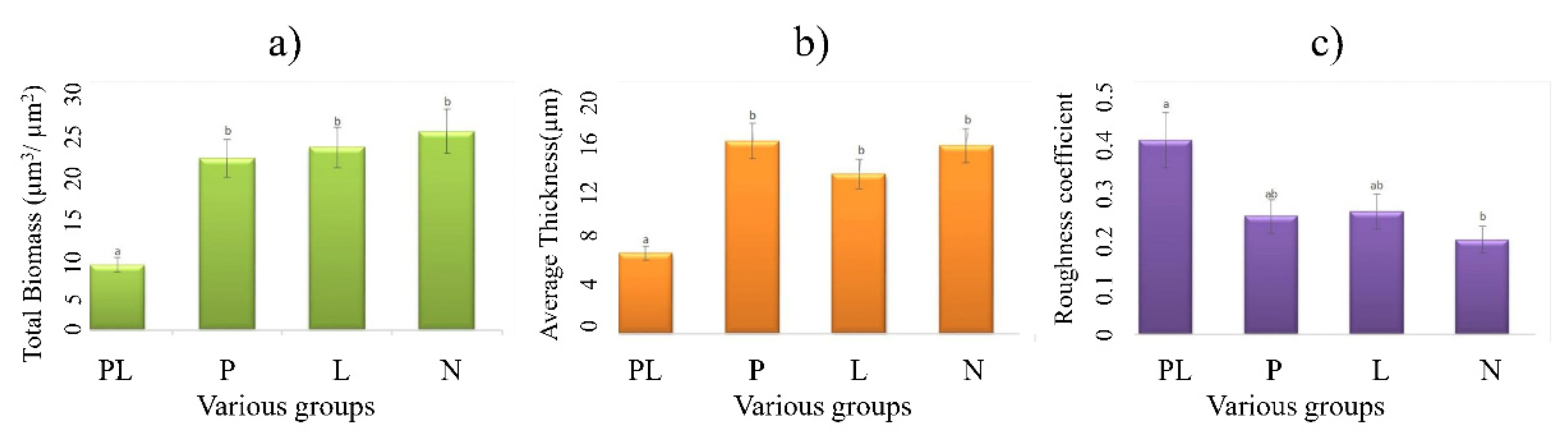
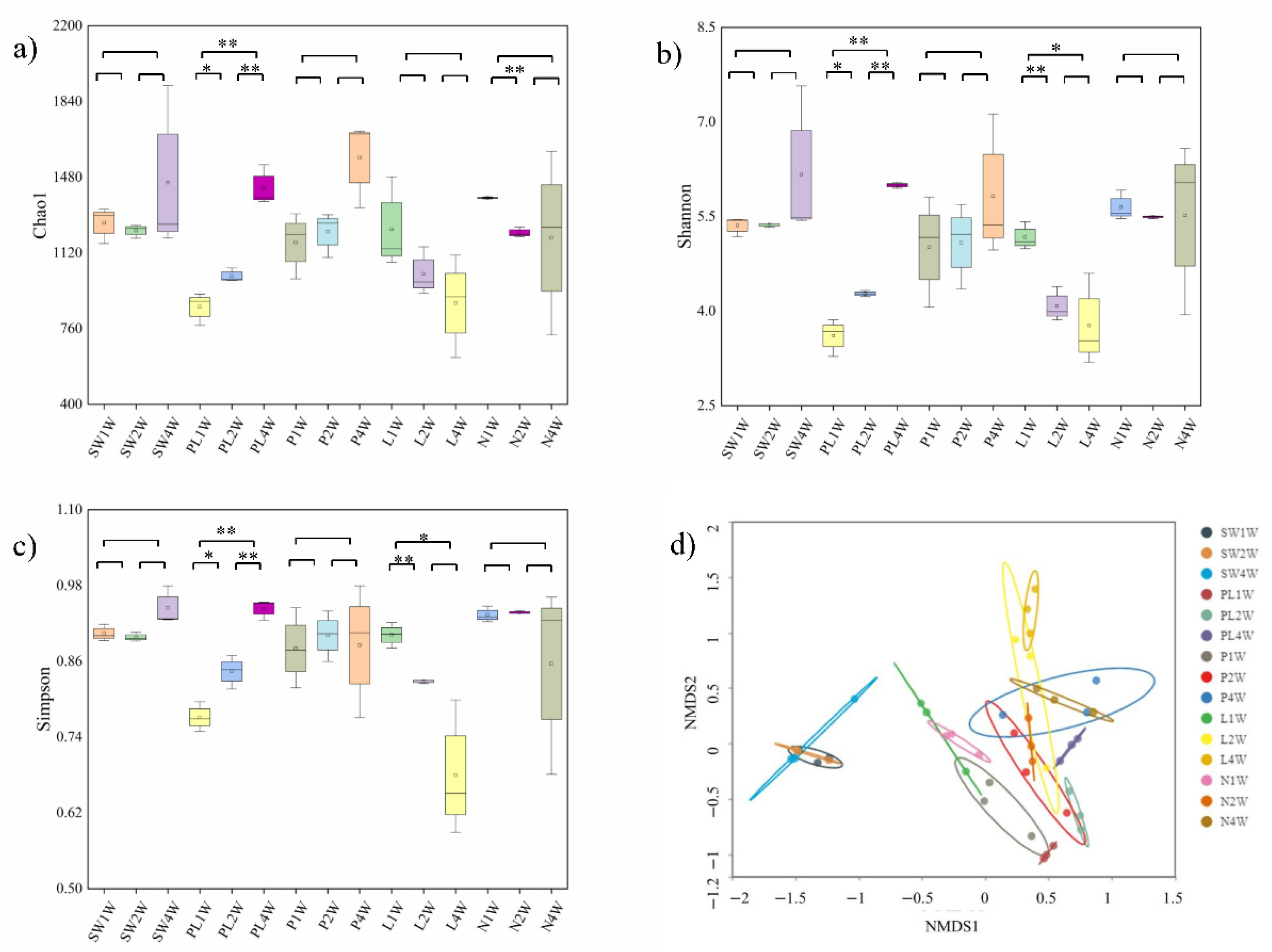
2.2. Effect of TRP on the Biodiversity of Biofilm
2.3. Effect of TRP on the Biodiversity of Biofilm
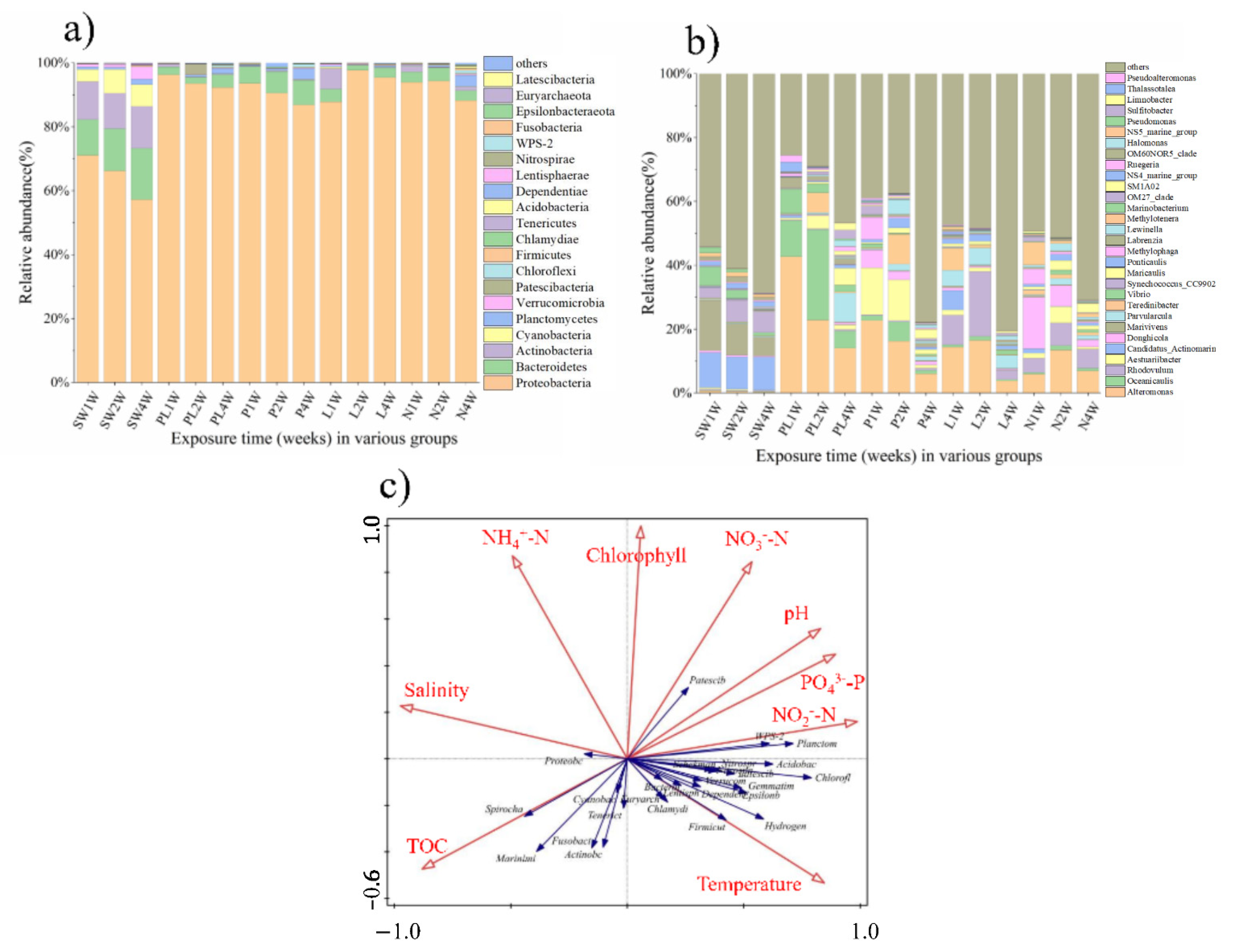
2.4. Taxonomic Composition
2.5. Correlation Analysis of Environmental Factors
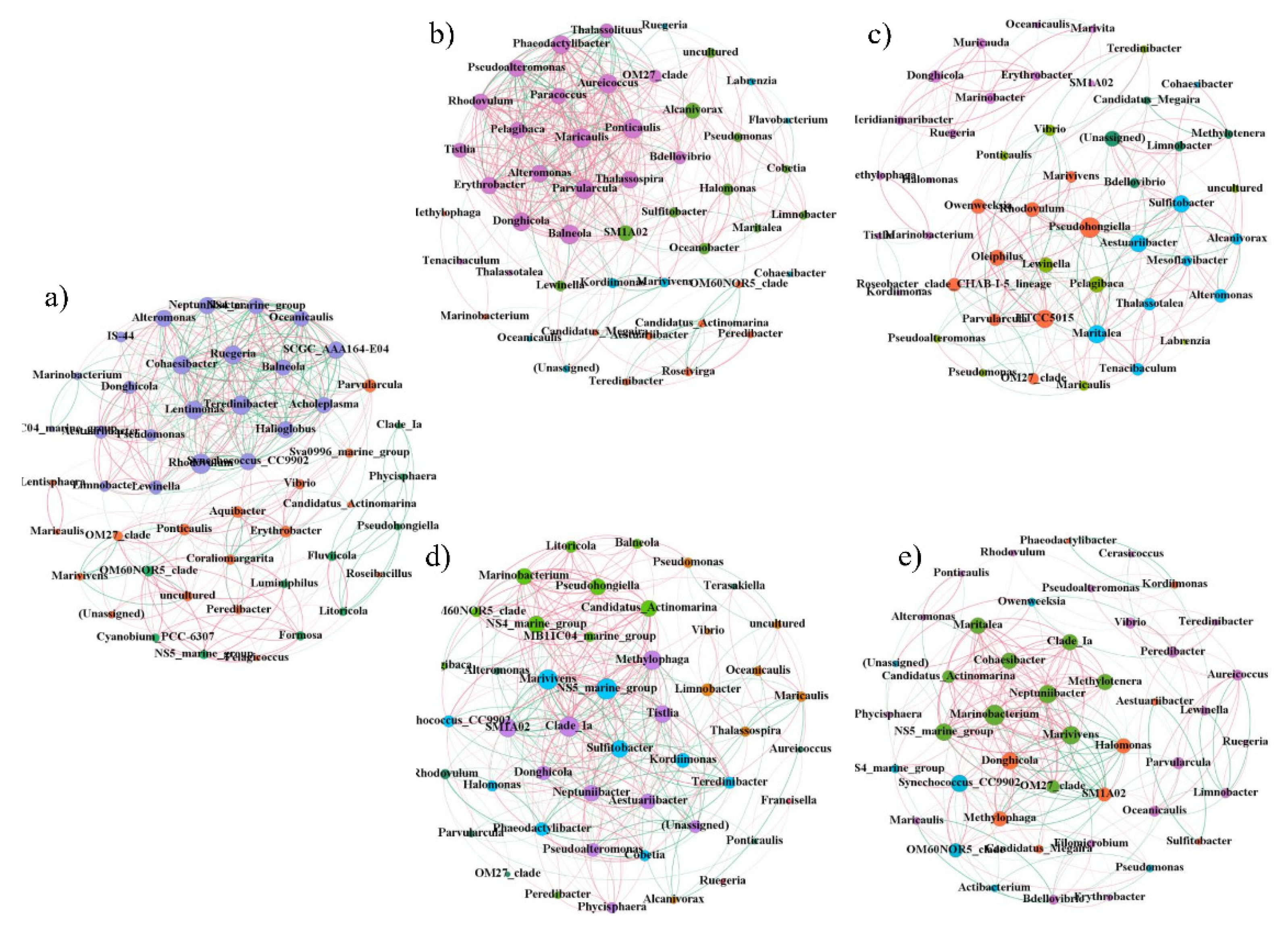
2.6. Network Analysis of Biofilm Microorganisms
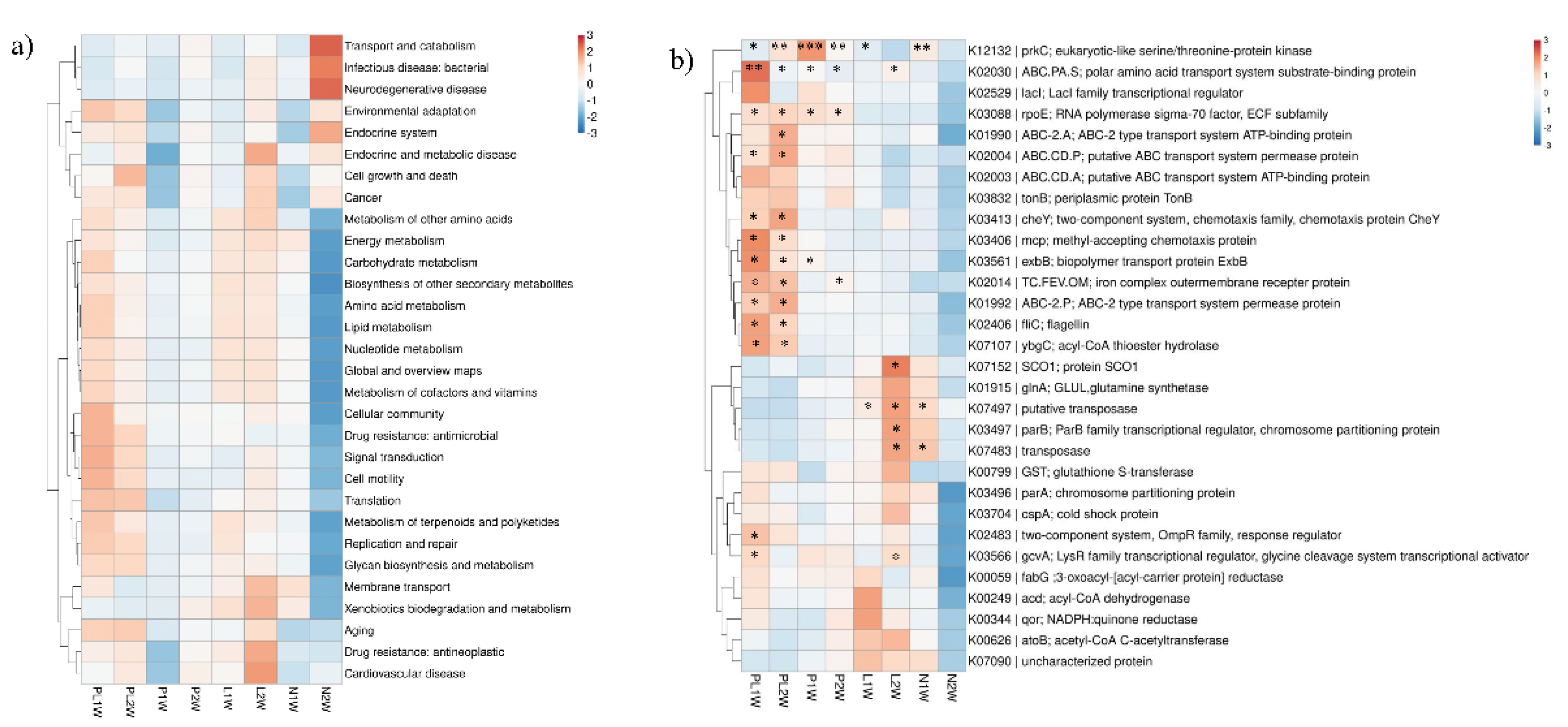
2.7. Biofilm Microbial Functions based on Metagenomic Data
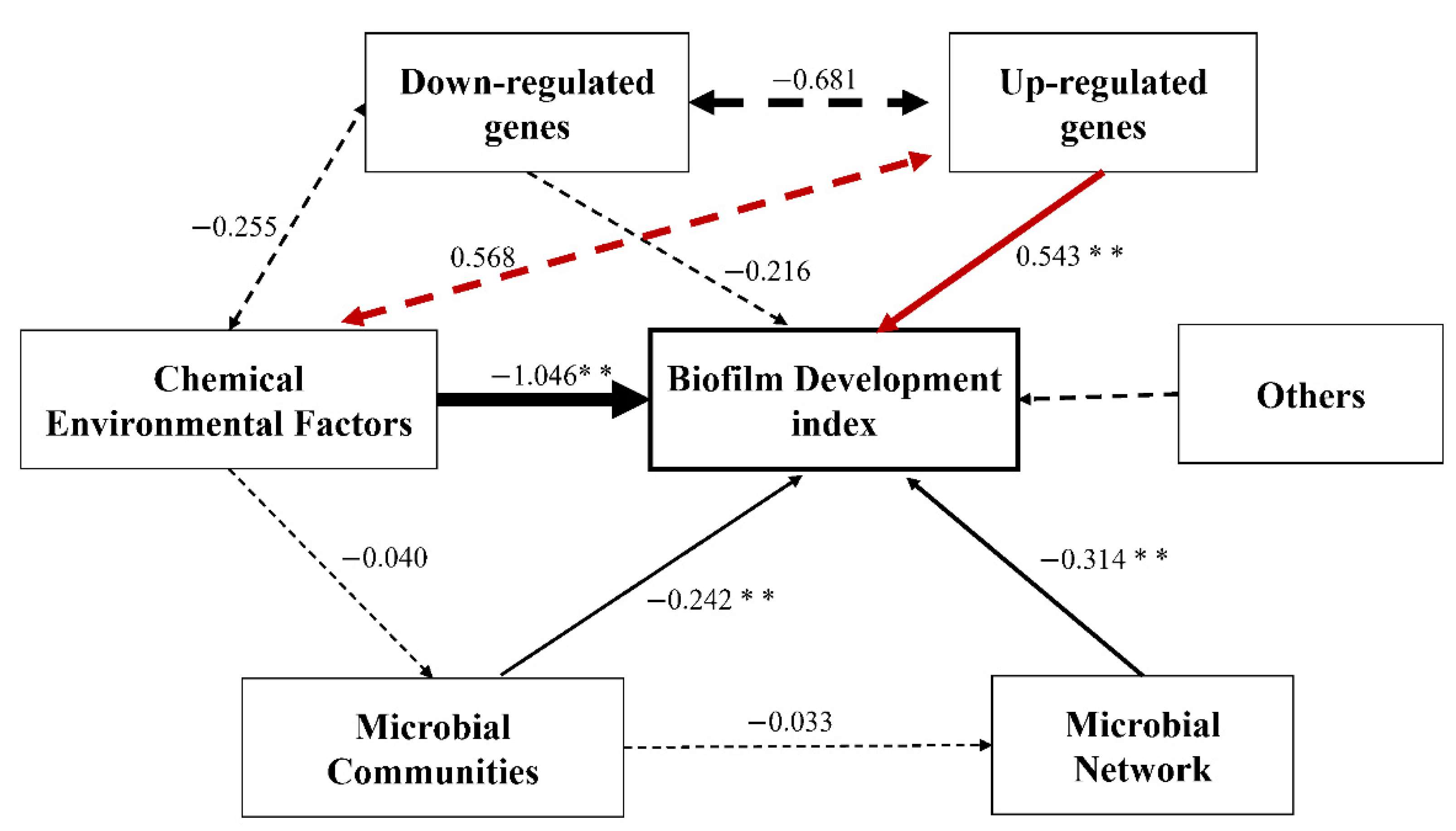
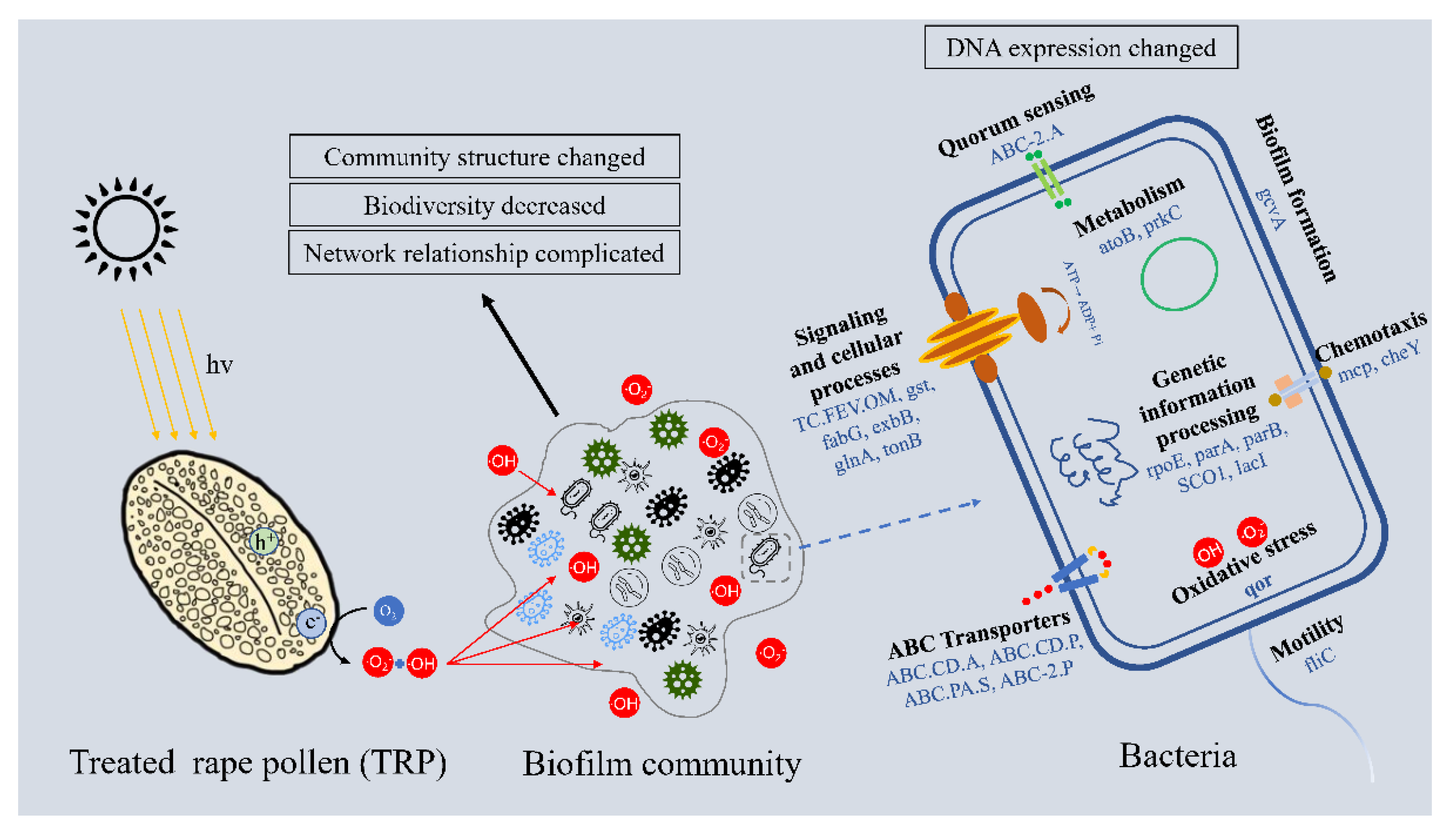
2.8. Linkage of TRP to Biofilm Inhibition
3. Discussion
3.1. Material Characteristics
3.2. Anti-Biofilm Profiles of TRP
3.3. Anti-Biofilm Mechanism at Functional Gene Level
4. Materials and Methods
4.1. Preparation and Characterization of TRP
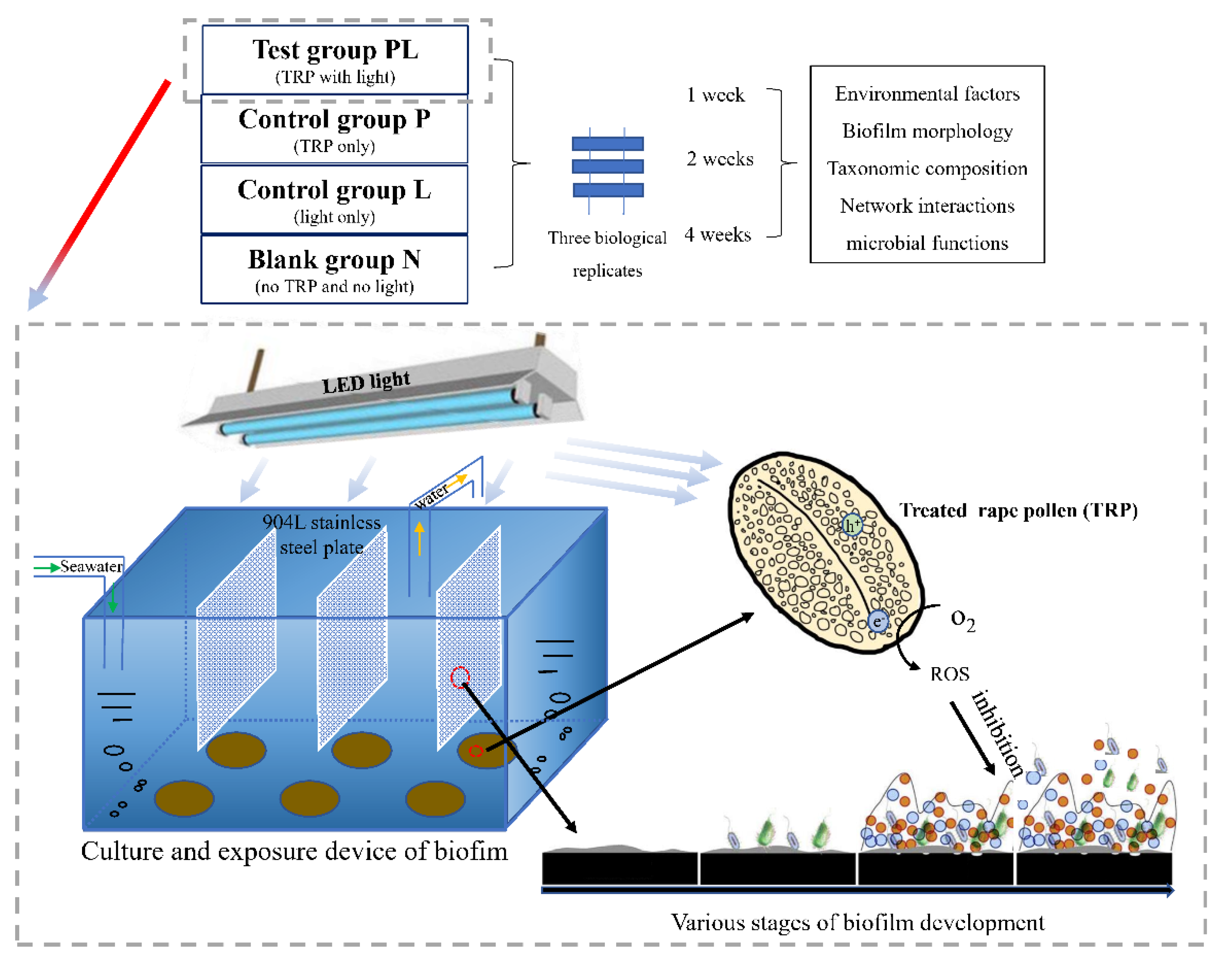
4.2. Experimental Design
4.3. Environmental Parameters
4.4. Biofilm Profiles
4.5. DNA Extraction and Amplification
4.6. 16S rRNA Gene Sequencing and Analyses
4.7. Metagenomic Sequencing and Analyses
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fitridge, I.; Dempster, T.; Guenther, J.; Nys, R.D. The impact and control of biofouling in marine aquaculture: A review. Biofouling 2012, 28, 649–669. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.J.; Chen, A.R.; Pan, Z.C.; Ma, R.J. Two-dimensional numerical model and fast estimation method for calculating crevice corrosion of cross-sea bridges. Constr. Build. Mater. 2019, 206, 683–693. [Google Scholar] [CrossRef]
- Qu, G.; Cai, Y.; Lv, P.; Ma, X.; Xie, R.; Xu, Y.; Ning, P. Effect of EM microbial agent on aerobic composting for dairy cattle manure. Int. J. Environ. Sci. Technol. 2019, 16, 6945–6958. [Google Scholar] [CrossRef]
- Xue, J.H. Asymptotic analysis for buckling of undersea corroded pipelines. J. Press. Vessel Technol. 2008, 130, 021705. [Google Scholar] [CrossRef]
- Ferrario, J.; Caronni, S.; Ambrogi, A.O.; Marchini, A. Role of commercial harbours and recreational marinas in the spread of non-indigenous fouling species. Biofouling 2017, 33, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.X.; Li, Y.N.; Ladewiga, B.P. A review of reverse osmosis membrane fouling and control strategies. Sci. Total Environ. 2017, 595, 567–583. [Google Scholar] [CrossRef]
- Antunes, J.; Leão, P.; Vasconcelos, V. Marine biofilms: Diversity of communities and of chemical cues. Environ. Microbiol. Rep. 2019, 11, 287–305. [Google Scholar] [CrossRef]
- Dobretsov, S.; Rittschof, D. Love at first taste: Induction of larval settlement by marine microbes. Int. J. Mol. Sci. 2020, 21, 731. [Google Scholar] [CrossRef] [Green Version]
- Balasubramanian, S.; Othman, E.M.; Kampik, D.; Stopper, H.; Hentschel, U.; Ziebuhr, W.; Oelschlaeger, T.A.; Abdelmohsen, U.R. Marine sponge-derived Streptomyces sp. SBT343 extract inhibits staphylococcal biofilm formation. Front. Microbiol. 2017, 8, 236. [Google Scholar] [CrossRef] [Green Version]
- Balasubramanian, S.; Skaf, O.; Holzgrabe, U.; Bharti, R.; Förstner, K.U.; Ziebuhr, W.; Humeida, U.H.; Abdelmohsen, U.R.; Oelschlaeger, T.A. A new bioactive compound from the marine sponge-derived Streptomyces sp. SBT348 inhibits staphylococcal growth and biofilm formation. Front. Microbiol. 2018, 9, 1473. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, M.L.; Schott, S.S.; Grigoryan, B.; Rufin, M.A.; Ngo, B.K.D.; Vanderwal, L.; Stafslien, S.J.; Grunlan, M.A. Anti-protein and anti-bacterial behavior of amphiphilic silicones. Polym. Chem-uk. 2017, 8, 5239–5251. [Google Scholar] [CrossRef] [PubMed]
- Peres, R.S.; Armelin, E.; Moreno, J.A.; Aleman, C.; Ferreira, C.A. Transport and antifouling properties of papain-based antifouling coatings. Appl. Surf. Sci. 2015, 341, 75–85. [Google Scholar] [CrossRef]
- Qin, L.G.; Hafezi, M.; Yang, H.; Dong, G.N.; Zhang, Y.L. Constructing a dual-function surface by microcasting and nanospraying for efficient drag reduction and potential antifouling capabilities. Micromachines 2019, 10, 490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rijavec, T.; Zrimec, J.; Spanning, R.V.; Lapanje, A. Natural microbial communities can be manipulated by artificially constructed biofilms. Adv. Sci. 2019, 6, 1901408. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Jiang, X.H.; Yu, L.M.; Xia, S.W.; Yu, X.L. Synthesis and quantum chemical calculation of benzamide derivatives containing capsaicin and their bacteriostatic and antifouling properties. J. Chin. Chem. Soc. 2015, 62, 861–870. [Google Scholar] [CrossRef]
- Castaneda, H.; Benetton, X.D. SRB-biofilm influence in active corrosion sites formed at the steel-electrolyte interface when exposed to artificial seawater conditions. Corros. Sci. 2008, 50, 1169–1183. [Google Scholar] [CrossRef]
- Thomas, K.V.; Brooks, S. The environmental fate and effects of antifouling paint biocides. Biofouling 2010, 26, 73–88. [Google Scholar] [CrossRef]
- Xia, D.; Shen, Z.; Huang, G.; Wang, W.; Yu, J.C.; Wong, P.K. Red phosphorus: An earth-abundant elemental photocatalyst for “green” bacterial inactivation under visible light. Environ. Sci. Technol. 2015, 49, 6264–6273. [Google Scholar] [CrossRef]
- Sun, H.; Jiang, Z.; Wu, D.; Ye, L.; Wang, T.; Wang, B.; An, T.; Wong, P.K. Defect-type-dependent near-infrared-driven photocatalytic bacterial inactivation by defective Bi2S3 nanorods. ChemSusChem 2019, 12, 890–897. [Google Scholar] [CrossRef]
- Hu, Z.; Shen, Z.; Yu, J.C. Converting carbohydrates to carbon-based photocatalysts for environmental treatment. Environ. Sci. Technol. 2017, 51, 7076–7083. [Google Scholar] [CrossRef]
- Jiang, Z.F.; Sun, H.L.; Wang, T.Q.; Wang, B.; Wei, W.; Li, H.M.; Yuan, S.Q.; An, T.C.; Zhao, H.J.; Yu, J.G.; et al. Nature-based catalyst for visible-light-driven photocatalytic CO2 reduction. Energy Environ. Sci. 2018, 11, 2382–2389. [Google Scholar] [CrossRef] [Green Version]
- Xiao, K.M.; Wang, T.Q.; Sun, M.Z.; Hanif, A.; Gu, Q.F.; Tian, B.B.; Jiang, Z.F.; Wang, B.; Sun, H.L.; Shang, J.; et al. Photocatalytic bacterial inactivation by a rape pollen-MoS2 biohybrid catalyst: Synergetic effects and inactivation mechanisms. Environ. Sci. Technol. 2020, 54, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Qiu, B.C.; Zhu, Q.H.; Du, M.M.; Fan, L.G.; Xing, M.Y.; Zhang, J.L. Efficient solar light harvesting CdS/Co9S8 hollow cubes for Z-scheme photocatalytic water splitting. Angew. Chem. Int. Ed. 2017, 56, 2684–2688. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.C.; Heil, T.; Sun, J.K.; Schmidt, B.V.K.J. Dispersed nano-MOFs via a stimuli-responsive biohybrid-system with enhanced photocatalytic performance. Mater. Horiz. 2019, 6, 802–809. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Jiang, Z.F.; Yu, J.C. Treated rape pollen: A metal-free visible-light-driven photocatalyst from nature for efficient water disinfection. J. Mater. Chem. A. 2019, 7, 9335–9344. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.J.; Nuccio, E.E.; Shi, Z.J.; He, Z.L.; Zhou, J.Z.; Firestone, M.K. The interconnected rhizosphere: High network complexity dominates rhizosphere assemblages. Ecol. Lett. 2016, 19, 926–936. [Google Scholar] [CrossRef] [Green Version]
- David, B.; Stefanie, W. Deciphering microbial interactions and detecting keystone species with co-occurrence networks. Front. Microbiol. 2014, 5, 219. [Google Scholar]
- Song, Y.; Zhang, S.J.; Zeng, Y.H.; Zhu, J.M.; Du, X.P.; Cai, Z.H.; Zhou, J. The Rhodamine isothiocyanate analogue as a quorum sensing inhibitor has the potential to control microbially-induced biofouling. Mar. Drugs 2020, 18, 484. [Google Scholar] [CrossRef]
- Packiavathy, I.A.S.; Maruthamuthu, S.; Gnanaselvan, G.; Manoharan, S.; Paul, J.B.J.; Annapoorani, A.; Kannappan, A.; Ravi, A.V. The control of microbially induced corrosion by methyl eugenol—A dietary phytochemical with quorum sensing inhibitory potential. Bioelectrochemistry 2019, 128, 186–192. [Google Scholar] [CrossRef]
- Ding, W.; Zhang, W.P.; Alikunhi, N.M.; Batang, Z.; Pei, B.T.; Wang, R.J.; Chen, L.G.; Al-Suwailem, A.; Qian, P.Y. Metagenomic analysis of zinc surface-associated marine biofilms. Microb. Ecol. 2019, 77, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.L.; Maki, J.S.; Rittschof, D.; Teo, S.L.M. Early marine bacterial biofilm on a copper-based antifouling paint. Int. Biodeterior. Biodegrad. 2013, 83, 71–76. [Google Scholar] [CrossRef]
- Cusick, K.D.; Dale, J.R.; Fitzgerald, L.A.; Little, B.J.; Biffinger, J.C. Adaptation to copper stress influences biofilm formation in Alteromonas macleodii. Biofouling 2017, 33, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Dang, H.Y.; Li, T.G.; Chen, M.N.; Huang, G.Q. Cross-Ocean distribution of Rhodobacterales bacteria as primary surface colonizers in temperate coastal marine waters. Appl. Environ. Microbiol. 2008, 74, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Kesy, K.; Oberbeckmann, S.; Kreikemeyer, B.; Labrenz, M. Spatial environmental heterogeneity determines young biofilm assemblages on microplastics in Baltic Sea mesocosms. Front. Microbiol. 2019, 10, 1665. [Google Scholar] [CrossRef] [Green Version]
- Brislawn, C.J.; Graham, E.B.; Dana, K.; Ihard, P.; Fansler, S.J.; Chrisler, W.B.; Cliff, J.B.; Stegen, J.C.; Moran, J.J.; Bernstein, H.C. Forfeiting the priority effect: Turnover defines biofilm community succession. ISME J. 2019, 13, 1865–1877. [Google Scholar] [CrossRef] [Green Version]
- Polz, M.F.; Cordero, O.X. Bacterial evolution: Genomics of metabolic trade-offs. Nat. Microbiol. 2016, 1, 16181. [Google Scholar] [CrossRef]
- Wu, X.; Pan, J.; Li, M.; Li, Y.; Bartlam, M.; Wang, Y. Selective enrichment of bacterial pathogens by microplastic biofilm. Water Res. 2019, 165, 114979. [Google Scholar] [CrossRef]
- Grosskopf, T.; Soyer, O.S. Microbial diversity arising from thermodynamic constraints. ISME J. 2016, 10, 2725–2733. [Google Scholar] [CrossRef]
- Santolini, M.; Barabasi, A.L. Predicting perturbation patterns from the topology of biological networks. Proc. Natl. Acad. Sci. USA 2018, 115, E6375–E6383. [Google Scholar] [CrossRef] [Green Version]
- Zaquin, T.; Zaslansky, P.; Pinkas, I.; Mass, T. Simulating bleaching: Long-term adaptation to the dark reveals phenotypic plasticity of the mediterranean sea coral oculina patagonica. Front. Mar. Sci. 2019, 6, 662. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.M.; Zhou, J.; Wang, H.; Chen, G.F.; Cai, Z.H. The ecological network approach to algal-bacterial relationships: Review and prospects. Chin. Sci. Bull. 2021, 66, 4378–4394. [Google Scholar] [CrossRef]
- Lobelle, D.; Cunliffe, M. Early microbial biofilm formation on marine plastic debris. Mar. Pollut. Bull. 2011, 62, 197–200. [Google Scholar] [CrossRef] [PubMed]
- Abed, R.M.M.; Fahdi, D.A.; Muthukrishnan, T. Short-term succession of marine microbial fouling communities and the identification of primary and secondary colonizers. Biofouling 2019, 35, 526–540. [Google Scholar] [CrossRef]
- Xu, R.H.; Zhang, S.Q.; Meng, F.G. Large-sized planktonic bioaggregates possess high biofilm formation potentials: Bacterial succession and assembly in the biofilm metacommunity. Water Res. 2020, 170, 115307. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, J.; He, R.; Hu, H.; Wu, B.; Ren, H. Bacterial assembly during the initial adhesion phase in wastewater treatment biofilms. Water Res. 2020, 184, 116147. [Google Scholar] [CrossRef]
- Ross, B.N.; Whiteley, M. Ignoring social distancing: Advances in understanding multi-species bacterial interactions. Fac. Rev. 2020, 9, 23. [Google Scholar] [CrossRef]
- Karimi, B.; Maron, P.A.; Boure, N.C.P.; Bernard, N.; Gilbert, D.; Ranjard, L. Microbial diversity and ecological networks as indicators of environmental quality. Environ. Chem. Lett. 2017, 15, 265–281. [Google Scholar] [CrossRef]
- Greenblum, S.; Turnbaugh, P.J.; Borenstein, E. Metagenomic systems biology of the human gut microbiome reveals topological shifts associated with obesity and inflammatory bowel disease. Proc. Natl. Acad. Sci. USA 2012, 109, 594–599. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wu, H.; Shen, Y.; Wang, C.; Wang, P.; Zhang, W.; Gao, Y.; Niu, L. Statistical determination of crucial taxa indicative of pollution gradients in sediments of Lake Taihu, China. Environ. Pollut. 2019, 246, 753–762. [Google Scholar] [CrossRef]
- Zhang, X.L.; Qi, M.; Li, Q.H.; Cui, Z.D.; Yang, Q. Maricaulis alexandrii sp. nov., a novel active bioflocculants-bearing and dimorphic prosthecate bacterium isolated from marine phycosphere. Antonie Van Leeuwenhoek 2021, 114, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Yu, M.; Liu, J.; Lin, H.; Liang, J.; Zhang, X.H. Role of RpoN from Labrenzia aggregata LZB033 (Rhodobacteraceae) in formation of flagella and biofilms, motility, and environmental adaptation. Appl. Environ. Microbiol. 2019, 85, e02844-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morohoshi, T.; Yokoyama, Y.; Ouchi, M.; Kato, N.; Ikeda, T. Motility and the expression of the flagellin protein FliC are negatively regulated by quorum sensing in Edwardsiella tarda. J. Biosci. Bioeng. 2009, 108, 314–318. [Google Scholar] [CrossRef] [PubMed]
- Bora, T.; Sathe, P.; Laxman, K.; Dobretsov, S.; Dutta, J. Defect engineered visible light active ZnO nanorods for photocatalytic treatment of water. Catal. Today 2017, 284, 11–18. [Google Scholar] [CrossRef]
- Tout, J.; Jeffries, T.C.; Petrou, K.; Tyson, G.W.; Webster, N.S.; Garren, M.; Stocker, R.; Ralph, P.J.; Seymour, J.R. Chemotaxis by natural populations of coral reef bacteria. ISME J. 2015, 9, 1764–1777. [Google Scholar] [CrossRef]
- Dai, F.; Li, Y.; Shao, Y.N.; Li, C.H.; Zhang, W.W. FliC of Vibrio splendidus-related strain involved in adhesion to Apostichopus japonicus. Microb. Pathog. 2020, 149, 104503. [Google Scholar] [CrossRef]
- Yang, X.H.; Thornburg, T.; Suo, Z.Y.; Jun, S.M.; Robison, A.; Li, J.Q.; Lim, T.; Cao, L.; Hoyt, T.; Avci, R.; et al. Flagella overexpression attenuates Salmonella pathogenesis. PLoS ONE 2012, 7, e46828. [Google Scholar] [CrossRef]
- Choi, Y.; Kim, S.; Hwang, H.; Kim, K.-P.; Kang, D.H.; Ryu, S. Plasmid-encoded MCP is involved in virulence, motility, and biofilm formation of Cronobacter sakazakii ATCC 29544. Infect. Immun. 2015, 83, 197–204. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Xu, T.T.; Fossheim, L.E.; Zhang, X.H. Flic, a flagellin protein, is essential for the growth and virulence of fish pathogen Edwardsiella tarda. PLoS ONE 2012, 7, e45070. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, E.J.; Marcano, J.; Camilli, A. Isolation of Streptococcus pneumoniae biofilm mutants and their characterization during nasopharyngeal colonization. Infect. Immun. 2008, 76, 5049–5061. [Google Scholar] [CrossRef] [Green Version]
- Klauck, G.; Serra, D.O.; Possling, A.; Hengge, R. Spatial organization of different sigma factor activities and c-di-GMP signalling within the three-dimensional landscape of a bacterial biofilm. Open Biol. 2018, 8, 180066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Song, S.; Sheng, L.; Zhu, L.; Kim, J.-S.; Wood, T.K. Substrate binding protein DppA1 of ABC transporter DppBCDF increases biofilm formation in Pseudomonas aeruginosa by inhibiting Pf5 prophage lysis. Front. Microbiol. 2018, 9, 30. [Google Scholar] [CrossRef] [PubMed]
- Maddocks, S.E.; Oyston, P.C. Structure and function of the LysR-type transcriptional regulator (LTTR) family proteins. Microbiology 2008, 154, 3609–3623. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.; Krishnan, G.; Goumnerov, B.; Tsongalis, J.; Tompkins, R.; Rahme, L.G. A quorum sensing-associated virulence gene of Pseudomonas aeruginosa encodes a LysR-like transcription regulator with a unique self-regulatory mechanism. Proc. Natl. Acad. Sci. USA 2001, 98, 14613–14618. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Kim, J.G.; Kang, Y.; Jang, J.Y.; Jog, G.J.; Lim, J.Y.; Kim, S.; Suga, H.; Nagamatsu, T.; Hwang, I. Quorum sensing and the LysR-type transcriptional activator ToxR regulate toxoflavin biosynthesis and transport in Burkholderia glumae. Mol. Microbiol. 2004, 54, 921–934. [Google Scholar] [CrossRef] [PubMed]
- Sperandio, V.; Li, C.C.; Kaper, J.B. Quorum-sensing Escherichia coli regulator A: A regulator of the LysR family involved in the regulation of the locus of enterocyte effacement pathogenicity island in enterohemorrhagic E. coli. Infect. Immun. 2002, 70, 3085–3093. [Google Scholar] [CrossRef] [Green Version]
- Ross, D.; Kepa, J.K.; Winski, S.L.; Beall, H.D.; Anwar, A.; Siegel, D. NAD(P)H:quinone oxidoreductase 1 (NQO1): Chemoprotection, bioactivation, gene regulation and genetic polymorphisms. Chem. Biol. Interact. 2000, 129, 77–97. [Google Scholar] [CrossRef]
- Kostova, A.T.D.; Talalay, P. Persuasive evidence that quinone reductase type 1 (DT diaphorase) protects cells against the toxicity of electrophiles and reactive forms of oxygen. Free. Radic. Bio. Med. 2000, 29, 231–240. [Google Scholar] [CrossRef]
- Imlay, J.A. Where in the world do bacteria experience oxidative stress? Environ. Microbiol. 2019, 21, 521–530. [Google Scholar] [CrossRef]
- Dong, T.G.; Dong, S.Q.; Catalano, C.; Moore, R.; Liang, X.Y.; Mekalanos, J.J. Generation of reactive oxygen species by lethal attacks from competing microbes. Proc. Natl. Acad. Sci. USA 2015, 112, 2181–2186. [Google Scholar] [CrossRef] [Green Version]
- Mcdowell, R.E.; Amsler, C.D.; Dickinson, D.A.; Mcclintock, J.B.; Baker, B.J. Reactive oxygen species and the antarctic macroalgal wound response. J. Phycol. 2014, 50, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, S.R.; Moran, J.J.; Stegen, J.C.; Renslow, R.S.; Hutchison, J.R.; Cole, J.K.; Dohnalkova, A.C.; Tremblay, J.; Singh, K.; Malfatti, S.A.; et al. The epsomitic phototrophic microbial mat of Hot Lake, Washington: Community structural responses to seasonal cycling. Front. Microbiol. 2013, 4, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindemann, S.R.; Mobberley, J.M.; Cole, J.K.; Markillie, L.M.; Taylor, R.C.; Huang, E.; Chrisler, W.B.; Wiley, H.S.; Lipton, M.S.; Nelson, W.C.; et al. Predicting species-resolved macronutrient acquisition during succession in a model phototrophic biofilm using an integrated ‘omics approach. Front. Microbiol. 2017, 8, 1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baruah, S.; Jaisa, M.; Dutta, J. Development of a visible light active photocatalytic portable water purification unit using ZnO nanorods. Catal. Sci. Technol. 2012, 2, 918–921. [Google Scholar] [CrossRef]
- Jones, N.; Ray, B.; Ranjit, K.T.; Adhar, C.; Mannajones, N. Antibacterial activity of ZnO nanoparticle suspensions on a broad spectrum of microorganisms. FEMS Microbiol. Lett. 2008, 279, 71–76. [Google Scholar] [CrossRef] [Green Version]
- Heydorn, A.; Nielsen, A.T.; Hentzer, M.; Sternberg, C.; Givskov, M.; Ersbøll, B.K.; Heydorn, A.; Sternberg, C.; Nielsen, A.T.; Molin, S. Quantification of biofilm structures by the novel computer program COMSTAT. Microbiology 2000, 146, 2395–2407. [Google Scholar] [CrossRef] [Green Version]
- Hentzer, M.; Teitzel, G.M.; Balzer, G.J.; Heydorn, A.; Molin, S.; Givskov, M.; Parsek, M.R. Alginate overproduction affects Pseudomonas aeruginosa biofilm structure and function. J. Bacteriol. 2001, 183, 5395–5401. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.J.; Zeng, Y.H.; Zhu, J.M.; Cai, Z.H.; Zhou, J. The structure and assembly mechanisms of plastisphere microbial community in natural marine environment. J. Hazard. Mater. 2022, 421, 126780. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Oberbeckmann, S.; Kreikemeyer, B.; Labrenz, M. Environmental factors support the formation of specific bacterial assemblages on microplastics. Front. Microbiol. 2018, 8, 2709. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, L.Z.; Wang, P.F.; Hou, J.; Yao, Y.; Liu, Z.L.; Liu, S.Q.; Li, T.F. Distinct community structure and microbial functions of biofilms colonizing microplastics. Sci. Total Environ. 2019, 650, 2395–2402. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Chen, G.F.; Ying, K.Z.; Jin, H.; Song, J.T.; Cai, Z.H. Phycosphere microbial succession patterns and assembly mechanisms in a marine dinoflagellate bloom. Appl. Environ. Microb. 2019, 85, e00319–e00349. [Google Scholar] [CrossRef] [Green Version]
- Reshef, D.N.; Reshef, Y.A.; Finucane, H.K.; Grossman, S.R.; Mcvean, G.; Turnbaugh, P.J.; Lander, E.S.; Mitzenmacher, M.; Sabeti, P.C. Detecting novel associations in large data sets. Science 2011, 334, 1518–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolge, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Li, D.H.; Liu, C.M.; Luo, R.B.; Sadakane, K.; Lam, T.W. MEGAHIT: An ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef] [Green Version]
- Li, D.H.; Luo, R.B.; Liu, C.M.; Leung, C.M.; Ting, H.F.; Sadakane, K.; Yamashita, H.; Lam, T.W. MEGAHIT v1.0: A fast and scalable metagenome assembler driven by advanced methodologies and community practices. Methods 2016, 102, 3–11. [Google Scholar] [CrossRef]
- Peng, Y.; Leung, H.C.M.; Yiu, S.M.; Chin, F.Y.L. IDBA-UD: A de novo assembler for single-cell and metagenomic sequencing data with highly uneven depth. Bioinformatics 2012, 28, 1420–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, H.B.; Almeida, M.; Juncker, A.S.; Rasmussen, S.; Li, J.H.; Sunagawa, S.; Plichta, D.R.; Gautier, L.; Pedersen, A.G.; Le Chatelier, E.; et al. Identification and assembly of genomes and genetic elements in complex metagenomic samples without using reference genomes. Nat. Biotechnol. 2014, 32, 822–828. [Google Scholar] [CrossRef] [PubMed]
- Walt, A.J.V.D.; Goethem, M.W.V.; Ramond, J.B.; Makhalanyane, T.P.; Reva, O.; Cowan, D.A. Assembling metagenomes, one community at a time. BMC Genom. 2017, 18, 521. [Google Scholar]
- Lukashin, A.V.; Borodovsky, M. GeneMark.hmm: New solutions for gene finding. Nucleic Acids Res. 1998, 26, 1107–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Number of Nodes | Number of Edges | Positive Correlation | Negative Correlation | Average Degree | Average Path Length | Average Clustering Coefficient | Figure Density | Modularity Index |
|---|---|---|---|---|---|---|---|---|---|
| Group A (PL) | 48 | 488 | 70.90% | 20.10% | 10.167 | 2.835 | 0.649 | 0.216 | 0.281 |
| Group B (P) | 47 | 270 | 66.67% | 33.33% | 5.745 | 3.816 | 0.547 | 0.125 | 0.468 |
| Group C (L) | 46 | 372 | 62.37% | 37.63% | 8.087 | 2.456 | 0.525 | 0.180 | 0.346 |
| Group D (N) | 45 | 274 | 70.07% | 29.93% | 6.089 | 3.201 | 0.52 | 0.138 | 0.436 |
| Seawater | 49 | 442 | 56.11% | 43.89% | 9.02 | 2.934 | 0.554 | 0.188 | 0.398 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.-C.; Wang, B.; Zeng, Y.-H.; Cai, Z.-H.; Zhou, J. The Microbial Mechanisms of a Novel Photosensitive Material (Treated Rape Pollen) in Anti-Biofilm Process under Marine Environment. Int. J. Mol. Sci. 2022, 23, 3837. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23073837
Li Q-C, Wang B, Zeng Y-H, Cai Z-H, Zhou J. The Microbial Mechanisms of a Novel Photosensitive Material (Treated Rape Pollen) in Anti-Biofilm Process under Marine Environment. International Journal of Molecular Sciences. 2022; 23(7):3837. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23073837
Chicago/Turabian StyleLi, Qing-Chao, Bo Wang, Yan-Hua Zeng, Zhong-Hua Cai, and Jin Zhou. 2022. "The Microbial Mechanisms of a Novel Photosensitive Material (Treated Rape Pollen) in Anti-Biofilm Process under Marine Environment" International Journal of Molecular Sciences 23, no. 7: 3837. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23073837