
Titanium Surface Characteristics Induce the Specific Reprogramming of Toll-like Receptor Signaling in Macrophages


 ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
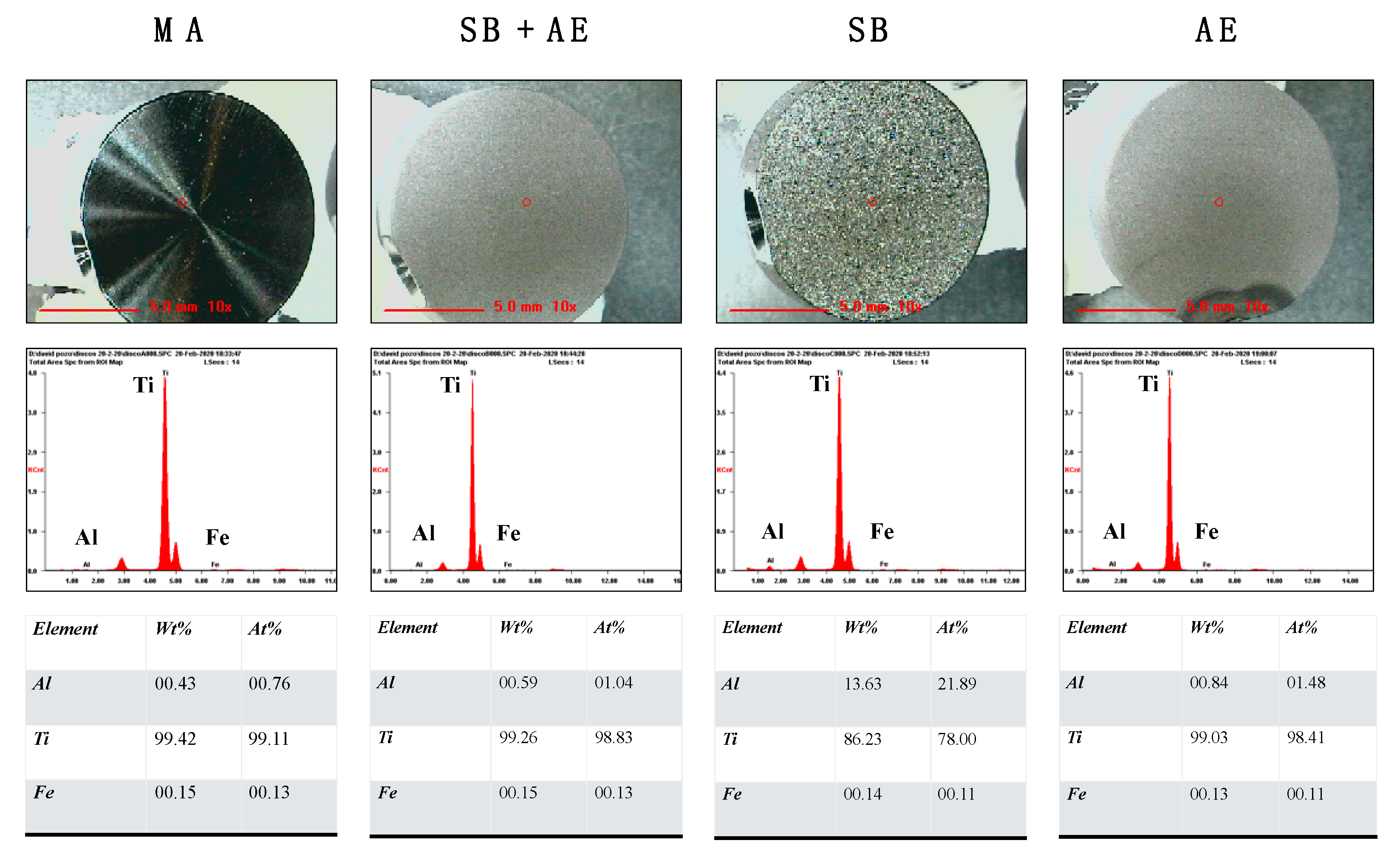
2.1. Characterization of Ti-Discs
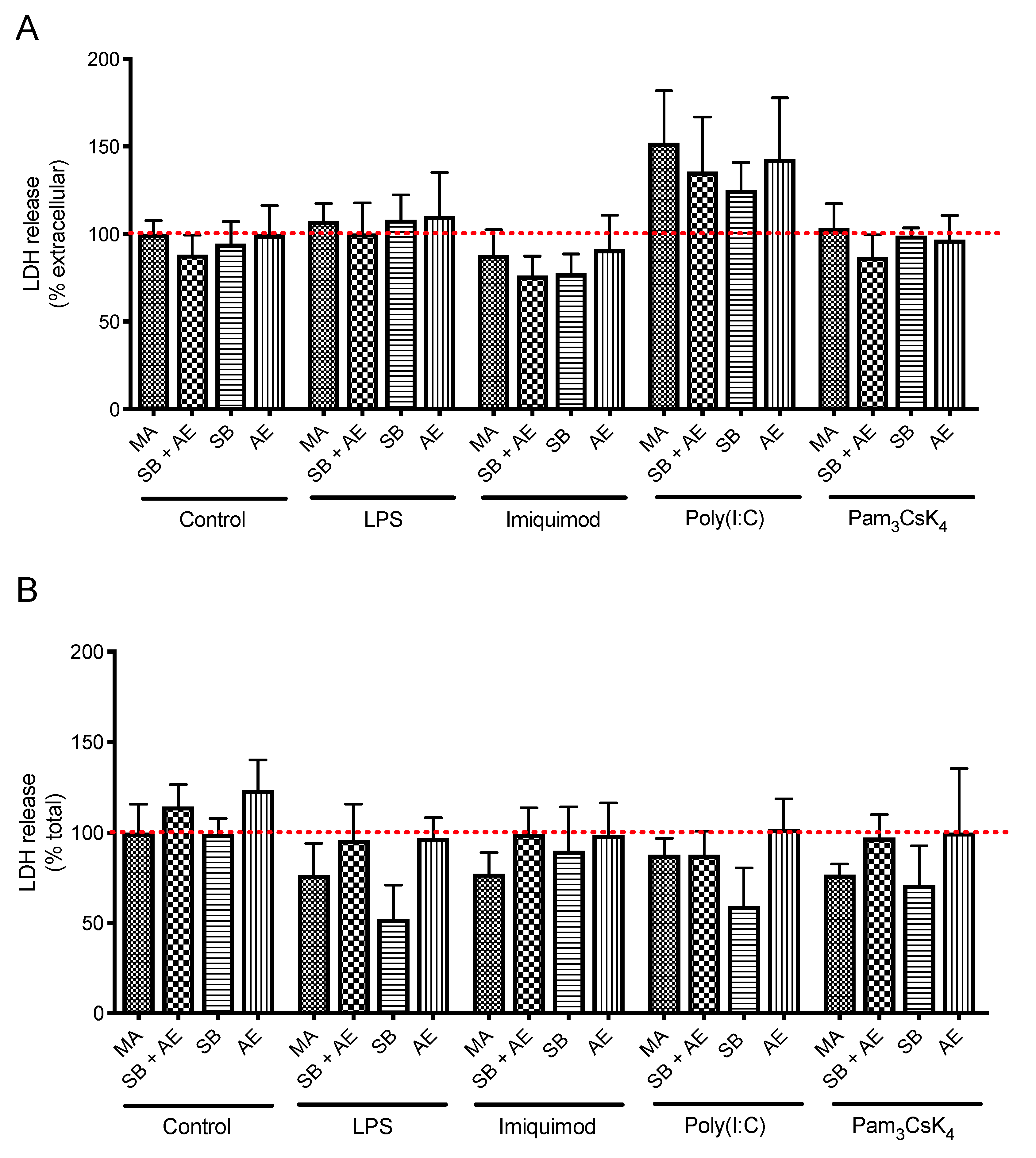
2.2. Cell Viability of Resting and TLR-Stimulated Macrophage Cultures in Different Ti-Discs
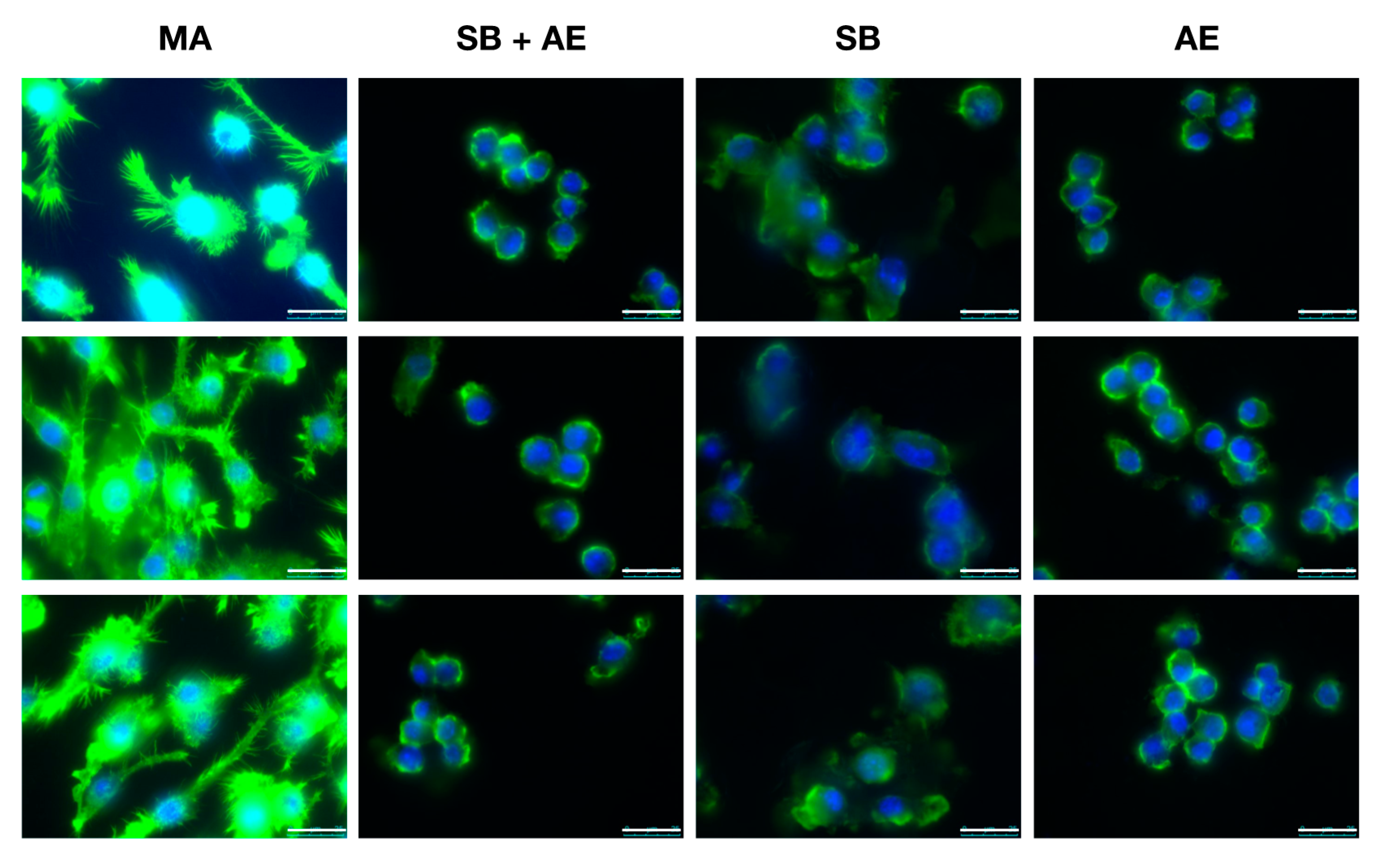
2.3. Macrophage Cell Morphology in Different Ti-Discs Cultures
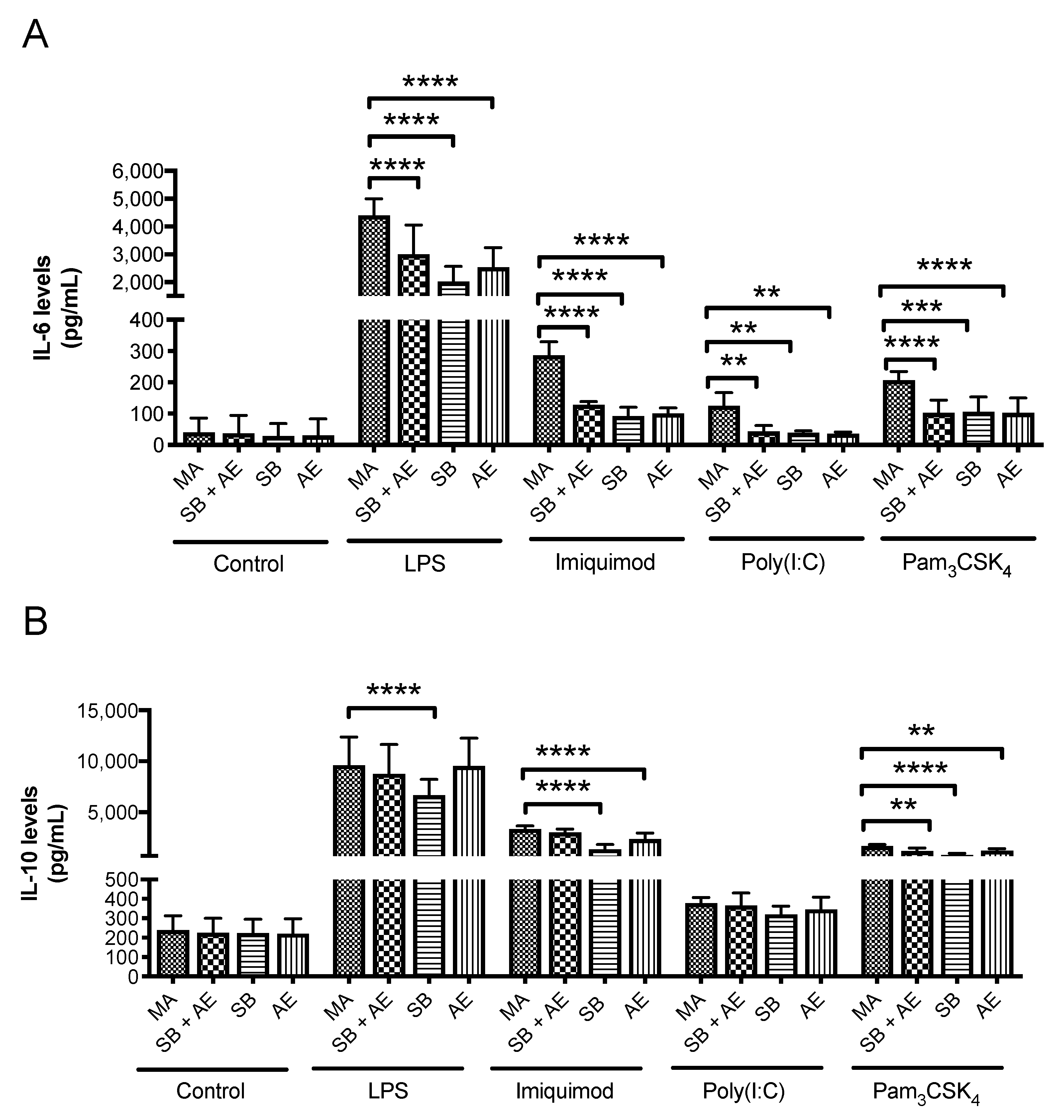
2.4. Determination of Cytokine Levels
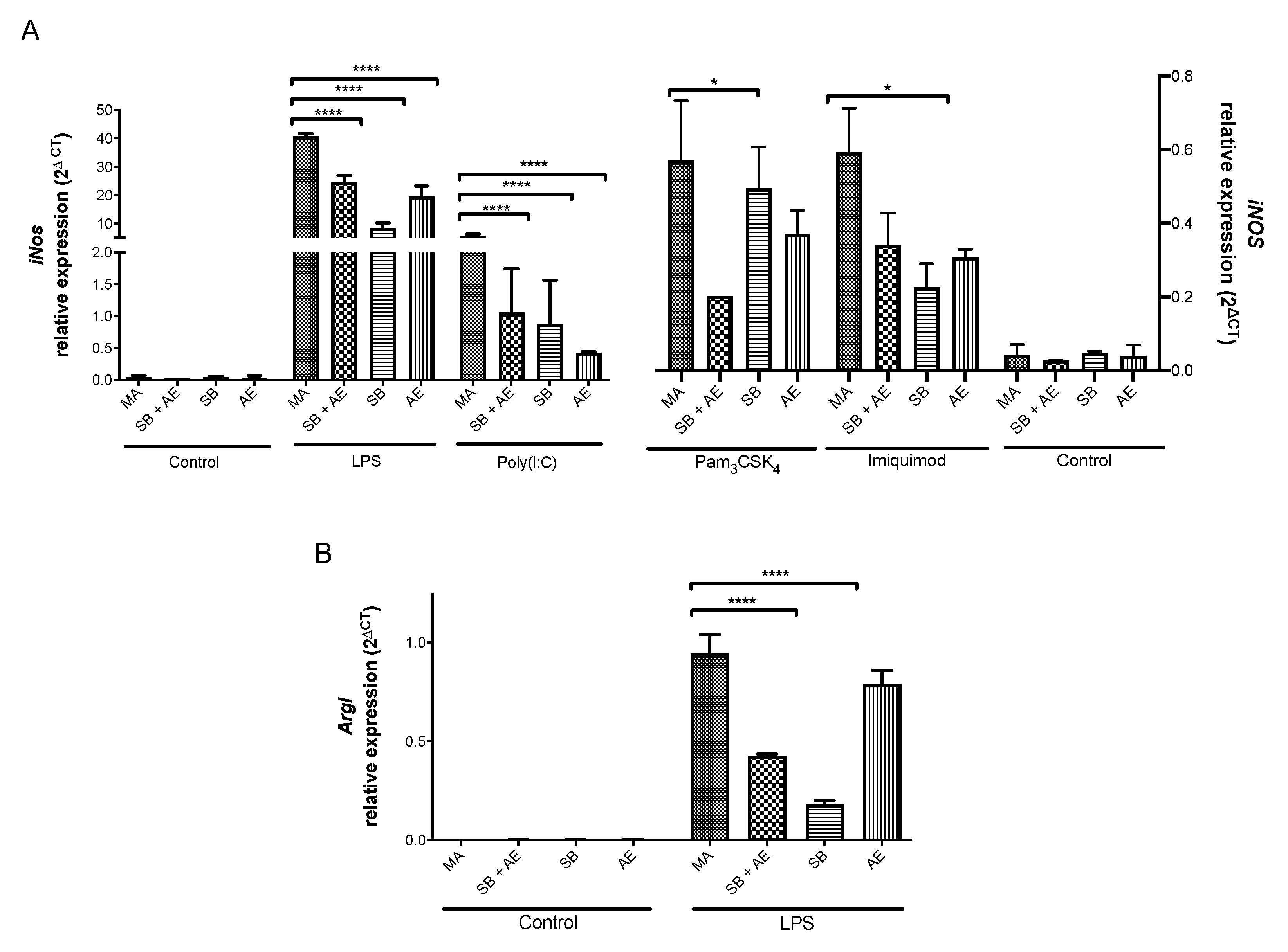
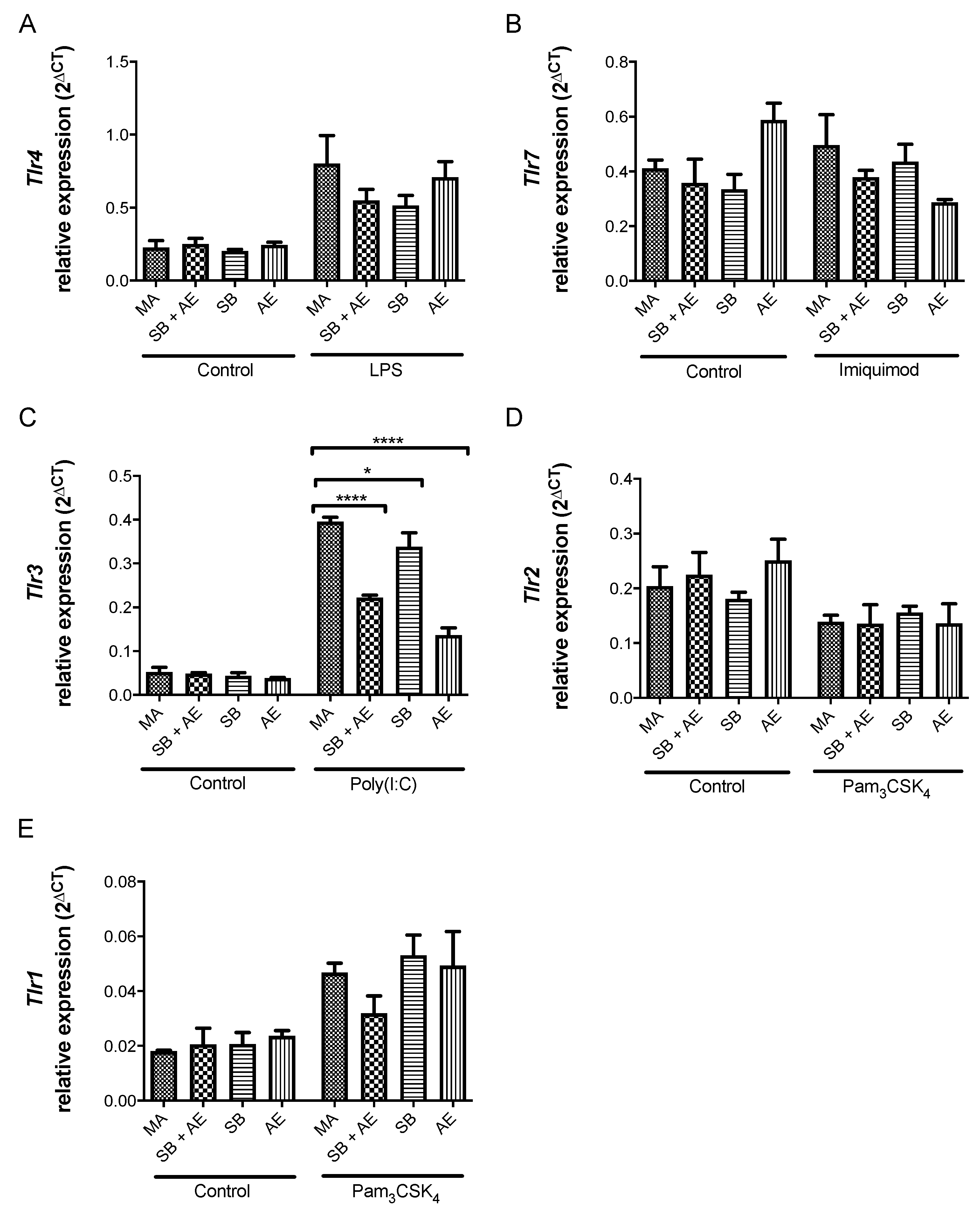
2.5. Gene Expression of iNos/ArgI and Tlr Receptors in Macrophages Exposed to Different Ti-Discs
3. Materials and Methods
3.1. Titanium-Based Materials
3.2. Chemical Composition of Ti-Discs
3.3. Scanning Electron Microscopy
3.4. Surface Imaging, Analysis and Metrology by Interferometer
3.5. Murine Macrophage Cell Culture
3.6. Cell Morphology by Fluorescence Microscopy
3.7. TLR Stimulation
3.8. Cell Viability and Plasma Membrane Integrity by LDH Assay
3.9. Cytokine Release
3.10. Relative Gene Expression of mRNAs
3.11. Data Analysis
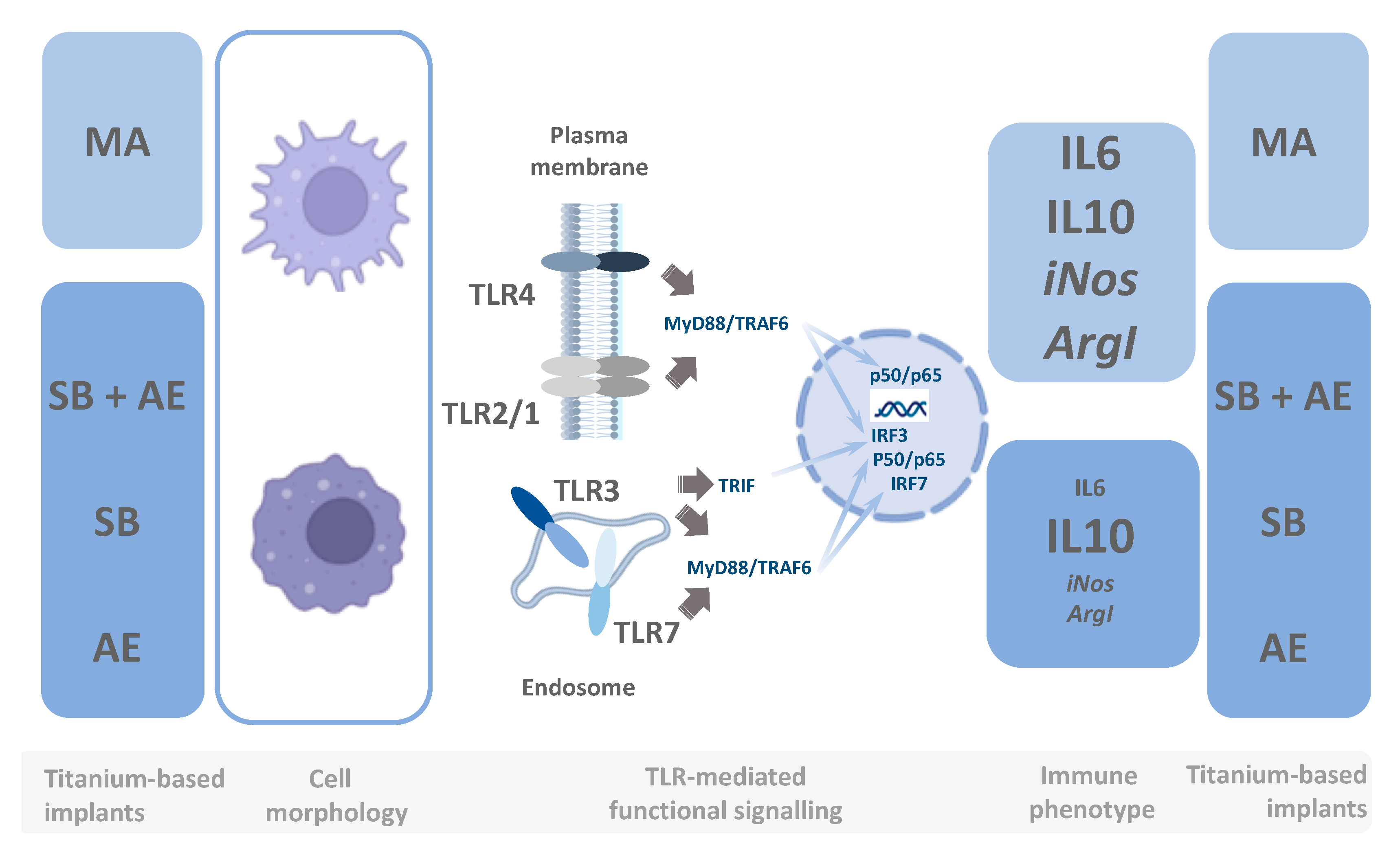
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ryu, H.; Seo, M.; Rogers, J.A. Bioresorbable Metals for Biomedical Applications: From Mechanical Components to Electronic Devices. Adv. Health. Mater. 2021, 10, 2002236. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Bathaei, M.J.; Istif, E.; Beker, L. A Review of Bioresorbable Implantable Medical Devices: Materials, Fabrication, and Implementation. Adv. Health. Mater. 2020, 9, 2000790. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Wu, Z.; Xu, K.; Wang, X.; Wang, S.; Qiu, H.; Li, X.; Chen, J. Multifunctional Coatings of Titanium Implants Toward Promoting Osseointegration and Preventing Infection: Recent Developments. Front. Bioeng. Biotechnol. 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Roehling, S.; Schlegel, K.A.; Woelfler, H.; Gahlert, M. Zirconia compared to titanium dental implants in preclinical studies—A systematic review and meta-analysis. Clin. Oral Implant. Res. 2019, 30, 365–395. [Google Scholar] [CrossRef] [PubMed]
- Tchinda, A.P.; Pierson, G.; Kouitat-Njiwa, R.; Bravetti, P. The Surface Conditions and Composition of Titanium Alloys in Implantology: A Comparative Study of Dental Implants of Different Brands. Materials 2022, 15, 1018. [Google Scholar] [CrossRef] [PubMed]
- Gil, J.; Manero, J.M.; Ruperez, E.; Velasco-Ortega, E.; Jiménez-Guerra, A.; Ortiz-García, I.; Monsalve-Guil, L. Mineralization of Titanium Surfaces: Biomimetic Implants. Materials 2021, 14, 2879. [Google Scholar] [CrossRef] [PubMed]
- Velasco-Ortega, E.; Ortiz-Garcia, I.; Jiménez-Guerra, A.; Núñez-Márquez, E.; Moreno-Muñoz, J.; Rondón-Romero, J.; Cabanillas-Balsera, D.; Gil, J.; Muñoz-Guzón, F.; Monsalve-Guil, L. Osseointegration of Sandblasted and Acid-Etched Implant Surfaces. A Histological and Histomorphometric Study in the Rabbit. Int. J. Mol. Sci. 2021, 22, 8507. [Google Scholar] [CrossRef]
- Trindade, R.; Albrektsson, T.; Galli, S.; Prgomet, Z.; Tengvall, P.; Wennerberg, A. Osseointegration and foreign body reaction: Titanium implants activate the immune system and suppress bone resorption during the first 4 weeks after implantation. Clin. Implant. Dent. Relat. Res. 2018, 20, 82–91. [Google Scholar] [CrossRef]
- Trindade, R.; Albrektsson, T.; Tengvall, P.; Wennerberg, A. Foreign Body Reaction to Biomaterials: On Mechanisms for Buildup and Breakdown of Osseointegration. Clin. Implant. Dent. Relat. Res. 2016, 18, 192–203. [Google Scholar] [CrossRef]
- Albrektsson, T.; Wennerberg, A. On osseointegration in relation to implant surfaces. Clin. Implant. Dent. Relat. Res. 2019, 21, 4–7. [Google Scholar] [CrossRef] [Green Version]
- Wennerberg, A.; Albrektsson, T.; Chrcanovic, B. Long-term clinical outcome of implants with different surface modifications. Eur. J. Oral Implant. 2018, 11, S123–S136. [Google Scholar]
- Chen, X.; Li, H.S.; Yin, Y.; Feng, Y.; Tan, X.W. Macrophage Proinflammatory Response to the Titanium Alloy Equipment in Dental Implantation. Genet. Mol. Res. 2015, 14, 9155–9162. [Google Scholar] [CrossRef] [PubMed]
- Ottria, L.; Lauritano, D.; Andreasi Bassi, M.; Palmieri, A.; Candotto, V.; Tagliabue, A.; Tettamanti, L. Mechanical, Chemicaand Biological Aspects of Titanium and Titanium Alloys in Implant Dentistry. J. Biol. Regul. Homeost. Agents 2018, 32, 81–90. [Google Scholar] [PubMed]
- Stich, T.; Alagboso, F.; Křenek, T.; Kovářík, T.; Alt, V.; Docheva, D. Implant-Bone-Interface: Reviewing the Impact of Titanium Surface Modifications on Osteogenic Processes In Vitro and In Vivo. Bioeng. Transl. Med. 2022, 7, e10239. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Liang, H.; Liu, X.; Wu, J.; Yang, C.; Wong, T.M.; Kwan, K.Y.H.; Cheung, K.M.C.; Wu, S.; Yeung, K.W.K. Regulation of macrophage polarization through surface topography design to facilitate implant-to-bone osteointegration. Sci. Adv. 2021, 7, eabf6654. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, Z.; Xu, Z.; Zhang, Y.; Ye, H.; Wang, X. Macrophage responses to selective laser-melted Ti-6Al-4V scaffolds of different pore geometries and the corresponding osteoimmunomodulatory effects toward osteogenesis. J. Biomed. Mater. Res. Part A 2022, 110, 873–883. [Google Scholar] [CrossRef]
- Zhang, T.; Jiang, M.; Yin, X.; Yao, P.; Sun, H. The role of autophagy in the process of osseointegration around titanium implants with micro-nano topography promoted by osteoimmunity. Sci. Rep. 2021, 11, 18418. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, T.; Jiang, M.; Yin, X.; Luo, X.; Sun, H. Effect of the immune responses induced by implants in a integrated three-dimensional micro-nano topography on osseointegration. J. Biomed. Mater. Res. Part A 2021, 109, 1429–1440. [Google Scholar] [CrossRef]
- Dai, X.; Wei, Y.; Zhang, X.; Meng, S.; Mo, X.; Liu, X.; Deng, X.; Zhang, L.; Deng, X. Attenuating Immune Response of Macrophage by Enhancing Hydrophilicity of Ti Surface. J. Nanomater. 2015, 2015, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Hamlet, S.; Alfarsi, M.; George, R.; Ivanovski, S. The effect of hydrophilic titanium surface modification on macrophage inflammatory cytokine gene expression. Clin. Oral Implant. Res. 2012, 23, 584–590. [Google Scholar] [CrossRef]
- Hamlet, S.M.; Lee, R.S.; Moon, H.; Alfarsi, M.A.; Ivanovski, S. Hydrophilic titanium surface-induced macrophage modulation promotes pro-osteogenic signalling. Clin. Oral Implant. Res. 2019, 30, 1085–1096. [Google Scholar] [CrossRef] [PubMed]
- Gao, A.; Liao, Q.; Xie, L.; Wang, G.; Zhang, W.; Wu, Y.; Li, P.; Guan, M.; Pan, H.; Tong, L.; et al. Tuning the surface immunomodulatory functions of polyetheretherketone for enhanced osseointegration. Biomaterials 2020, 230, 119642. [Google Scholar] [CrossRef] [PubMed]
- Novak, T.; Hamedi, M.; Bergmeier, L.A.; Fortune, F.; Hagi-Pavli, E. Saliva and Serum Cytokine Profiles during Oral Ulceration in Behçet’s Disease. Front. Immunol. 2021, 12, 724900. [Google Scholar] [CrossRef] [PubMed]
- Barutta, F.; Bellini, S.; Durazzo, M.; Gruden, G. Novel Insight into the Mechanisms of the Bidirectional Relationship between Diabetes and Periodontitis. Biomedicines 2022, 10, 178. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G.; Chavakis, T. Local and Systemic Mechanisms Linking Periodontal Disease and Inflammatory Comorbidities. Nat. Rev. Immunol. 2021, 21, 426–440. [Google Scholar] [CrossRef] [PubMed]
- Cheah, C.W.; Al-Maleki, A.R.; Vaithilingam, R.D.; Vadivelu, J.; Sockalingam, S.; Baharuddin, N.A.; Bartold, P.M. Associations between inflammation-related LL-37 with subgingival microbial dysbiosis in rheumatoid arthritis patients. Clin. Oral Investig. 2022, 1–12. [Google Scholar] [CrossRef]
- Konkel, J.E.; O’Boyle, C.; Krishnan, S. Distal Consequences of Oral Inflammation. Front. Immunol. 2019, 10, 1403. [Google Scholar] [CrossRef] [Green Version]
- Sabharwal, A.; Stellrecht, E.; Scannapieco, F.A. Associations between dental caries and systemic diseases: A scoping review. BMC Oral Health 2021, 21, 1–35. [Google Scholar] [CrossRef]
- Zengeler, K.E.; Lukens, J.R. Innate Immunity at the Crossroads of Healthy Brain Maturation and Neurodevelopmental Disorders. Nat. Rev. Immunol. 2021, 21, 454–468. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. Signaling to NF-ΚB by Toll-like Receptors. Trends Mol. Med. 2007, 13, 460–469. [Google Scholar] [CrossRef]
- Romo, M.R.; Pérez-Martínez, D.; Ferrer, C.C. Innate immunity in vertebrates: An overview. Immunology 2016, 148, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, K.A.; Kagan, J.C. Toll-like Receptors and the Control of Immunity. Cell 2020, 180, 1044–1066. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wu, M. Pattern recognition receptors in health and diseases. Signal Transduct. Target. Ther. 2021, 6, 291. [Google Scholar] [CrossRef] [PubMed]
- Cejudo-Guillén, M.; Ramiro-Gutiérrez, M.; Labrador-Garrido, A.; Cuenca, A.D.; Pozo, D. Nanoporous silica microparticle interaction with toll-like receptor agonists in macrophages. Acta Biomater. 2012, 8, 4295–4303. [Google Scholar] [CrossRef]
- Castillo, P.M.; Herrera, J.L.; Fernandez-Montesinos, R.; Caro, C.; Zaderenko, A.P.; Mejías, J.A.; Pozo, D. Tiopronin monolayer-protected silver nanoparticles modulate IL-6 secretion mediated by Toll-like receptor ligands. Nanomedicine 2008, 3, 627–635. [Google Scholar] [CrossRef]
- Kasemo, B.; Gold, J. Implant Surfaces and Interface Processes. Adv. Dent. Res. 1999, 13, 8–20. [Google Scholar] [CrossRef]
- Prasad, A.; Alizadeh, E. Cell Form and Function: Interpreting and Controlling the Shape of Adherent Cells. Trends Biotechnol. 2019, 37, 347–357. [Google Scholar] [CrossRef]
- Heinrich, F.; Lehmbecker, A.; Raddatz, B.B.; Kegler, K.; Tipold, A.; Stein, V.M.; Kalkuhl, A.; Deschl, U.; Baumgärtner, W.; Ulrich, R.; et al. Morphologic, phenotypic, and transcriptomic characterization of classically and alternatively activated canine blood-derived macrophages in vitro. PLoS ONE 2017, 12, e0183572. [Google Scholar] [CrossRef]
- Sigola, L.B.; Fuentes, A.-L.; Millis, L.M.; Vapenik, J.; Murira, A. Effects of Toll-like receptor ligands on RAW 264.7 macrophage morphology and zymosan phagocytosis. Tissue Cell 2016, 48, 389–396. [Google Scholar] [CrossRef]
- Pegueroles, M.; Aparicio, C.; Bosio, M.; Engel, E.; Gil, F.J.; Planell, J.A.; Altankov, G. Spatial organization of osteoblast fibronectin matrix on titanium surfaces: Effects of roughness, chemical heterogeneity and surface energy. Acta Biomater. 2010, 6, 291–301. [Google Scholar] [CrossRef]
- Yang, D.; Wan, Y. Molecular Determinants for the Polarization of Macrophage and Osteoclast. Semin. Immunopathol. 2019, 41, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Macrophage Polarization: Different Gene Signatures in M1(Lps+) vs. Classically and M2(LPS-) vs. Alternatively Activated Macrophages. Front. Immunol. 2019, 10, 1084. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, C. Nitric Oxide Synthase in Innate and Adaptive Immunity: An Update. Trends Immunol. 2015, 36, 161–178. [Google Scholar] [CrossRef] [PubMed]
- Roodveldt, C.; Labrador-Garrido, A.; Gonzalez-Rey, E.; Lachaud, C.C.; Guilliams, T.; Fernandez-Montesinos, R.; Benitez-Rondan, A.; Robledo, G.; Hmadcha, A.; Delgado, M.; et al. Preconditioning of Microglia by α-Synuclein Strongly Affects the Response Induced by Toll-like Receptor (TLR) Stimulation. PLoS ONE 2013, 8, e79160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, H.; Ando, T.; Ogiso, H.; Arioka, Y.; Seishima, M. Inhibition of induced nitric oxide synthase enhances the anti-tumor effects on cancer immunotherapy using TLR7 agonist in mice. Cancer Immunol. Immunother. 2015, 64, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Brieger, A.; Rink, L.; Haase, H. Differential Regulation of TLR-Dependent MyD88 and TRIF Signaling Pathways by Free Zinc Ions. J. Immunol. 2013, 191, 1808–1817. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.; Wang, H.; Hajishengallis, G.N.; Martin, M. TLR-Signaling Networks: An Integration of Adaptor Molecules, Kinases, and Cross-Talk. J. Dent. Res. 2011, 90, 417–427. [Google Scholar] [CrossRef]
- Martins, B.R.; Pinto, T.S.; Fernandes, C.J.D.C.; Bezerra, F.; Zambuzzi, W.F. PI3K/AKT signaling drives titanium-induced angiogenic stimulus. J. Mater. Sci. Mater. Med. 2021, 32, 1–9. [Google Scholar] [CrossRef]
- He, Y.; Li, Z.; Ding, X.; Xu, B.; Wang, J.; Li, Y.; Chen, F.; Meng, F.; Song, W.; Zhang, Y. Nanoporous titanium implant surface promotes osteogenesis by suppressing osteoclastogenesis via integrin β1/FAKpY397/MAPK pathway. Bioact. Mater. 2022, 8, 109–123. [Google Scholar] [CrossRef]
- Jin, Y.; Liu, Y.; Nelin, L.D. Extracellular Signal-regulated Kinase Mediates Expression of Arginase II but Not Inducible Nitric-oxide Synthase in Lipopolysaccharide-stimulated Macrophages. J. Biol. Chem. 2015, 290, 2099–2111. [Google Scholar] [CrossRef] [Green Version]
- Komal, A.; Noreen, M.; El-Kott, A.F. TLR3 Agonists: RGC100, ARNAX, and Poly-IC: A Comparative Review. Immunol. Res. 2021, 69, 312–322. [Google Scholar] [CrossRef]
- Jelinek, I.; Leonard, J.N.; Price, G.E.; Brown, K.N.; Meyer-Manlapat, A.; Goldsmith, P.K.; Wang, Y.; Venzon, D.; Epstein, S.L.; Segal, D.M. TLR3-Specific Double-Stranded RNA Oligonucleotide Adjuvants Induce Dendritic Cell Cross-Presentation, CTL Responses, and Antiviral Protection. J. Immunol. 2011, 186, 2422–2429. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ti-Disc | MA | SB + AE | SB | AE |
|---|---|---|---|---|
| Ra (µm) | 0.026 ± 0.012 | 1.235 ± 0.024 | 0.932 ± 0.523 | 0.603 ± 0.181 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Sánchez, Z.; Areal-Quecuty, V.; Jimenez-Guerra, A.; Cabanillas-Balsera, D.; Gil, F.J.; Velasco-Ortega, E.; Pozo, D. Titanium Surface Characteristics Induce the Specific Reprogramming of Toll-like Receptor Signaling in Macrophages. Int. J. Mol. Sci. 2022, 23, 4285. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23084285
González-Sánchez Z, Areal-Quecuty V, Jimenez-Guerra A, Cabanillas-Balsera D, Gil FJ, Velasco-Ortega E, Pozo D. Titanium Surface Characteristics Induce the Specific Reprogramming of Toll-like Receptor Signaling in Macrophages. International Journal of Molecular Sciences. 2022; 23(8):4285. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23084285
Chicago/Turabian StyleGonzález-Sánchez, Zaira, Victoria Areal-Quecuty, Alvaro Jimenez-Guerra, Daniel Cabanillas-Balsera, Francisco Javier Gil, Eugenio Velasco-Ortega, and David Pozo. 2022. "Titanium Surface Characteristics Induce the Specific Reprogramming of Toll-like Receptor Signaling in Macrophages" International Journal of Molecular Sciences 23, no. 8: 4285. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23084285