
Having a Same Type IIS Enzyme’s Restriction Site on Guide RNA Sequence Does Not Affect Golden Gate (GG) Cloning and Subsequent CRISPR/Cas Mutagenesis


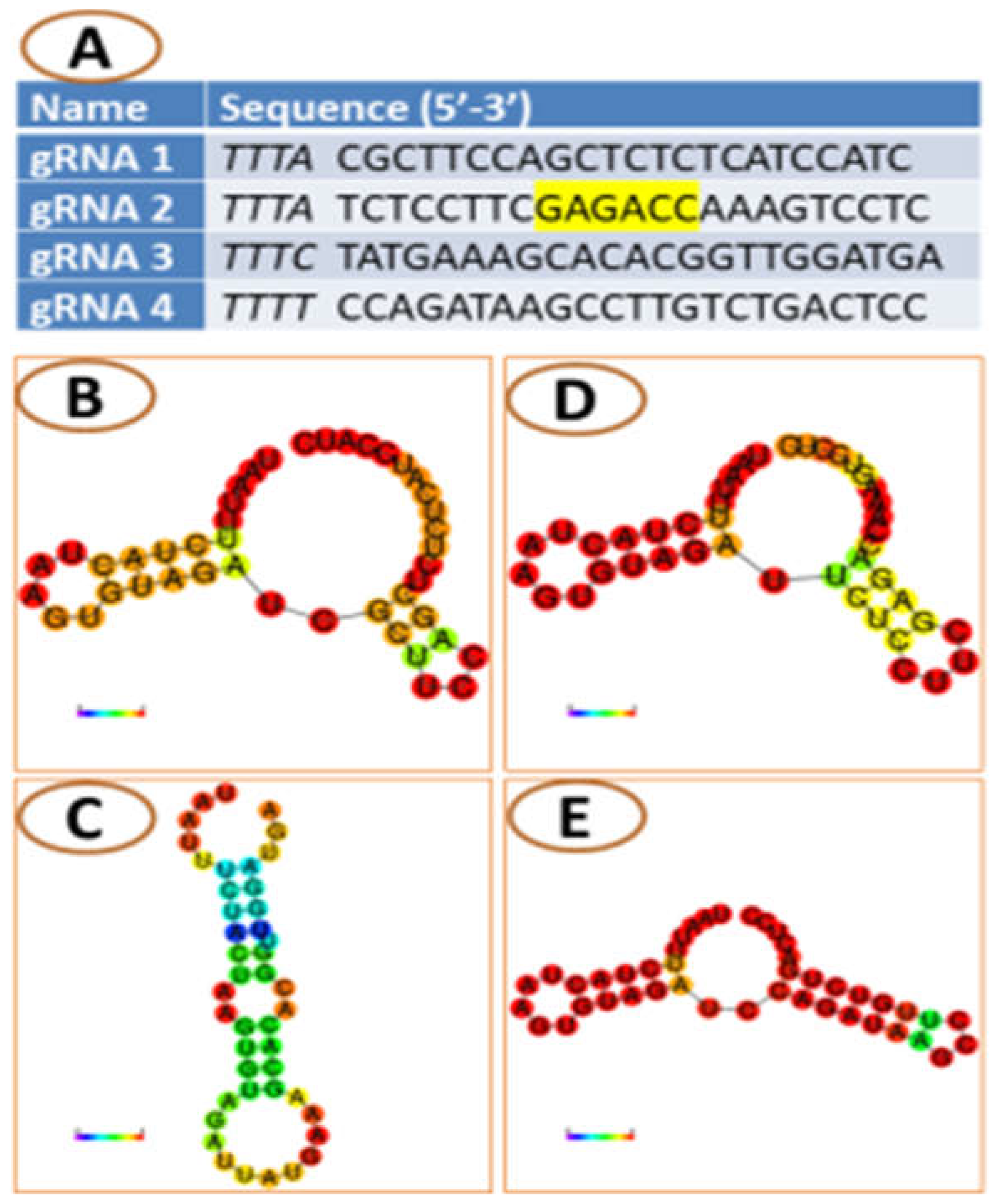{kind=link}
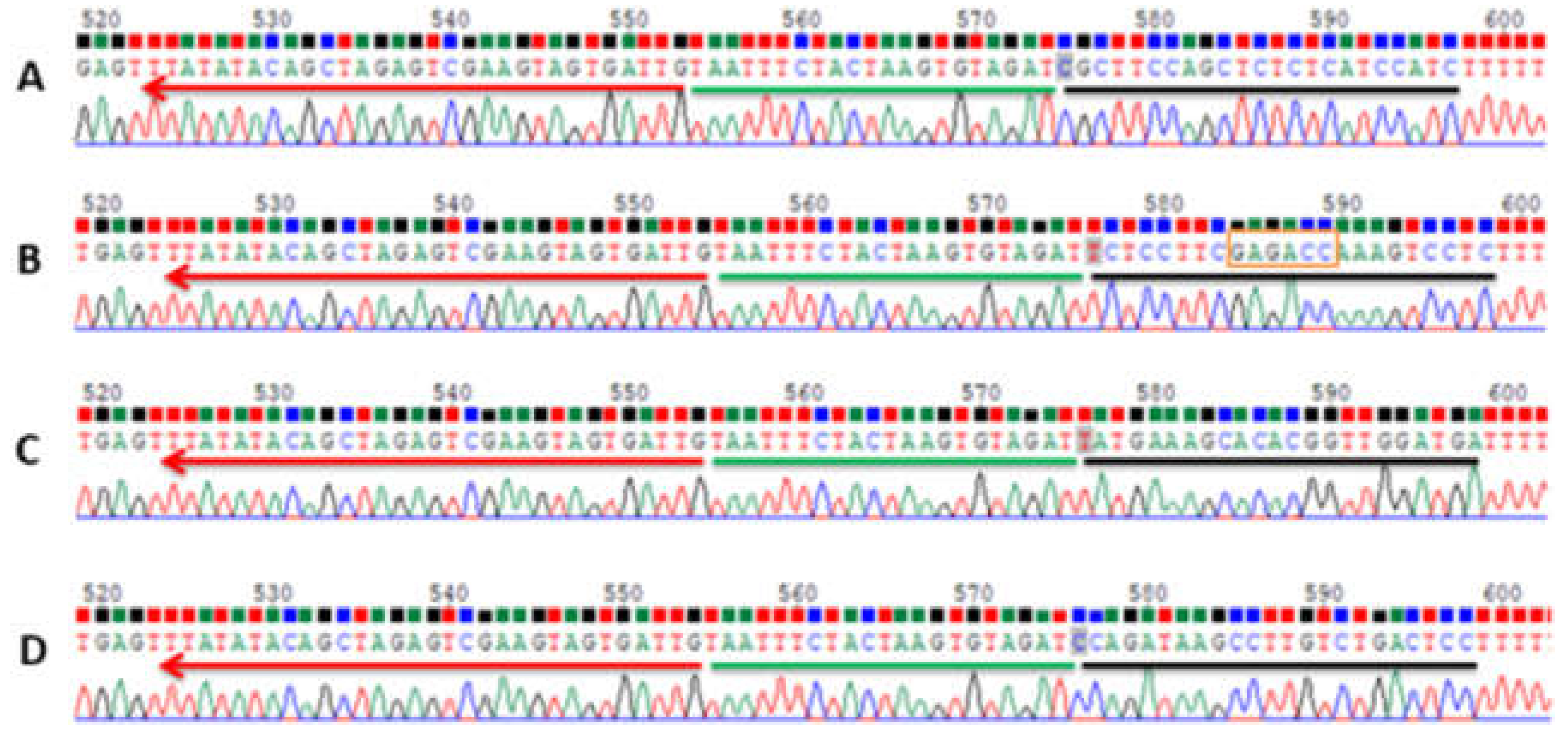{kind=link}
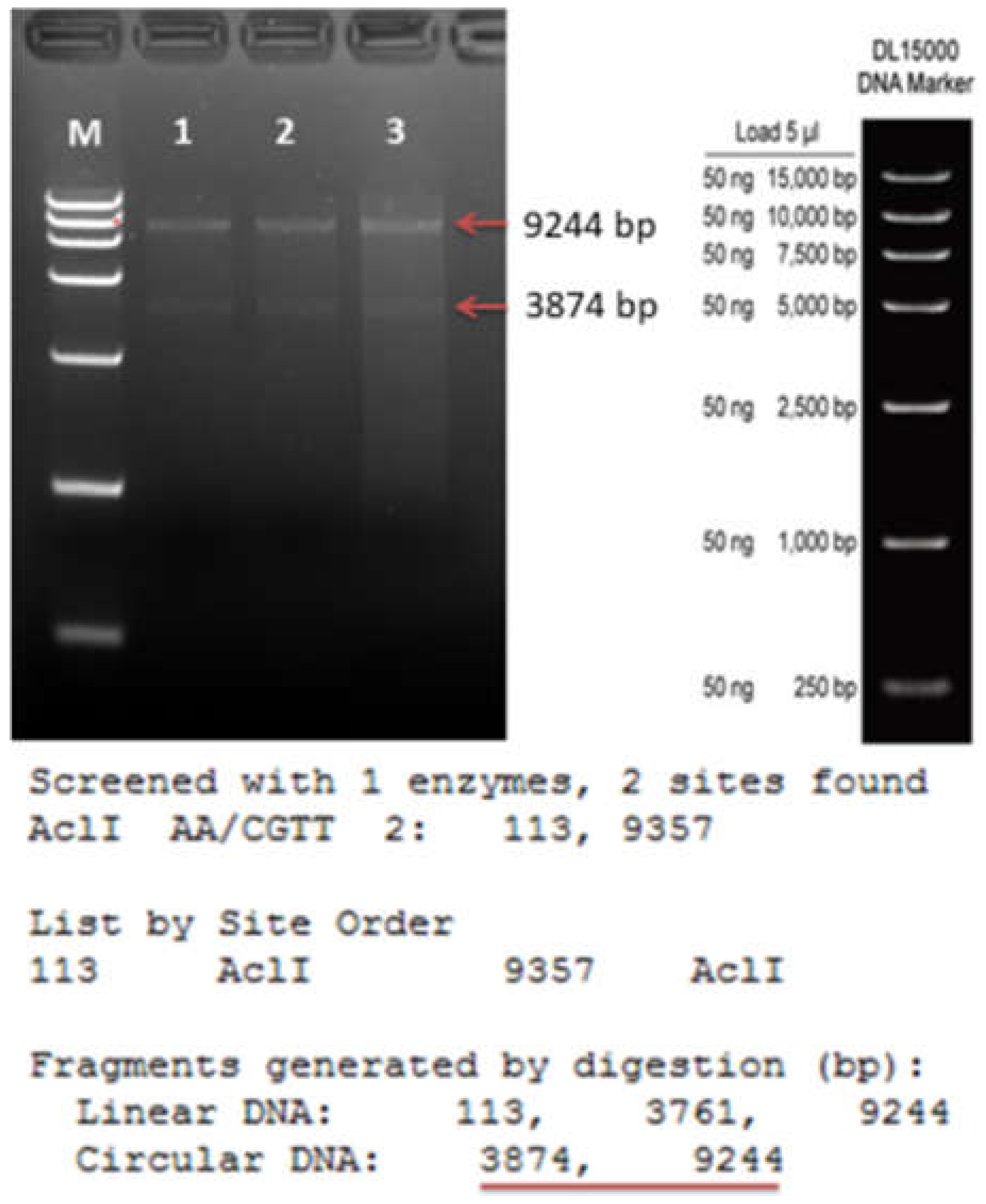{kind=link}
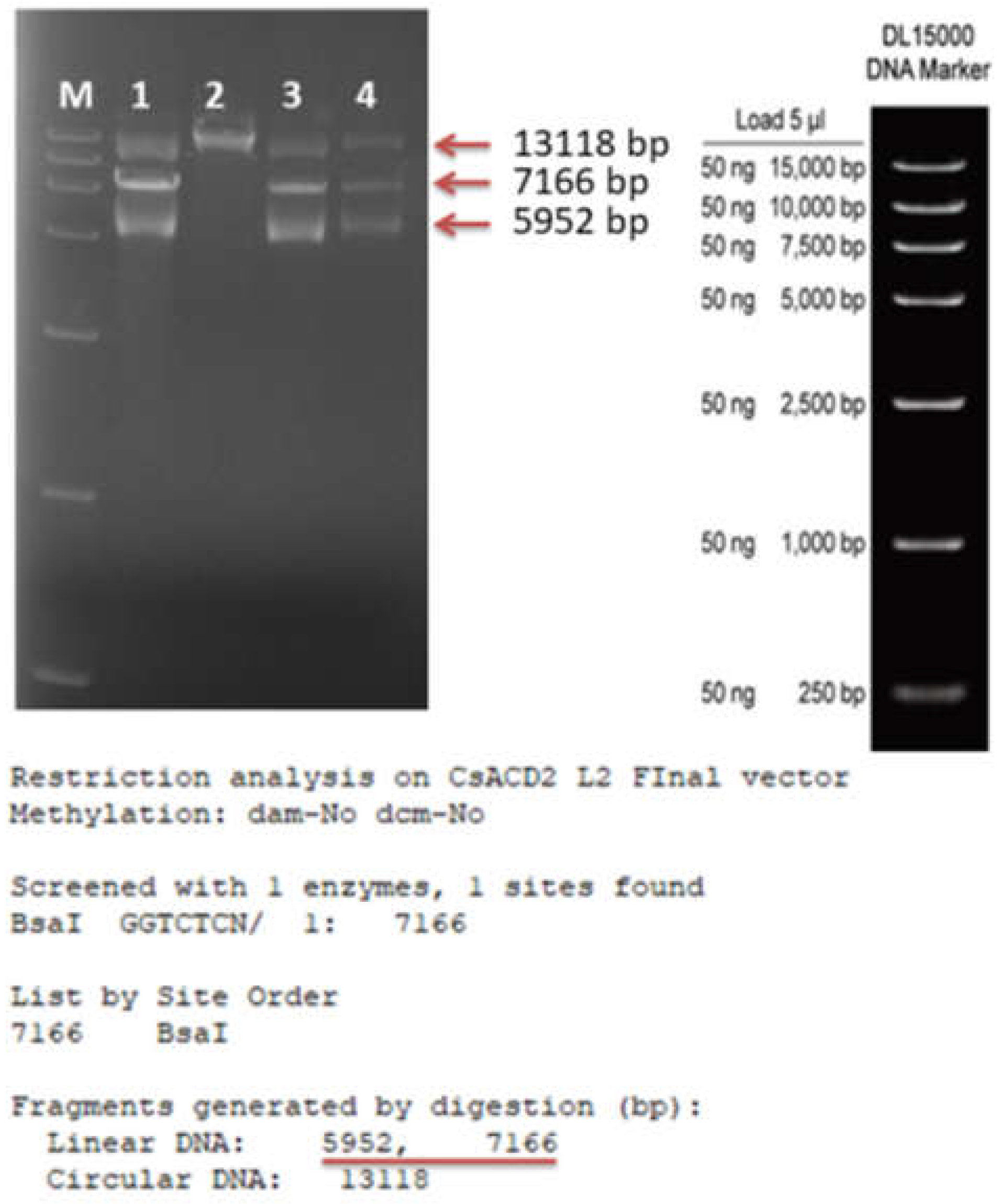{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
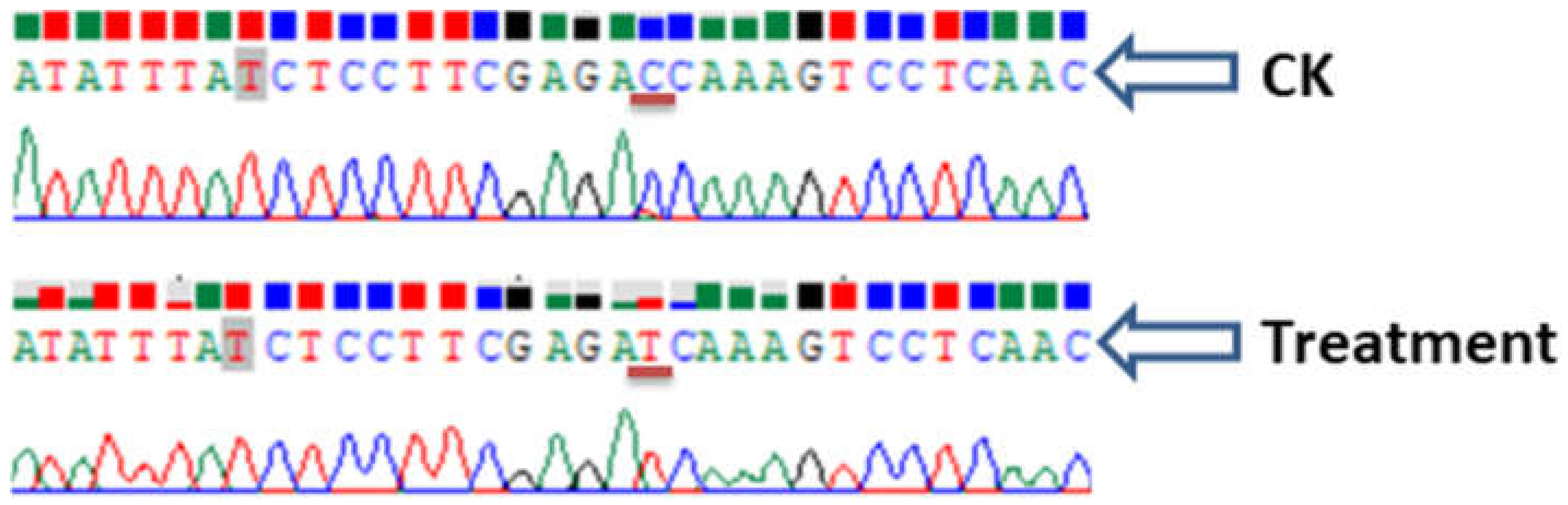
2. Results
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moniruzzaman, M.; Zhong, Y.; Yan, H.; Yuanda, L.; Jiang, B.; Zhong, G. Exploration of susceptible genes with clustered regularly interspaced short palindromic repeats–tissue-specific knockout (crispr-tsko) to enhance host resistance. Crit. Rev. Plant Sci. 2020, 39, 387–417. [Google Scholar] [CrossRef]
- Jaganathan, D.; Ramasamy, K.; Sellamuthu, G.; Jayabalan, S.; Venkataraman, G. Crispr for crop improvement: An update review. Front. Plant Sci. 2018, 9, 985. [Google Scholar] [CrossRef] [PubMed]
- Moniruzzaman, M.; Zhong, Y.; Huang, Z.; Yan, H.; Yuanda, L.; Jiang, B.; Zhong, G. Citrus cell suspension culture establishment, maintenance, efficient transformation and regeneration to complete transgenic plant. Plants 2021, 10, 664. [Google Scholar] [CrossRef] [PubMed]
- Manghwar, H.; Li, B.; Ding, X.; Hussain, A.; Lindsey, K.; Zhang, X.; Jin, S. Crispr/cas systems in genome editing: Methodologies and tools for sgrna design, off-target evaluation, and strategies to mitigate off-target effects. Adv. Sci. 2020, 7, 1902312. [Google Scholar] [CrossRef] [PubMed]
- Schindele, P.; Wolter, F.; Puchta, H. Crispr guide rna design guidelines for efficient genome editing. In RNA Tagging: Methods and Protocols; Heinlein, M., Ed.; Springer: New York, NY, USA, 2020; pp. 331–342. [Google Scholar]
- Weber, E.; Engler, C.; Gruetzner, R.; Werner, S.; Marillonnet, S. A modular cloning system for standardized assembly of multigene constructs. PLoS ONE 2011, 6, e16765. [Google Scholar] [CrossRef] [PubMed]
- Hahn, F.; Korolev, A.; Sanjurjo Loures, L.; Nekrasov, V. A modular cloning toolkit for genome editing in plants. BMC Plant Biol. 2020, 20, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, D.; O’Callaghan, C.A. Metclo: Methylase-assisted hierarchical DNA assembly using a single type iis restriction enzyme. Nucleic Acids Res. 2018, 46, e113. [Google Scholar] [CrossRef] [PubMed]
- Marillonnet, S.; Grützner, R. Synthetic DNA assembly using golden gate cloning and the hierarchical modular cloning pipeline. Curr. Protoc. Mol. Biol. 2020, 130, e115. [Google Scholar] [CrossRef] [PubMed]
- Molla, K.A.; Yang, Y. Crispr/cas-mediated base editing: Technical considerations and practical applications. Trends Biotechnol. 2019, 37, 1121–1142. [Google Scholar] [CrossRef] [PubMed]
- Monsur, M.B.; Shao, G.; Lv, Y.; Ahmad, S.; Wei, X.; Hu, P.; Tang, S. Base editing: The ever expanding clustered regularly interspaced short palindromic repeats (crispr) tool kit for precise genome editing in plants. Genes 2020, 11, 466. [Google Scholar] [CrossRef] [PubMed]
- Carson, S.; Miller, H.B.; Witherow, D.S. Lab Session 2—Purification and Digestion of Plasmid (Vector) DNA. In Molecular Biology Techniques, 3rd ed.; Carson, S., Miller, H.B., Witherow, D.S., Eds.; Academic Press: San Diego, CA, USA, 2012; pp. 11–20. [Google Scholar]
- Zhang, F.; LeBlanc, C.; Irish, V.F.; Jacob, Y. Rapid and efficient crispr/cas9 gene editing in citrus using the yao promoter. Plant Cell Rep. 2017, 36, 1883–1887. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Orbović, V.; Wang, N. Crispr-lbcas12a-mediated modification of citrus. Plant Biotechnol. J. 2019, 17, 1928–1937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Z.; Zhang, L.; Coaker, G.; Ma, W.; He, S.-Y.; Wang, N. Citrus csacd2 is a target of candidatus liberibacter asiaticus in huanglongbing disease. Plant Physiol. 2020, 184, 792–805. [Google Scholar] [CrossRef] [PubMed]
- Concordet, J.-P.; Haeussler, M. Crispor: Intuitive guide selection for crispr/cas9 genome editing experiments and screens. Nucleic Acids Res. 2018, 46, W242–W245. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moniruzzaman, M.; Zhong, Y.; Huang, Z.; Zhong, G. Having a Same Type IIS Enzyme’s Restriction Site on Guide RNA Sequence Does Not Affect Golden Gate (GG) Cloning and Subsequent CRISPR/Cas Mutagenesis. Int. J. Mol. Sci. 2022, 23, 4889. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094889
Moniruzzaman M, Zhong Y, Huang Z, Zhong G. Having a Same Type IIS Enzyme’s Restriction Site on Guide RNA Sequence Does Not Affect Golden Gate (GG) Cloning and Subsequent CRISPR/Cas Mutagenesis. International Journal of Molecular Sciences. 2022; 23(9):4889. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094889
Chicago/Turabian StyleMoniruzzaman, M., Yun Zhong, Zhifeng Huang, and Guangyan Zhong. 2022. "Having a Same Type IIS Enzyme’s Restriction Site on Guide RNA Sequence Does Not Affect Golden Gate (GG) Cloning and Subsequent CRISPR/Cas Mutagenesis" International Journal of Molecular Sciences 23, no. 9: 4889. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094889