
Direct Injection of Recombinant AAV-Containing Solution into the Oviductal Lumen of Pregnant Mice Caused In Situ Infection of Both Preimplantation Embryos and Oviductal Epithelium

 ,
,
Abstract
:1. Introduction
2. Results
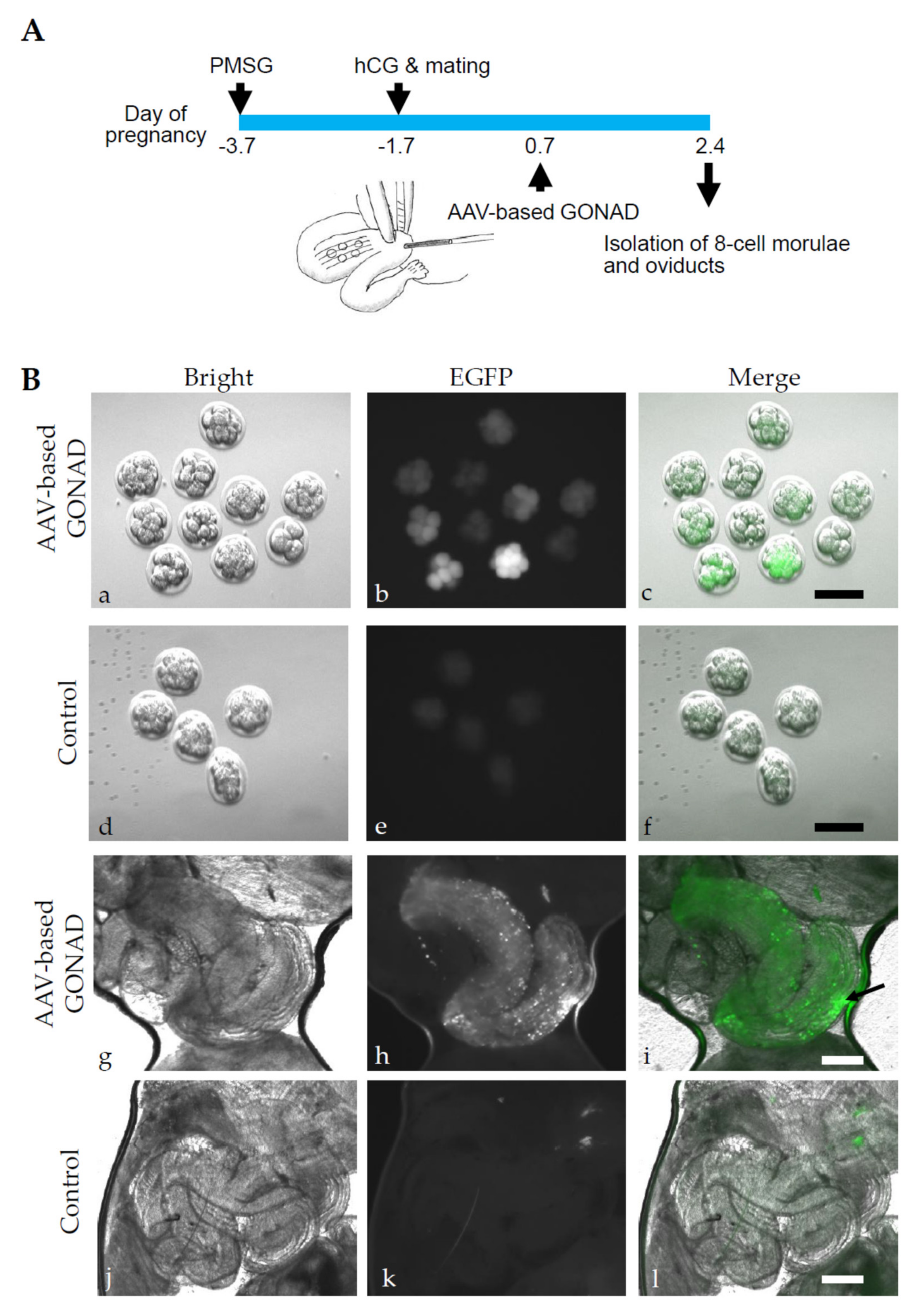
2.1. Experiment 1: AVV-Based GONAD Using Serotype 6 of rAAV
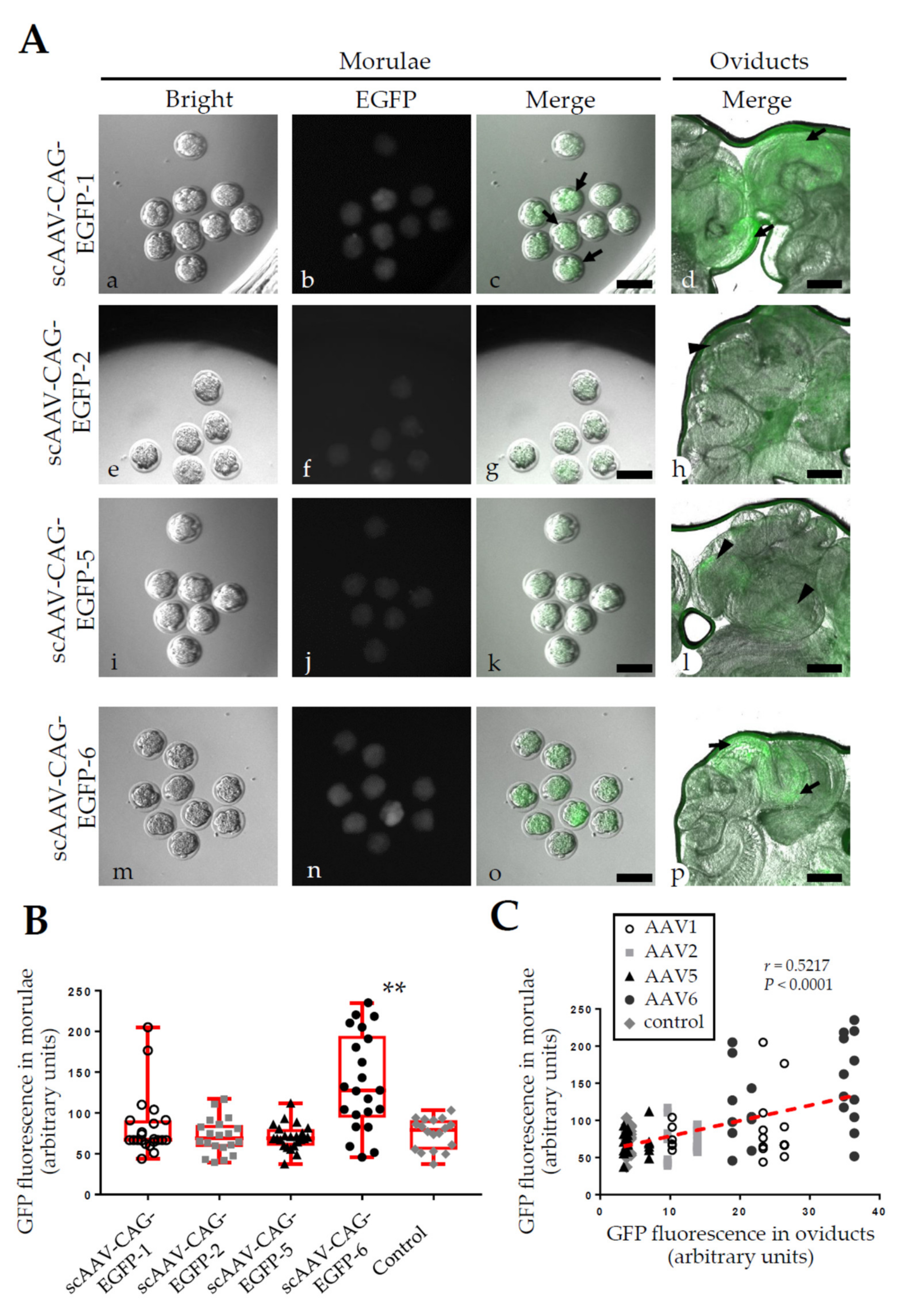
2.2. Experiment 2: AVV-Based GONAD Using Various Serotypes of rAAV
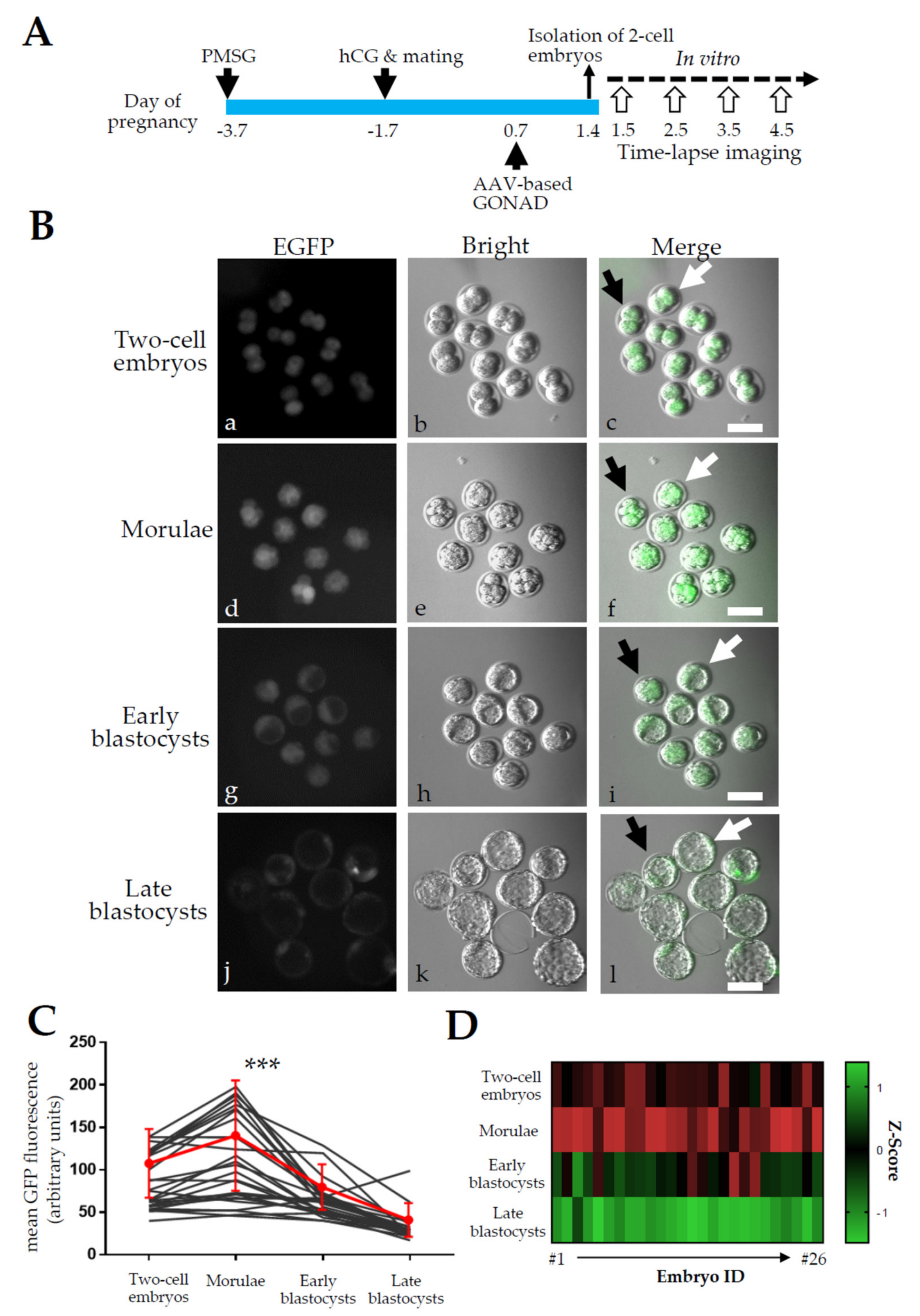
2.3. Experiment 3: Expression Analysis of EGFP in Zygotes after AVV-Based GONAD
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Mouse and Superovulation Induction
5.2. RAAV Vectors
5.3. RAAV Vector Production and Purification
5.4. Transduction of Cells with rAAVs
5.5. AAV-Based GONAD
5.6. Embryo Collection and Oviduct Processing
5.7. Observation of Fluorescence and Quantitative Evaluation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AAV | Adeno-associated virus |
| AD | Adenovirus |
| CRISPR/Cas9 | Clustered regularly interspaced short palindrome repeats/CRISPR-associated nuclease 9 |
| CMV | Cytomegalovirus |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DPBS | Dulbecco’s modified Ca2+, Mg2+-free phosphate-buffered saline |
| DSB | Double-stranded break |
| EGFP | Enhanced green fluorescent protein |
| EP | Electroporation |
| ET | Egg transfer |
| FBS | Fetal bovine serum |
| GC | Genome copies |
| GFP | Green fluorescent protein |
| GM | Genetically modified |
| GONAD | Genome-editing via Oviductal Nucleic Acid Delivery |
| gRNA | Guide RNA |
| hCG | Human chorionic gonadotrophin |
| HDR | Homology-directed repair |
| i-GONAD | Improved GONAD |
| kb | Kilobase |
| MI | Microinjection |
| MWNTs | Multiwall carbon nanotubes |
| NmeCas9 | Neisseria meningitidis Cas9 |
| PMSG | Pregnant mare serum gonadotropin |
| rAAV | Recombinant AAV |
| rAAV-6 | Serotype 6 rAAV |
| sgRNA | Single guide RNA |
| RNP | Ribonucleoprotein |
| ROIs | Regions of interest |
| RV | Retrovirus |
| scAAV | Self-complementary AAV |
| Tyr | Tyrosinase |
| WT | Wild-type |
| ZP | Zona pellucida |
References
- Harrison, M.M.; Jenkins, B.V.; O’Connor-Giles, K.M.; Wildonger, J.A. CRISPR view of development. Genes Dev. 2014, 28, 1859–1872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, P.D.; Lander, E.S.; Zhang, F. Development and applications of CRISPR-Cas9 for genome engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef] [Green Version]
- Sonoda, E.; Hochegger, H.; Saberi, A.; Taniguchi, Y.; Takeda, S. Differential usage of non-homologous end-joining and homologous recombination in double strand break repair. DNA Repair 2006, 5, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Sonntag, F.; Schmidt, K.; Kleinschmidt, J.A. A viral assembly factor promotes AAV2 capsid formation in the nucleolus. Proc. Natl. Acad. Sci. USA 2010, 107, 10220–10225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrari, F.K.; Samulski, T.; Shenk, T.; Samulski, R.J. Second-strand synthesis is a rate-limiting step for efficient transduction by recombinant adeno-associated virus vectors. J. Virol. 1996, 70, 3227–3234. [Google Scholar] [CrossRef] [Green Version]
- McCarty, D.M.; Monahan, P.E.; Samulski, R.J. Self-complementary recombinant adeno-associated virus (scAAV) vectors promote efficient transduction independently of DNA synthesis. Gene Ther. 2001, 8, 1248–1254. [Google Scholar] [CrossRef] [Green Version]
- Choi, V.W.; Samulski, R.J.; McCarty, D.M. Effects of adeno-associated virus DNA hairpin structure on recombination. J. Virol. 2005, 79, 6801–6807. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, N.; Mizutani, E.; Sato, H.; Kasai, M.; Ogawa, A.; Suchy, F.; Yamaguchi, T.; Nakauchi, H. Intra-embryo gene cassette knockin by CRISPR/Cas9-mediated genome editing with adeno-associated viral vector. iScience 2018, 9, 286–297. [Google Scholar] [CrossRef] [Green Version]
- Yoon, Y.; Wang, D.; Tai, P.W.L.; Riley, J.; Gao, G.; Rivera-Perez, J.A. Streamlined ex vivo and in vivo genome editing in mouse embryos using recombinant adeno-associated viruses. Nat. Commun. 2018, 9, 412. [Google Scholar] [CrossRef]
- Mizuno, N.; Mizutani, E.; Sato, H.; Kasai, M.; Nakauchi, H.; Yamaguchi, T. CRISPR/Cas9 + AAV-mediated intra-embryonic gene knocking in mice. Bio-Protocol 2019, 9, e3295. [Google Scholar] [CrossRef]
- Edraki, A.; Mir, A.; Ibraheim, R.; Gainetdinov, I.; Yoon, Y.; Song, C.Q.; Cao, Y.; Gallant, J.; Xue, W.; Rivera-Perez, J.A.; et al. A compact, high-accuracy Cas9 with a dinucleotide PAM for in vivo genome editing. Mol. Cell 2019, 73, 714–726.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Sun, S.; Moonen, D.; Lee, C.; Lee, A.Y.; Schaffer, D.V.; He, L. CRISPR-READI: Efficient generation of knockin mice by CRISPR RNP electroporation and AAV donor infection. Cell Rep. 2019, 27, 3780–3789.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romeo, C.; Chen, S.H.; Goulding, E.; Van Gorder, L.; Schwartz, M.; Walker, M.; Scott, G.; Scappini, E.; Ray, M.; Martin, N.P. AAV diffuses across zona pellucida for effortless gene delivery to fertilized eggs. Biochem. Biophys. Res. Commun. 2020, 526, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Niu, Y.; Ren, L.; Kang, Y.; Tai, P.W.L.; Si, C.; Mendonca, C.A.; Ma, H.; Gao, G.; Ji, W. Gene delivery to nonhuman primate preimplantation embryos using recombinant adeno-associated virus. Adv. Sci. 2019, 6, 1900440. [Google Scholar] [CrossRef]
- Takahashi, G.; Gurumurthy, C.B.; Wada, K.; Miura, H.; Sato, M.; Ohtsuka, M. GONAD: Genome-editing via Oviductal Nucleic Acids Delivery system: A novel microinjection independent genome engineering method in mice. Sci. Rep. 2015, 5, 11406. [Google Scholar] [CrossRef] [Green Version]
- Ohtsuka, M.; Sato, M.; Miura, H.; Takabayashi, S.; Matsuyama, M.; Koyano, T.; Arifin, N.; Nakamura, S.; Wada, K.; Gurumurthy, C.B. i-GONAD: A robust method for in situ germline genome engineering using CRISPR nucleases. Genome Biol. 2018, 19, 25. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.; Takabayashi, S.; Akasaka, E.; Nakamura, S. Recent advances and future perspectives of in vivo targeted delivery of genome-editing reagents to germ cells, embryos, and fetuses in mice. Cells 2020, 9, 799. [Google Scholar] [CrossRef] [Green Version]
- Sato, M. Intraoviductal introduction of plasmid DNA and subsequent electroporation for efficient in vivo gene transfer to murine oviductal epithelium. Mol. Reprod. Dev. 2005, 71, 321–330. [Google Scholar] [CrossRef]
- Wassarman, P.M.; Mortillo, S. Structure of the mouse egg extracellular coat, the zona pellucida. Int. Rev. Cytol. 1991, 130, 85–110. [Google Scholar] [CrossRef]
- Wassarman, P.M. Zona pellucida glycoproteins. J. Biol. Chem. 2008, 283, 24285–24289. [Google Scholar] [CrossRef] [Green Version]
- Gordon, J.W.; Scangos, G.A.; Plotkin, D.J.; Barbosa, J.A.; Ruddle, F.H. Genetic transformation of mouse embryos by microinjection of purified DNA. Proc. Natl. Acad. Sci. USA 1980, 77, 7380–7384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakim, B.A.; Tyagi, V.; Agnihotri, S.K.; Amar Nath, A.; Agrawal, A.K.; Jain, A.; Singh, D.; Konwar, R.; Sachdev, M. Electroporation of mouse follicles, oocytes and embryos without manipulating zona pellucida. J. Dev. Biol. 2021, 9, 13. [Google Scholar] [CrossRef] [PubMed]
- Grabarek, J.B.; Plusa, B.; Glover, D.M.; Zernicka-Goetz, M. Efficient delivery of dsRNA into zona-enclosed mouse oocytes and preimplantation embryos by electroporation. Genesis 2002, 32, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ding, T.; Brown, N.; Yamamoto, Y.; Prince, L.S.; Reese, J.; Paria, B.C. Zonula occludens-1 (ZO-1) is involved in morula to blastocyst transformation in the mouse. Dev. Biol. 2008, 318, 112–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, H.; Wu, Y.; Zhang, Y. Efficient delivery of DNA and morpholinos into mouse preimplantation embryos by electroporation. PLoS ONE 2012, 7, e43748. [Google Scholar] [CrossRef]
- Martin, N.P.; Myers, P.; Goulding, E.; Chen, S.H.; Walker, M.; Porter, T.M.; Van Gorder, L.; Mathew, A.; Gruzdev, A.; Romeo, C. En masse lentiviral gene delivery to mouse fertilized eggs via laser perforation of zona pellucida. Transgenic Res. 2018, 27, 39–49. [Google Scholar] [CrossRef]
- Lois, C.; Hong, E.J.; Pease, S.; Brown, E.J.; Baltimore, D. Germline transmission and tissue-specific expression of transgenes delivered by lentiviral vectors. Science 2002, 295, 868–872. [Google Scholar] [CrossRef] [Green Version]
- Pfeifer, A.; Ikawa, M.; Dayn, Y.; Verma, I.M. Transgenesis by lentiviral vectors: Lack of gene silencing in mammalian embryonic stem cells and preimplantation embryos. Proc. Natl. Acad. Sci. USA 2002, 99, 2140–2145. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, M.M.; Rosenkranz, A.A.; Smirnova, O.A.; Nikitin, V.A.; Sobolev, A.S.; Landa, V.; Naroditsky, B.S.; Ernst, L.K. Receptor-mediated transport of foreign DNA into preimplantation mammalian embryos. Mol. Reprod. Dev. 1999, 54, 112–120. [Google Scholar] [CrossRef]
- Joo, J.Y.; Lee, J.; Ko, H.Y.; Lee, Y.S.; Lim, D.-H.; Kim, E.-Y.; Cho, S.; Hong, K.-S.; Ko, J.J.; Lee, S.; et al. Microinjection free delivery of miRNA inhibitor into zygotes. Sci. Rep. 2014, 4, 5417. [Google Scholar] [CrossRef] [Green Version]
- Munk, M.; Ladeira, L.O.; Carvalho, B.C.; Camargo, L.S.A.; Raposo, N.R.B.; Serapião, R.V.; Quintão, C.C.R.; Silva, S.R.; Soares, J.S.; Jorio, A.; et al. Efficient delivery of DNA into bovine preimplantation embryos by multiwall carbon nanotubes. Sci. Rep. 2016, 6, 33588. [Google Scholar] [CrossRef] [PubMed]
- McCarty, D.M. Self-complementary AAV vectors; advances and applications. Mol. Ther. 2008, 16, 1648–1656. [Google Scholar] [CrossRef] [PubMed]
- Botquin, V.; Cid-Arregui, A.; Schlehofer, J.R. Adeno-associated virus type 2 interferes with early development of mouse embryos. J. Gen. Virol. 1994, 75 (Pt 10), 2655–2662. [Google Scholar] [CrossRef]
- Miller, D.J. Regulation of sperm function by oviduct fluid and the epithelium: Insight into the role of glycans. Reprod. Domest. Anim. 2015, 50 (Suppl. S2), 31–39. [Google Scholar] [CrossRef] [PubMed]
- Saint-Dizier, M.; Schoen, J.; Chen, S.; Banliat, C.; Mermillod, P. Composing the early embryonic microenvironment: Physiology and regulation of oviductal secretions. Int. J. Mol. Sci. 2020, 21, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujihara, Y.; Noda, T.; Kobayashi, K.; Oji, A.; Kobayashi, S.; Matsumura, T.; Larasati, T.; Oura, S.; Kojima-Kita, K.; Yu, Z.; et al. Identification of multiple male reproductive tract-specific proteins that regulate sperm migration through the oviduct in mice. Proc. Natl. Acad. Sci. USA 2019, 116, 18498–18506. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.F.; Chan, J.Y.C.; Lau, K.F.; Lee, W.C.; Miller, C.C.J.; Anderton, B.H.; Shaw, P.C. Molecular cloning and expression analysis of human glycogen synthase kinase-3α promoter. Mol. Brain Res. 2001, 84, 150–157. [Google Scholar] [CrossRef]
- Merlen, Y.; Bleau, G. Organization of a gene coding for an oviduct-specific glycoprotein (oviductin) in the hamster. Mol. Reprod. Dev. 2000, 57, 238–246. [Google Scholar] [CrossRef]
- Relloso, M.; Esponda, P. In vivo gene transfer to the mouse oviduct epithelium. Fertil. Steril. 1998, 70, 366–368. [Google Scholar] [CrossRef]
- Relloso, M.; Esponda, P. In-vivo transfection of the female reproductive tract epithelium. Mol. Hum. Reprod. 2000, 6, 1099–1105. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Zhang, F.; Gao, G. CRISPR-based therapeutic genome editing: Strategies and in vivo delivery by AAV vectors. Cell 2020, 181, 136–150. [Google Scholar] [CrossRef] [PubMed]
- Ibraheim, R.; Song, C.-Q.; Mir, A.; Amrani, N.; Xue, W.; Sontheimer, E.J. All-in-one adeno-associated virus delivery and genome editing by Neisseria meningitidis Cas9 in vivo. Genome Biol. 2018, 19, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanlon, K.S.; Kleinstiver, B.P.; Garcia, S.P.; Zaborowski, M.P.; Volak, A.; Spirig, S.E.; Muller, A.; Sousa, A.A.; Tsai, S.Q.; Bengtsson, N.E.; et al. High levels of AAV vector integration into CRISPR-induced DNA breaks. Nat. Commun. 2019, 10, 4439. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Ohtsuka, M.; Nakamura, S. Intraoviductal instillation of a solution as an effective route for manipulating preimplantation mammalian embryos in vivo. In New Insights into Theriogenology; Payan-Carreira, R., Ed.; InTechOpen: Rijeka, Croatia, 2018; pp. 135–150. [Google Scholar] [CrossRef] [Green Version]
- Niwa, H.; Yamamura, K.; Miyazaki, J. Efficient selection for high-expression transfectants with a novel eukaryotic vector. Gene 1991, 108, 193–199. [Google Scholar] [CrossRef]
- Sato, M.; Miyagasako, R.; Takabayashi, S.; Ohtsuka, M.; Hatada, I.; Horii, T. Sequential i-GONAD: An improved in vivo technique for CRISPR/Cas9-based genetic manipulations in mice. Cells 2020, 9, 546. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Experiment | AAV Titer (GC/mL) | Number of Animals Examined | Number of Oviducts Examined | Number of Embryos Examined |
|---|---|---|---|---|
| Experiments 1 & 2 | ||||
| scAAV-CAG-EGFP-1 | 2.1 × 1012 | 2 | 3 | 22 |
| scAAV-CAG-EGFP-2 | 3.4 × 1012 | 2 | 2 | 20 |
| scAAV-CAG-EGFP-5 | 1.9 × 1012 | 3 | 3 | 23 |
| scAAV-CAG-EGFP-6 | 3.4 × 1012 | 4 | 5 | 32 |
| Control (uninfected) | n/a | 3 | 3 | 23 |
| Experiments 3 | ||||
| scAAV-CAG-EGFP-6 | 6.0 × 1012 | 3 | 3 | 26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sato, M.; Sato-Yamamoto, N.; Wakita, A.; Haraguchi, M.; Shimonishi, M.; Okuno, H. Direct Injection of Recombinant AAV-Containing Solution into the Oviductal Lumen of Pregnant Mice Caused In Situ Infection of Both Preimplantation Embryos and Oviductal Epithelium. Int. J. Mol. Sci. 2022, 23, 4897. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094897
Sato M, Sato-Yamamoto N, Wakita A, Haraguchi M, Shimonishi M, Okuno H. Direct Injection of Recombinant AAV-Containing Solution into the Oviductal Lumen of Pregnant Mice Caused In Situ Infection of Both Preimplantation Embryos and Oviductal Epithelium. International Journal of Molecular Sciences. 2022; 23(9):4897. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094897
Chicago/Turabian StyleSato, Masahiro, Nami Sato-Yamamoto, Ai Wakita, Misako Haraguchi, Manabu Shimonishi, and Hiroyuki Okuno. 2022. "Direct Injection of Recombinant AAV-Containing Solution into the Oviductal Lumen of Pregnant Mice Caused In Situ Infection of Both Preimplantation Embryos and Oviductal Epithelium" International Journal of Molecular Sciences 23, no. 9: 4897. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094897