
Contrasting Health Effects of Bacteroidetes and Firmicutes Lies in Their Genomes: Analysis of P450s, Ferredoxins, and Secondary Metabolite Clusters

 , , , and
, , , and
Abstract
:1. Introduction
2. Results and Discussion
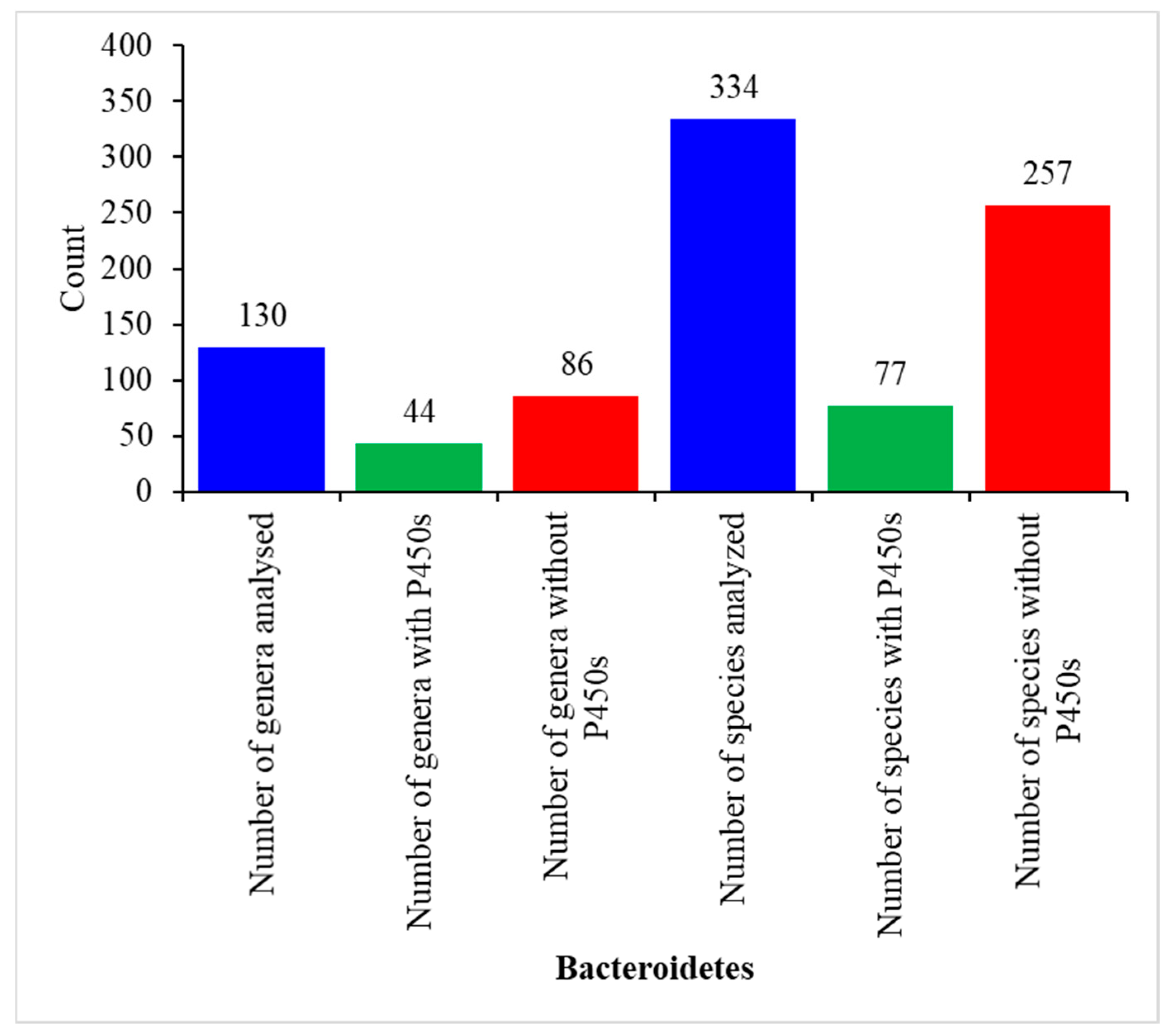
2.1. Only a Few Bacteroidetes Species Have P450s
2.2. Bacteroidetes Species Have the Highest P450 Diversity
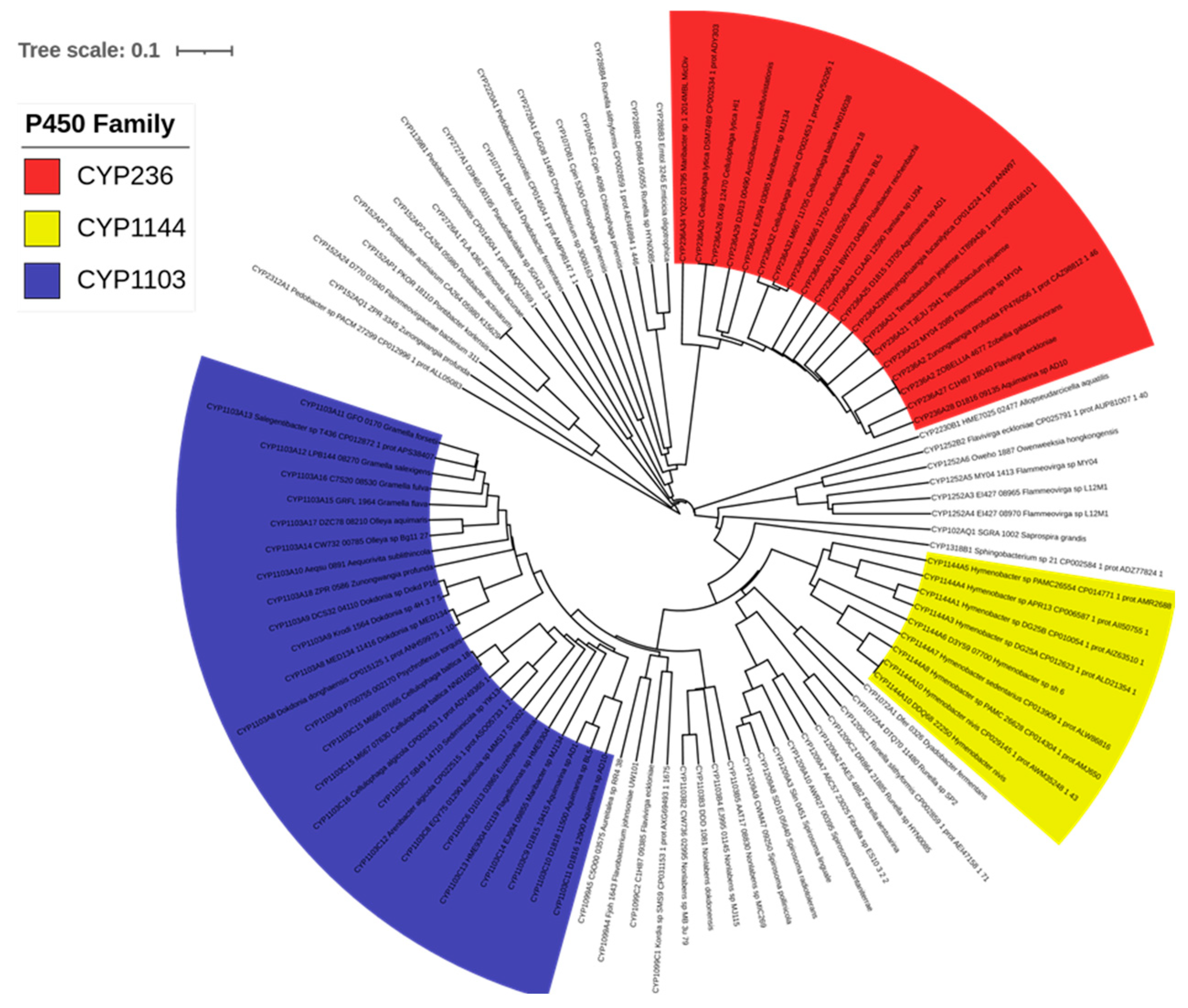
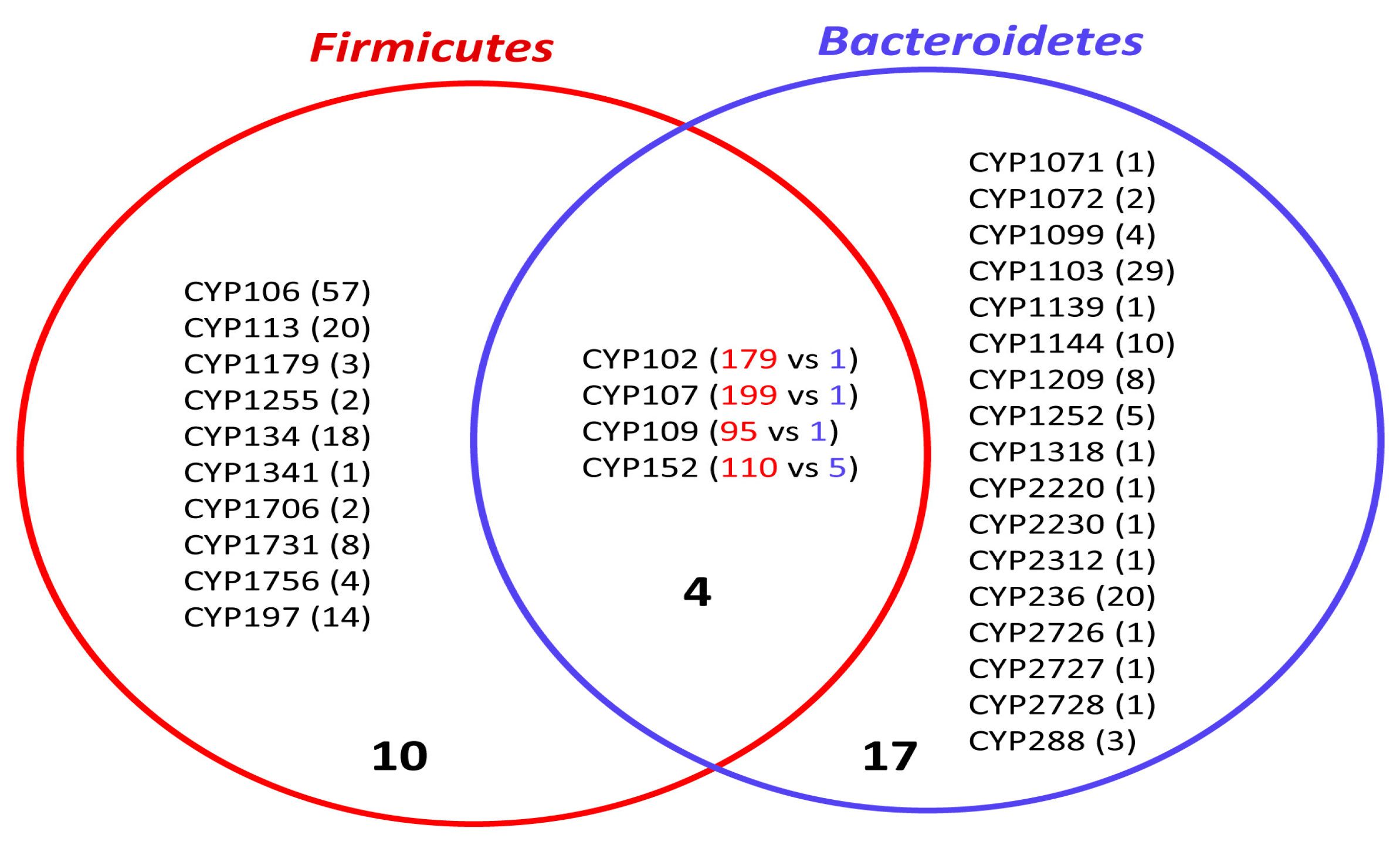
2.3. Bacteroidetes-, and Firmicutes-Species Have Diverse P450 Families in Their Genome
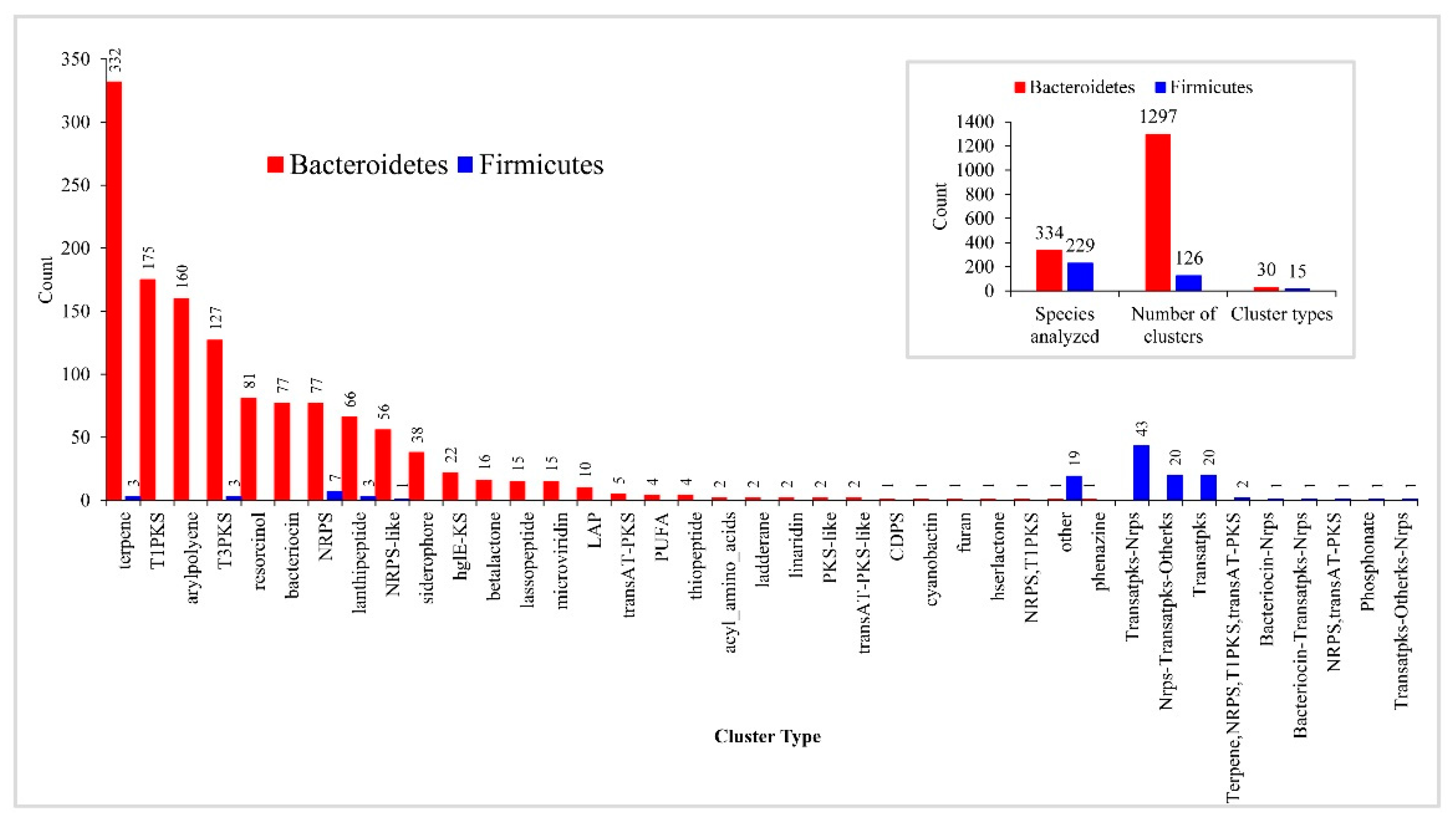
2.4. Bacteroidetes Species Have a Large and Diverse Number of Secondary Metabolite BGCs
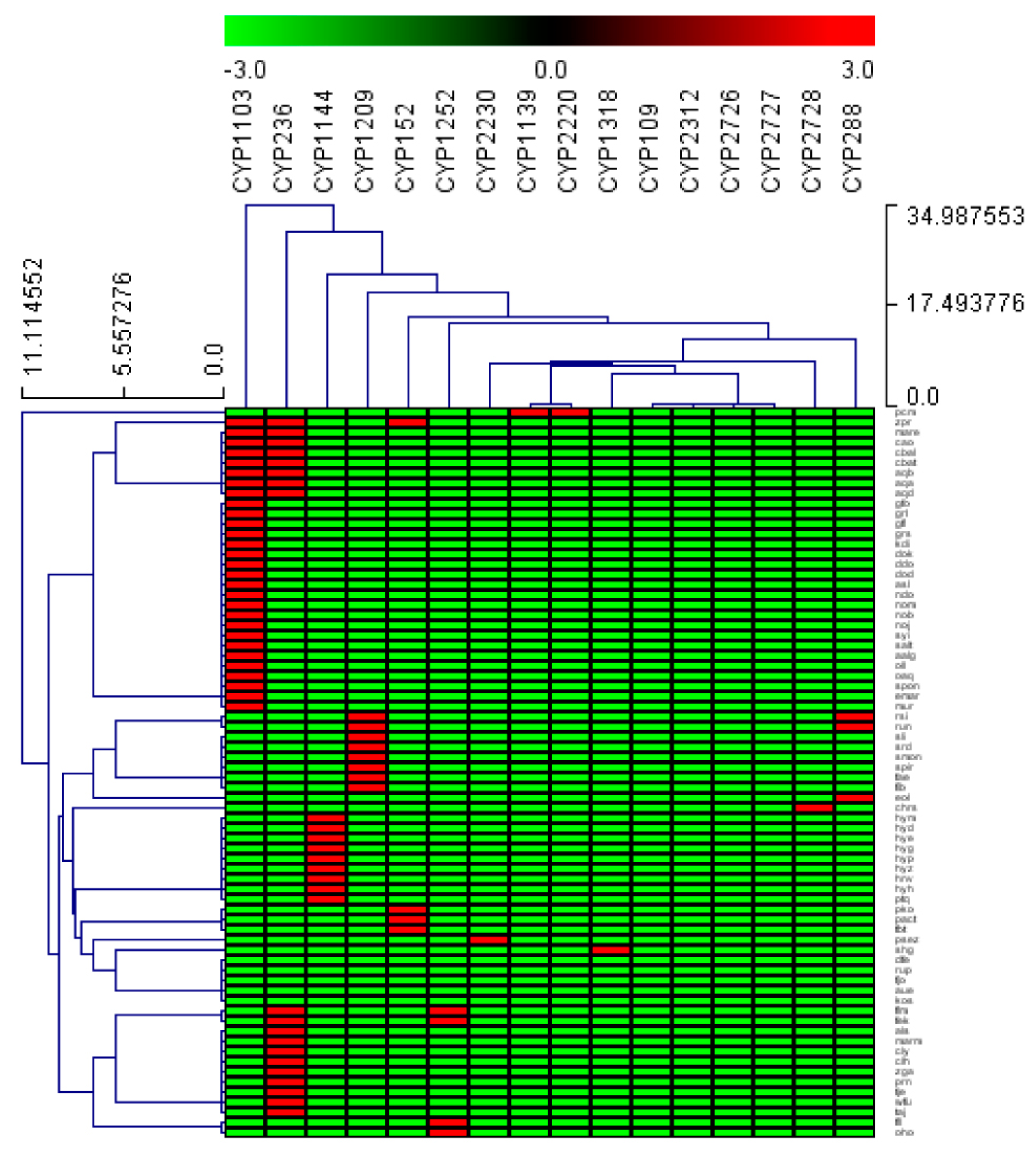
2.5. Bacteroidetes Species P450s Has a Minor Role in Secondary Metabolism
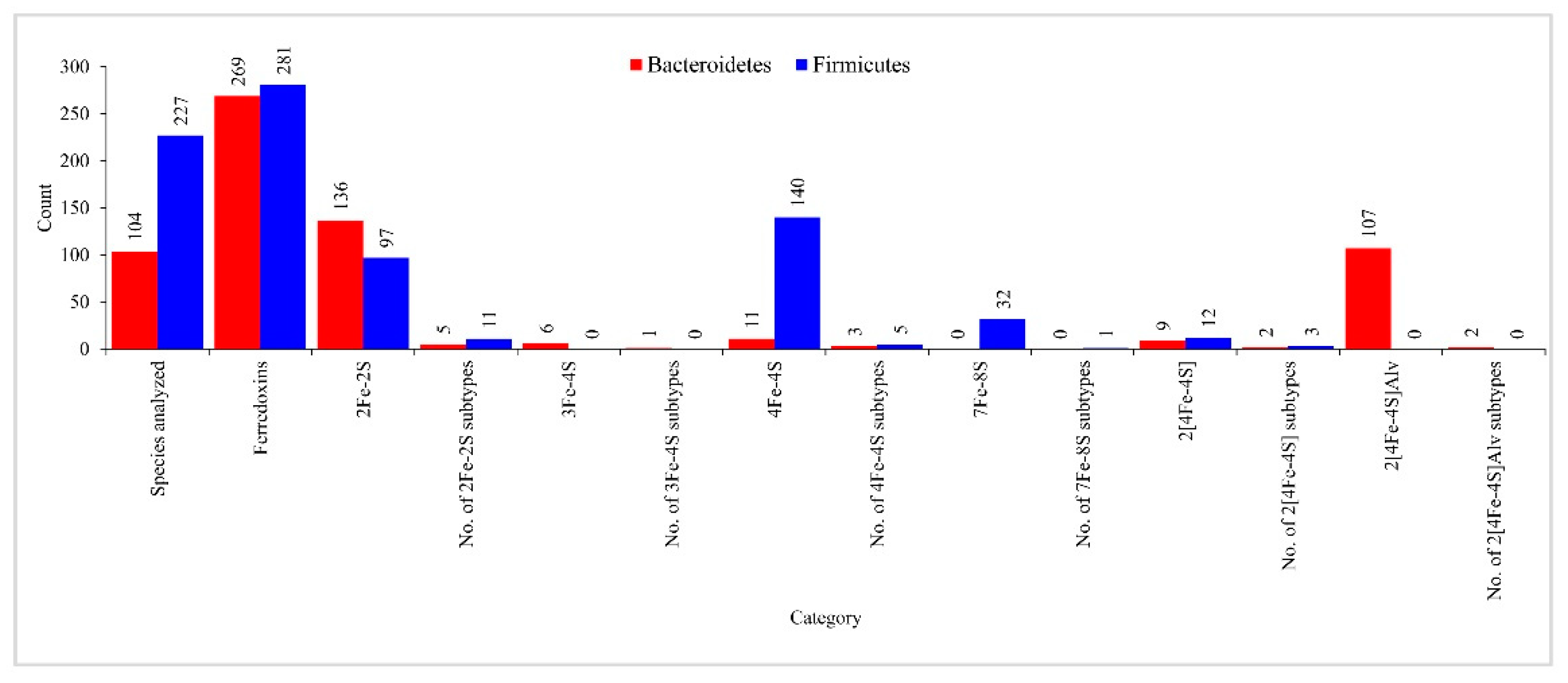
2.6. Bacteroidetes- and Firmicutes-Species Have Highly Diverse Ferredoxins in Their Genomes
3. Materials and Methods
3.1. Species and Database
3.2. Genome Data Mining and Annotation of P450s
3.3. Genome Data Mining and Annotation of Ferredoxins
3.4. Phylogenetic Analysis of P450s
3.5. Generation of P450 Profile Heat-Maps
3.6. smBGCs Analysis and P450s Identification
3.7. Data Analysis
3.8. Comparative Analysis of P450s, Ferredoxins, and smBGCs Data
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lapébie, P.; Lombard, V.; Drula, E.; Terrapon, N.; Henrissat, B. Bacteroidetes use thousands of enzyme combinations to break down glycans. Nat. Commun. 2019, 10, 2043. [Google Scholar] [CrossRef] [PubMed]
- McKee, L.S.; La Rosa, S.L.; Westereng, B.; Eijsink, V.G.; Pope, P.B.; Larsbrink, J. Polysaccharide degradation by the Bacteroidetes: Mechanisms and nomenclature. Environ. Microbiol. Rep. 2021, 13, 559–581. [Google Scholar] [CrossRef] [PubMed]
- Larsbrink, J.; McKee, L.S. Bacteroidetes bacteria in the soil: Glycan acquisition, enzyme secretion, and gliding motility. Adv. Appl. Microbiol. 2020, 110, 63–98. [Google Scholar] [PubMed]
- Johnson, E.; Heaver, S.; Walters, W.; Ley, R. Microbiome and metabolic disease: Revisiting the bacterial phylum Bacteroidetes. J. Mol. Med. 2017, 95, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.; Bai, Y.; Zha, L.; Ullah, N.; Ullah, H.; Shah, S.R.H.; Sun, H.; Zhang, C. Mechanism of the Gut Microbiota Colonization Resistance and Enteric Pathogen Infection. Front. Cell. Infect. Microbiol. 2021, 11, 1273. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Sato, Y.; Furumichi, M.; Morishima, K.; Tanabe, M. New approach for understanding genome variations in KEGG. Nucleic Acids Res. 2019, 47, D590–D595. [Google Scholar] [CrossRef] [Green Version]
- Maria Carmen, C.; Seppo, S.; Vinderola, G. Chapter 11-Postbiotics: Defining the impact of inactivated microbes and their metabolites on promotion of health. In The Human Microbiome in Early Life; Koren, O., Rautava, S., Eds.; Academic Press: Cambridge, CA, USA, 2021; pp. 257–268. [Google Scholar]
- Thomas, F.; Barbeyron, T.; Michel, G. Evaluation of reference genes for real-time quantitative PCR in the marine flavobacterium Zobellia galactanivorans. J. Microbiol. Methods 2011, 84, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Rojas, J.; Ambika Manirajan, B.; Ratering, S.; Suarez, C.; Schnell, S. Draft Genome Sequences of Spirosoma agri KCTC 52727 and Spirosoma terrae KCTC 52035. Microbiol. Resour. Announc. 2020, 9, e00317–e00320. [Google Scholar] [CrossRef] [PubMed]
- Bokhari, S.; Abbas, N.; Singh, M.; Cindrich, R.B.; Zeana, C. Empedobacter brevis bacteremia in a patient infected with HIV: Case report and review of literature. Case Rep. Infect. Dis. 2015, 2015, 813528. [Google Scholar] [PubMed] [Green Version]
- Sharma, D.; Patel, A.; Soni, P.; Sharma, P.; Gupta, B. Empedobacter brevis meningitis in a neonate: A very rare case of neonatal meningitis and literature review. Case Rep. Pediatr. 2016, 2016, 7609602. [Google Scholar] [PubMed] [Green Version]
- Shanson, D.C. Microbiology in Clinical Practice; Butterworth-Heinemann: Oxford, UK, 2014. [Google Scholar]
- Kämpfer, P.; Matthews, H.; Glaeser, S.P.; Martin, K.; Lodders, N.; Faye, I. Elizabethkingia anophelis sp. nov., isolated from the midgut of the mosquito Anopheles gambiae. Int. J. Syst. Evol. Microbiol. 2011, 61, 2670–2675. [Google Scholar] [CrossRef]
- Gershenzon, J.; Dudareva, N. The function of terpene natural products in the natural world. Nat. Chem. Biol. 2007, 3, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Belimov, A.; Hontzeas, N.; Safronova, V.; Demchinskaya, S.; Piluzza, G.; Bullitta, S.; Glick, B. Cadmium-tolerant plant growth-promoting bacteria associated with the roots of Indian mustard (Brassica juncea L. Czern.). Soil Biol. Biochem. 2005, 37, 241–250. [Google Scholar] [CrossRef]
- Katz, L.; Baltz, R.H. Natural product discovery: Past, present, and future. J. Ind. Microbiol. Biotechnol. 2016, 43, 155–176. [Google Scholar] [CrossRef]
- McBride, M.J.; Xie, G.; Martens, E.C.; Lapidus, A.; Henrissat, B.; Rhodes, R.G.; Goltsman, E.; Wang, W.; Xu, J.; Hunnicutt, D.W. Novel features of the polysaccharide-digesting gliding bacterium Flavobacterium johnsoniae as revealed by genome sequence analysis. Appl. Environ. Microbiol. 2009, 75, 6864–6875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schöner, T.A.; Fuchs, S.W.; Schönau, C.; Bode, H.B. Initiation of the flexirubin biosynthesis in Chitinophaga pinensis. Microb. Biotechnol. 2014, 7, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, D.K.; Dahal, R.H.; Kim, D.-U.; Kim, J. Flavobacterium sandaracinum sp. nov., Flavobacterium caseinilyticum sp. nov., and Flavobacterium hiemivividum sp. nov., novel psychrophilic bacteria isolated from Arctic soil. Int. J. Syst. Evol. Microbiol. 2020, 70, 2269–2280. [Google Scholar] [CrossRef]
- Walsh, C.J.; Guinane, C.M.; Hill, C.; Ross, R.P.; O’Toole, P.W.; Cotter, P.D. In silico identification of bacteriocin gene clusters in the gastrointestinal tract, based on the Human Microbiome Project’s reference genome database. BMC Microbiol. 2015, 15, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, K.; Bao, Y.; Zhao, G. Effects of dietary crude protein and tannic acid on rumen fermentation, rumen microbiota and nutrient digestion in beef cattle. Arch. Anim. Nutr. 2019, 73, 30–43. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The influence of probiotics on the firmicutes/bacteroidetes ratio in the treatment of obesity and inflammatory bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef] [PubMed]
- Manor, O.; Dai, C.L.; Kornilov, S.A.; Smith, B.; Price, N.D.; Lovejoy, J.C.; Gibbons, S.M.; Magis, A.T. Health and disease markers correlate with gut microbiome composition across thousands of people. Nat. Commun. 2020, 11, 5206. [Google Scholar] [CrossRef] [PubMed]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The firmicutes/bacteroidetes ratio: A relevant marker of gut dysbiosis in obese patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef] [PubMed]
- Mahowald, M.A.; Rey, F.E.; Seedorf, H.; Turnbaugh, P.J.; Fulton, R.S.; Wollam, A.; Shah, N.; Wang, C.; Magrini, V.; Wilson, R.K. Characterizing a model human gut microbiota composed of members of its two dominant bacterial phyla. Proc. Natl. Acad. Sci. USA 2009, 106, 5859–5864. [Google Scholar] [CrossRef] [Green Version]
- Ravcheev, D.A.; Thiele, I. Comparative genomic analysis of the human gut microbiome reveals a broad distribution of metabolic pathways for the degradation of host-synthetized mucin glycans and utilization of mucin-derived monosaccharides. Front. Genet. 2017, 8, 111. [Google Scholar] [CrossRef] [Green Version]
- Medvecky, M.; Cejkova, D.; Polansky, O.; Karasova, D.; Kubasova, T.; Cizek, A.; Rychlik, I. Whole genome sequencing and function prediction of 133 gut anaerobes isolated from chicken caecum in pure cultures. BMC Genom. 2018, 19, 561. [Google Scholar] [CrossRef] [Green Version]
- Almeida, A.; Mitchell, A.L.; Boland, M.; Forster, S.C.; Gloor, G.B.; Tarkowska, A.; Lawley, T.D.; Finn, R.D. A new genomic blueprint of the human gut microbiota. Nature 2019, 568, 499–504. [Google Scholar] [CrossRef] [Green Version]
- Padayachee, T.; Nzuza, N.; Chen, W.; Nelson, D.R.; Syed, K. Impact of lifestyle on cytochrome P450 monooxygenase repertoire is clearly evident in the bacterial phylum Firmicutes. Sci. Rep. 2020, 10, 13982. [Google Scholar] [CrossRef]
- Nzuza, N.; Padayachee, T.; Chen, W.; Gront, D.; Nelson, D.R.; Syed, K. Diversification of Ferredoxins across Living Organisms. Curr. Issues Mol. Biol. 2021, 43, 1374–1390. [Google Scholar] [CrossRef]
- Nelson, D.R. Cytochrome P450 diversity in the tree of life. Biochim. Biophys. Acta Proteins Proteom. 2018, 1866, 141–154. [Google Scholar] [CrossRef]
- Lamb, D.C.; Follmer, A.H.; Goldstone, J.V.; Nelson, D.R.; Warrilow, A.G.; Price, C.L.; True, M.Y.; Kelly, S.L.; Poulos, T.L.; Stegeman, J.J. On the occurrence of cytochrome P450 in viruses. Proc. Natl. Acad. Sci. USA 2019, 116, 12343–12352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syed, K.; Shale, K.; Pagadala, N.S.; Tuszynski, J. Systematic identification and evolutionary analysis of catalytically versatile cytochrome P450 monooxygenase families enriched in model basidiomycete fungi. PLoS ONE 2014, 9, e86683. [Google Scholar] [CrossRef] [PubMed]
- Khumalo, M.J.N.N.; Padayachee, T.; Chen, W.; Yu, J.-H.; Nelson, D.; Syed, K. Comprehensive analyses of cytochrome P450 monoxygenases and secondary metabolite biosynthetic gene clusters in Cyanobacteria. Int. J. Mol. Sci. 2020, 21, 656. [Google Scholar] [CrossRef] [Green Version]
- Kgosiemang, I.K.R.; Syed, K.; Mashele, S.S. Comparative genomics and evolutionary analysis of cytochrome P450 monooxygenases in fungal subphylum Saccharomycotina. J. Pure Appl. Microbiol. 2014, 8, 12. [Google Scholar]
- Ngwenya, M.L.; Chen, W.; Basson, A.K.; Shandu, J.S.; Yu, J.H.; Nelson, D.R.; Syed, K. Blooming of unusual cytochrome P450s by tandem duplication in the pathogenic fungus Conidiobolus coronatus. Int. J. Mol. Sci. 2018, 19, 1711. [Google Scholar] [CrossRef] [Green Version]
- Nzuza, N.; Padayachee, T.; Syed, P.R.; Kryś, J.D.; Chen, W.; Gront, D.; Nelson, D.R.; Syed, K. Ancient Bacterial Class Alphaproteobacteria Cytochrome P450 Monooxygenases Can Be Found in Other Bacterial Species. Int. J. Mol. Sci. 2021, 22, 5542. [Google Scholar] [CrossRef] [PubMed]
- Msomi, N.N.; Padayachee, T.; Nzuza, N.; Syed, P.R.; Kryś, J.D.; Chen, W.; Gront, D.; Nelson, D.R.; Syed, K. In silico analysis of P450s and their role in secondary metabolism in the bacterial class Gammaproteobacteria. Molecules 2021, 26, 1538. [Google Scholar] [CrossRef] [PubMed]
- Akapo, O.O.; Padayachee, T.; Chen, W.; Kappo, A.P.; Yu, J.H.; Nelson, D.R.; Syed, K. Distribution and Diversity of Cytochrome P450 Monooxygenases in the Fungal Class Tremellomycetes. Int. J. Mol. Sci. 2019, 20, 2889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qhanya, L.B.; Matowane, G.; Chen, W.; Sun, Y.; Letsimo, E.M.; Parvez, M.; Yu, J.H.; Mashele, S.S.; Syed, K. Genome-wide annotation and comparative analysis of cytochrome P450 monooxygenases in Basidiomycete biotrophic plant pathogens. PLoS ONE 2015, 10, e0142100. [Google Scholar]
- Malinga, N.A.; Nzuza, N.; Padayachee, T.; Syed, P.R.; Karpoormath, R.; Gront, D.; Nelson, D.R.; Syed, K. An Unprecedented Number of Cytochrome P450s Are Involved in Secondary Metabolism in Salinispora Species. Microorganisms 2022, 10, 871. [Google Scholar] [CrossRef]
- Chiliza, Z.E.; Martínez-Oyanedel, J.; Syed, K. An overview of the factors playing a role in cytochrome P450 monooxygenase and ferredoxin interactions. Biophys. Rev. 2020, 12, 1217–1222. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Du, L.; Bernhardt, R. Redox partners: Function modulators of bacterial P450 enzymes. Trends Microbiol. 2020, 28, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Podust, L.M.; Sherman, D.H. Diversity of P450 enzymes in the biosynthesis of natural products. Nat. Prod. Rep. 2012, 29, 1251–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greule, A.; Stok, J.E.; De Voss, J.J.; Cryle, M.J. Unrivalled diversity: The many roles and reactions of bacterial cytochromes P450 in secondary metabolism. Nat. Prod. Rep. 2018, 35, 757–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glowacki, R.W.; Martens, E.C. In sickness and health: Effects of gut microbial metabolites on human physiology. PLoS Pathog. 2020, 16, e1008370. [Google Scholar] [CrossRef] [Green Version]
- Sharon, G.; Garg, N.; Debelius, J.; Knight, R.; Dorrestein, P.C.; Mazmanian, S.K. Specialized metabolites from the microbiome in health and disease. Cell Metab. 2014, 20, 719–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Man, A.W.; Zhou, Y.; Xia, N.; Li, H. Involvement of gut microbiota, microbial metabolites and interaction with polyphenol in host immunometabolism. Nutrients 2020, 12, 3054. [Google Scholar] [CrossRef] [PubMed]
- Senate, L.M.; Tjatji, M.P.; Pillay, K.; Chen, W.; Zondo, N.M.; Syed, P.R.; Mnguni, F.C.; Chiliza, Z.E.; Bamal, H.D.; Karpoormath, R.; et al. Similarities, variations, and evolution of cytochrome P450s in Streptomyces versus Mycobacterium. Sci. Rep. 2019, 9, 3962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mnguni, F.C.; Padayachee, T.; Chen, W.; Gront, D.; Yu, J.-H.; Nelson, D.R.; Syed, K. More P450s are involved in secondary metabolite biosynthesis in Streptomyces compared to Bacillus, Cyanobacteria and Mycobacterium. Int. J. Mol. Sci. 2020, 21, 4814. [Google Scholar] [CrossRef] [PubMed]
- Parvez, M.; Qhanya, L.B.; Mthakathi, N.T.; Kgosiemang, I.K.; Bamal, H.D.; Pagadala, N.S.; Xie, T.; Yang, H.; Chen, H.; Theron, C.W.; et al. Molecular evolutionary dynamics of cytochrome P450 monooxygenases across kingdoms: Special focus on mycobacterial P450s. Sci. Rep. 2016, 6, 33099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, D.R.; Kamataki, T.; Waxman, D.J.; Guengerich, F.P.; Estabrook, R.W.; Feyereisen, R.; Gonzalez, F.J.; Coon, M.J.; Gunsalus, I.C.; Gotoh, O.; et al. The P450 superfamily: Update on new sequences, gene mapping, accession numbers, early trivial names of enzymes, and nomenclature. DNA Cell Biol. 1993, 12, 1–51. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R. Cytochrome P450 nomenclature, 2004. Methods Mol. Biol. 2006, 320, 1–10. [Google Scholar] [PubMed]
- Nelson, D.R. Cytochrome P450 nomenclature. Methods Mol. Biol. 1998, 107, 15–24. [Google Scholar] [PubMed]
- Blin, K.; Pascal Andreu, V.; de Los Santos, E.L.C.; Del Carratore, F.; Lee, S.Y.; Medema, M.H.; Weber, T. The antiSMASH database version 2: A comprehensive resource on secondary metabolite biosynthetic gene clusters. Nucleic Acids Res. 2019, 47, D625–D630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotoh, O. Substrate recognition sites in cytochrome P450 family 2 (CYP2) proteins inferred from comparative analyses of amino acid and coding nucleotide sequences. J. Biol. Chem. 1992, 267, 83–90. [Google Scholar] [CrossRef]
- Syed, K.; Mashele, S.S. Comparative analysis of P450 signature motifs EXXR and CXG in the large and diverse kingdom of fungi: Identification of evolutionarily conserved amino acid patterns characteristic of P450 family. PLoS ONE 2014, 9, e95616. [Google Scholar]
- Sayers, E.W.; Beck, J.; Bolton, E.E.; Bourexis, D.; Brister, J.R.; Canese, K.; Comeau, D.C.; Funk, K.; Kim, S.; Klimke, W. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2021, 49, D10. [Google Scholar] [CrossRef]
- wwPDB Consortium. Protein Data Bank: The single global archive for 3D macromolecular structure data. Nucleic Acids Res. 2019, 47, D520–D528. [Google Scholar] [CrossRef] [Green Version]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef]
- Mitchell, A.L.; Attwood, T.K.; Babbitt, P.C.; Blum, M.; Bork, P.; Bridge, A.; Brown, S.D.; Chang, H.-Y.; El-Gebali, S.; Fraser, M.I. InterPro in 2019: Improving coverage, classification and access to protein sequence annotations. Nucleic Acids Res. 2019, 47, D351–D360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, I.J.; Bennett, G.N.; Silberg, J.J. Evolutionary relationships between low potential ferredoxin and flavodoxin electron carriers. Front. Energy Res. 2019, 7, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Kuma, K.; Toh, H.; Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005, 33, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Boc, A.; Diallo, A.B.; Makarenkov, V. T-REX: A web server for inferring, validating and visualizing phylogenetic trees and networks. Nucleic Acids Res. 2012, 40, W573–W579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kryś, J.D.; Gront, D. VisuaLife: Library for interactive visualization in rich web applications. Bioinformatics 2021, 37, 3662–3663. [Google Scholar] [CrossRef] [PubMed]
- Howe, E.A.; Sinha, R.; Schlauch, D.; Quackenbush, J. RNA-Seq analysis in MeV. Bioinformatics 2011, 27, 3209–3210. [Google Scholar] [CrossRef] [Green Version]
- Cai, K.; Tonelli, M.; Frederick, R.O.; Markley, J.L. Human Mitochondrial Ferredoxin 1 (FDX1) and Ferredoxin 2 (FDX2) Both Bind Cysteine Desulfurase and Donate Electrons for Iron–Sulfur Cluster Biosynthesis. Biochemistry 2017, 56, 487–499. [Google Scholar] [CrossRef]
- Pochapsky, T.C.; Jain, N.U.; Kuti, M.; Lyons, T.A.; Heymont, J. A Refined Model for the Solution Structure of Oxidized Putidaredoxin. Biochemistry 1999, 38, 4681–4690. [Google Scholar] [CrossRef]
- Koksharova, O.A.; Klint, J.; Rasmussen, U. The first protein map of Synechococcus sp. strain PCC 7942. Microbiology 2006, 75, 664–672. [Google Scholar] [CrossRef]
- Lau, I.C.; Feyereisen, R.; Nelson, D.R.; Bell, S.G. Analysis and preliminary characterisation of the cytochrome P450 monooxygenases from Frankia sp. EuI1c (Frankia inefficax sp.). Arch. Biochem. Biophys. 2019, 669, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Child, S.A.; Bradley, J.M.; Pukala, T.L.; Svistunenko, D.A.; Le Brun, N.E.; Bell, S.G. Electron transfer ferredoxins with unusual cluster binding motifs support secondary metabolism in many bacteria. Chem. Sci. 2018, 9, 7948–7957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentley, S.D.; Chater, K.F.; Cerdeño-Tárraga, A.-M.; Challis, G.L.; Thomson, N.R.; James, K.D.; Harris, D.E.; Quail, M.A.; Kieser, H.; Harper, D.; et al. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2). Nature 2002, 417, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Ugalde, S.O.; de Koning, C.P.; Wallraven, K.; Bruyneel, B.; Vermeulen, N.P.E.; Grossmann, T.N.; Bitter, W.; Commandeur, J.N.M.; Vos, J.C. Linking cytochrome P450 enzymes from Mycobacterium tuberculosis to their cognate ferredoxin partners. Appl. Microbiol. Biotechnol. 2018, 102, 9231–9242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLean, K.J.; Warman, A.J.; Seward, H.E.; Marshall, K.R.; Girvan, H.M.; Cheesman, M.R.; Waterman, M.R.; Munro, A.W. Biophysical characterization of the sterol demethylase P450 from Mycobacterium tuberculosis, its cognate ferredoxin, and their interactions. Biochemistry 2006, 45, 8427–8443. [Google Scholar] [CrossRef] [PubMed]
- Frazão, C.; Aragão, D.; Coelho, R.; Leal, S.S.; Gomes, C.M.; Teixeira, M.; Carrondo, M.A. Crystallographic analysis of the intact metal centres [3Fe–4S] 1+/0 and [4Fe–4S] 2+/1+ in a Zn2+-containing ferredoxin. FEBS Lett. 2008, 582, 763–767. [Google Scholar] [CrossRef] [Green Version]
- Macedo-Ribeiro, S.; Martins, B.M.; Pereira, P.B.; Buse, G.; Huber, R.; Soulimane, T. New insights into the thermostability of bacterial ferredoxins: High-resolution crystal structure of the seven-iron ferredoxin from Thermus thermophilus. JBIC J. Biol. Inorg. Chem. 2001, 6, 663–674. [Google Scholar] [CrossRef]
- Saridakis, E.; Giastas, P.; Efthymiou, G.; Thoma, V.; Moulis, J.-M.; Kyritsis, P.; Mavridis, I.M. Insight into the protein and solvent contributions to the reduction potentials of [4Fe–4S] 2+/+ clusters: Crystal structures of the Allochromatium vinosum ferredoxin variants C57A and V13G and the homologous Escherichia coli ferredoxin. JBIC J. Biol. Inorg. Chem. 2009, 14, 783–799. [Google Scholar] [CrossRef]
- Dauter, Z.; Wilson, K.S.; Sieker, L.C.; Meyer, J.; Moulis, J.-M. Atomic resolution (0.94 Å) structure of Clostridium acidurici ferredoxin. Detailed geometry of [4Fe-4S] clusters in a protein. Biochemistry 1997, 36, 16065–16073. [Google Scholar] [CrossRef]
- Saeki, K.; Suetsugu, Y.; Tokuda, K.-I.; Miyatake, Y.; Young, D.; Marrs, B.; Matsubara, H. Genetic analysis of functional differences among distinct ferredoxins in Rhodobacter capsulatus. J. Biol. Chem. 1991, 266, 12889–12895. [Google Scholar] [CrossRef]
- Moulis, J.M.; Sieker, L.C.; Wilson, K.S.; Dauter, Z. Crystal structure of the 2 [4Fe-4S] ferredoxin from Chromatium vinosum: Evolutionary and mechanistic inferences for [3/4Fe-4S] ferredoxins. Protein Sci. 1996, 5, 1765–1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unciuleac, M.; Boll, M.; Warkentin, E.; Ermler, U. Crystallization of 4-hydroxybenzoyl-CoA reductase and the structure of its electron donor ferredoxin. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, 60, 388–391. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species (Source) | Secondary Metabolite | Biological Function | References |
|---|---|---|---|
| Spirosoma spp. and Sphingobacterium sp. 21 | Terpene | Neuroprotective, anti-tumorigenic, and anti-inflammatory | [9,14] |
| Flavobacterium spp. | Siderophore | Iron scavenger and transporter | [15] |
| Spirosoma spp.; Pedobacter cryoconitis; Sphingobacterium sp. 21 and Chitinophaga pinesis | T1PKS | Useful as agrochemical and pharmaceutical compounds | [9,16] |
| Chinophaga pinesis; Flavobacterium johnsoniae UW10 | Arylpolyene | Shields bacteria from reactive oxidation | [17,18] |
| Spirosoma spp.; Flavobacterium spp. | T3PKS | Involved in hypocrellin and kanamycin synthesis | [9,19] |
| Spirosoma spp.; Bacteroides spp. | Bacteriocin | Anti-bacterial activity | [9,20] |
| Prevotella spp. | Resorcinol | Used to treat skin disorders | [21] |
| Spirosoma spp. | NRPs | Antibiotic, immunosuppressant, and cytotoxic properties | [9] |
| Spirosoma spp. | Lanthipeptide | Antimicrobial, antifungal, and antiviral activities | [9] |
| Bacteroidetes Species | Firmicutes Species | Gammaproteobacterial Species | Streptomyces Species | Mycobacterial Species | Cyanobacterial Species | |
|---|---|---|---|---|---|---|
| Total no. of species analyzed | 334 | 972 | 1 261 | 203 | 60 | 114 |
| No. of species with P450s | 77 | 229 | 169 | 203 | 60 | 114 |
| No. of P450s | 98 | 712 | 277 | 5460 | 1784 | 341 |
| No. of families | 21 | 14 | 84 | 253 | 77 | 36 |
| No. of subfamilies | 28 | 53 | 105 | 698 | 132 | 79 |
| Dominant P450 family | CYP1103 | CYP107 | CYP133 | CYP107 | CYP125 | CYP110 |
| Average no. of P450s | 1 | 3 | 0.2 | 27 | 30 | 3 |
| P450 diversity percentage | 0.28 | 0.01 | 0.18 | 0.02 | 0.07 | 0.09 |
| No. of P450s part of BGCs | 8 | 126 | 49 | 1231 | 204 | 27 |
| Percentage of P450s part of BGCs | 8 | 18 | 18 | 23 | 11 | 8 |
| Reference(s) | This work | [30] | [39] | [50,51] | [50,52] | [35] |
| P450 Family | Count | Percentage | P450 Subfamily | Count |
|---|---|---|---|---|
| CYP1103 | 29 | 30 | A | 13 |
| B | 4 | |||
| C | 12 | |||
| CYP236 | 20 | 20 | A | 20 |
| CYP1144 | 10 | 10 | A | 10 |
| CYP1209 | 8 | 8 | A | 6 |
| C | 2 | |||
| CYP1252 | 5 | 5 | A | 4 |
| B | 1 | |||
| CYP152 | 5 | 5 | A | 1 |
| A.P. | 3 | |||
| AQ | 1 | |||
| CYP1099 | 4 | 4 | A | 2 |
| C | 2 | |||
| CYP288 | 3 | 3 | B | 3 |
| CYP1072 | 2 | 2 | A | 2 |
| CYP102 | 1 | 1 | AQ | 1 |
| CYP1071 | 1 | 1 | A | 1 |
| CYP107 | 1 | 1 | DB | 1 |
| CYP109 | 1 | 1 | AE | 1 |
| CYP1139 | 1 | 1 | B | 1 |
| CYP1318 | 1 | 1 | B | 1 |
| CYP2220 | 1 | 1 | A | 1 |
| CYP2230 | 1 | 1 | B | 1 |
| CYP2312 | 1 | 1 | A | 1 |
| CYP2726 | 1 | 1 | A | 1 |
| CYP2727 | 1 | 1 | A | 1 |
| CYP2728 | 1 | 1 | A | 1 |
| Species Name | P450 | Cluster Type |
|---|---|---|
| Chitinophaga pinensis | CYP109 | terpene |
| CYP107 | NRPS | |
| Pedobacter cryoconitis | CYP1139 | lanthipeptide |
| Sphingobacterium sp. 21 | CYP1318 | NRPS-like |
| Spirosoma linguale | CYP1209 | terpene |
| Spirosoma radiotolerans | CYP1209 | terpene |
| Spirosoma montaniterrae | CYP1209 | terpene |
| Spirosoma pollinicola | CYP1209 | terpene |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nkosi, B.V.Z.; Padayachee, T.; Gront, D.; Nelson, D.R.; Syed, K. Contrasting Health Effects of Bacteroidetes and Firmicutes Lies in Their Genomes: Analysis of P450s, Ferredoxins, and Secondary Metabolite Clusters. Int. J. Mol. Sci. 2022, 23, 5057. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095057
Nkosi BVZ, Padayachee T, Gront D, Nelson DR, Syed K. Contrasting Health Effects of Bacteroidetes and Firmicutes Lies in Their Genomes: Analysis of P450s, Ferredoxins, and Secondary Metabolite Clusters. International Journal of Molecular Sciences. 2022; 23(9):5057. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095057
Chicago/Turabian StyleNkosi, Bridget Valeria Zinhle, Tiara Padayachee, Dominik Gront, David R. Nelson, and Khajamohiddin Syed. 2022. "Contrasting Health Effects of Bacteroidetes and Firmicutes Lies in Their Genomes: Analysis of P450s, Ferredoxins, and Secondary Metabolite Clusters" International Journal of Molecular Sciences 23, no. 9: 5057. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095057