
Function Analysis of the PR55/B Gene Related to Self-Incompatibility in Chinese Cabbage Using CRISPR/Cas9


Abstract
:1. Introduction
2. Results
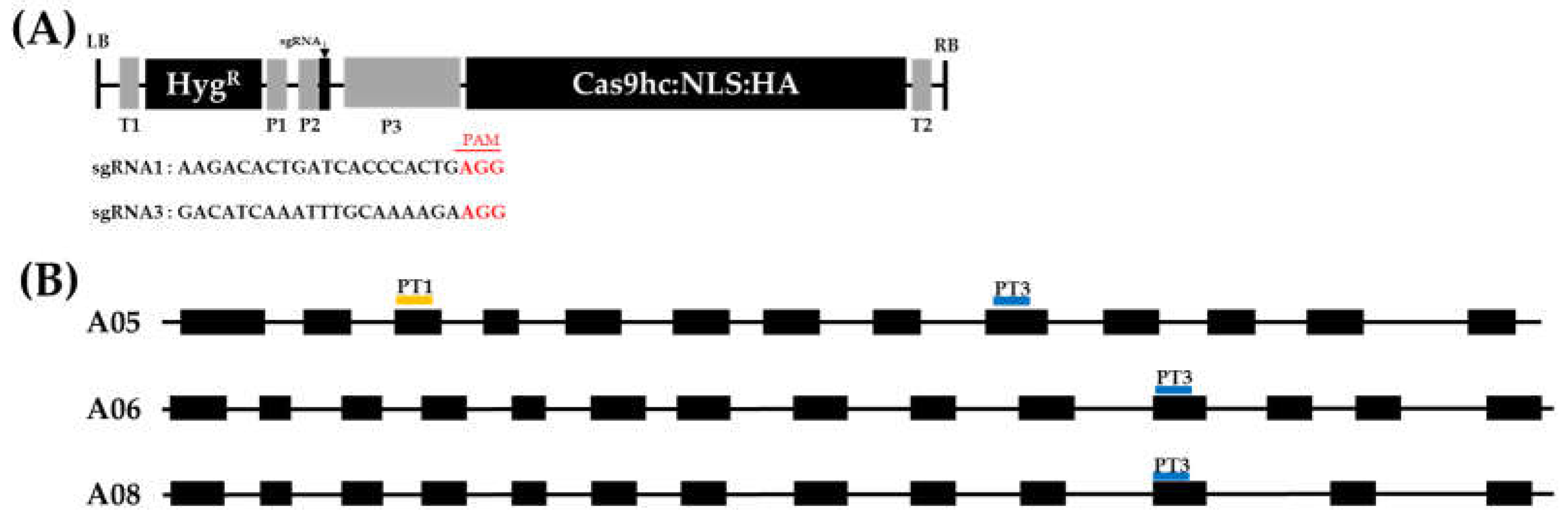
2.1. Vector Construction and Agrobacterium-Mediated Transformation of Chinese Cabbage
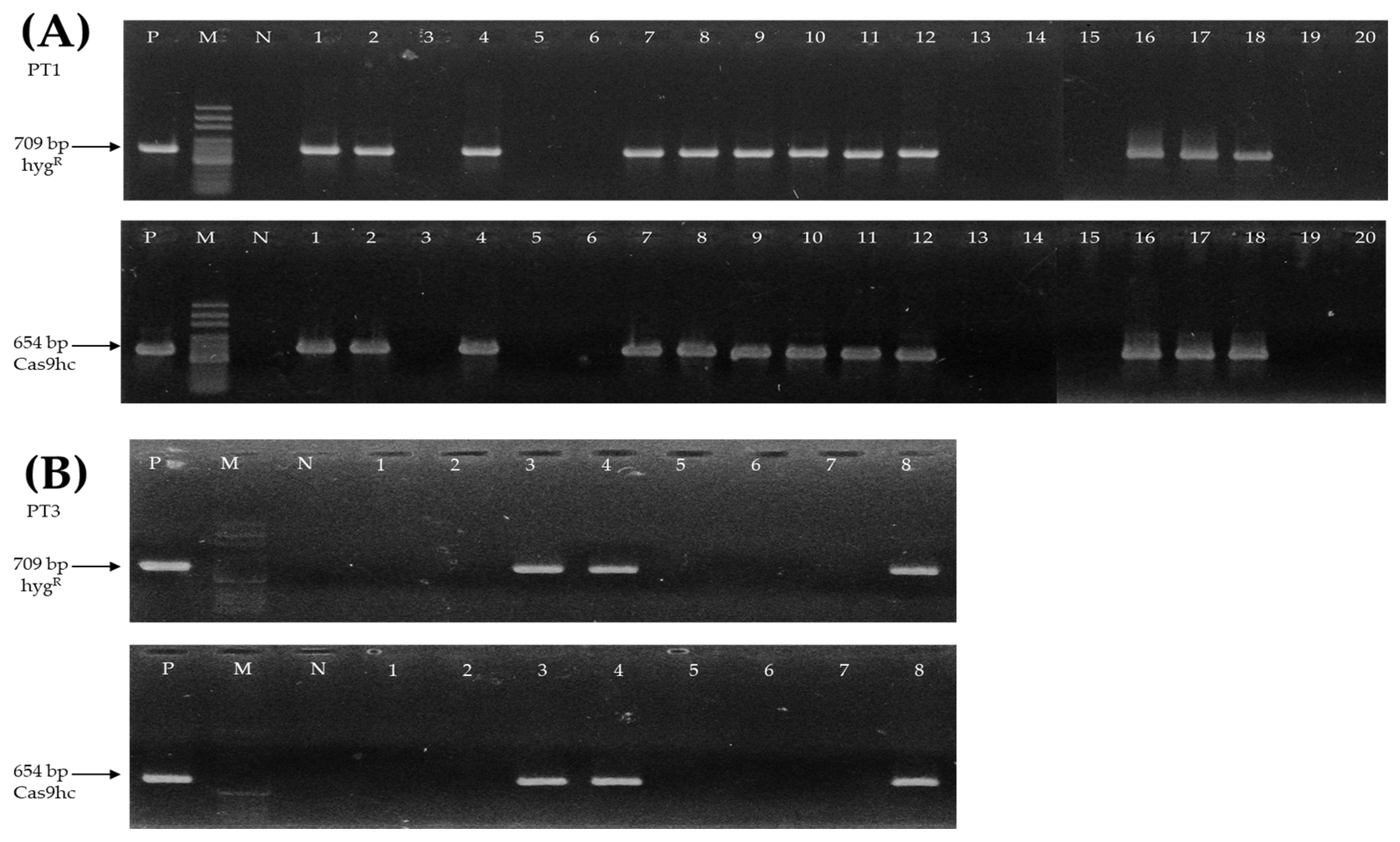
2.2. Selection and Fertility Analysis of T0 Gene-Edited Chinese Cabbage Lines
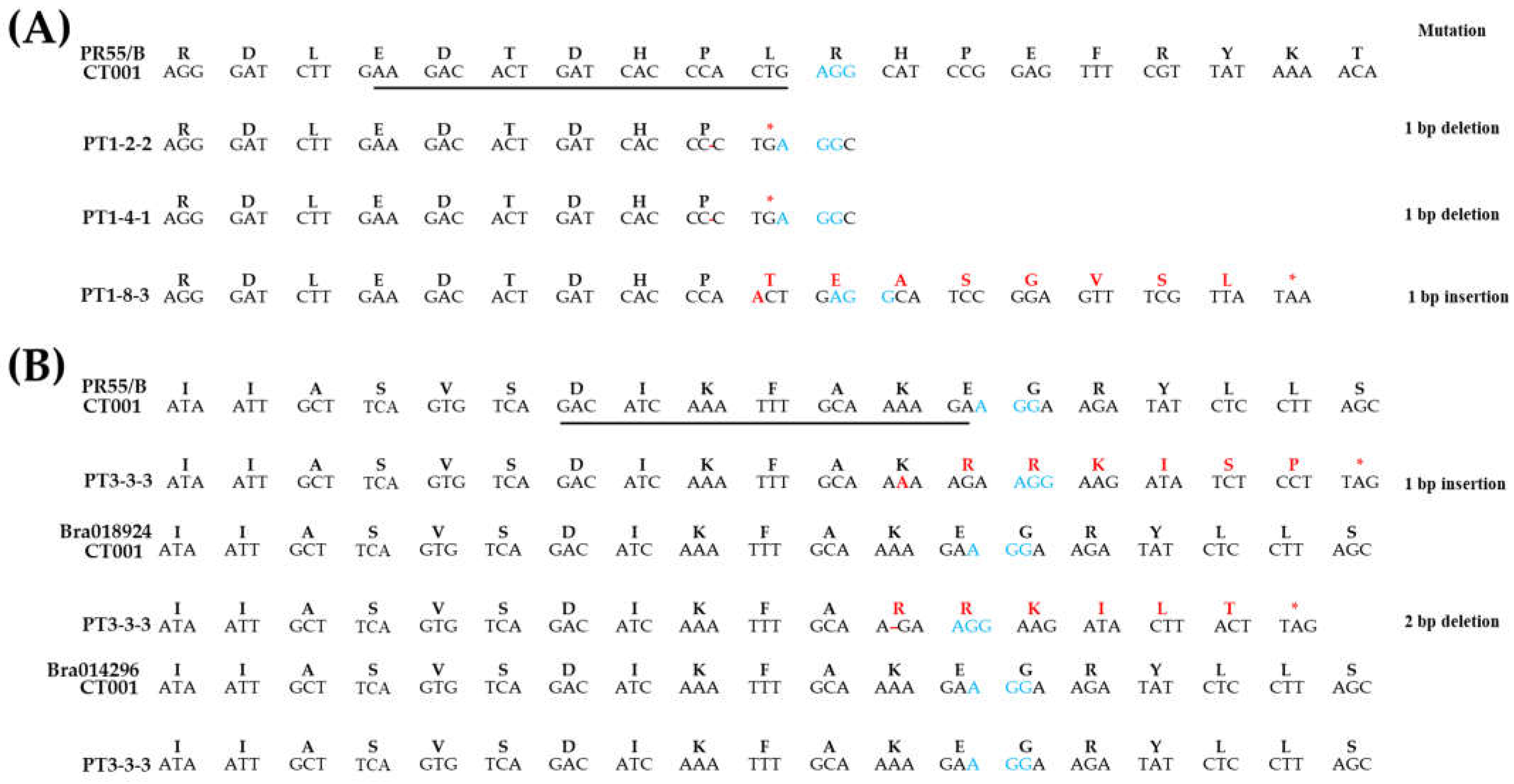
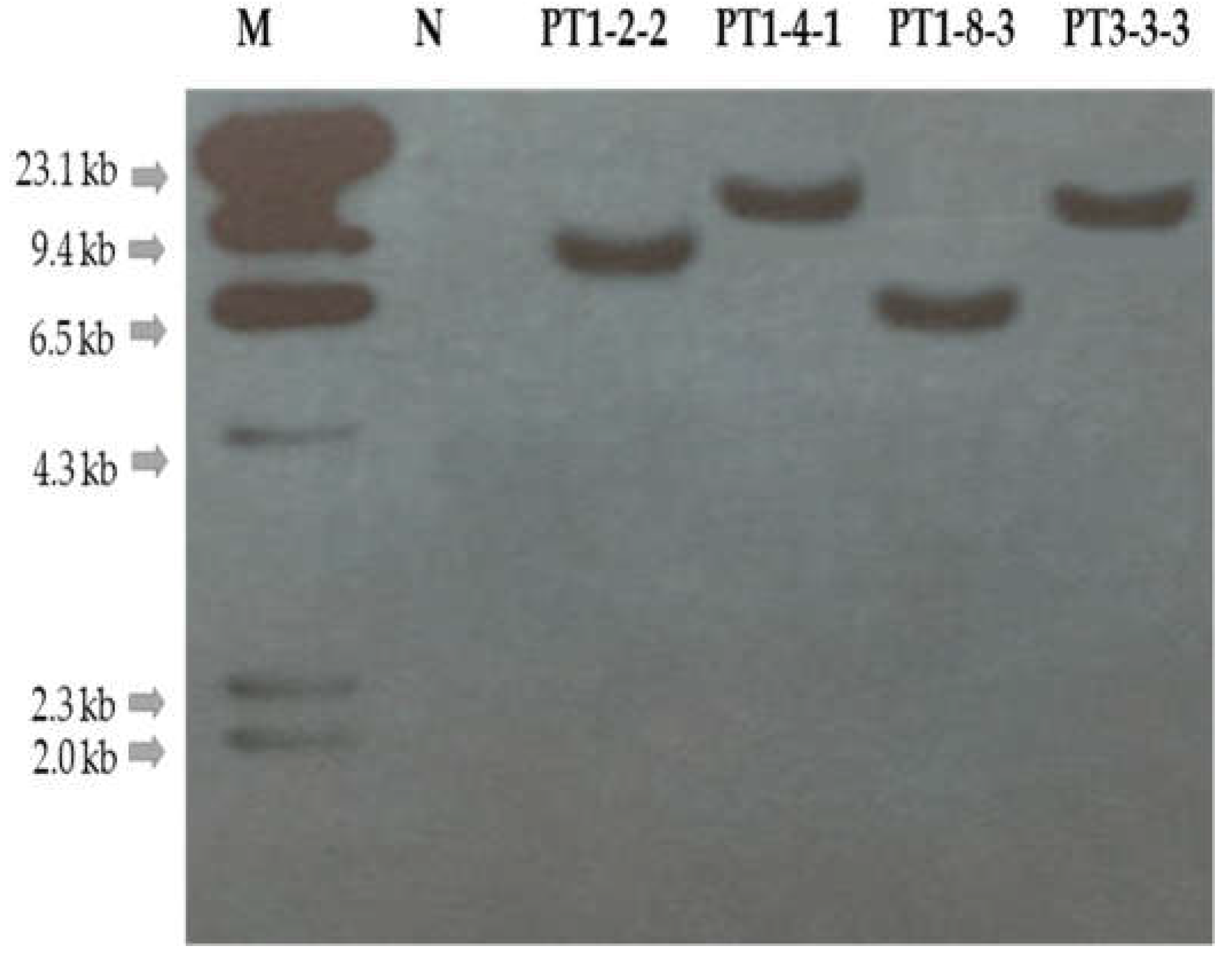
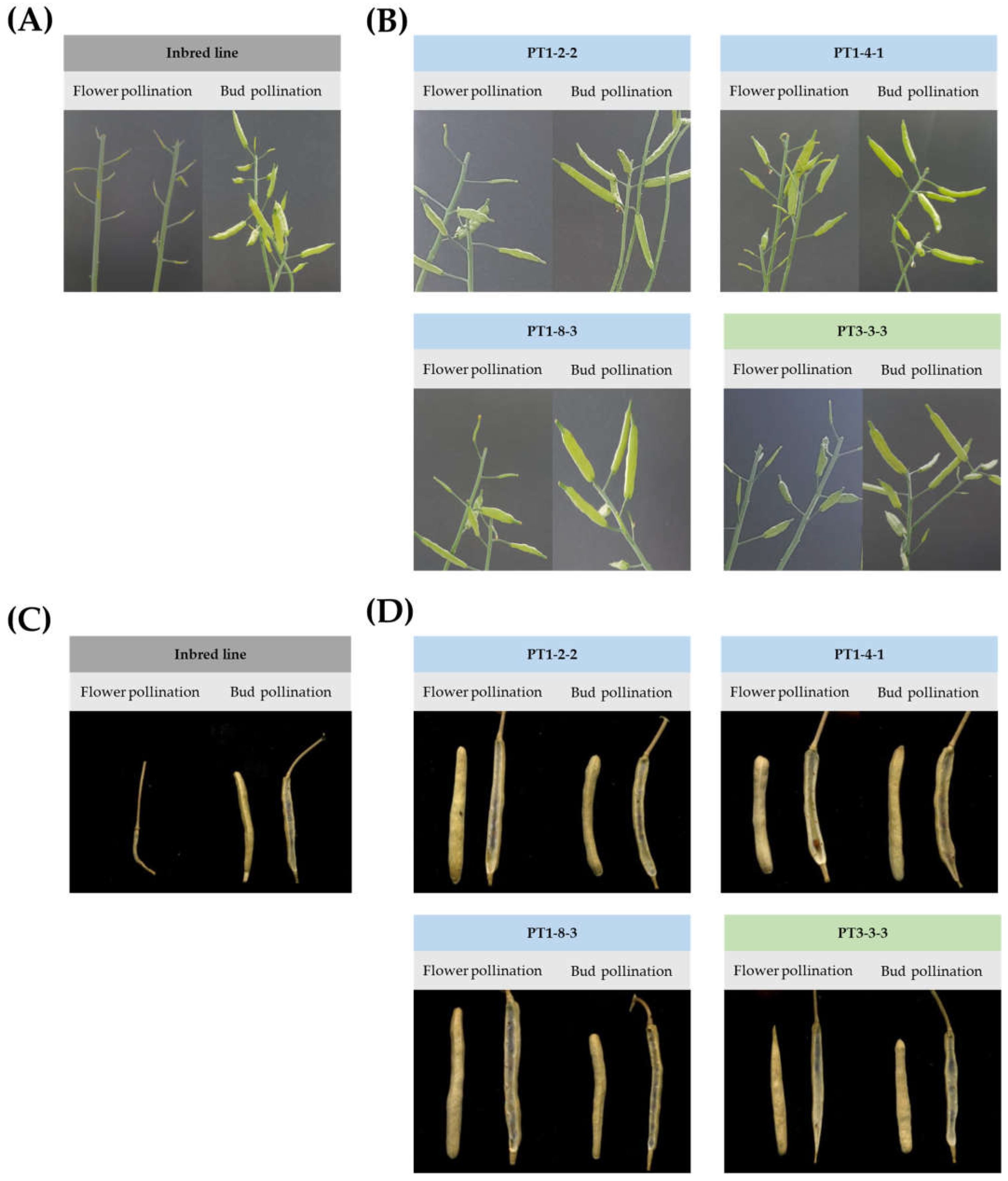
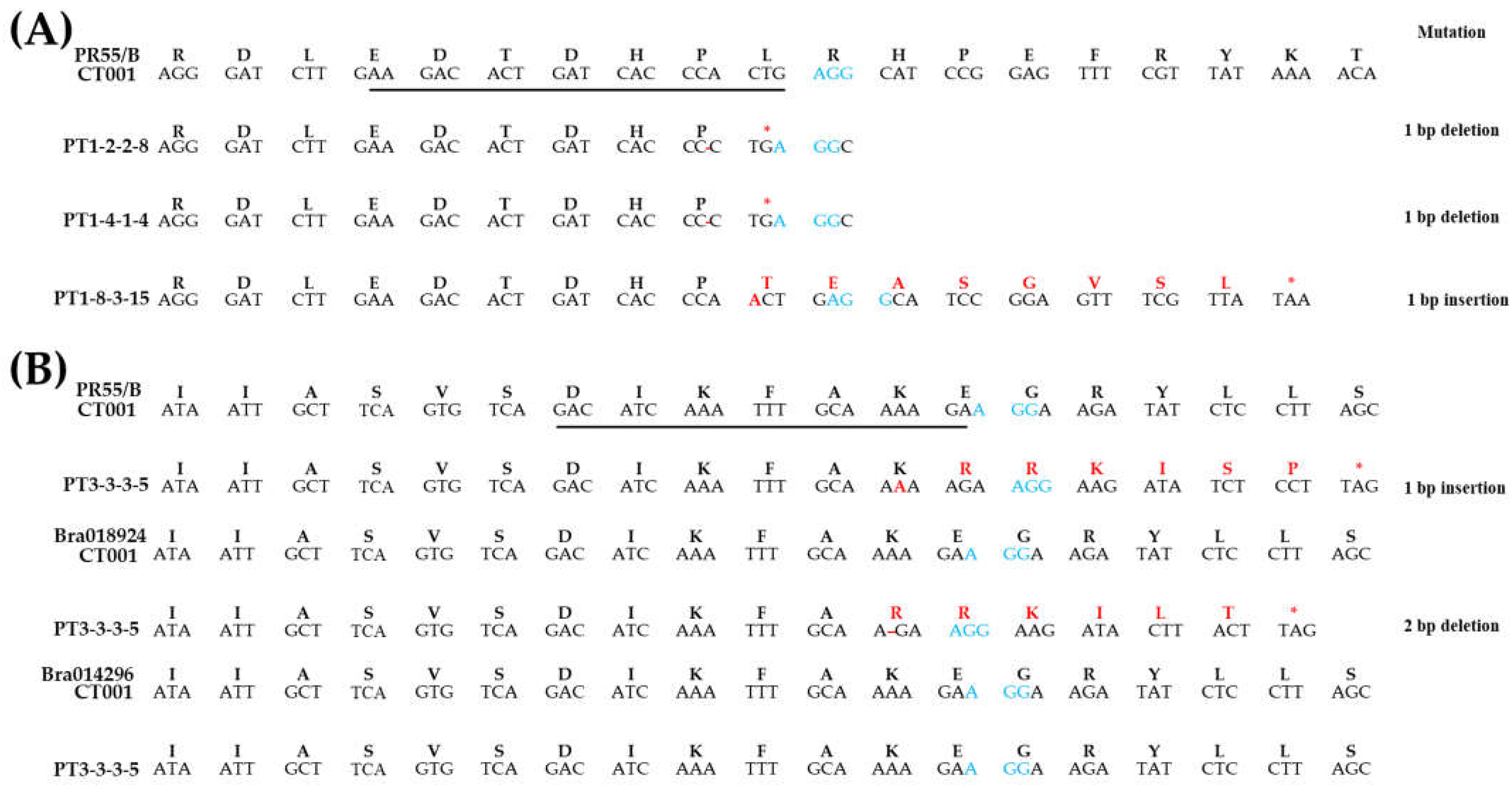
2.3. Confirmation of Sequence Change and Fertility Analysis in T1 Gene-Edited Chinese Cabbage Lines
2.4. Inheritance of Sequence Change in T-DNA-Free T2 Gene-Edited Chinese Cabbage Lines
3. Discussion
4. Materials and Methods
4.1. Target Site Selection and Construction of PR55/B Gene-Editing Vectors
4.2. PCR and RT-PCR Analysis of Gene-Edited Chinese Cabbage Lines
4.3. Southern Hybridization Analysis for Identifying the Number of T-DNA Copies
4.4. Self-Compatibility Analysis of Gene-Edited Chinese Cabbage Lines
4.5. Sequence Change Analysis of Gene-Edited Chinese Cabbage Lines
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Takayama, S.; Isogai, A. Self-incompatibility in plants. Annu. Rev. Plant Biol. 2005, 56, 467–489. [Google Scholar] [CrossRef] [Green Version]
- Hiscock, S.J.; Allen, A.M. Diverse cell signalling pathways regulate pollen-stigma interactions: The search for consensus. New Phytol. 2008, 179, 286–317. [Google Scholar] [CrossRef]
- Igic, B.; Lande, R.; Kohn, J.R. Loss of self-incompatibility and its evolutionary consequences. Int. J. Plant Sci. 2008, 169, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Vanoosthuyse, V.; Tichtinsky, G.; Dumas, C.; Gaude, T.; Cock, J.M. Interaction of calmodulin, a sorting nexin and kinase-associated protein phosphatase with the Brassica oleracea S locus receptor kinase. Plant Physiol. 2003, 133, 919–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehgal, N.; Singh, S. Progress on deciphering the molecular aspects of cell-to-cell communication in Brassica self-incompatibility response. 3 Biotech 2018, 8, 1–17. [Google Scholar] [CrossRef]
- Janssens, V.; Goris, J. Protein phosphatase 2A: A highly regulated family of serine/threonine phosphatases implicated in cell growth and signalling. Biochem. J. 2001, 353, 417–439. [Google Scholar] [CrossRef] [PubMed]
- Rundle, S.J.; Nasrallah, M.E.; Nasrallah, J.B. Effects of inhibitors of protein serine/threonine phosphatases on pollination in Brassica. Plant Physiol. 1993, 103, 1165–1171. [Google Scholar] [CrossRef] [Green Version]
- Ku, J.J.; Park, Y.H.; Park, Y.D. A non-antibiotic selection system uses the phosphomannose-isomerase (PMI) gene for Agrobacterium-mediated transformation of Chinese cabbage. J. Plant Biol. 2006, 49, 115–122. [Google Scholar] [CrossRef]
- Yu, J.G.; Lee, G.H.; Kim, J.S.; Shim, E.J.; Park, Y.D. An insertional mutagenesis system for analyzing the Chinese cabbage genome using Agrobacterium T-DNA. Mol. Cells. 2010, 29, 267–275. [Google Scholar] [CrossRef]
- Singh, V.K.; Khan, A.W.; Saxena, R.K.; Sinha, P.; Kale, S.M.; Parupalli, S.; Kumar, V.; Chitikineni, A.; Vechalapu, S.; Kumar, C.V.S.; et al. Indel-seq: A fast-forward genetics approach for identification of trait-associated putative candidate genomic regions and its application in pigeonpea (Cajanus cajan). Plant Biotechnol. J. 2017, 15, 906–914. [Google Scholar] [CrossRef] [Green Version]
- Gajewska, P.; Janiak, A.; Kwasniewski, M.; Kędziorski, P.; Szarejko, I. Forward genetics approach reveals a mutation in bHLH transcription factor-encoding gene as the best candidate for the root hairless phenotype in barley. Front. Plant Sci. 2018, 9, 1229. [Google Scholar] [CrossRef]
- Gilchrist, E.; Haughn, G. Reverse genetics techniques: Engineering loss and gain of gene function in plants. Brief. Funct. Genom. 2010, 9, 103–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloyd, J.; Meinke, D. A comprehensive dataset of genes with a loss-of-function mutant phenotype in Arabidopsis. Plant Physiol. 2012, 158, 1115–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, S.; Park, S.; Jeong, D.H.; Lee, D.Y.; Kang, H.G.; Yu, J.H.; Hur, J.H.; Kim, S.R.; Kim, Y.H.; Lee, M.O.; et al. Generation and analysis of end sequence database for T-DNA tagging lines in rice. Plant Physiol. 2003, 133, 2040–2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.G.; Lee, G.H.; Park, Y.D. Comparison of RNA interference-mediated gene silencing and T-DNA integration techniques for gene function analysis in Chinese cabbage. Hortic. Sci. Technol. 2012, 30, 734–742. [Google Scholar]
- Jinek, M.; Chylinski, K.; Ronfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E.A. programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Jinek, M.; Jiang, F.; Taylor, D.W.; Sternberg, S.H.; Kaya, E.; Ma, E.; Anders, C.; Hauer, M.; Zhou, K.; Lin, S.; et al. Structures of Cas9 endonucleases reveal RNA-mediated conformational activation. Science 2014, 343, 1247997. [Google Scholar] [CrossRef] [Green Version]
- Nishimasu, H.; Ran, F.A.; Hsu, P.D.; Konermann, S.; Shelata, S.I.; Dohmae, N.; Ishitani, R.; Zhang, F.; Nureki, O. Crystal structure of Cas9 in complex with guide RNA and target DNA. Cell 2014, 156, 935–949. [Google Scholar] [CrossRef] [Green Version]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex genome engineering using CRISPR/Cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [Green Version]
- Mali, P.; Yang, L.; Esvelt, K.M.; Aach, J.; Guell, M.; DiCarlo, J.E.; Julie, E.N.; Church, G.M. RNA-guided human genome engineering via Cas9. Science 2013, 339, 823–826. [Google Scholar] [CrossRef] [Green Version]
- Hrusha, A.; Krawitz, P.; Rechenberg, A.; Heinrich, V.; Hecht, J.; Haass, C.; Schmid, B. Efficient CRISPR/Cas9 genome editing with low off-target effects in zebrafish. Development 2013, 140, 4982–4987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedland, A.E.; Tzur, Y.B.; Esvelt, K.M.; Colaiacovo, M.P.; Church, G.M.; Calarco, J.A. Heritable genome editing in C. elegans via a CRISPR-Cas9 system. Nat. Methods 2013, 10, 741–743. [Google Scholar]
- Zhou, H.; He, M.; Li, J.; Chen, L.; Huang, Z.; Zheng, S.; Zhu, L.; Ni, E.; Jiang, D.; Zhao, B.; et al. Development of commercial thermos-sensitive genic male sterile rice accelerates hybrid rice breeding using the CRISPR/Cas9-mediated TMS5 editing system. Sci. Rep. 2016, 6, 37395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, X.; Li, W.; Liu, Y.; Tan, M.; Ganal, M.; Chetelat, R.T. A farnesyl pyrophosphate synthase gene expressed in pollen functions in S-RNase-independent unilateral incompatibility. Plant J. 2018, 93, 417–430. [Google Scholar] [CrossRef]
- Nekrasov, V.; Staskawicz, B.; Weigel, D.; Jones, J.D.; Kamoun, S. Targeted mutagenesis in the model plant Nicotiana benthamiana using Cas9 RNA-guided endonuclease. Nat. Biotechnol. 2013, 31, 691–693. [Google Scholar] [CrossRef]
- Mao, Y.; Zhang, H.; Xu, N.; Zhang, B.; Gou, F.; Zhu, J.K. Application of the CRISPR-Cas system for efficient genome engineering in plants. Mol. Plant 2013, 6, 2008–2011. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.G.; Kim, D.H.; Park, C.G.; Yeo, J.H.; Ahn, Y.S.; Park, H.K. Effect on Breaking of Self-Incompatibility by Old-Flower Pollination, NaCl and CO2 Treatment in Astragalus membranaceus Bunge. Korean J. Plant Res. 2009, 22, 467–472. [Google Scholar]
- Lao, X.; Suwabe, K.; Niikura, S.; Kakita, M.; Iwano, M.; Takayama, S. Physiological and genetic analysis of CO2-induced breakdown of self-incompatibility in Brassica rapa. J. Exp. Bot. 2014, 65, 939–951. [Google Scholar] [CrossRef] [Green Version]
- Lv, H.; Miyaji, N.; Osabe, K.; Akter, A.; Mehraj, H.; Shea, D.J.; Fujimoto, R. The importance of genetic and epigenetic research in the Brassica vegetables in the face of climate change. In Genomic Designing of Climate-Smart Vegetable Crops; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 161–255. [Google Scholar]
- Muñoz-Sanz, J.V.; Zuriaga, E.; Cruz-García, F.; McClure, B.; Romero, C. Self-(In) compatibility systems: Target traits for crop-production, plant breeding, and biotechnology. Front. Plant Sci. 2020, 11, 195. [Google Scholar] [CrossRef] [Green Version]
- Nasrallah, J.B.; Kao, T.H.; Goldberg, M.L.; Nasrallah, M.E. A cDNA encoding an S-locus specific glycoprotein from Brassica oleracea. Nature 1985, 318, 263–267. [Google Scholar]
- Takasaki, T.; Hatakeyama, K.; Suzuki, G.; Watanabe, M.; Isogai, A.; Hinata, K. The S receptor kinase determines self-incompatibility in brassica stigma. Nature 2000, 403, 913–916. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Yang, Y.; Li, B.; Liu, Z.; Khan, F.; Zhang, T.; Zhou, G.; Tu, J.; Shen, J.; Yi, B.; et al. Functional analysis of M-locus protein kinase revealed a novel regulatory mechanism of self-incompatibility in Brassica napus L. Int. J. Mol. Sci. 2019, 20, 3303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azibi, T.; Hadj-Arab, H.; Lodé, M.; de Carvalho, J.F.; Trotoux, G.; Nègre, S.; Gilet, M.M.; Boutte, J.; Lucas, J.; Vekemans, X.; et al. Impact of whole genome triplication on the evolutionary history and the functional dynamics of regulatory genes involved in Brassica self-incompatibility signalling pathway. Plant Reprod. 2020, 33, 43–58. [Google Scholar] [CrossRef]
- Dou, S.; Zhang, T.; Tu, J.; Shen, J.; Yi, B.; Wen, J.; Fu, T.; Dai, C.; Ma, C. Generation of novel self-incompatible Brassica napus by CRISPR/Cas9. Plant Biotechnol. J. 2021, 19, 875. [Google Scholar] [CrossRef] [PubMed]
- Tung, H.L.; Alemany, S.; Cohen, P. The protein phosphatases involved in cellular regulation: 2. Purification, subunit structure and properties of protein phosphatases-2Ao, 2A1, and 2A2 from rabbit skeletal muscle. Eur. J. Biochem. 1985, 148, 253–263. [Google Scholar] [CrossRef]
- Seshacharyulu, P.; Pandey, P.; Datta, K.; Batra, S.K. Phosphatase: PP2A structural importance, regulation and its aberrant expression in cancer. Cancer Lett. 2013, 335, 9–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, P. The structure and regulation of protein phosphatases. Annu. Rev. Biochem. 1989, 58, 453–508. [Google Scholar] [CrossRef]
- Mayer-Jaekel, R.E.; Hemmings, B.A. Protein phosphatase 2A−a ‘menage a trois’. Trends Cell Biol. 1994, 4, 287–291. [Google Scholar] [CrossRef]
- Smith, R.D.; Walker, J.C. Plant protein phosphatases. Annu. Rev. Plant Biol. 1996, 47, 101–125. [Google Scholar] [CrossRef] [Green Version]
- Luan, S. Protein phosphatases in plants. Annu. Rev. Plant Biol. 2003, 54, 63–92. [Google Scholar] [CrossRef]
- MacKintosh, C.; Cohen, P. Identification of high levels of type 1 and type 2A protein phosphatases in high levels of type 1 and type 2A protein phosphatases in higher plants. Biochem. J. 1989, 262, 335–339. [Google Scholar] [CrossRef] [Green Version]
- Jagiello, I.; Donella-Deana, A.; Szczegielnia, J.; Pinna, L.A.; Muszyńska, G. Identification of protein phosphatase activities in maize seedlings. Biochim. Biophys. Acta—Mol. Cell Res. 1992, 1134, 129–136. [Google Scholar] [CrossRef]
- Bialojan, C.; Takai, A. Inhibitory effect of a marine-sponge toxin, okadaic acid, on protein phosphatases. Biochem. J. 1988, 256, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Scutt, C.P.; Fordham-Skelton, A.P.; Croy, R.R.D. Okadaic acid causes breakdown of self-incompatibility in Brassica oleracea: Evidence for the involvement of protein phosphatases in the incompatible response. Sex. Plant Reprod. 1993, 6, 282–285. [Google Scholar] [CrossRef]
- Lee, G.H.; Shin, N.R.; Park, Y.D. Reverse genetics analysis of the 55-kDa B regulatory subunit of 2A serine/threonine protein phosphatase (PP2A) related to self-incompatibility in Chinese cabbage. Hortic. Sci. Technol. 2020, 61, 127–137. [Google Scholar] [CrossRef]
- Yu, S.G.; Cho, N.H.; Kim, J.H.; Oh, T.R.; Kim, W.T. Suppression of DRR1 results in the accumulation of insoluble ubiquitinated proteins, which impairs drought stress tolerance. J. Integr. Plant Biol. 2021, 63, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Son, G.H.; Moon, J.; Shelake, R.M.; Vuong, U.T.; Ingle, R.A.; Gassmann, W.; Kim, J.Y.; Kim, S.H. Conserved Opposite Functions in Plant Resistance to Biotrophic and Necrotrophic Pathogens of the Immune Regulator SRFR1. Int. J. Mol. Sci. 2021, 22, 6427. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Liu, H.; Lin, Y.; Chen, J.; Fu, Y.; Lou, J.; Wang, F. In-frame and frame-shift editing of the EHd1 gene to develop ja-ponica rice with prolonged basic vegetative growth periods. Front. Plant Sci. 2020, 11, 307. [Google Scholar] [CrossRef] [Green Version]
- Fan, D.; Liu, T.; Li, C.; Jiao, B.; Li, S.; Hou, Y.; Luo, K. Efficient CRISPR/Cas9-mediated targeted mutagenesis in Populus in the first generation. Sci. Rep. 2015, 5, 1–7. [Google Scholar] [CrossRef]
- Enciso-Rodriguez, F.; Manrique-Carpintero, N.C.; Nadakuduti, S.S.; Buell, C.R.; Zarka, D.; Douches, D. Overcoming self-incompatibility in diploid potato using CRISPR-Cas9. Front. Plant Sci. 2019, 10, 376. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.S.; Feng, K.; Xiong, A.S. CRISPR/Cas9-mediated multiply targeted mutagenesis in orange and purple carrot plants. Mol. Biotechnol. 2019, 61, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Doll, N.M.; Gilles, L.M.; Gérentes, M.F.; Richard, C.; Just, J.; Fierlej, Y.; Borrelli, V.M.G.; Gendrot, G.; Ingram, G.C.; Rogowsky, P.M.; et al. Single and multiple gene knockouts by CRISPR–Cas9 in maize. Plant Cell Rep. 2019, 38, 487–501. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Liu, Y.; Guo, Y.; Duan, W.; Fan, P.; Li, S.; Liang, Z. Optimizing the CRISPR/Cas9 system for genome editing in grape by using grape promoters. Hortic. Res. 2021, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Liu, M.; Li, Q.; Si, J.; Ren, X.; Song, H. Efficient BoPDS gene editing in cabbage by the CRISPR/Cas9 system. Hortic. Plant J. 2019, 5, 164–169. [Google Scholar]
- Lopez-obando, M.; Hoffmann, B.; Géry, C.; Guyon-Debast, A.; Téoulé, E.; Rameau, C.; Bonhomme, S.; Nogué, F. Simple and efficient targeting of multiple genes through CRISPR-Cas9 in physcomitrella patens. G3 Genes Genomes Genet. 2016, 6, 3647–3653. [Google Scholar] [CrossRef] [Green Version]
- Shan, S.; Soltis, P.S.; Soltis, D.E.; Yang, B. Considerations in adapting CRISPR/Cas9 in nongenetic model plant systems. Appl. Plant Sci. 2020, 8, e11314. [Google Scholar] [CrossRef]
- Pramanik, D.; Shelake, R.M.; Park, J.; Kim, M.J.; Hwang, I.; Park, Y.; Kim, J.Y. CRISPR/Cas9-mediated generation of pathogen-resistant tomato against tomato yellow leaf curl virus and powdery mildew. Int. J. Mol. Sci. 2021, 22, 1878. [Google Scholar] [CrossRef]
- Elorriaga, E.; Klocko, A.L.; Ma, C.; Strauss, S.H. Variation in mutation spectra among CRISPR/Cas9 mutagenized poplars. Front. Plant Sci. 2018, 9, 594. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Guo, D.; Chang, Y.; You, C.; Li, X.; Dai, X.; Weng, Q.; Zhang, J.; Chen, G.; Li, X.; et al. Non-random distribution of T-DNA insertions at various levels of the genome hierarchy as revealed by analyzing 13 804 T-DNA flanking sequences from an enhancer-trap mutant library. Plant J. 2007, 9, 947–959. [Google Scholar] [CrossRef]
- Pla, M. Insert stability and transgenic plant risk. In Encyclopedia of Biotechnology in Agriculture and Food; CRC Press Inc.: London, UK, 2012. [Google Scholar]
- Forsbach, B.; Schubert, D.; Lechtenberg, B.; Gils, M.; Schmidt, R. A comprehensive characterization of single copy T-DNA insertions in the Arabidopsis thaliana genome. Plant Mol. Biol. 2003, 52, 161–176. [Google Scholar] [CrossRef]
- Peterson, B.A.; Haak, D.C.; Nishimura, M.T.; Teixeira, P.J.P.L.; James, S.R.; Dangl, J.L.; Nimchuk, Z.L. Genome-wide assessment of efficiency and specificity in CRISPR/Cas9 mediated multiple site targeting in Arabidopsis. PLoS ONE 2016, 11, e0162169. [Google Scholar] [CrossRef] [PubMed]
- Hahn, F.; Nekrasov, V. CRISPR/Cas precision: Do we need to worry about off-targeting in plants? Plant Cell Rep. 2018, 38, 437–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, L.; Ntui, V.O.; Tripathi, J.N. RNA interference and CRISPR/Cas9 applications for virus resistance. In CRISPR and RNAi Systems: Nanobiotechnology Approaches to Plant Breeding and Protection; Elsevier: Amsterdam, The Netherlands, 2021; pp. 162–182. [Google Scholar]
- Nishihara, M.; Higuchi, A.; Watanabe, A.; Tasaki, K. Application of the CRISPR/Cas9 system for modification of flower color in Torenia fournieri. BMC Plant Biol. 2018, 18, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charrier, A.; Vergne, E.; Dousset, N.; Richer, A.; Petiteau, A.; Chevreau, E. Efficient targeted mutagenesis in apple and first time edition of pear using the CRISPR-Cas9 system. Front. Plant Sci. 2019, 10, 40. [Google Scholar] [CrossRef] [Green Version]
- Zhou, E.; Zhang, Y.; Wang, H.; Jia, Z.; Wang, X.; Wen, J.; Shen, J.; Fu, T.; Yi, B. Identification and Characterization of the MIKC-Type MADS-Box Gene Family in Brassica napus and Its Role in Floral Transition. Int. J. Mol. Sci. 2022, 23, 4289. [Google Scholar] [CrossRef]
- Martínez, M.I.S.; Bracuto, V.; Koseoglou, E.; Appiano, M.; Jacobsen, E.; Visser, R.G.; Wolters, A.M.A.; Bai, Y. CRISPR/Cas9-targeted mutagenesis of the tomato susceptibility gene PMR4 for resistance against powdery mildew. BMC Plant Biol. 2020, 20, 1–13. [Google Scholar]
- Navet, N.; Tian, M.Y. Efficient targeted mutagenesis in allotetraploid sweet basil by CRISPR/Cas9. Plant Direct 2020, 4, e00233. [Google Scholar] [CrossRef]
- Wan, D.Y.; Guo, Y.; Cheng, Y.; Hu, Y.; Xiao, S.; Wang, Y.; Wen, Y.Q. CRISPR/Cas9-mediated mutagenesis of VvMLO3 results in enhanced resistance to powdery mildew in grapevine (Vitis vinifera). Hort. Res. 2020, 7, 116. [Google Scholar] [CrossRef]
- Kim, H.R.; Kim, S.T.; Ryu, J.H.; Choi, M.K.; Kweon, J.Y.; Kang, B.C.; Ahn, H.M.; Bae, S.J.; Kim, J.G.; Kim, J.S.; et al. A simple, flexible and high-throughput cloning system for plant genome editing via CRISPR-Cas system. J. Integr. Plant Biol. 2016, 58, 705–712. [Google Scholar] [CrossRef]
- Jyothishwaran, G.; Kotresha, D.; Selvaraj, T.; Srideshikan, S.H.; Rajvanshi, P.K.; Jayabaskaran, C. A modified freeze-thaw method for efficient transformation of Agrobacterium tumefaciens. Curr. Sci. 2007, 93, 770–772. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Lee, M.K.; Kim, H.S.; Kim, J.S.; Kim, S.H.; Park, Y.D. Agrobacterium-mediated transformation system for large-scale producion of transgenic Chinese cabbage (Brassica rapa L. ssp. pekinensis) plants for insertional mutagenesis. J. Plant Biol. 2004, 47, 300–306. [Google Scholar] [CrossRef]
- McCouch, S.R.; Kochert, G.; Yu, Z.H.; Wang, Z.Y.; Khush, G.S.; Coffman, W.R.; Tanksley, S.D. Molecular mapping of rice chromosomes. Theor. Appl. Genet. 1988, 76, 815–829. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Line | Methods of Pollination z | Pods | Seeds | Seed Fertility y |
|---|---|---|---|---|
| IB | FP | 0 | 0 | 0 |
| BP | 49 | 121 | 2.47 | |
| IB | FP | 0 | 0 | 0 |
| BP | 32 | 72 | 2.25 | |
| PT1-2 | FP | 32 | 79 | 2.47 |
| BP | 24 | 67 | 2.80 | |
| PT1-4 | FP | 48 | 76 | 1.58 |
| BP | 32 | 50 | 1.56 | |
| PT1-8 | FP | 26 | 64 | 2.46 |
| BP | 40 | 105 | 2.63 | |
| PT3-3 | FP | 40 | 72 | 1.80 |
| BP | 24 | 50 | 2.08 |
| Line | Methods of Pollination z | Pods | Seeds | Seed Fertility y |
|---|---|---|---|---|
| IB | FP | 0 | 0 | 0 |
| BP | 52 | 108 | 2.08 | |
| IB | FP | 0 | 0 | 0 |
| BP | 30 | 55 | 1.83 | |
| PT1-2-2 | FP | 30 | 52 | 1.73 |
| BP | 28 | 49 | 1.75 | |
| PT1-4-1 | FP | 43 | 70 | 1.63 |
| BP | 49 | 90 | 1.84 | |
| PT1-8-3 | FP | 30 | 55 | 1.83 |
| BP | 58 | 115 | 1.98 | |
| PT3-3-3 | FP | 40 | 72 | 1.80 |
| BP | 38 | 70 | 1.84 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, N.-R.; Shin, Y.-H.; Kim, H.-S.; Park, Y.-D. Function Analysis of the PR55/B Gene Related to Self-Incompatibility in Chinese Cabbage Using CRISPR/Cas9. Int. J. Mol. Sci. 2022, 23, 5062. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095062
Shin N-R, Shin Y-H, Kim H-S, Park Y-D. Function Analysis of the PR55/B Gene Related to Self-Incompatibility in Chinese Cabbage Using CRISPR/Cas9. International Journal of Molecular Sciences. 2022; 23(9):5062. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095062
Chicago/Turabian StyleShin, Na-Ri, Yun-Hee Shin, Han-Seul Kim, and Young-Doo Park. 2022. "Function Analysis of the PR55/B Gene Related to Self-Incompatibility in Chinese Cabbage Using CRISPR/Cas9" International Journal of Molecular Sciences 23, no. 9: 5062. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095062