
Cloning and Functional Characterization of Dog OCT1 and OCT2: Another Step in Exploring Species Differences in Organic Cation Transporters

 ,
,  and
and
Abstract
:1. Introduction
2. Results
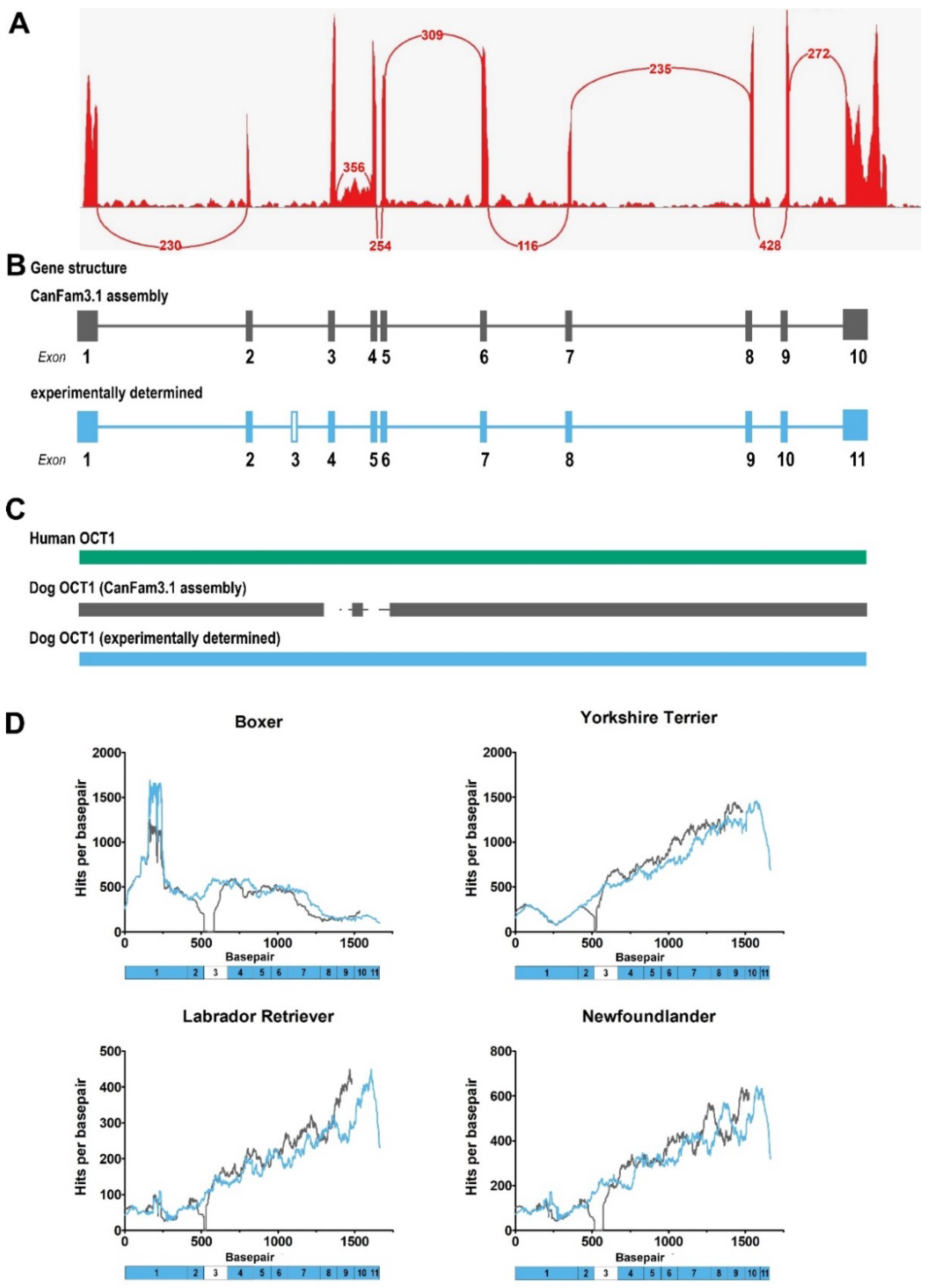
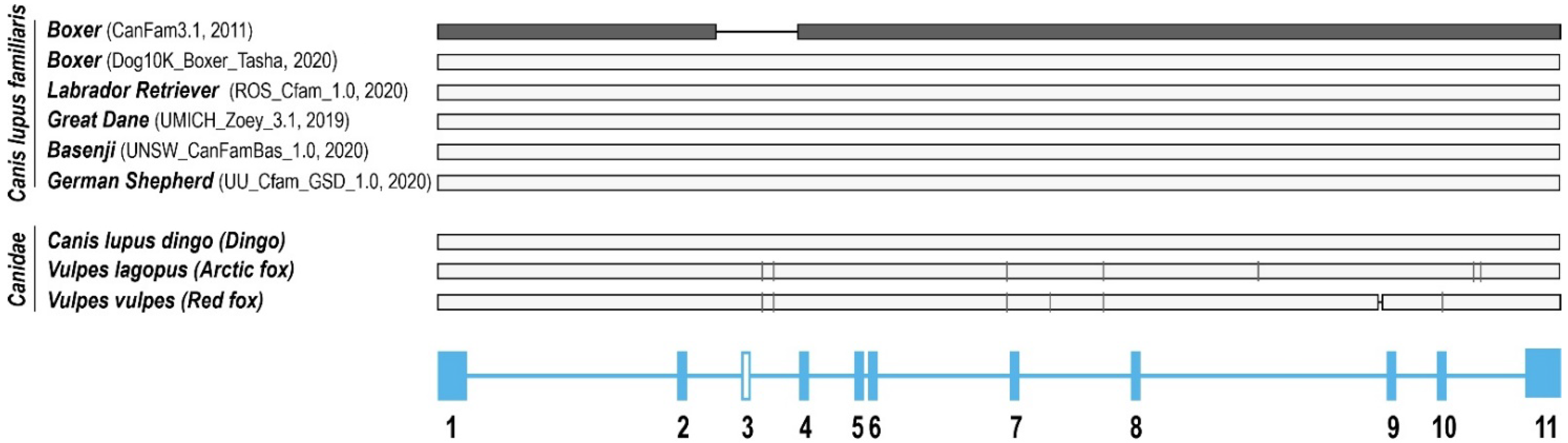
2.1. Gene Structure of Dog OCT1 and OCT2
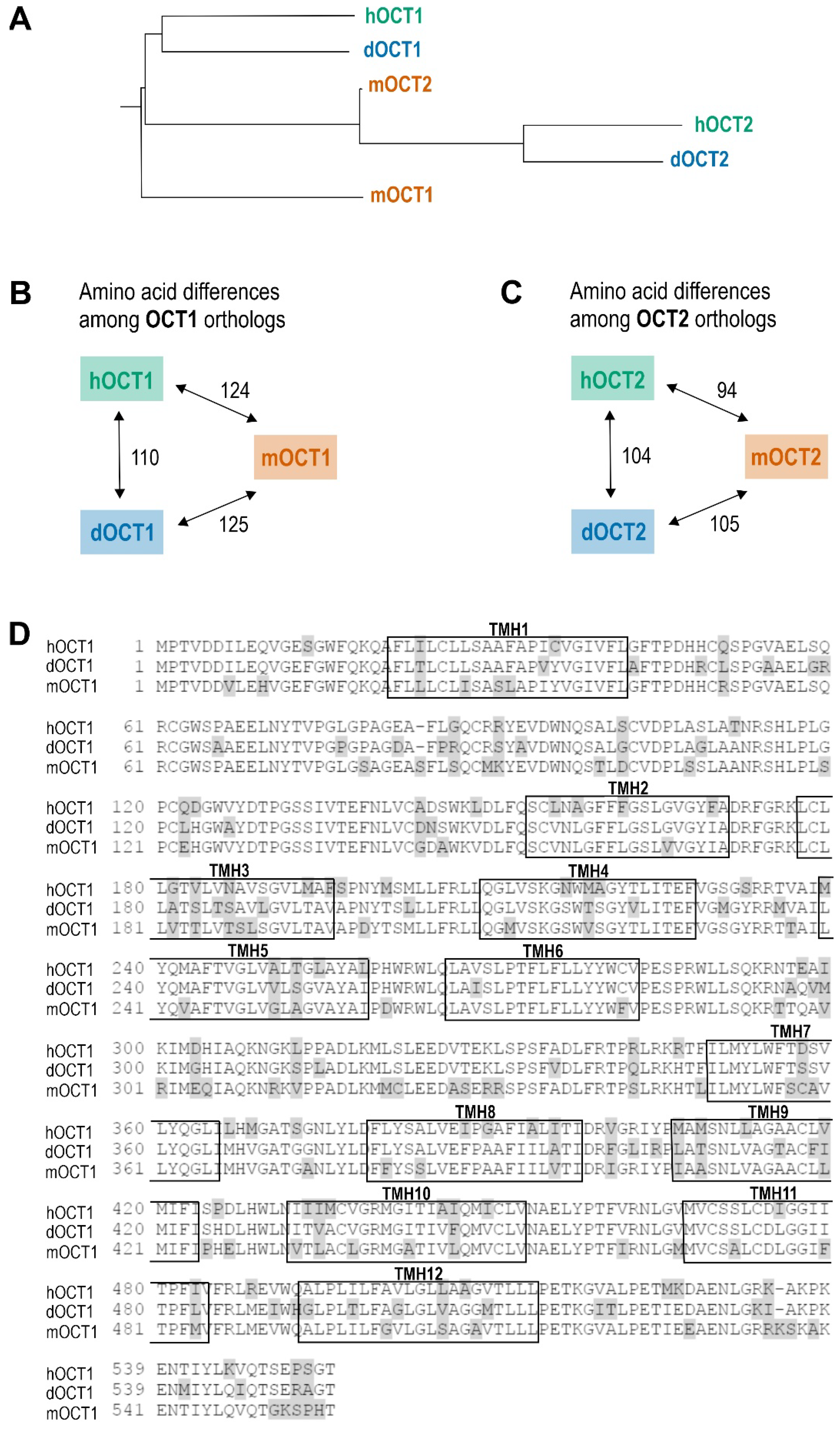
2.2. Cloning of Dog OCT1 and OCT2 mRNA Transcripts from Dog Liver and Kidney
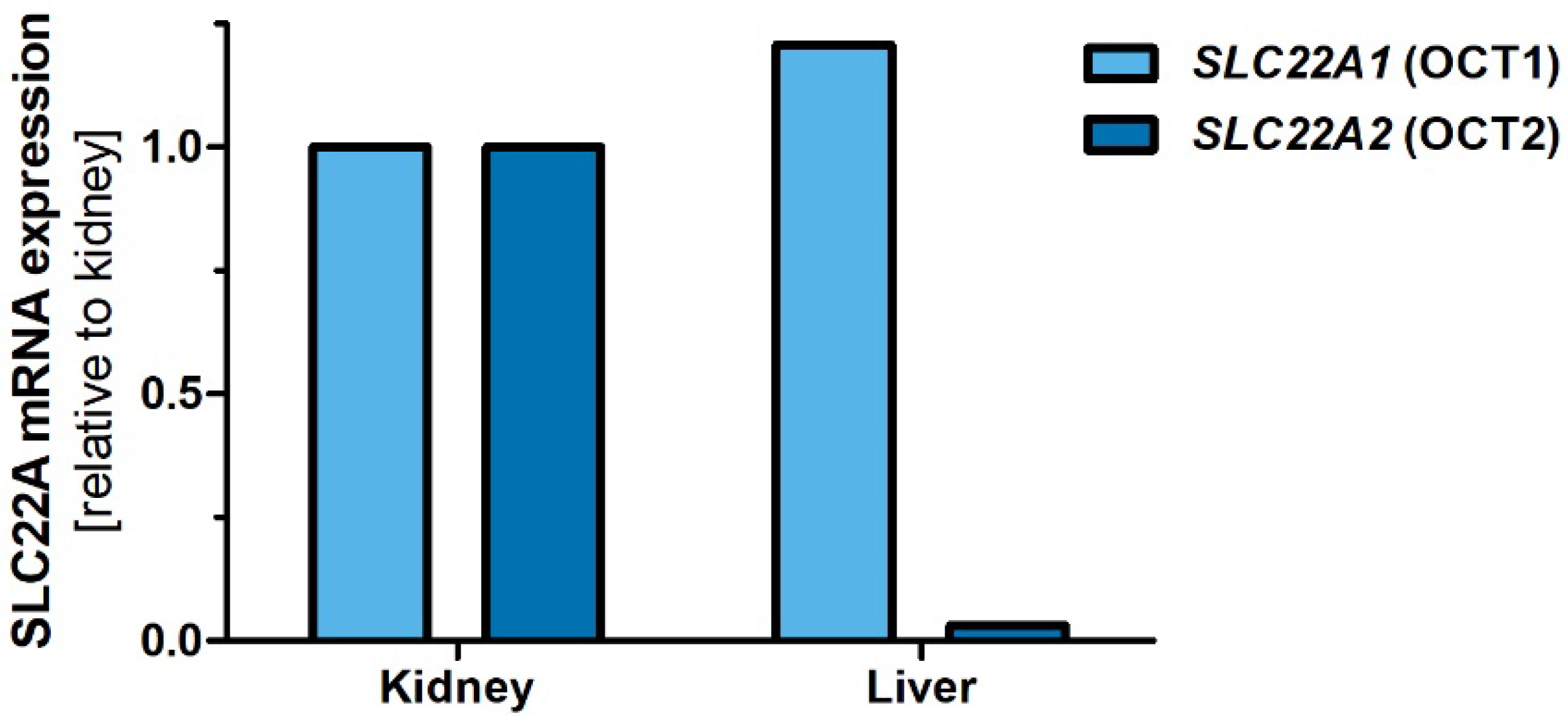
2.3. Organ Expression of Dog OCT1 and OCT2
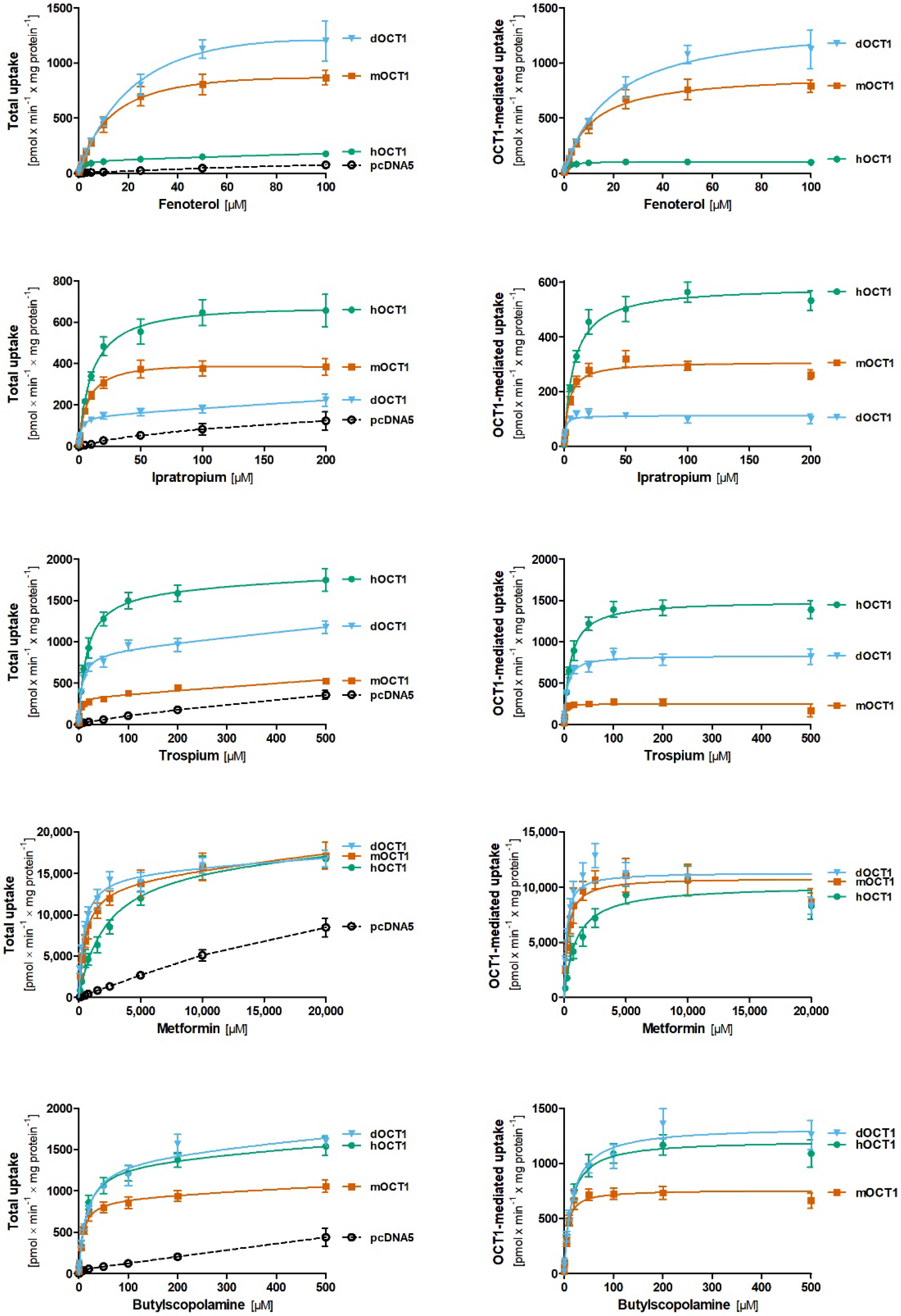
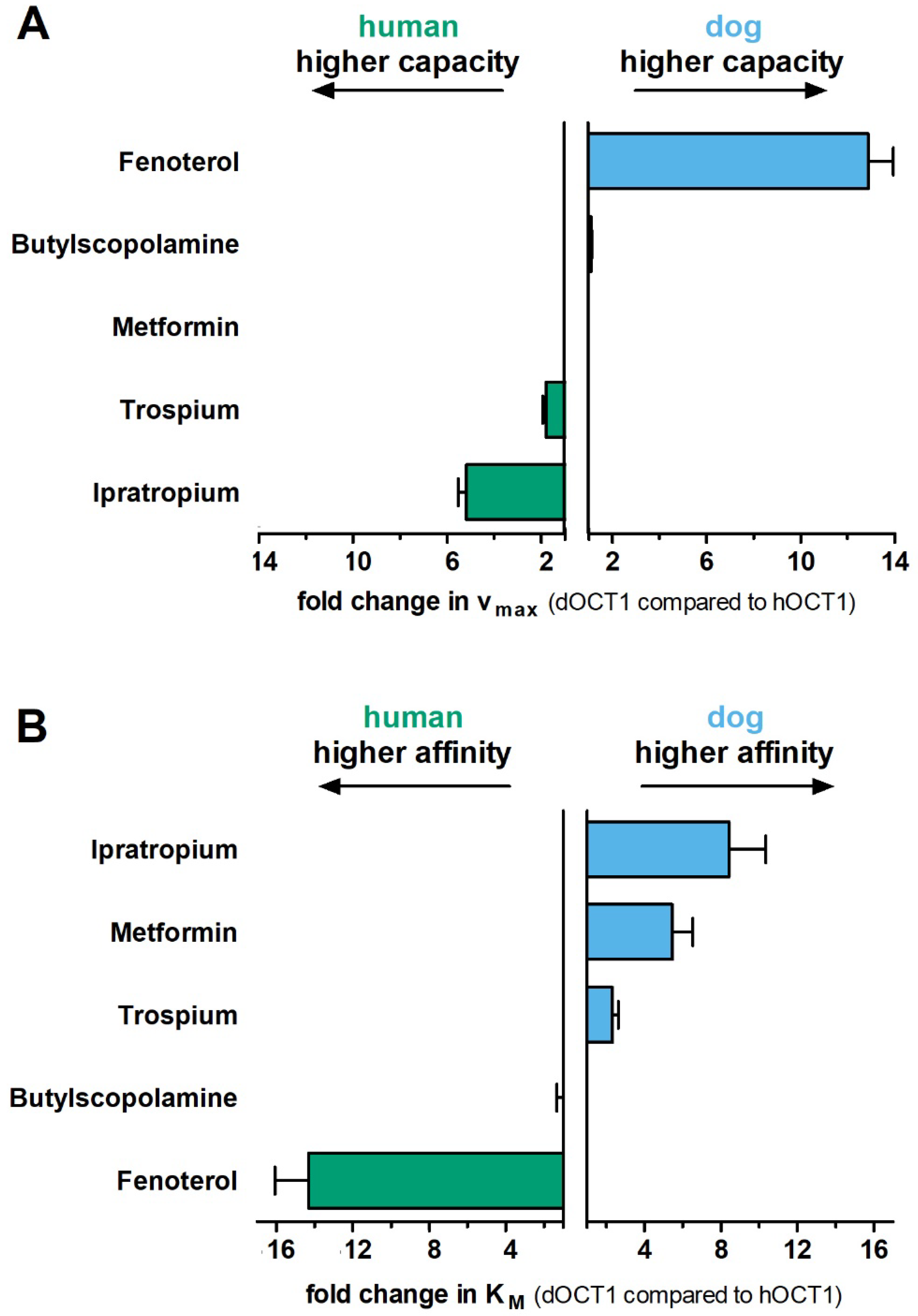
2.4. Comparative Characterization of Transport Kinetics between Human and Dog OCT1
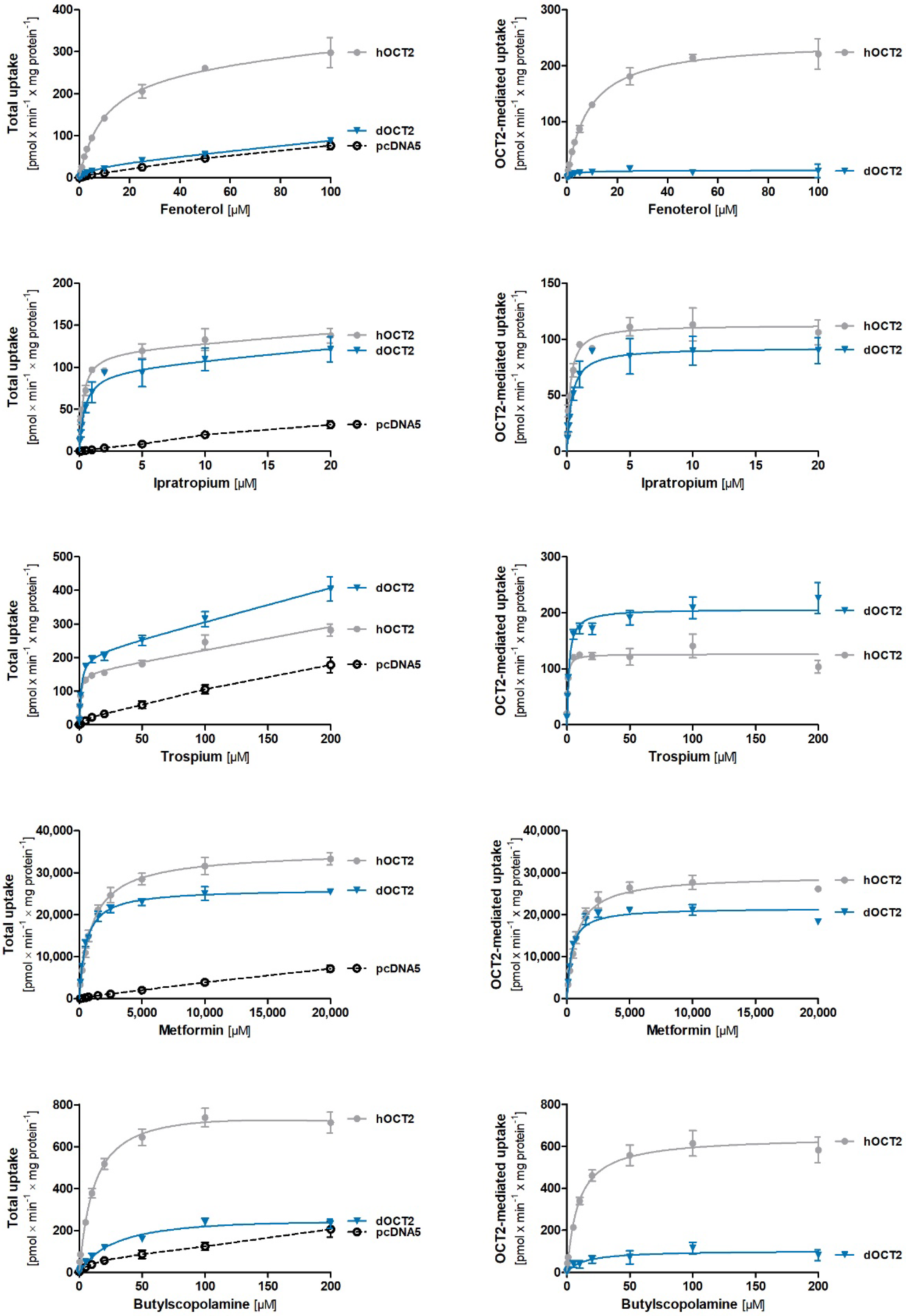
2.5. Comparative Characterization of Transport Kinetics between Human and Dog OCT2
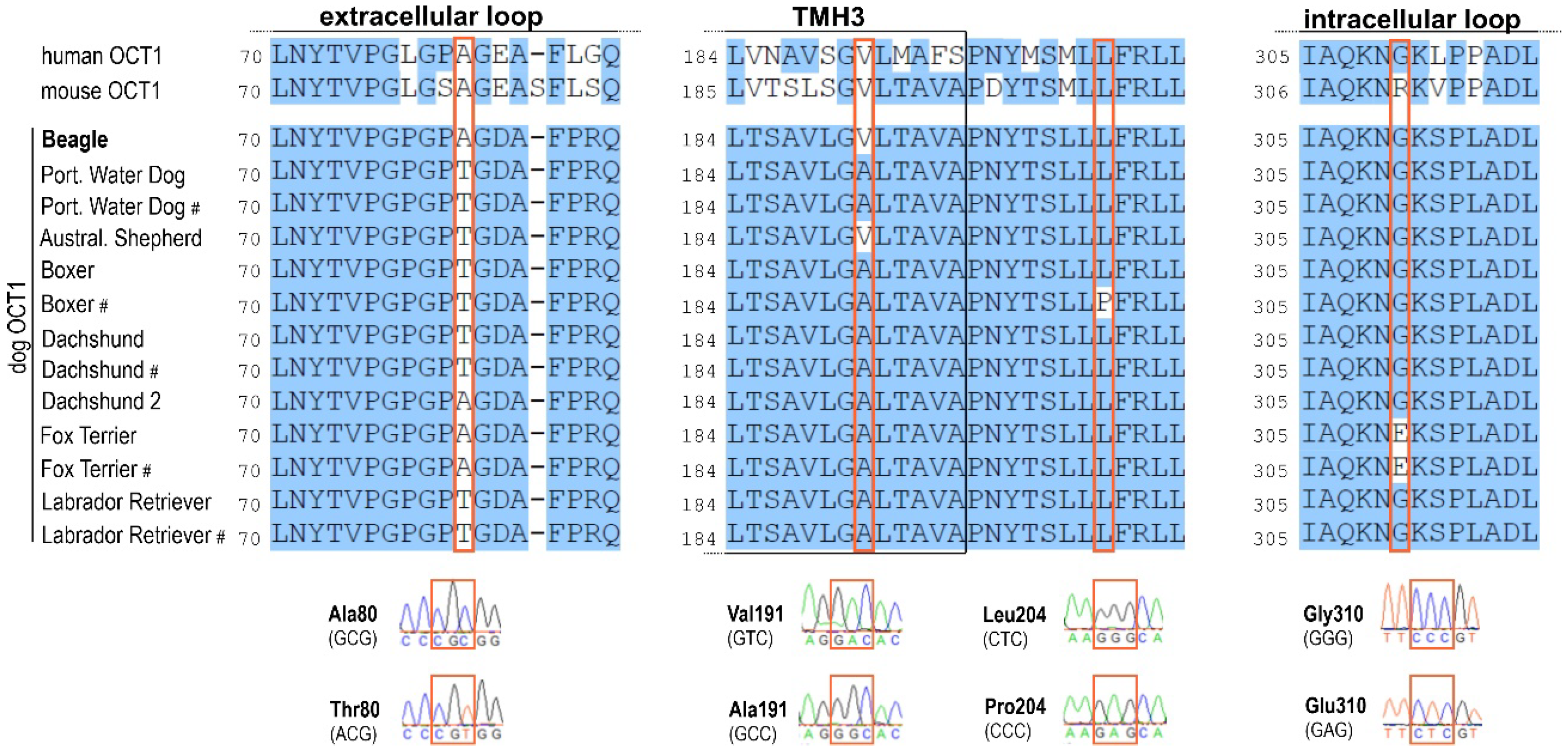
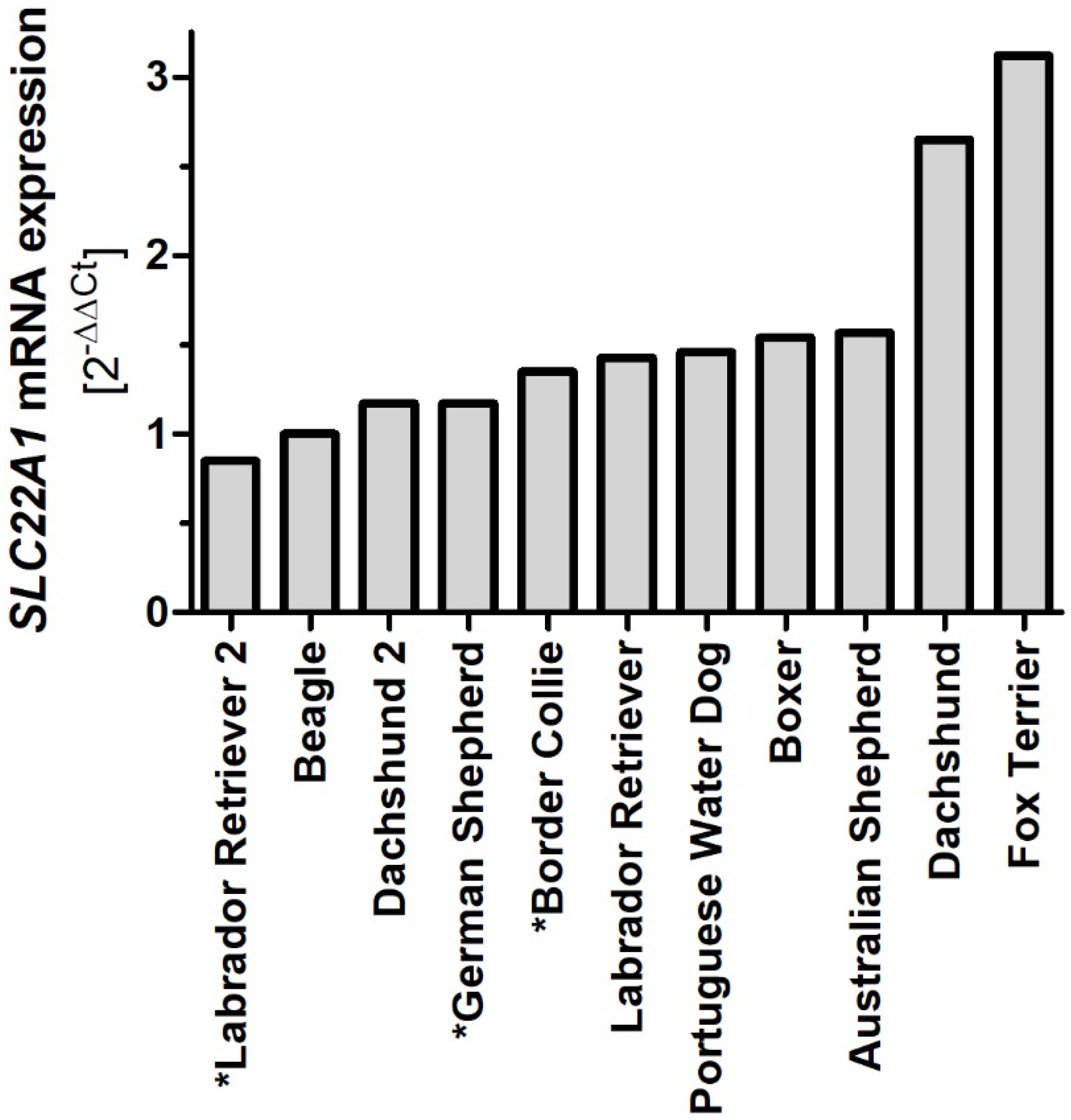
2.6. Variability of OCT1 Sequence and Expression among Different Dog Breeds
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cloning Dog OCT1 and OCT2 from Dog Liver and Kidney
4.3. mRNA Expression Analysis of Dog OCT1 and OCT2
4.4. Cell Lines and Cell Culturing
4.5. Cellular Uptake Experiments
4.6. Quantification of Intracellular Substrate Concentration by LC-MS/MS
4.7. Mapping of RNA-Seq Datasets on Dog OCT1 and OCT2 Reference
4.8. Data Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, L.; Dresser, M.J.; Gray, A.T.; Yost, S.C.; Terashita, S.; Giacomini, K.M. Cloning and Functional Expression of a Human Liver Organic Cation Transporter. Mol. Pharmacol. 1997, 51, 913–921. [Google Scholar] [CrossRef]
- Gorboulev, V.; Ulzheimer, J.C.; Akhoundova, A.; Ulzheimer-Teuber, I.; Karbach, U.; Quester, S.; Baumann, C.; Lang, F.; Busch, A.E.; Koepsell, H. Cloning and Characterization of Two Human Polyspecific Organic Cation Transporters. DNA Cell Biol. 1997, 16, 871–881. [Google Scholar] [CrossRef]
- Gründemann, D.; Gorboulev, V.; Gambaryan, S.; Veyhl, M.; Koepsell, H. Drug excretion mediated by a new prototype of polyspecific transporter. Nature 1994, 372, 549–552. [Google Scholar] [CrossRef] [Green Version]
- Green, R.M.; Lo, K.; Sterritt, C.; Beier, D.R. Cloning and functional expression of a mouse liver organic cation transporter. Hepatology 1999, 29, 1556–1562. [Google Scholar] [CrossRef]
- Schmitt, A.; Mössner, R.; Gossmann, A.; Fischer, I.G.; Gorboulev, V.; Murphy, D.L.; Koepsell, H.; Lesch, K.P. Organic cation transporter capable of transporting serotonin is up-regulated in serotonin transporter-deficient mice. J. Neurosci. Res. 2003, 71, 701–709. [Google Scholar] [CrossRef]
- Wang, D.-S.; Jonker, J.W.; Kato, Y.; Kusuhara, H.; Schinkel, A.H.; Sugiyama, Y. Involvement of Organic Cation Transporter 1 in Hepatic and Intestinal Distribution of Metformin. J. Pharmacol. Exp. Ther. 2002, 302, 510–515. [Google Scholar] [CrossRef] [Green Version]
- Shu, Y.; Brown, C.; Castro, R.A.; Shi, R.J.; Lin, E.T.; Owen, R.P.; Sheardown, S.A.; Yue, L.; Burchard, E.G.; Brett, C.M.; et al. Effect of genetic variation in the organic cation transporter 1, OCT1, on metformin pharmacokinetics. Clin. Pharmacol. Ther. 2008, 83, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Tzvetkov, M.V.; Vormfelde, S.V.; Balen, D.; Meineke, I.; Schmidt, T.; Sehrt, D.; Sabolić, I.; Koepsell, H.; Brockmöller, J. The Effects of Genetic Polymorphisms in the Organic Cation Transporters OCT1, OCT2, and OCT3 on the Renal Clearance of Metformin. Clin. Pharmacol. Ther. 2009, 86, 299–306. [Google Scholar] [CrossRef]
- Tzvetkov, M.V.; Matthaei, J.; Pojar, S.; Faltraco, F.; Vogler, S.; Prukop, T.; Seitz, T.; Brockmöller, J. Increased Systemic Exposure and Stronger Cardiovascular and Metabolic Adverse Reactions to Fenoterol in Individuals with Heritable OCT1 Deficiency. Clin. Pharmacol. Ther. 2018, 103, 868–878. [Google Scholar] [CrossRef]
- Matthaei, J.; Kuron, D.; Faltraco, F.; Knoch, T.; Dos Santos Pereira, J.N.; Abu Abed, M.; Prukop, T.; Brockmöller, J.; Tzvetkov, M.V. OCT1 mediates hepatic uptake of sumatriptan and loss-of-function OCT1 polymorphisms affect sumatriptan pharmacokinetics. Clin. Pharmacol. Ther. 2016, 99, 633–641. [Google Scholar] [CrossRef]
- Tzvetkov, M.V.; Saadatmand, A.R.; Lotsch, J.; Tegeder, I.; Stingl, J.C.; Brockmöller, J. Genetically Polymorphic OCT1: Another Piece in the Puzzle of the Variable Pharmacokinetics and Pharmacodynamics of the Opioidergic Drug Tramadol. Clin. Pharmacol. Ther. 2011, 90, 143–150. [Google Scholar] [CrossRef]
- Chen, J.; Brockmöller, J.; Seitz, T.; König, J.; Tzvetkov, M.V.; Chen, X. Erratum to: Tropane alkaloids as substrates and inhibitors of human organic cation transporters of the SLC22 (OCT) and the SLC47 (MATE) families. Biol. Chem. 2017, 398, 813. [Google Scholar] [CrossRef]
- Bourdet, D.L.; Pritchard, J.B.; Thakker, D.R. Differential Substrate and Inhibitory Activities of Ranitidine and Famotidine toward Human Organic Cation Transporter 1 (hOCT1; SLC22A1), hOCT2 (SLC22A2), and hOCT3 (SLC22A3). J. Pharmacol. Exp. Ther. 2005, 315, 1288–1297. [Google Scholar] [CrossRef] [Green Version]
- Meyer, M.J.; Seitz, T.; Brockmöller, J.; Tzvetkov, M.V. Effects of genetic polymorphisms on the OCT1 and OCT2-mediated uptake of ranitidine. PLoS ONE 2017, 12, e0189521. [Google Scholar] [CrossRef] [Green Version]
- Hendrickx, R.; Johansson, J.G.; Lohmann, C.; Jenvert, R.-M.; Blomgren, A.; Börjesson, L.; Gustavsson, L. Identification of Novel Substrates and Structure–Activity Relationship of Cellular Uptake Mediated by Human Organic Cation Transporters 1 and 2. J. Med. Chem. 2013, 56, 7232–7242. [Google Scholar] [CrossRef]
- European Medicines Agency. Guideline on the Investigation of Drug Interactions; European Medicines Agency: London, UK, 2012.
- U.S. Food and Drug Administration. In Vitro Drug Interaction Studies—Cytochrome P450 Enzyme- and Transporter-Mediated Drug Interactions: Guidance for Industry; U.S. Food and Drug Administration: Silver Spring, MD, USA, 2020.
- Zamek-Gliszczynski, M.J.; Taub, M.E.; Chothe, P.P.; Chu, X.; Giacomini, K.M.; Kim, R.B.; Ray, A.S.; Stocker, S.L.; Unadkat, J.D.; Wittwer, M.B.; et al. Transporters in Drug Development: 2018 ITC Recommendations for Transporters of Emerging Clinical Importance. Clin. Pharmacol. Ther. 2018, 104, 890–899. [Google Scholar] [CrossRef]
- Zamek-Gliszczynski, M.J.; Giacomini, K.M.; Zhang, L. Emerging Clinical Importance of Hepatic Organic Cation Transporter 1 (OCT1) in Drug Pharmacokinetics, Dynamics, Pharmacogenetic Variability, and Drug Interactions. Clin. Pharmacol. Ther. 2018, 103, 758–760. [Google Scholar] [CrossRef]
- European Medicines Agency. ICH Guideline M3(R2) on Non-Clinical Safety Studies for the Conduct of Human Clinical Trials and Marketing Authorisation for Pharmaceuticals: Step 5; European Medicines Agency: London, UK, 2009.
- U.S. Food and Drug Administration. Federal Food, Drug, and Cosmetic Act: FFDCA, FD&C Act. In United States Code; U.S. Food and Drug Administration: Silver Spring, MD, USA, 1938. [Google Scholar]
- Khanna, C.; Lindblad-Toh, K.; Vail, D.; London, C.; Bergman, P.; Barber, L.; Breen, M.; Kitchell, B.; McNeil, E.; Modiano, J.F.; et al. The dog as a cancer model. Nat. Biotechnol. 2006, 24, 1065–1066. [Google Scholar] [CrossRef]
- European Pet Food Industry Federation. Facts & Figures 2020: European Overview; European Pet Food Industry Federation: Bruxelles, Belgium, 2020. [Google Scholar]
- American Veterinary Medical Association. AVMA Pet Ownership and Demographics Sourcebook: 2017–2018 Edition; American Veterinary Medical Association: Washington, DC, USA, 2018. [Google Scholar]
- Bleasby, K.; Castle, J.C.; Roberts, C.J.; Cheng, C.; Bailey, W.J.; Sina, J.F.; Kulkarni, A.V.; Hafey, M.J.; Evers, R.; Johnson, J.M.; et al. Expression profiles of 50 xenobiotic transporter genes in humans and pre-clinical species: A resource for investigations into drug disposition. Xenobiotica 2006, 36, 963–988. [Google Scholar] [CrossRef]
- Gui, C.; Hagenbuch, B. Cloning/characterization of the canine organic anion transporting polypeptide 1b4 (Oatp1b4) and classification of the canine OATP/SLCO members. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2010, 151, 393–399. [Google Scholar] [CrossRef] [Green Version]
- Shu, Y.; Bello, C.L.; Mangravite, L.M.; Feng, B.; Giacomini, K.M. Functional characteristics and steroid hormone-mediated regulation of an organic cation transporter in Madin-Darby canine kidney cells. J. Pharmacol. Exp. Ther. 2001, 299, 392–398. [Google Scholar]
- Meyer, M.J.; Tzvetkov, M.V. OCT1 Polyspecificity—Friend or Foe? Front. Pharmacol. 2021, 12, 698153. [Google Scholar] [CrossRef]
- Meyer, M.J.; Tuerkova, A.; Römer, S.; Wenzel, C.; Seitz, T.; Gaedcke, J.; Oswald, S.; Brockmöller, J.; Zdrazil, B.; Tzvetkov, M.V. Differences in Metformin and Thiamine Uptake between Human and Mouse Organic Cation Transporter 1: Structural Determinants and Potential Consequences for Intrahepatic Concentrations. Drug Metab. Dispos. 2020, 48, 1380–1392. [Google Scholar] [CrossRef]
- Meyer, M.J.; Schreier, P.C.F.; Basaran, M.; Vlasova, S.; Seitz, T.; Brockmöller, J.; Zdrazil, B.; Tzvetkov, M.V. Amino acids in transmembrane helix 1 confer major functional differences between human and mouse orthologs of the polyspecific membrane transporter OCT1. J. Biol. Chem. 2022, 101974. [Google Scholar] [CrossRef]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for functional genomics data sets—Update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [Green Version]
- Athar, A.; Füllgrabe, A.; George, N.; Iqbal, H.; Huerta, L.; Ali, A.; Snow, C.; Fonseca, N.; Petryszak, R.; Papatheodorou, I.; et al. ArrayExpress update—From bulk to single-cell expression data. Nucleic Acids Res. 2019, 47, D711–D715. [Google Scholar] [CrossRef]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef] [Green Version]
- Karakus, E.; Prinzinger, C.; Leiting, S.; Geyer, J. Sequencing of the Canine Cytochrome P450 CYP2C41 Gene and Genotyping of Its Polymorphic Occurrence in 36 Dog Breeds. Front. Vet. Sci. 2021, 8, 663175. [Google Scholar] [CrossRef]
- Nies, A.T.; Koepsell, H.; Winter, S.; Burk, O.; Klein, K.; Kerb, R.; Zanger, U.M.; Keppler, D.; Schwab, M.; Schaeffeler, E. Expression of organic cation transporters OCT1 (SLC22A1) and OCT3 (SLC22A3) is affected by genetic factors and cholestasis in human liver. Hepatology 2009, 50, 1227–1240. [Google Scholar] [CrossRef]
- Floerl, S.; Kuehne, A.; Hagos, Y. Functional and Pharmacological Comparison of Human, Mouse, and Rat Organic Cation Transporter 1 toward Drug and Pesticide Interaction. Int. J. Mol. Sci. 2020, 21, 6871. [Google Scholar] [CrossRef]
- Dresser, M.J.; Gray, A.T.; Giacomini, K.M. Kinetic and selectivity differences between rodent, rabbit, and human organic cation transporters (OCT1). J. Pharmacol. Exp. Ther. 2000, 292, 1146–1152. [Google Scholar]
- Morse, B.L.; Kolur, A.; Hudson, L.R.; Hogan, A.T.; Chen, L.H.; Brackman, R.M.; Sawada, G.A.; Fallon, J.K.; Smith, P.C.; Hillgren, K.M. Pharmacokinetics of Organic Cation Transporter 1 (OCT1) Substrates in Oct1/2 Knockout Mice and Species Difference in Hepatic OCT1-Mediated Uptake. Drug Metab. Dispos. 2020, 48, 93–105. [Google Scholar] [CrossRef]
- Meyer, M.J.; Neumann, V.E.; Friesacher, H.R.; Zdrazil, B.; Brockmöller, J.; Tzvetkov, M.V. Opioids as Substrates and Inhibitors of the Genetically Highly Variable Organic Cation Transporter OCT1. J. Med. Chem. 2019, 62, 9890–9905. [Google Scholar] [CrossRef]
- Mise, M.; Hashizume, T.; Matsumoto, S.; Terauchi, Y.; Fujii, T. Identification of non-functional allelic variant of CYP1A2 in dogs. Pharmacogenetics 2004, 14, 769–773. [Google Scholar] [CrossRef]
- Mise, M.; Yadera, S.; Matsuda, M.; Hashizume, T.; Matsumoto, S.; Terauchi, Y.; Fujii, T. Polymorphic expression of CYP1A2 leading to interindividual variability in metabolism of a novel benzodiazepine receptor partial inverse agonist in dogs. Drug Metab. Dispos. 2004, 32, 240–245. [Google Scholar] [CrossRef] [Green Version]
- Tenmizu, D.; Endo, Y.; Noguchi, K.; Kamimura, H. Identification of the novel canine CYP1A2 1117 C T SNP causing protein deletion. Xenobiotica 2004, 34, 835–846. [Google Scholar] [CrossRef]
- Geyer, J.; Janko, C. Treatment of MDR1 mutant dogs with macrocyclic lactones. Curr. Pharm. Biotechnol. 2012, 13, 969–986. [Google Scholar] [CrossRef] [Green Version]
- Mealey, K.L.; Bentjen, S.A.; Gay, J.M.; Cantor, G.H. Ivermectin sensitivity in collies is associated with a deletion mutation of the mdr1 gene. Pharmacogenetics 2001, 11, 727–733. [Google Scholar] [CrossRef]
- Geyer, J.; Döring, B.; Meerkamp, K.; Ugele, B.; Bakhiya, N.; Fernandes, C.F.; Godoy, J.R.; Glatt, H.; Petzinger, E. Cloning and Functional Characterization of Human Sodium-dependent Organic Anion Transporter (SLC10A6). J. Biol. Chem. 2007, 282, 19728–19741. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Tzvetkov, M.V.; Saadatmand, A.R.; Bokelmann, K.; Meineke, I.; Kaiser, R.; Brockmoller, J. Effects of OCT1 polymorphisms on the cellular uptake, plasma concentrations and efficacy of the 5-HT3 antagonists tropisetron and ondansetron. Pharm. J. 2012, 12, 22–29. [Google Scholar] [CrossRef]
- Seitz, T.; Stalmann, R.; Dalila, N.; Chen, J.; Pojar, S.; Dos Santos Pereira, J.N.; Krätzner, R.; Brockmöller, J.; Tzvetkov, M.V. Global genetic analyses reveal strong inter-ethnic variability in the loss of activity of the organic cation transporter OCT1. Genome Med. 2015, 7, 56. [Google Scholar] [CrossRef] [Green Version]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Sedlazeck, F.J.; Rescheneder, P.; Von Haeseler, A. NextGenMap: Fast and accurate read mapping in highly polymorphic genomes. Bioinformatics 2013, 29, 2790–2791. [Google Scholar] [CrossRef] [Green Version]
- Lindblad-Toh, K.; Wade, C.M.; Mikkelsen, T.S.; Karlsson, E.K.; Jaffe, D.B.; Kamal, M.; Clamp, M.; Chang, J.L.; Kulbokas, E.J.; Zody, M.C.; et al. Genome sequence, comparative analysis and haplotype structure of the domestic dog. Nature 2005, 438, 803–819. [Google Scholar] [CrossRef]
- Hoeppner, M.P.; Lundquist, A.; Pirun, M.; Meadows, J.R.S.; Zamani, N.; Johnson, J.; Sundström, G.; Cook, A.; Fitzgerald, M.G.; Swofford, R.; et al. An Improved Canine Genome and a Comprehensive Catalogue of Coding Genes and Non-Coding Transcripts. PLoS ONE 2014, 9, e91172. [Google Scholar] [CrossRef]
- Chen, J.; Swofford, R.; Johnson, J.; Cummings, B.B.; Rogel, N.; Lindblad-Toh, K.; Haerty, W.; Di Palma, F.; Regev, A. A quantitative framework for characterizing the evolutionary history of mammalian gene expression. Genome Res. 2019, 29, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| vmax [pmol × min−1 × mg Protein−1] ± SEM | KM [µM] ± SEM | CLint [µL × min−1 × mg Protein−1] ± SEM | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Substrate | dOCT1 | hOCT1 | mOCT1 | dOCT1 | hOCT1 | mOCT1 | dOCT1 | hOCT1 | mOCT1 |
| Fenoterol | 1346 ± 134 | 106 ± 11.8 *** | 912 ± 78.3 * | 16.8 ± 1.09 | 1.19 ± 0.09 *** | 10.7 ± 1.09 * | 80.7 ± 7.76 | 88.3 ± 3.85 | 88.5 ± 15.5 |
| Ipratropium | 113 ± 1.43 | 586 ± 42.7 *** | 309 ± 18.8 * | 1.02 ± 0.20 | 7.79 ± 0.48 *** | 3.77 ± 0.31 ** | 122 ± 28.1 | 75.7 ± 6.33 | 83.6 ± 10.4 |
| Trospium | 835 ± 64.9 | 1503 ± 103 *** | 255 ± 33.2 *** | 5.72 ± 0.30 | 13.3 ± 1.58 ** | 1.92 ± 0.85 | 146 ± 10.3 | 117 ± 12.7 | 180 ± 37.5 |
| Metformin | 11,373 ± 714 | 10,465 ± 1306 | 10,891 ± 1249 | 204 ± 41.8 | 1090 ± 297 | 298 ± 46.9 * | 63.1 ± 12.0 | 11.0 ± 2.05 ** | 38.7 ± 6.23 |
| Butylscopolamine | 1383 ± 112 | 1249 ± 117 | 778 ± 59.2 * | 18.7 ± 3.32 | 15.0 ± 1.67 | 6.60 ± 1.13 * | 80.5 ± 12.3 | 84.4 ± 6.76 | 131 ± 25.6 |
| vmax [pmol × min−1 × mg Protein−1] ± SEM | KM [µM] ± SEM | CLint [µL × min−1 × mg Protein−1] ± SEM | ||||
|---|---|---|---|---|---|---|
| Substrate | dOCT2 | hOCT2 | dOCT2 | hOCT2 | dOCT2 | hOCT2 |
| Fenoterol | n.d. | 248 ± 21.7 | n.d. | 8.81 ± 0.55 | n.d. | 28.0 ± 0.95 |
| Ipratropium | 93.2 ± 12.6 | 114 ± 10.5 | 0.35 ± 0.02 | 0.24 ± 0.04 * | 265 ± 35.3 | 477 ± 31.9 * |
| Trospium | 224 ± 14.6 | 119 ± 13.1 ** | 2.18 ± 0.27 | 0.49 ± 0.15 ** | 106 ± 8.78 | 278 ± 40.5 * |
| Metformin | 21,578 ± 606 | 29,650 ± 1457 * | 365 ± 11.5 | 766 ± 95.4 * | 59.2 ± 2.76 | 39.9 ± 5.27 * |
| Butylscopolamine | n.d. | 616 ± 52.0 | n.d. | 7.62 ± 0.85 | n.d. | 81.6 ± 2.16 |
| Primer | Sequence (5′-3′) | Annealing Temperature [°C] |
|---|---|---|
| dOCT1_HindIII_for dOCT1_EcoRV_rev | GTGATGAAGCTTCTGGCTCCGTTATGCCCACCG | 70 |
| CCGAGCGATATCTCTCTCTCAGGTGCCGGCACG | ||
| dOCT2_for dOCT2_rev | AGCATCGGAAGCTTTCCTGCCTCCGGAGATAATGCCAACT | 50 |
| GTATGGAGGATATCAGCTCCCTACCTCTGCATGTTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meyer, M.J.; Falk, S.; Römer, S.; Prinzinger, C.; Tacke, S.; Geyer, J.; Simm, S.; Tzvetkov, M.V. Cloning and Functional Characterization of Dog OCT1 and OCT2: Another Step in Exploring Species Differences in Organic Cation Transporters. Int. J. Mol. Sci. 2022, 23, 5100. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095100
Meyer MJ, Falk S, Römer S, Prinzinger C, Tacke S, Geyer J, Simm S, Tzvetkov MV. Cloning and Functional Characterization of Dog OCT1 and OCT2: Another Step in Exploring Species Differences in Organic Cation Transporters. International Journal of Molecular Sciences. 2022; 23(9):5100. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095100
Chicago/Turabian StyleMeyer, Marleen Julia, Simon Falk, Sarah Römer, Clarissa Prinzinger, Sabine Tacke, Joachim Geyer, Stefan Simm, and Mladen Vassilev Tzvetkov. 2022. "Cloning and Functional Characterization of Dog OCT1 and OCT2: Another Step in Exploring Species Differences in Organic Cation Transporters" International Journal of Molecular Sciences 23, no. 9: 5100. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095100