
Activation of Free Fatty Acid Receptor 4 (FFA4) Ameliorates Ovalbumin-Induced Allergic Asthma by Suppressing Activation of Dendritic and Mast Cells in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
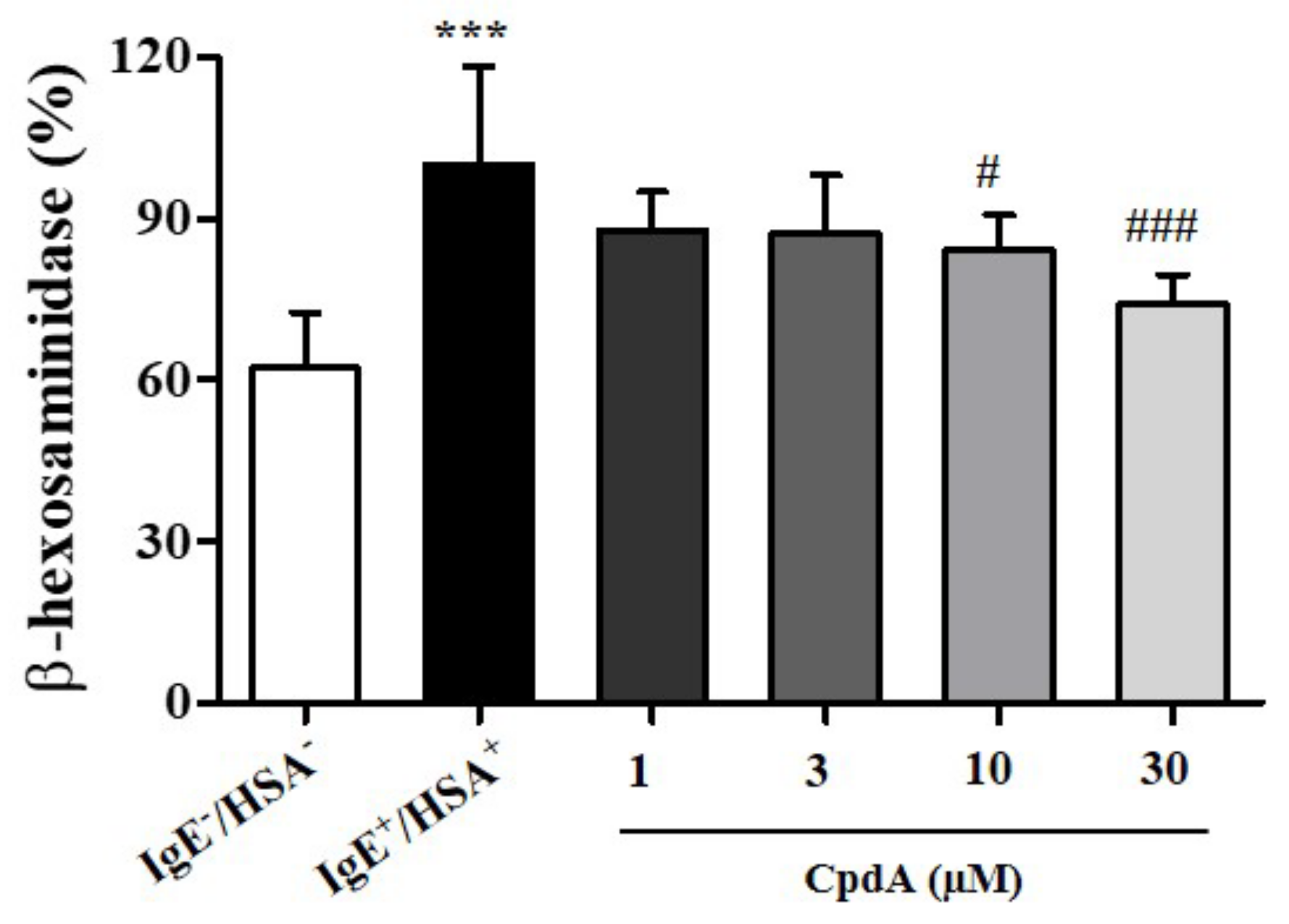
2.1. Compound A Suppressed Degranulation of Mast Cells
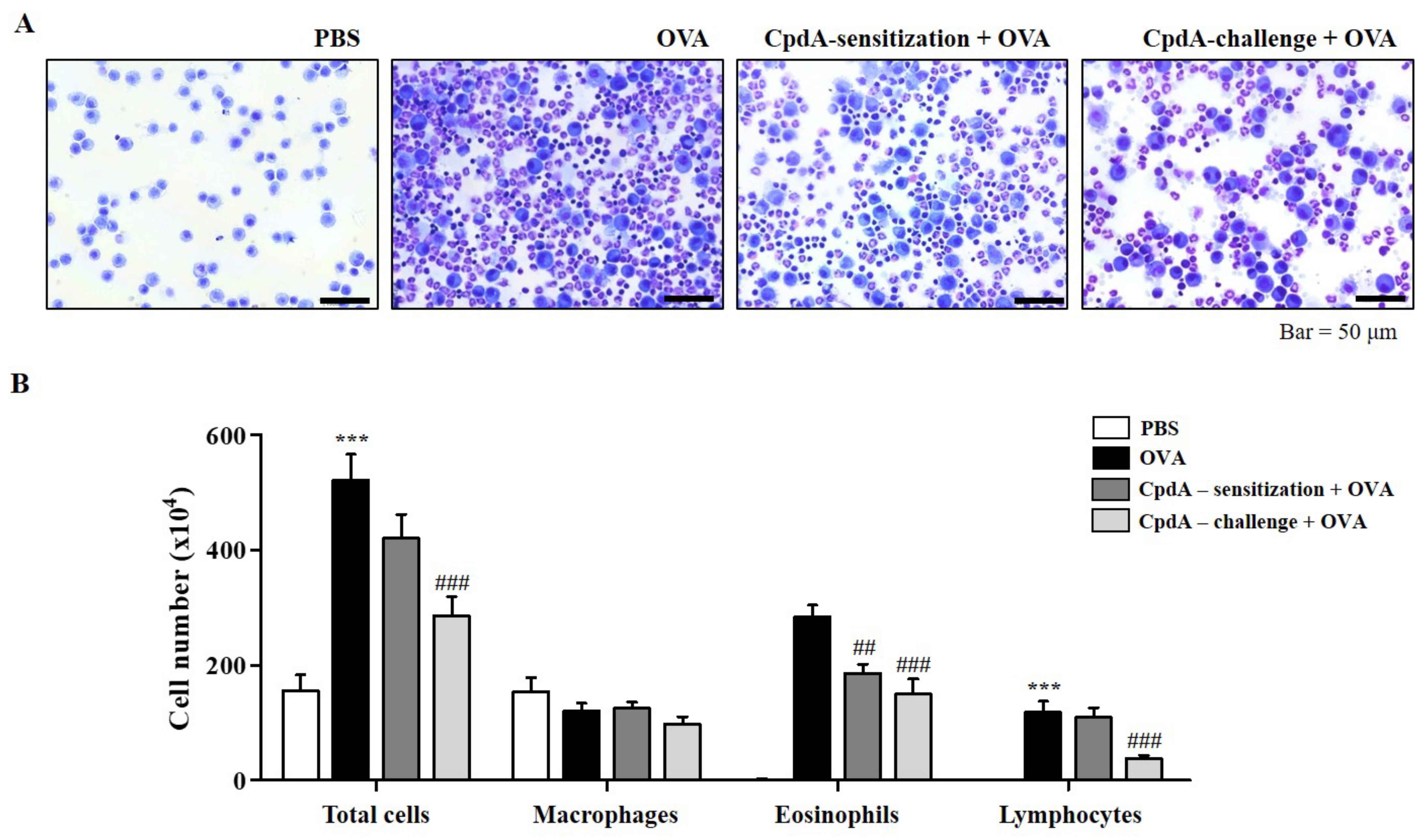
2.2. Compound A Ameliorated Asthma-Induced Influx of Immune Cells in FFA4-WT Mice
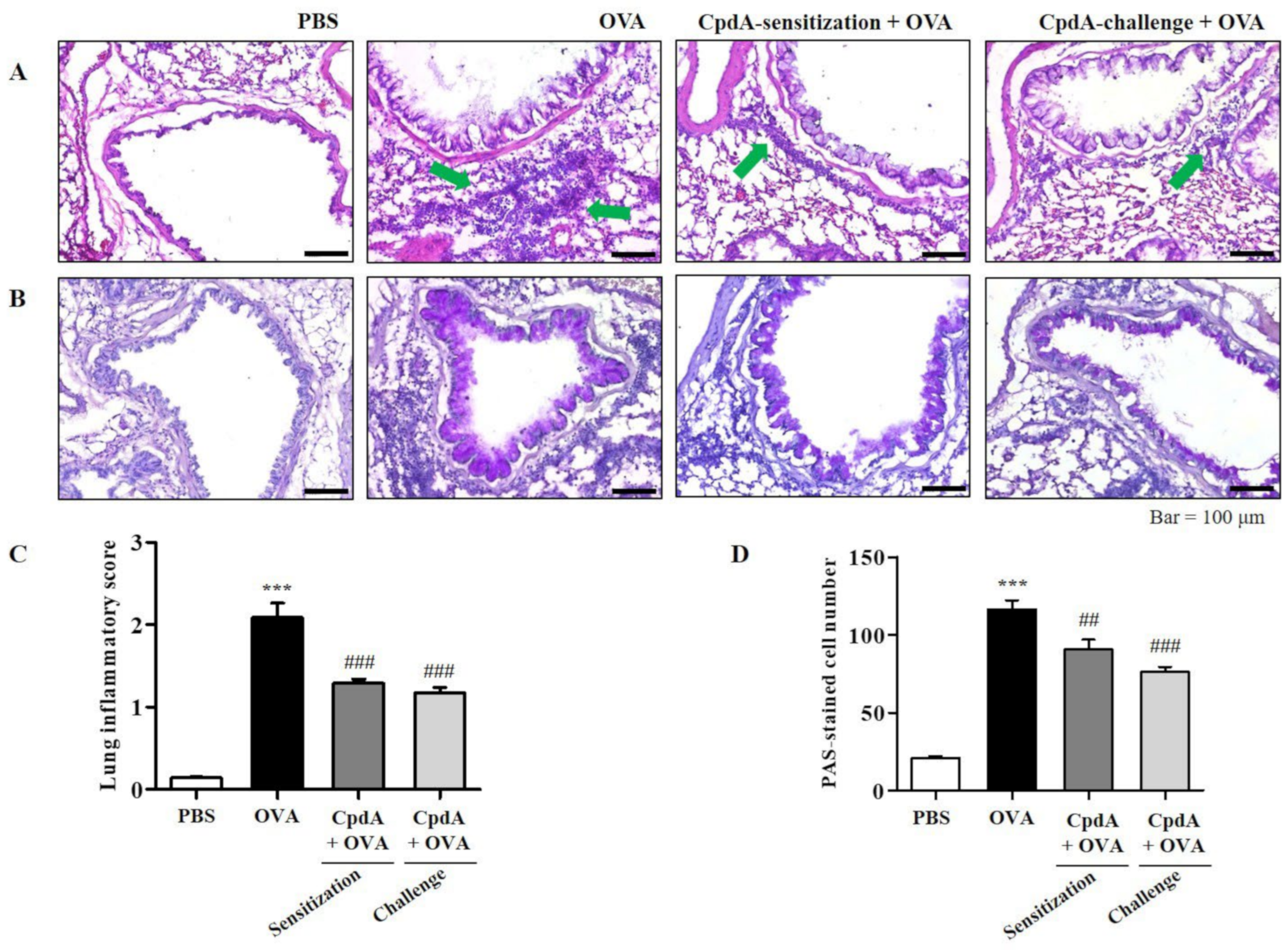
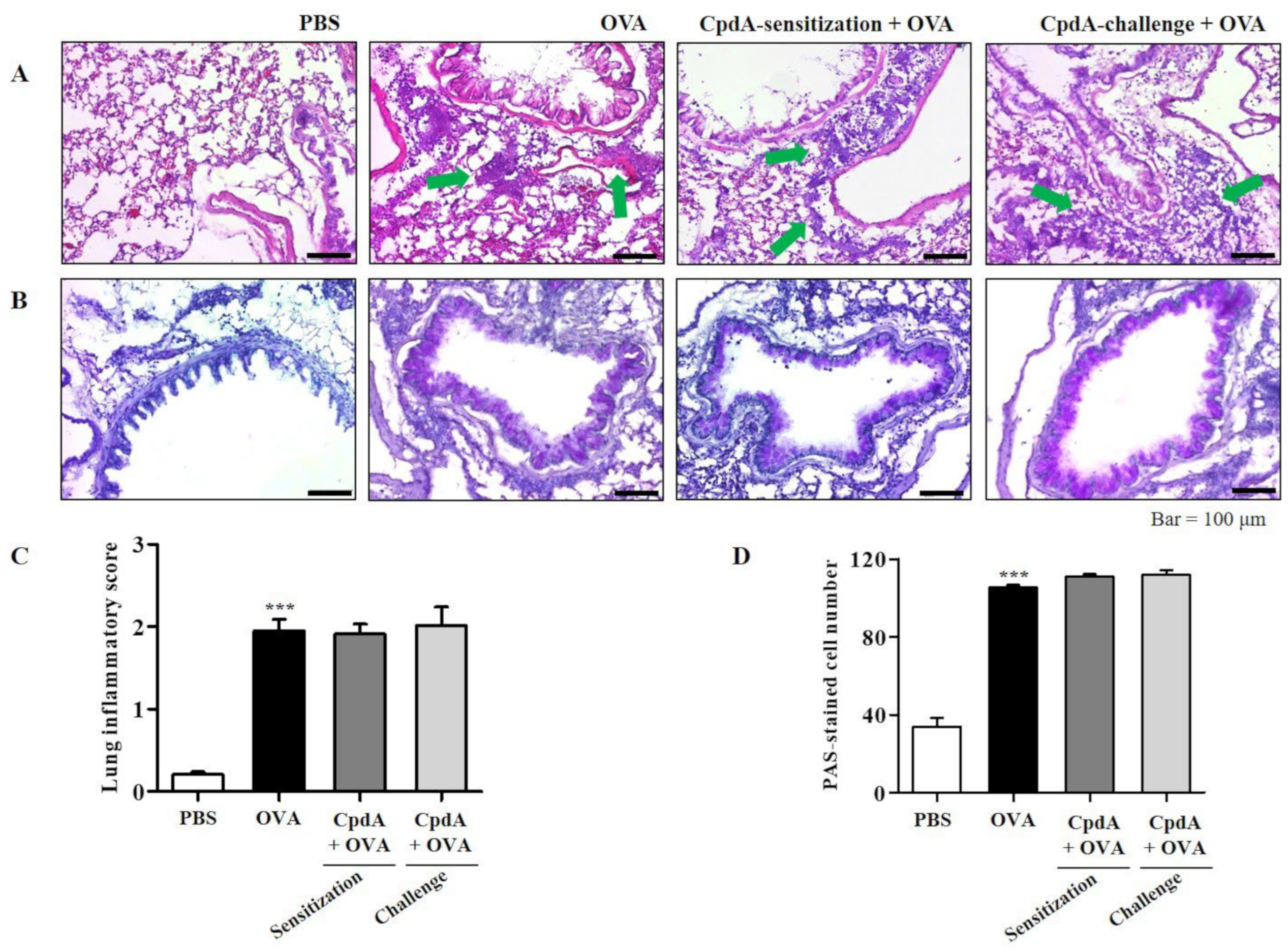
2.3. Compound A Ameliorated Asthma-Induced Inflammation and Mucin Secretion in the Lungs of FFA4-WT Mice
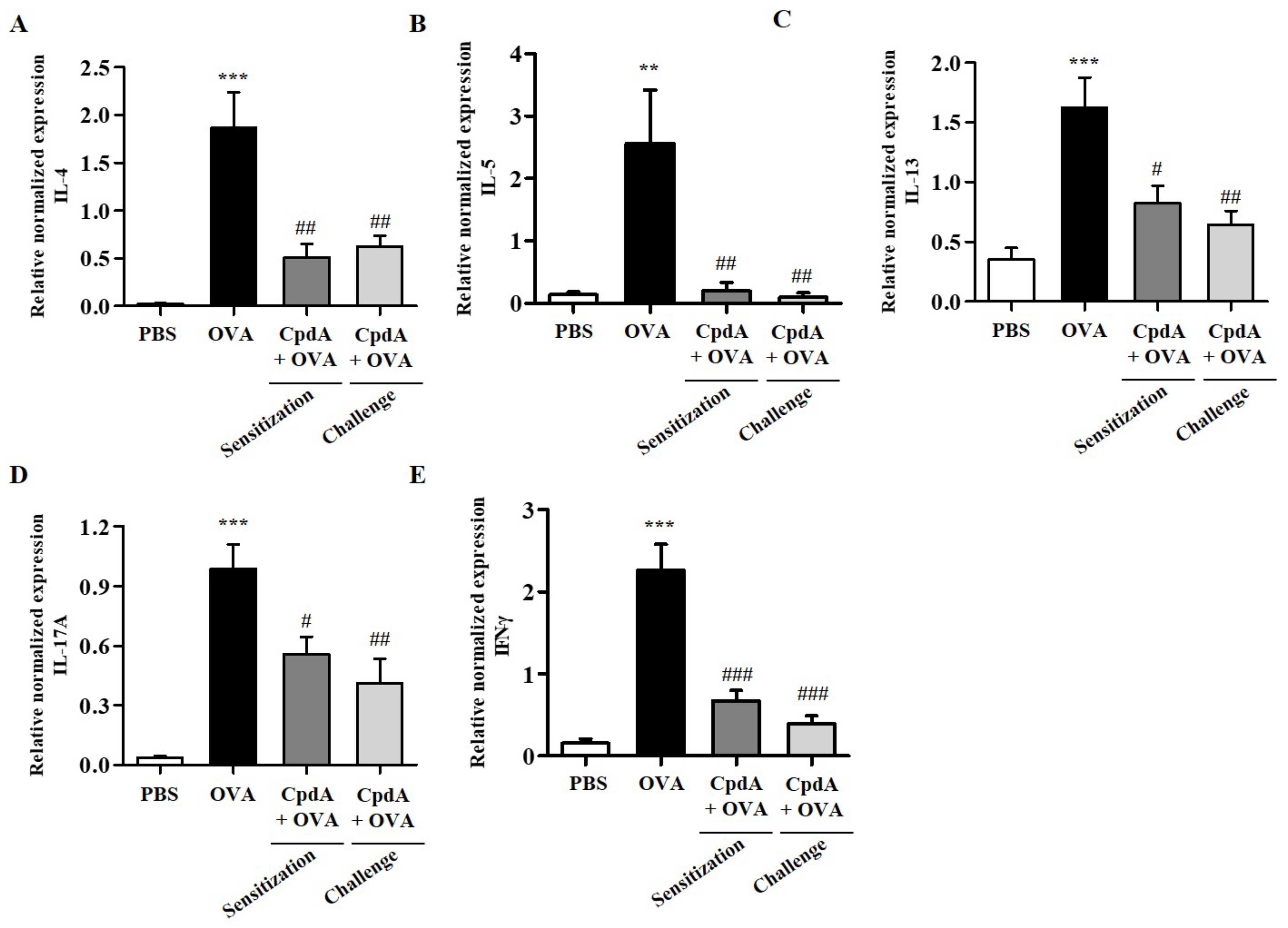
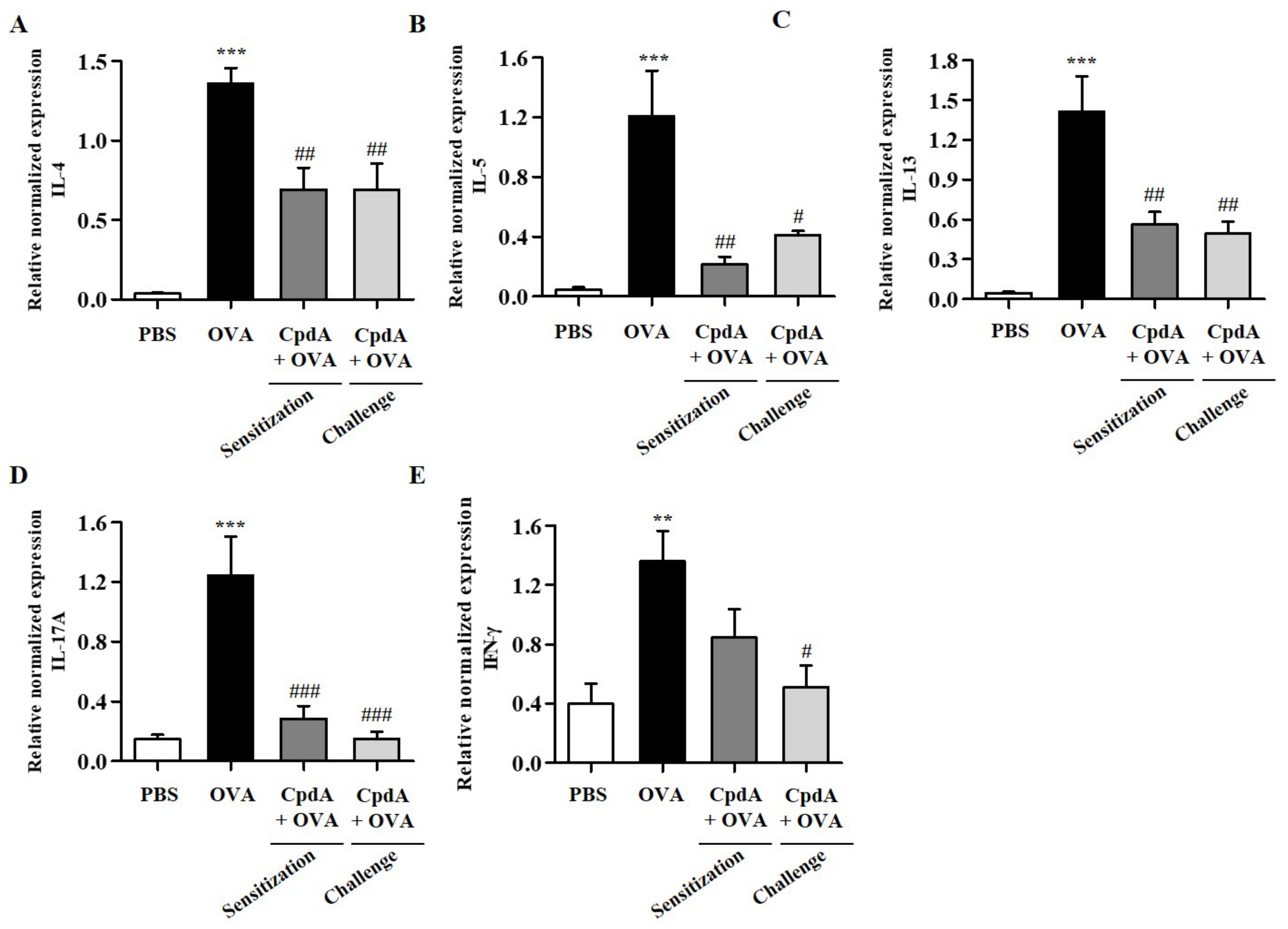
2.4. Compound A Ameliorated Asthma-Induced Cytokine Expression in FFA4-WT Mice
2.5. Compound A Did Not Ameliorate Asthma-Induced Influx of Immune Cells in FFA4-KO Mice
2.6. Compound A Did Not Ameliorate Asthma-Induced Inflammation and Mucin Secretion in the Lungs of FFA4-KO Mice
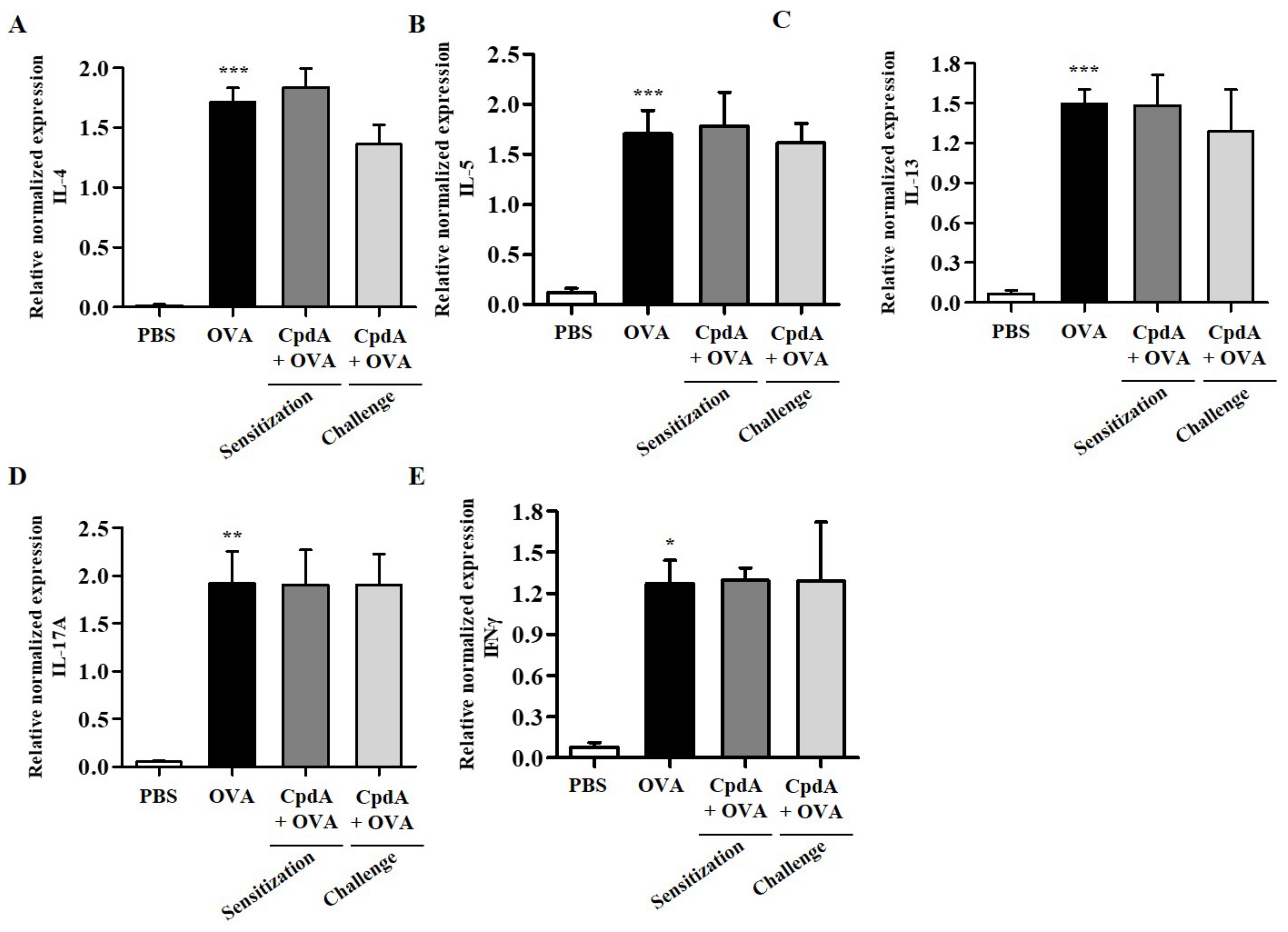
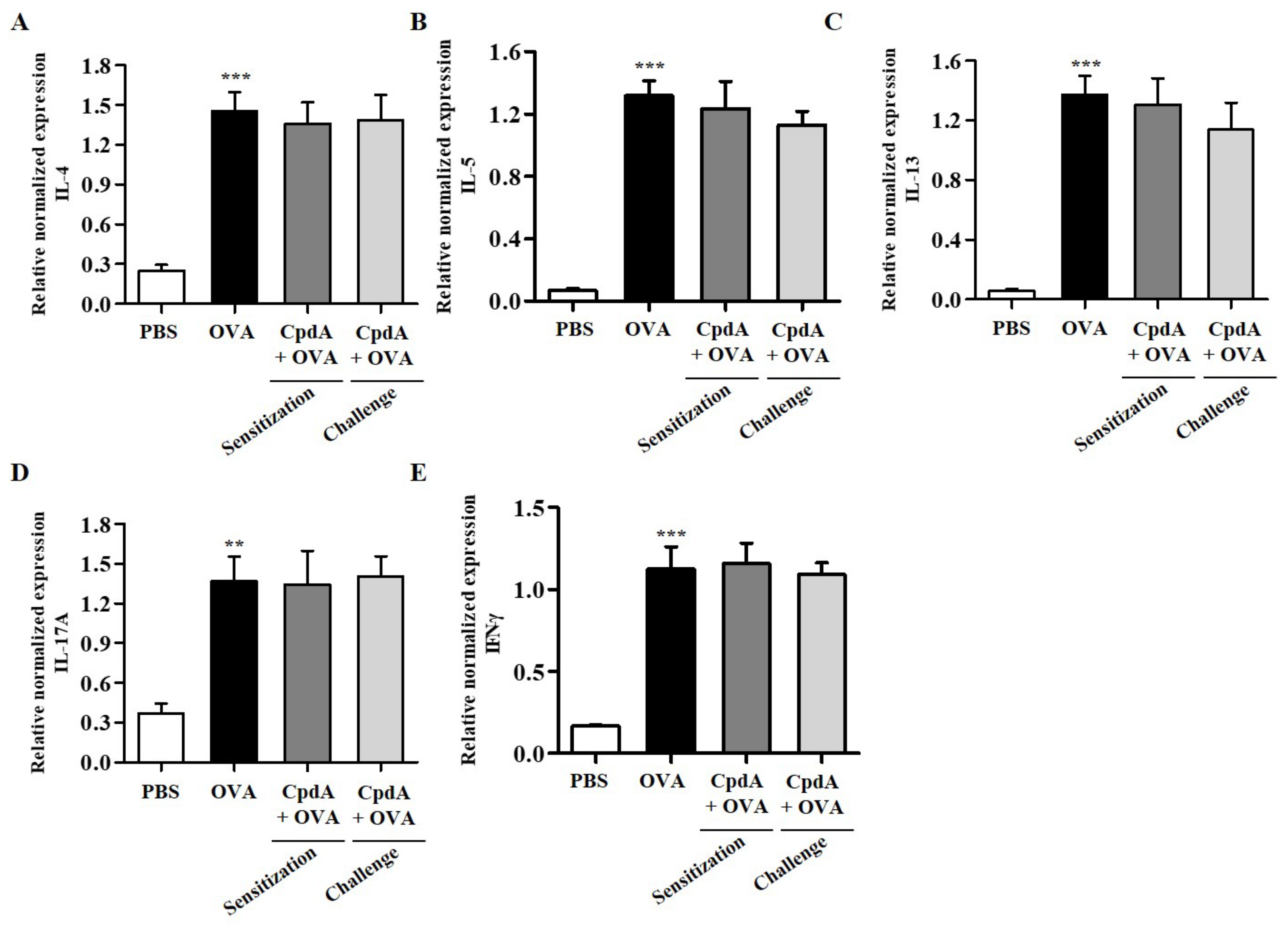
2.7. Compound A Did Not Ameliorate Asthma-Induced Cytokine Expression in FFA4-KO Mice
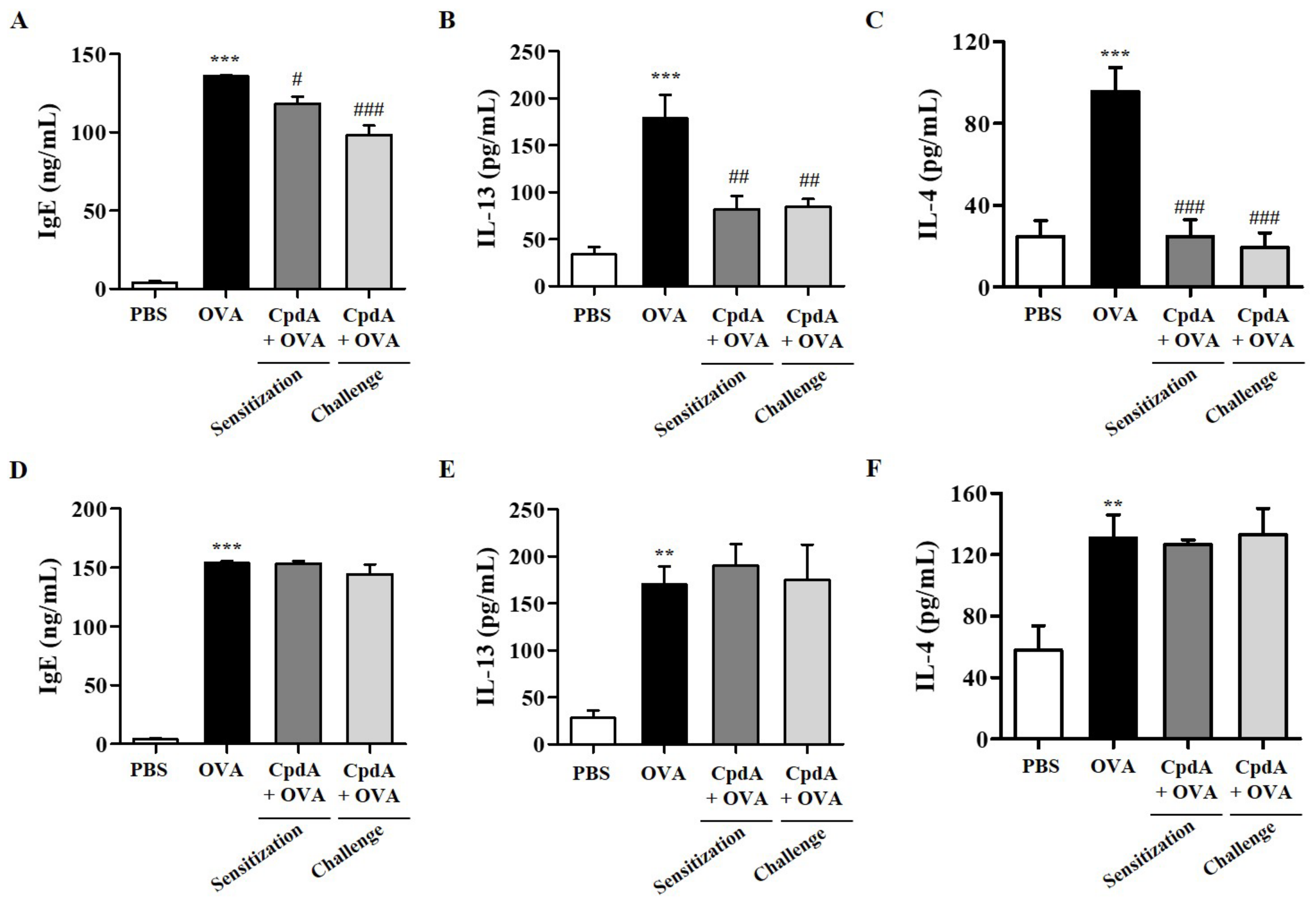
2.8. Changes of Serum IgE and BALF Cytokine Levels by OVA and Compound A in Mice
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals
4.3. Cell Culture
4.4. Assessment of Degranulation
4.5. Asthma Induction in Mice and Administration of Compound A
4.6. Cell Counting and Analysis in BALF
4.7. Histological Examination of the Lungs
4.8. Quantitative Real-Time PCR
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barnes, P.J. Targeting cytokines to treat asthma and chronic obstructive pulmonary disease. Nat. Rev. Immunol. 2018, 18, 454–466. [Google Scholar] [CrossRef] [PubMed]
- Shehibi, N.M.A.A.; Ibrahim, K.M.S.; Salam, M.M.A.; Mohamed, R.M. Assessment of Response to Anti Leukotriene Among Children with Bronchial Asthma. Eur. J. Mol. Clin. Med. 2021, 8, 3000–3008. [Google Scholar]
- Horrobin, D.F. Low prevalences of coronary heart disease (CHD), psoriasis, asthma and rheumatoid arthritis in Eskimos: Are they caused by high dietary intake of eicosapentaenoic acid (EPA), a genetic variation of essential fatty acid (EFA) metabolism or a combination of both? Med. Hypotheses 1987, 22, 421–428. [Google Scholar] [PubMed]
- Schubert, R.; Kitz, R.; Beermann, C.; Rose, M.; Lieb, A.; Sommerer, P.; Moskovits, J.; Alberternst, H.; Böhles, H.; Schulze, J. Effect of n–3 polyunsaturated fatty acids in asthma after low-dose allergen challenge. Int. Arch. Allergy Immunol. 2009, 148, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Dry, J.; Vincent, D. Effect of a fish oil diet on asthma: Results of a 1-year double-blind study. Int. Arch. Allergy Appl. Immunol. 1991, 95, 156–157. [Google Scholar] [CrossRef]
- Nagakura, T.; Matsuda, S.; Shichijyo, K.; Sugimoto, H.; Hata, K. Dietary supplementation with fish oil rich in omega-3 polyunsaturated fatty acids in children with bronchial asthma. Eur. Respir. J. 2000, 16, 861–865. [Google Scholar] [CrossRef] [Green Version]
- Thien, F.C.; Mencia-Huerta, J.M.; Lee, T.H. Dietary fish oil effects on seasonal hay fever and asthma in pollen-sensitive subjects. Am. Rev. Respir. Dis. 1993, 147, 1138–1143. [Google Scholar] [CrossRef]
- Mickleborough, T.D.; Rundell, K.W. Dietary polyunsaturated fatty acids in asthma- and exercise-induced bronchoconstriction. Eur. J. Clin. Nutr. 2005, 59, 1335–1346. [Google Scholar] [CrossRef] [Green Version]
- Im, D.-S. Omega-3 fatty acids in anti-inflammation (pro-resolution) and GPCRs. Prog. Lipid Res. 2012, 51, 232–237. [Google Scholar] [CrossRef]
- Hirasawa, A.; Tsumaya, K.; Awaji, T.; Katsuma, S.; Adachi, T.; Yamada, M.; Sugimoto, Y.; Miyazaki, S.; Tsujimoto, G. Free fatty acids regulate gut incretin glucagon-like peptide-1 secretion through GPR120. Nat. Med. 2005, 11, 90–94. [Google Scholar] [CrossRef]
- Oh, D.Y.; Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Im, D.S. FFA4 (GPR120) as a fatty acid sensor involved in appetite control, insulin sensitivity and inflammation regulation. Mol. Asp. Med. 2018, 64, 92–108. [Google Scholar] [CrossRef] [PubMed]
- Talukdar, S.; Olefsky, J.M.; Osborn, O. Targeting GPR120 and other fatty acid-sensing GPCRs ameliorates insulin resistance and inflammatory diseases. Trends Pharmacol. Sci. 2011, 32, 543–550. [Google Scholar] [CrossRef] [Green Version]
- Moniri, N.H. Free-fatty acid receptor-4 (GPR120): Cellular and molecular function and its role in metabolic disorders. Biochem. Pharmacol. 2016, 110–111, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Yore, M.M.; Syed, I.; Moraes-Vieira, P.M.; Zhang, T.; Herman, M.A.; Homan, E.A.; Patel, R.T.; Lee, J.; Chen, S.; Peroni, O.D.; et al. Discovery of a class of endogenous mammalian lipids with anti-diabetic and anti-inflammatory effects. Cell 2014, 159, 318–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konno, Y.; Ueki, S.; Takeda, M.; Kobayashi, Y.; Tamaki, M.; Moritoki, Y.; Oyamada, H.; Itoga, M.; Kayaba, H.; Omokawa, A.; et al. Functional analysis of free fatty acid receptor GPR120 in human eosinophils: Implications in metabolic homeostasis. PLoS ONE 2015, 10, e0120386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, D.Y.; Walenta, E.; Akiyama, T.E.; Lagakos, W.S.; Lackey, D.; Pessentheiner, A.R.; Sasik, R.; Hah, N.; Chi, T.J.; Cox, J.M.; et al. A Gpr120-selective agonist improves insulin resistance and chronic inflammation in obese mice. Nat. Med. 2014, 20, 942–947. [Google Scholar] [CrossRef] [Green Version]
- Cox, J.M.; Chu, H.D.; Chelliah, M.V.; Debenham, J.S.; Eagen, K.; Lan, P.; Lombardo, M.; London, C.; Plotkin, M.A.; Shah, U.; et al. Design, Synthesis, and Evaluation of Novel and Selective G-protein Coupled Receptor 120 (GPR120) Spirocyclic Agonists. ACS Med. Chem. Lett. 2017, 8, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Kolkhir, P.; Elieh-Ali-Komi, D.; Metz, M.; Siebenhaar, F.; Maurer, M. Understanding human mast cells: Lesson from therapies for allergic and non-allergic diseases. Nat. Rev. Immunol. 2021, 22, 294–308. [Google Scholar] [CrossRef]
- Carroll-Portillo, A.; Spendier, K.; Pfeiffer, J.; Griffiths, G.; Li, H.; Lidke, K.A.; Oliver, J.M.; Lidke, D.S.; Thomas, J.L.; Wilson, B.S. Formation of a mast cell synapse: FcεRI membrane dynamics upon binding mobile or immobilized ligands on surfaces. J. Immunol. 2010, 184, 1328–1338. [Google Scholar] [CrossRef] [Green Version]
- Jiménez, M.; Cervantes-García, D.; Córdova-Dávalos, L.E.; Pérez-Rodríguez, M.J.; Gonzalez-Espinosa, C.; Salinas, E. Responses of mast cells to pathogens: Beneficial and detrimental roles. Front. Immunol. 2021, 12, 685865. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ilarraza, R.; Tancowny, B.P.; Alam, S.B.; Kulka, M. Disrupted Lipid Raft Shuttling of FcεRI by n-3 Polyunsaturated Fatty Acid Is Associated With Ligation of G Protein-Coupled Receptor 120 (GPR120) in Human Mast Cell Line LAD2. Front. Nutr. 2020, 7, 253. [Google Scholar] [CrossRef] [PubMed]
- Hammad, H.; Lambrecht, B.N. The basic immunology of asthma. Cell 2021, 184, 1469–1485. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Athari, S.S.; Mehrabi Nasab, E.; Zhao, L. PI3K/AKT/mTOR and TLR4/MyD88/NF-κB signaling inhibitors attenuate pathological mechanisms of allergic asthma. Inflammation 2021, 44, 1895–1907. [Google Scholar] [CrossRef] [PubMed]
- Johansson, S.; Lönnqvist, A.; Ostman, S.; Sandberg, A.S.; Wold, A.E. Long-chain polyunsaturated fatty acids are consumed during allergic inflammation and affect T helper type 1 (Th1)- and Th2-mediated hypersensitivity differently. Clin. Exp. Immunol. 2010, 160, 411–419. [Google Scholar] [CrossRef]
- Arm, J.P.; Horton, C.E.; Spur, B.W.; Mencia-Huerta, J.-M.; Lee, T.H. The effects of dietary supplementation with fish oil lipids on the airways response to inhaled allergen in bronchial asthma. Am. J. Respir. Crit. Care Med. 1989, 139, 1395–1400. [Google Scholar] [CrossRef]
- Yokoyama, A.; Hamazaki, T.; Ohshita, A.; Kohno, N.; Sakai, K.; Zhao, G.D.; Katayama, H.; Hiwada, K. Effect of aerosolized docosahexaenoic acid in a mouse model of atopic asthma. Int. Arch. Allergy Immunol. 2000, 123, 327–332. [Google Scholar] [CrossRef]
- Giudetti, A.M.; Cagnazzo, R. Beneficial effects of n-3 PUFA on chronic airway inflammatory diseases. Prostaglandins Other Lipid Mediat. 2012, 99, 57–67. [Google Scholar] [CrossRef]
- Levy, B.D.; Kohli, P.; Gotlinger, K.; Haworth, O.; Hong, S.; Kazani, S.; Israel, E.; Haley, K.J.; Serhan, C.N. Protectin D1 is generated in asthma and dampens airway inflammation and hyperresponsiveness. J. Immunol. 2007, 178, 496–502. [Google Scholar] [CrossRef] [Green Version]
- Aoki, H.; Hisada, T.; Ishizuka, T.; Utsugi, M.; Kawata, T.; Shimizu, Y.; Okajima, F.; Dobashi, K.; Mori, M. Resolvin E1 dampens airway inflammation and hyperresponsiveness in a murine model of asthma. Biochem. Biophys. Res. Commun. 2008, 367, 509–515. [Google Scholar] [CrossRef]
- Aoki, H.; Hisada, T.; Ishizuka, T.; Utsugi, M.; Ono, A.; Koga, Y.; Sunaga, N.; Nakakura, T.; Okajima, F.; Dobashi, K.; et al. Protective effect of resolvin E1 on the development of asthmatic airway inflammation. Biochem. Biophys. Res. Commun. 2010, 400, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Haworth, O.; Cernadas, M.; Yang, R.; Serhan, C.N.; Levy, B.D. Resolvin E1 regulates interleukin 23, interferon-gamma and lipoxin A4 to promote the resolution of allergic airway inflammation. Nat. Immunol. 2008, 9, 873–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Ma, D.W.; Kang, J.X.; Kulka, M. n-3 Polyunsaturated fatty acids inhibit Fc ε receptor I-mediated mast cell activation. J. Nutr. Biochem. 2015, 26, 1580–1588. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Lee, Y.E.; Yeon, J.W.; Go, G.-Y.; Byun, J.; Lee, K.; Lee, H.K.; Hur, J.K.; Jang, M.; Kim, T.H. A novel therapeutic modality using CRISPR-engineered dendritic cells to treat allergies. Biomaterials 2021, 273, 120798. [Google Scholar] [CrossRef]
- Zeyda, M.; Säemann, M.D.; Stuhlmeier, K.M.; Mascher, D.G.; Nowotny, P.N.; Zlabinger, G.J.; Waldhäusl, W.; Stulnig, T.M. Polyunsaturated fatty acids block dendritic cell activation and function independently of NF-κB activation. J. Biol. Chem. 2005, 280, 14293–14301. [Google Scholar] [CrossRef] [Green Version]
- Weatherill, A.R.; Lee, J.Y.; Zhao, L.; Lemay, D.G.; Youn, H.S.; Hwang, D.H. Saturated and polyunsaturated fatty acids reciprocally modulate dendritic cell functions mediated through TLR4. J. Immunol. 2005, 174, 5390–5397. [Google Scholar] [CrossRef]
- Wang, H.; Hao, Q.; Li, Q.-R.; Yan, X.-W.; Ye, S.; Li, Y.-S.; Li, N.; Li, J.-S. ω-3 Polyunsaturated fatty acids affect lipopolysaccharide-induced maturation of dendritic cells through mitogen-activated protein kinases p38. Nutrition 2007, 23, 474–482. [Google Scholar] [CrossRef]
- Draper, E.; Reynolds, C.M.; Canavan, M.; Mills, K.H.; Loscher, C.E.; Roche, H.M. Omega-3 fatty acids attenuate dendritic cell function via NF-κB independent of PPARγ. J. Nutr. Biochem. 2011, 22, 784–790. [Google Scholar] [CrossRef]
- Zhao, C.; Zhou, J.; Meng, Y.; Shi, N.; Wang, X.; Zhou, M.; Li, G.; Yang, Y. DHA sensor GPR120 in host defense exhibits the dual characteristics of regulating dendritic cell function and skewing the balance of Th17/Tregs. Int. J. Biol. Sci. 2020, 16, 374. [Google Scholar] [CrossRef] [Green Version]
- Teague, H.; Rockett, B.D.; Harris, M.; Brown, D.A.; Shaikh, S.R. Dendritic cell activation, phagocytosis and CD 69 expression on cognate T cells are suppressed by n-3 long-chain polyunsaturated fatty acids. Immunology 2013, 139, 386–394. [Google Scholar] [CrossRef]
- Feng, C.; Li, L.; Li, Q.; Switzer, K.; Liu, M.; Han, S.; Zheng, B. Docosahexaenoic acid ameliorates autoimmune inflammation by activating GPR120 signaling pathway in dendritic cells. Int. Immunopharmacol. 2021, 97, 107698. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.P.; Park, S.J.; Kang, S.; Koh, J.M.; Sato, K.; Chung, H.Y.; Okajima, F.; Im, D.S. omega-3 Polyunsaturated fatty acids accelerate airway repair by activating FFA4 in club cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2017, 312, L835–L844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Su, M.; Lee, B.K.; Kim, M.J.; Jung, J.H.; Im, D.S. Suppressive Effect of 4-Hydroxy-2-(4-Hydroxyphenethyl) Isoindoline-1,3-Dione on Ovalbumin-Induced Allergic Asthma. Biomol. Ther. 2018, 26, 539–545. [Google Scholar] [CrossRef]
- Kim, M.J.; Im, D.S. Suppressive effects of type I angiotensin receptor antagonists, candesartan and irbesartan on allergic asthma. Eur. J. Pharmacol. 2019, 852, 25–33. [Google Scholar] [CrossRef]
- Park, S.J.; Im, D.S. Blockage of sphingosine-1-phosphate receptor 2 attenuates allergic asthma in mice. Br. J. Pharmacol. 2019, 176, 938–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heo, J.Y.; Im, D.S. Anti-allergic effects of salvianolic acid A and tanshinone IIA from Salvia miltiorrhiza determined using in vivo and in vitro experiments. Int. Immunopharmacol. 2019, 67, 69–77. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Son, S.-E.; Koh, J.-M.; Im, D.-S. Activation of Free Fatty Acid Receptor 4 (FFA4) Ameliorates Ovalbumin-Induced Allergic Asthma by Suppressing Activation of Dendritic and Mast Cells in Mice. Int. J. Mol. Sci. 2022, 23, 5270. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095270
Son S-E, Koh J-M, Im D-S. Activation of Free Fatty Acid Receptor 4 (FFA4) Ameliorates Ovalbumin-Induced Allergic Asthma by Suppressing Activation of Dendritic and Mast Cells in Mice. International Journal of Molecular Sciences. 2022; 23(9):5270. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095270
Chicago/Turabian StyleSon, So-Eun, Jung-Min Koh, and Dong-Soon Im. 2022. "Activation of Free Fatty Acid Receptor 4 (FFA4) Ameliorates Ovalbumin-Induced Allergic Asthma by Suppressing Activation of Dendritic and Mast Cells in Mice" International Journal of Molecular Sciences 23, no. 9: 5270. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095270