
Regulation and Role of Neuron-Derived Hemoglobin in the Mouse Hippocampus


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
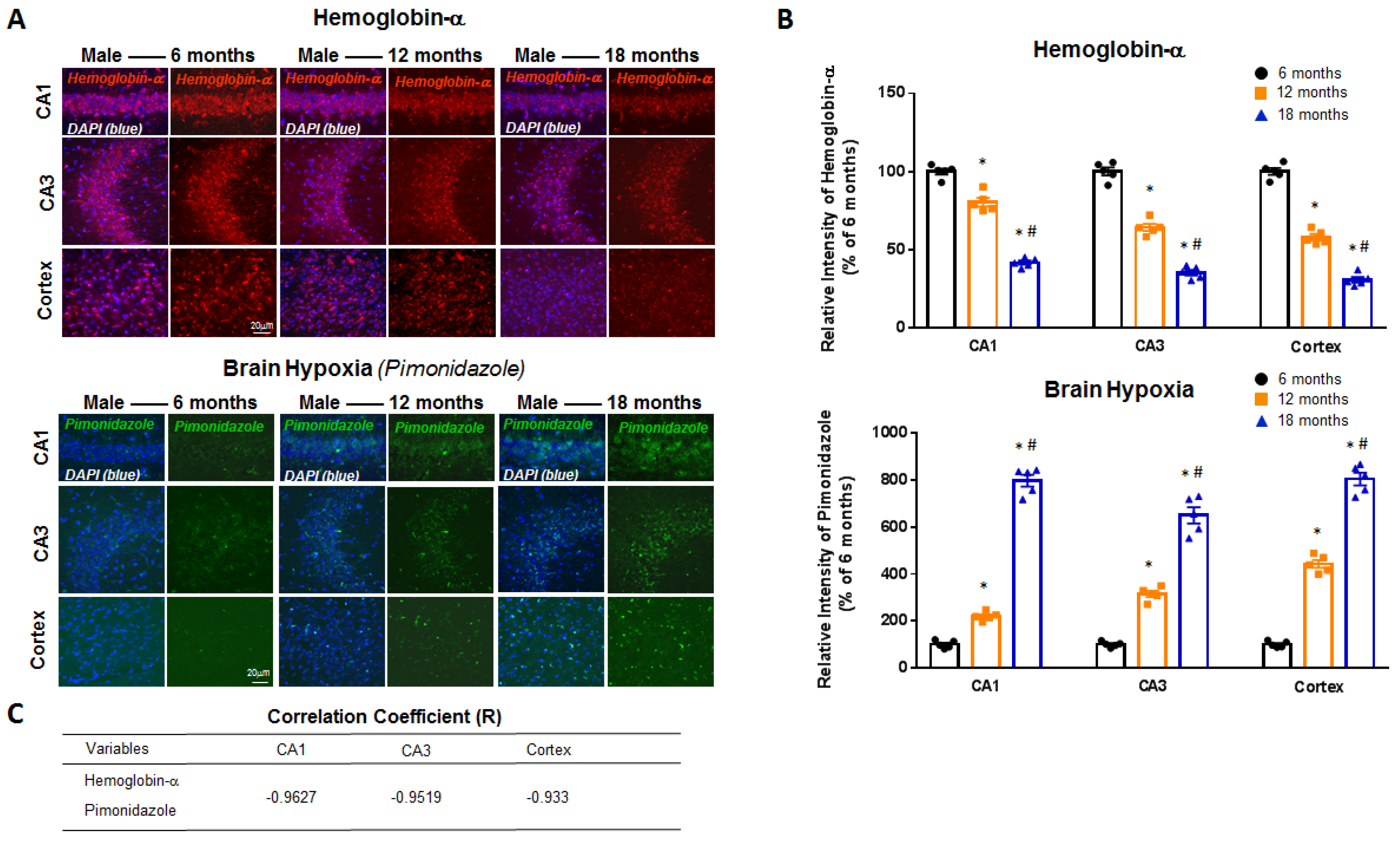
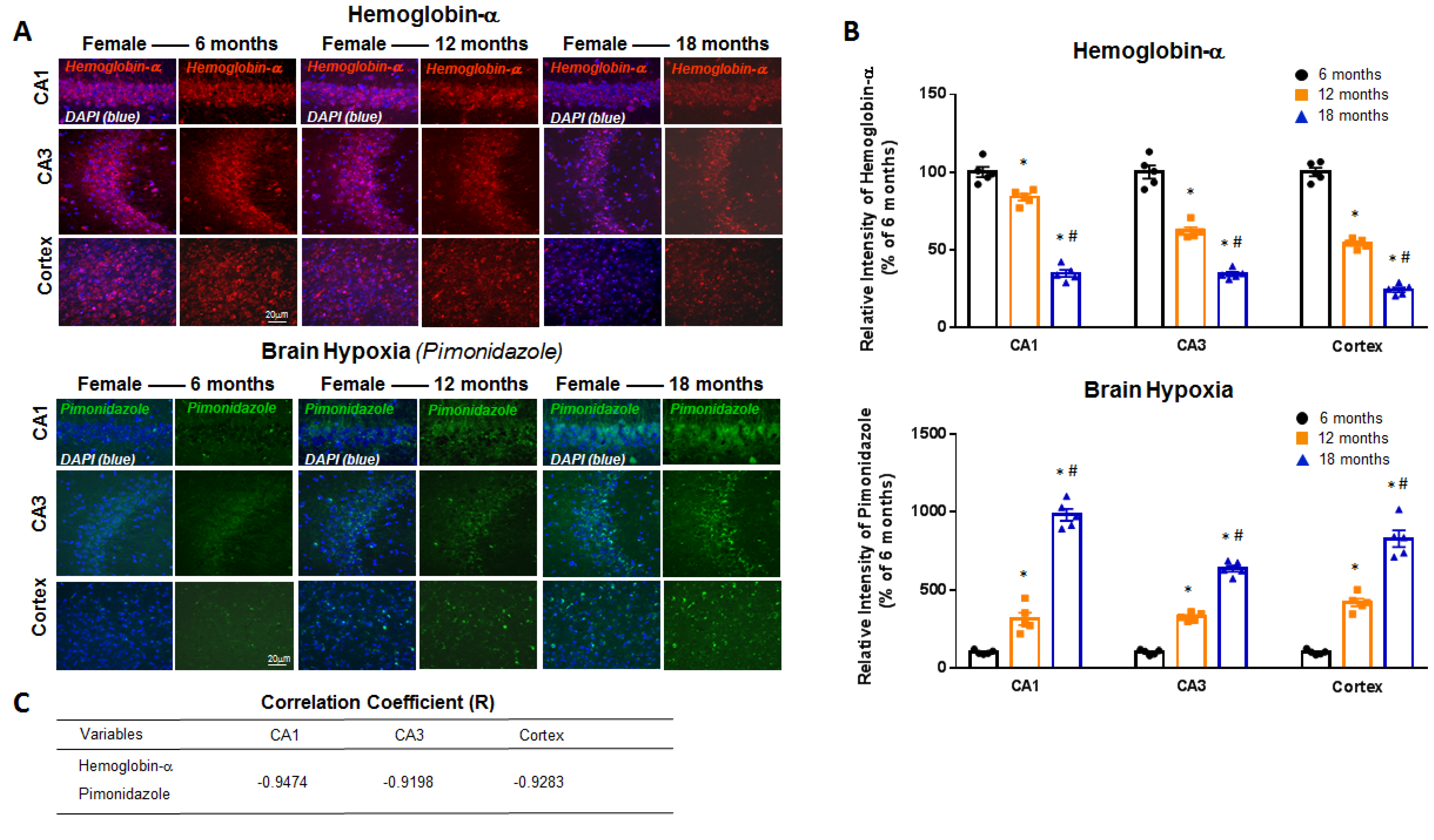
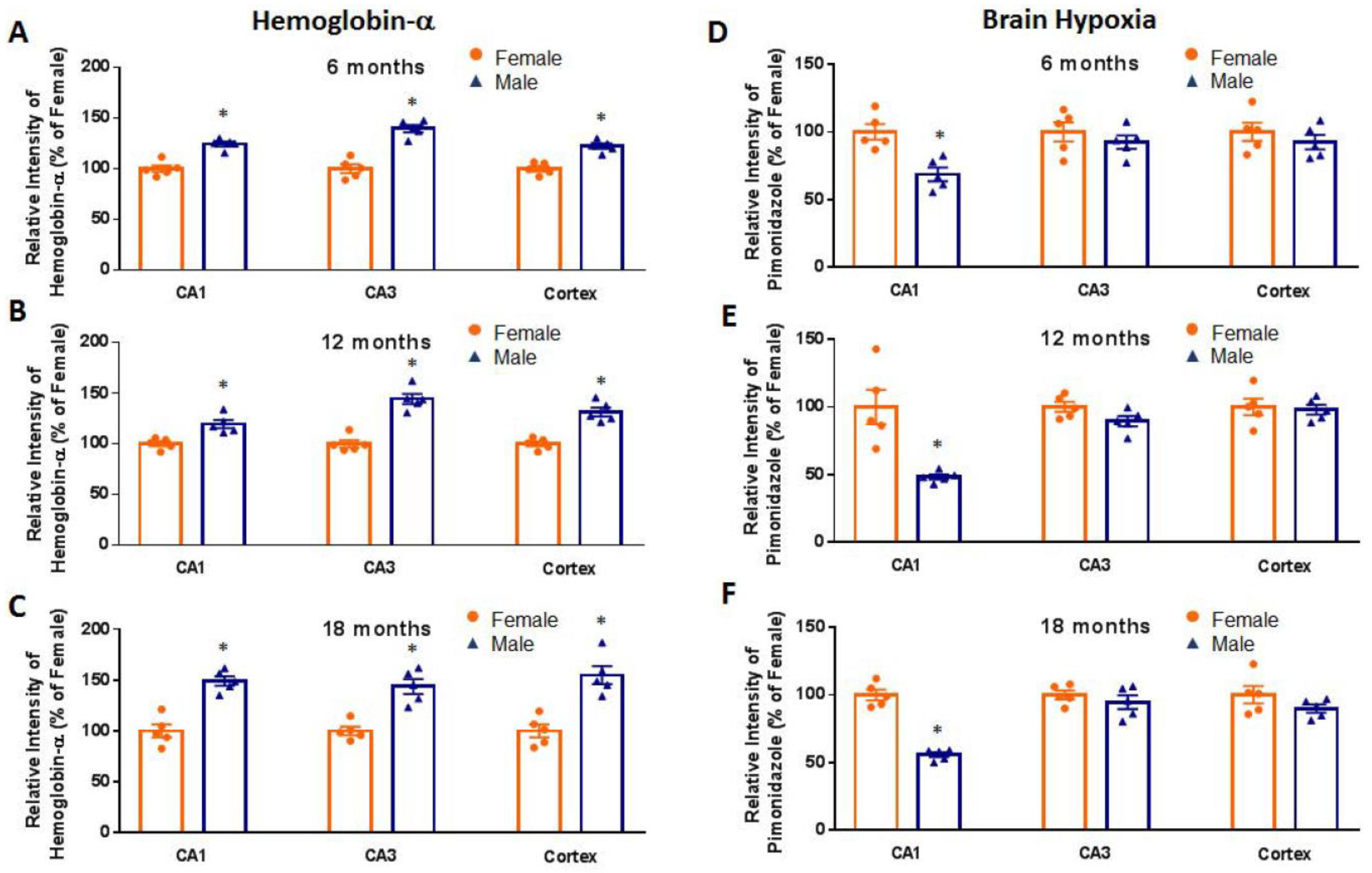
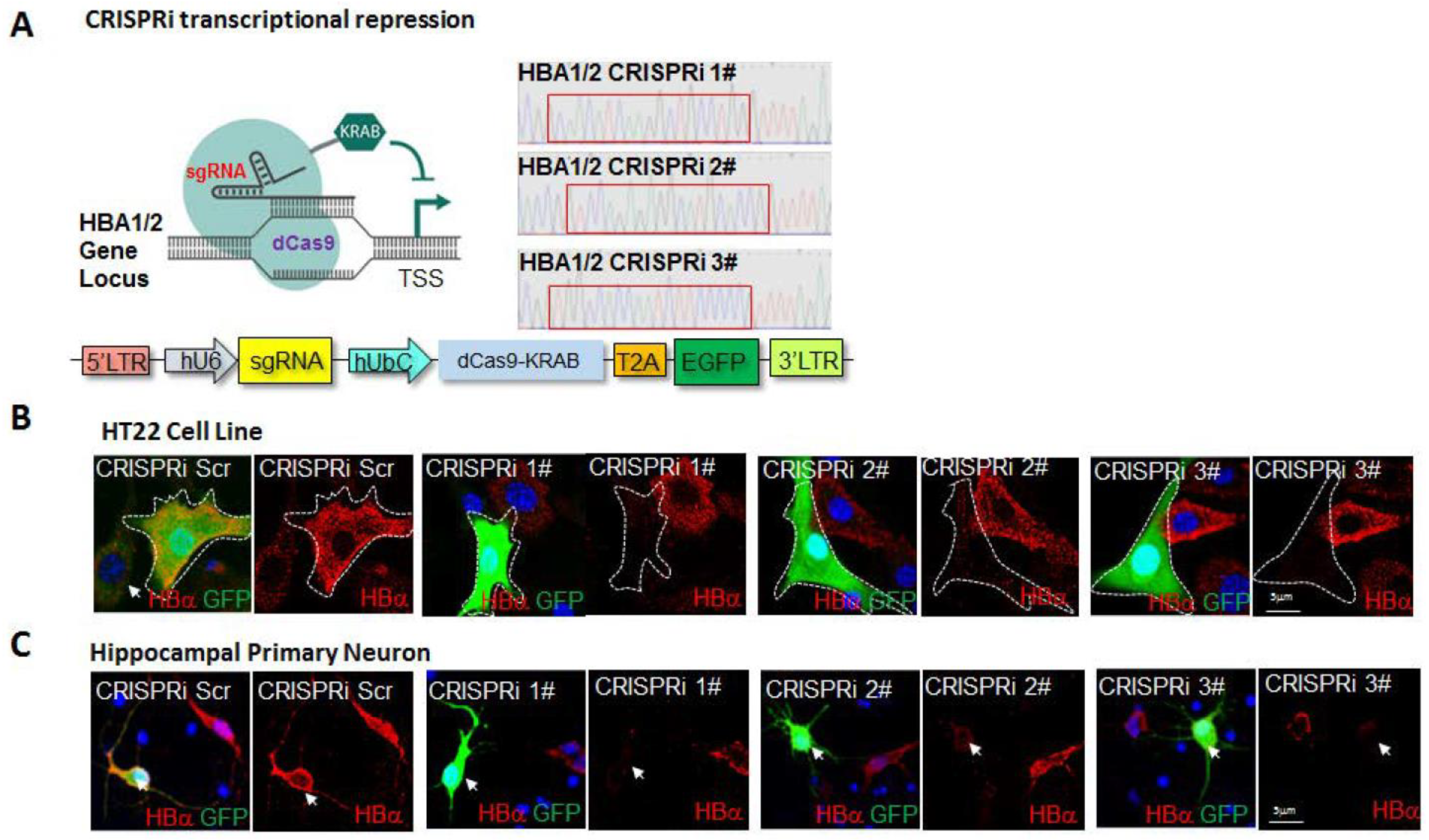
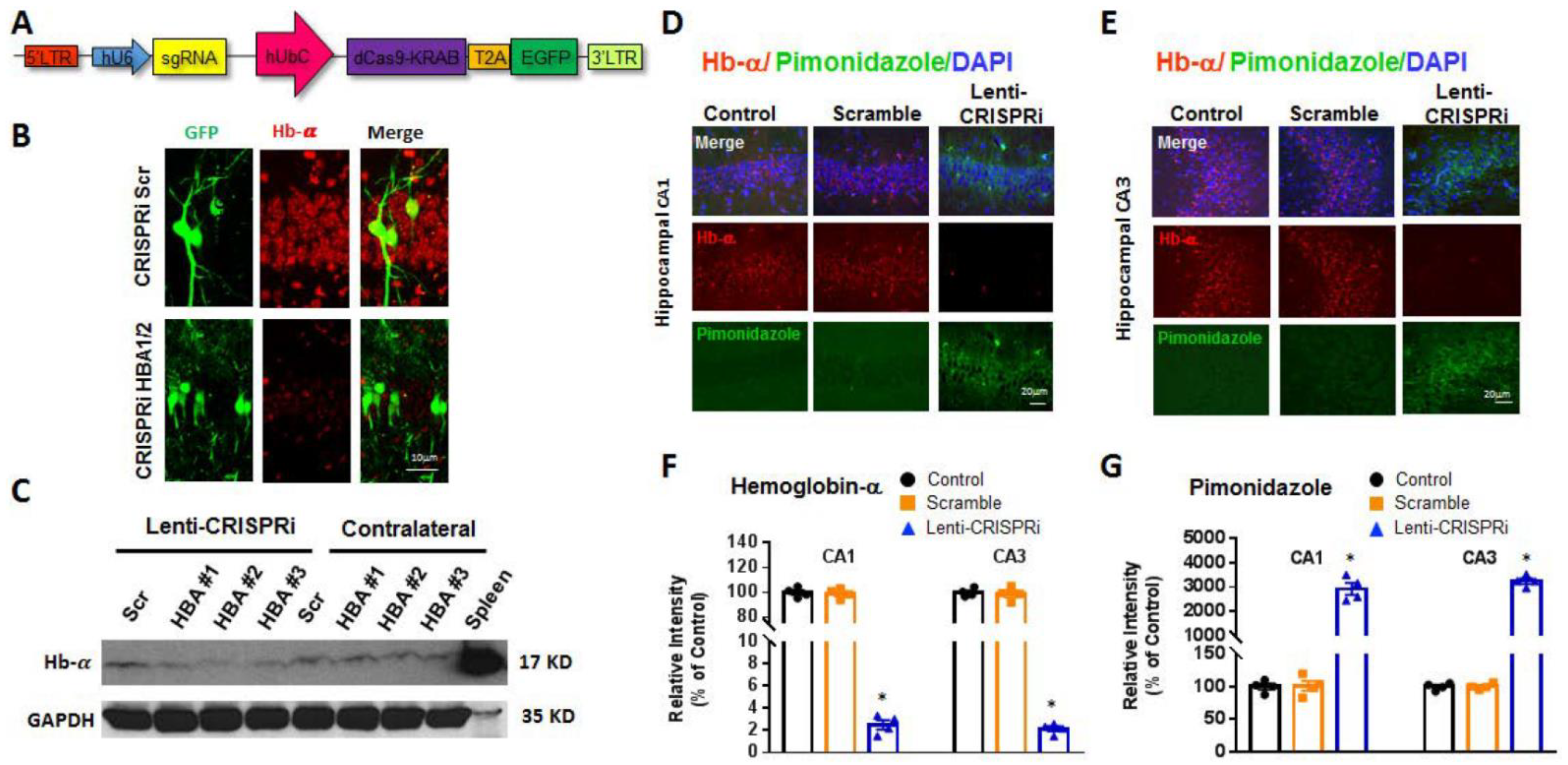
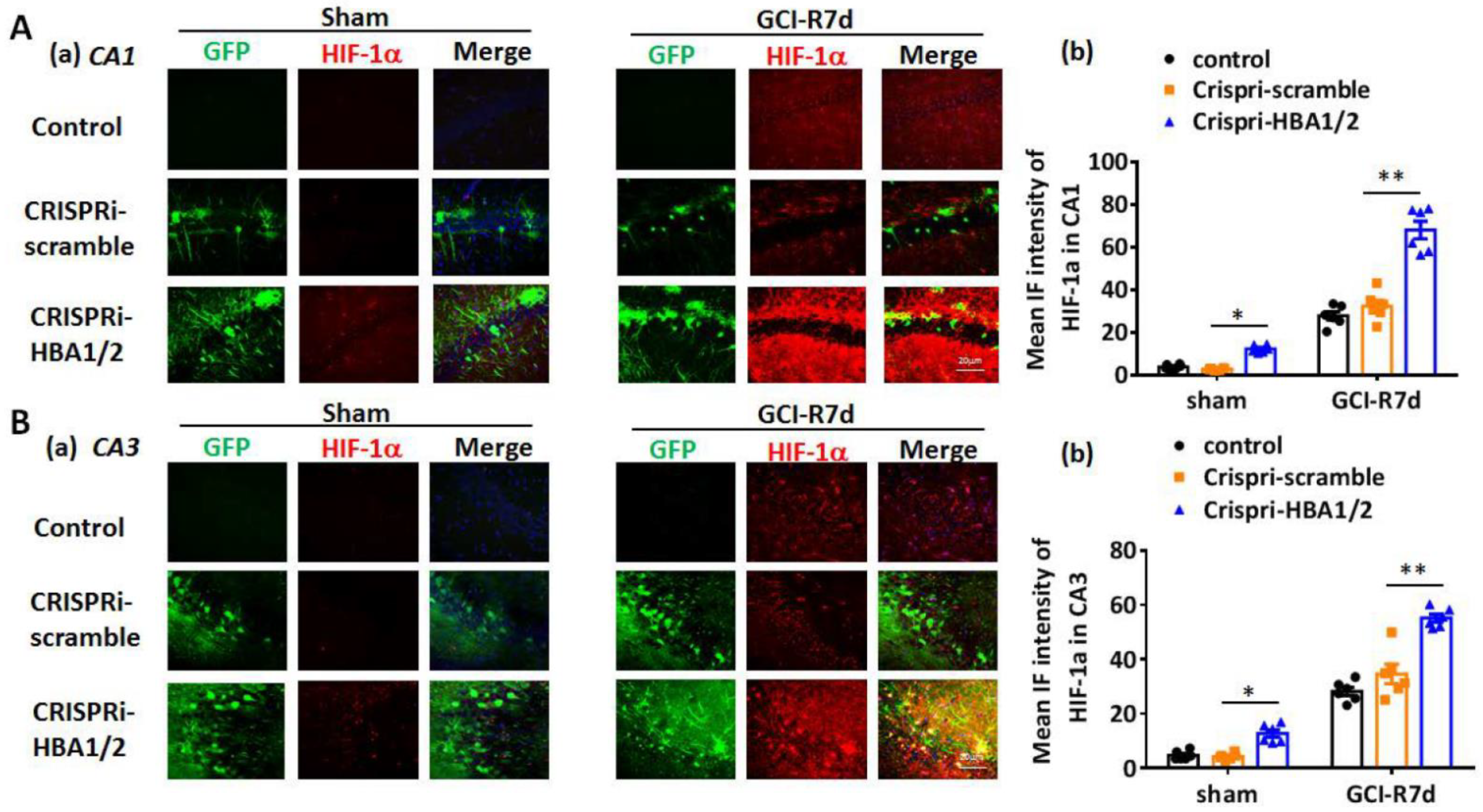
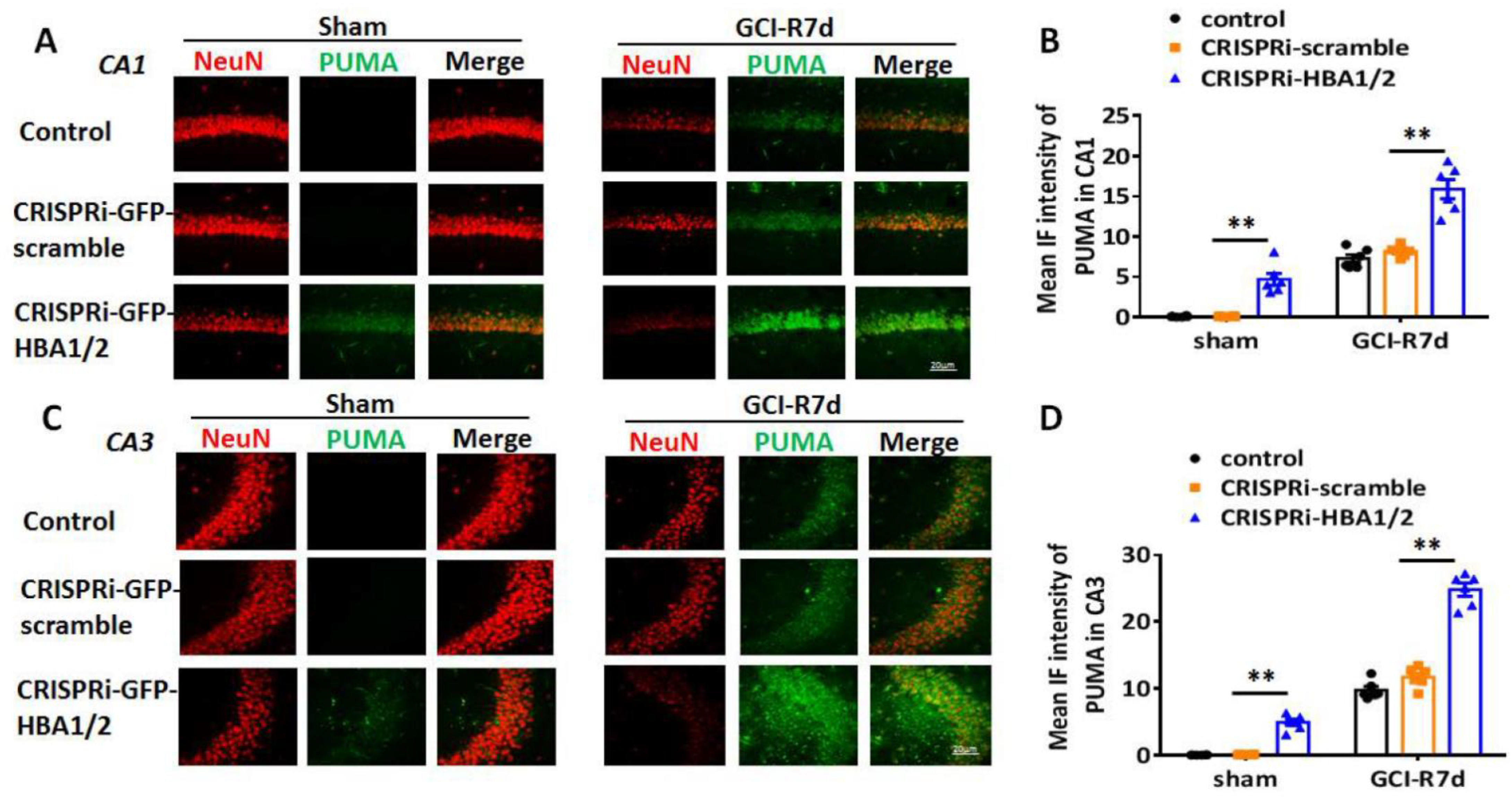
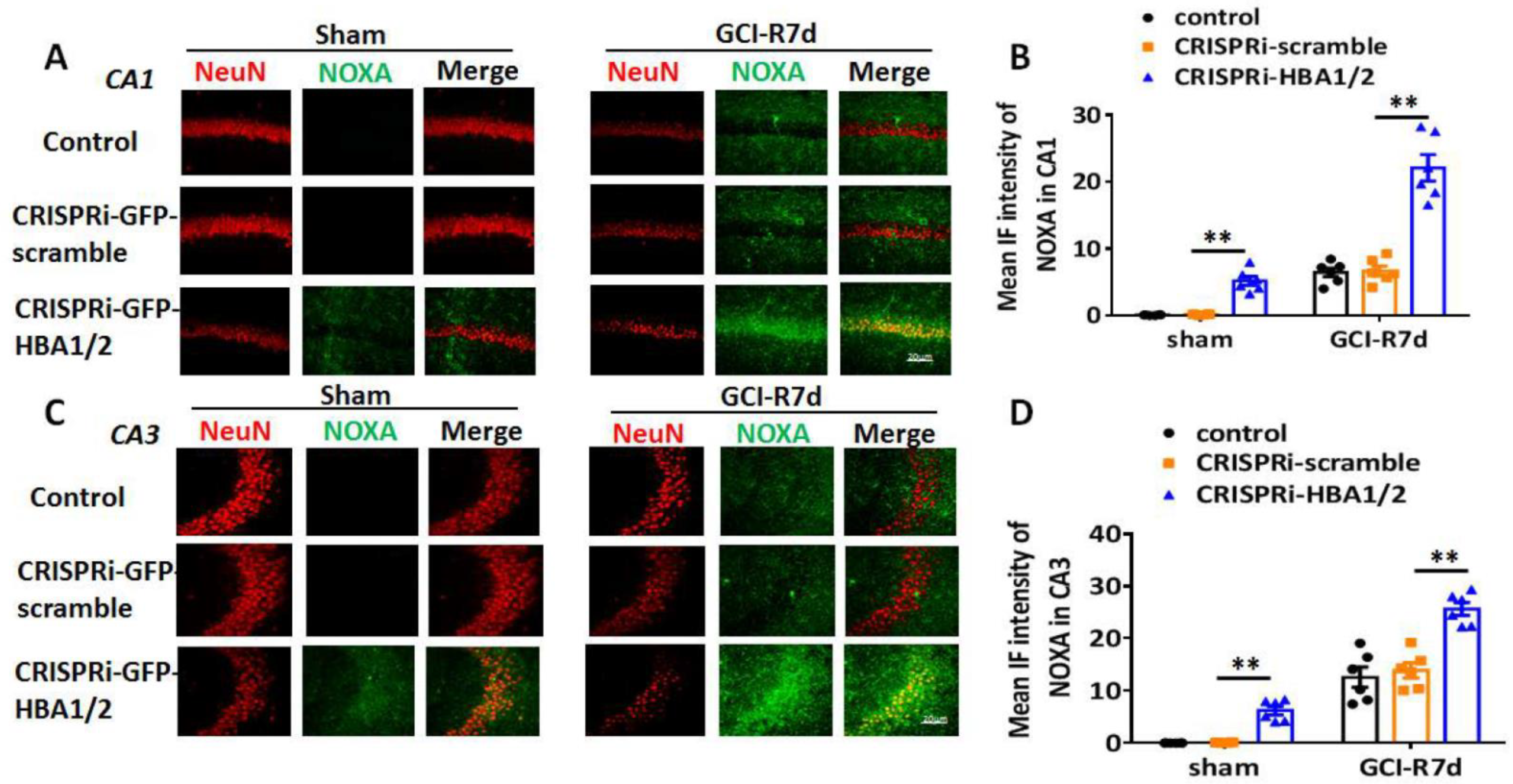
2. Results
3. Discussion
4. Materials and Methods
Animals
5. Plasmid Construction and sgRNA Design
6. Lentivirus Production
7. Cell Culture and Transfection
8. Lentivirus Infusion
9. Hypoxia
10. Global Cerebral Ischemia
11. Brain Section Collection and Immunohistochemistry (IHC) Analysis
12. Tissue Lysates Preparation and Western Blot Analysis
13. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gell, D.A. Structure and function of haemoglobins. Blood Cells Mol. Dis. 2018, 70, 13–42. [Google Scholar] [CrossRef] [PubMed]
- Wittenberg, J.B.; Wittenberg, B.A. Mechanisms of Cytoplasmic Hemoglobin and Myoglobin Function. Annu. Rev. Biophys. Biophys. Chem. 1990, 19, 217–241. [Google Scholar] [CrossRef] [PubMed]
- Gracco, V.L.; Lofqvist, A. Speech motor coordination and control: Evidence from lip, jaw, and laryngeal movements. J. Neurosci. 1994, 14 Pt 1, 6585–6597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, F.C.; Ackers, G.K. Quaternary enhancement in binding of oxygen by human hemoglobin. Proc. Natl. Acad. Sci. USA 1979, 76, 273–277. [Google Scholar] [CrossRef] [Green Version]
- Newton, D.A.; Rao, K.M.K.; Dluhy, R.A.; Baatz, J.E. Hemoglobin Is Expressed by Alveolar Epithelial Cells. J. Biol. Chem. 2006, 281, 5668–5676. [Google Scholar] [CrossRef] [Green Version]
- Dassen, H.; Kamps, R.; Punyadeera, C.; Dijcks, F.; De Goeij, A.; Ederveen, A.; Dunselman, G.; Groothuis, P. Haemoglobin expression in human endometrium. Hum. Reprod. 2008, 23, 635–641. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Zeng, M.; Stamler, J.S. Hemoglobin induction in mouse macrophages. Proc. Natl. Acad. Sci. USA 1999, 96, 6643–6647. [Google Scholar] [CrossRef] [Green Version]
- Tezel, T.H.; Geng, L.; Lato, E.B.; Schaal, S.; Liu, Y.; Dean, D.; Klein, J.B.; Kaplan, H.J.; Ellis, D.Z.; Dismuke, W.M.; et al. Synthesis and Secretion of Hemoglobin by Retinal Pigment Epithelium. Investig. Opthalmol. Vis. Sci. 2009, 50, 1911–1913. [Google Scholar] [CrossRef]
- Schelshorn, D.W.; Schneider, A.; Kuschinsky, W.; Weber, D.; Krüger, C.; Dittgen, T.; Bürgers, H.F.; Sabouri, F.; Gassler, N.; Bach, A.; et al. Expression of Hemoglobin in Rodent Neurons. J. Cereb. Blood Flow Metab. 2009, 29, 585–595. [Google Scholar] [CrossRef] [Green Version]
- Richter, F.; Meurers, B.H.; Zhu, C.; Medvedeva, V.P.; Chesselet, M.F. Neurons express hemoglobin alpha- and beta-chains in rat and human brains. J. Comp. Neurol. 2009, 515, 538–547. [Google Scholar] [CrossRef] [Green Version]
- Biagioli, M.; Pinto, M.; Cesselli, D.; Zaninello, M.; Lazarevic, D.; Roncaglia, P.; Simone, R.; Vlachouli, C.; Plessy, C.; Bertin, N.; et al. Unexpected expression of alpha- and beta-globin in mesencephalic dopaminergic neurons and glial cells. Proc. Natl. Acad. Sci. USA 2009, 106, 15454–15459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrer, I.; Gomez, A.; Carmona, M.; Huesa, G.; Porta, S.; Riera-Codina, M.; Biagioli, M.; Gustincich, S.; Aso, E. Neuronal hemoglobin is reduced in Alzheimer’s disease, argyrophilic grain disease, Parkinson’s disease, and dementia with Lewy bodies. J. Alzheimer’s Dis. 2011, 23, 537–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walser, M.; Svensson, J.; Karlsson, L.; Motalleb, R.; Åberg, M.; Kuhn, H.G.; Isgaard, J.; Åberg, N.D. Growth Hormone and Neuronal Hemoglobin in the Brain—Roles in Neuroprotection and Neurodegenerative Diseases. Front. Endocrinol. 2021, 11, 606089. [Google Scholar] [CrossRef] [PubMed]
- Kirino, T.; Tamura, A.; Sano, K. Selective Vulnerability of the Hippocampus to Ischemia—Reversible and Irreversible Types of Ischemic Cell Damage. Prog. Brain Res. 1985, 63, 39–58. [Google Scholar] [CrossRef] [PubMed]
- Aminova, L.R.; Siddiq, A.; Ratan, R.R. Antioxidants, HIF Prolyl Hydroxylase Inhibitors or Short Interfering RNAs to BNIP3 or PUMA, Can Prevent Prodeath Effects of the Transcriptional Activator, HIF-1α, in a Mouse Hippocampal Neuronal Line. Antioxid. Redox Signal. 2008, 10, 1989–1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-Y.; Ahn, H.-J.; Ryu, J.-H.; Suk, K.; Park, J.-H. BH3-only Protein Noxa Is a Mediator of Hypoxic Cell Death Induced by Hypoxia-inducible Factor 1α. J. Exp. Med. 2003, 199, 113–124. [Google Scholar] [CrossRef]
- Noto, T.; Furuichi, Y.; Ishiye, M.; Matsuoka, N.; Aramori, I.; Mutoh, S.; Yanagihara, T.; Manabe, N. Temporal and Topographic Profiles of Tissue Hypoxia Following Transient Focal Cerebral Ischemia in Rats. J. Veter.-Med. Sci. 2006, 68, 803–807. [Google Scholar] [CrossRef] [Green Version]
- Aguilera, K.; Brekken, R. Hypoxia Studies with Pimonidazole in vivo. Bio. Protoc. 2014, 4, e1254. [Google Scholar] [CrossRef] [Green Version]
- Raleigh, J.A.; Koch, C.J. Importance of thiols in the reductive binding of 2-nitroimidazoles to macromolecules. Biochem. Pharmacol. 1990, 40, 2457–2464. [Google Scholar] [CrossRef]
- Savell, K.E.; Day, J.J. Applications of CRISPR/Cas9 in the Mammalian Central Nervous System. Yale J. Biol. Med. 2017, 90, 567–581. [Google Scholar]
- Zheng, Y.; Shen, W.; Zhang, J.; Yang, B.; Liu, Y.N.; Qi, H.; Yu, X.; Lu, S.Y.; Chen, Y.; Xu, Y.Z.; et al. CRISPR interference-based specific and efficient gene inactivation in the brain. Nat. Neurosci. 2018, 21, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L. Expression of hypoxia-inducible factor 1: Mechanisms and consequences. Biochem. Pharmacol. 2000, 59, 47–53. [Google Scholar] [CrossRef]
- Wang, G.L.; Jiang, B.-H.; Rue, E.A.; Semenza, G.L. Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS heterodimer regulated by cellular O2 tension. Proc. Natl. Acad. Sci. USA 1995, 92, 5510–5514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiryu-Seo, S.; Hirayama, T.; Kato, R.; Kiyama, H. Noxa Is a Critical Mediator of p53-Dependent Motor Neuron Death after Nerve Injury in Adult Mouse. J. Neurosci. 2005, 25, 1442–1447. [Google Scholar] [CrossRef] [PubMed]
- Niizuma, K.; Endo, H.; Nito, C.; Myer, D.J.; Chan, P.H. Potential Role of PUMA in Delayed Death of Hippocampal CA1 Neurons After Transient Global Cerebral Ischemia. Stroke 2009, 40, 618–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, W.G. The sex difference in haemoglobin levels in adults—Mechanisms, causes, and consequences. Blood Rev. 2014, 28, 41–47. [Google Scholar] [CrossRef]
- Marti, H.H.; Wenger, R.H.; Rivas, L.A.; Straumann, U.; Digicaylioglu, M.; Henn, V.; Yonekawa, Y.; Bauer, C.; Gassmann, M. Erythropoietin gene expression in human, monkey and murine brain. Eur. J. Neurosci. 1996, 8, 666–676. [Google Scholar] [CrossRef]
- Wang, F.; Fried, W. Renal and extrarenal erythropoietin production in male and female rats of various ages. J. Lab. Clin. Med. 1972, 79, 181–186. [Google Scholar]
- Snyder, B.; Wu, H.-K.; Tillman, B.; Floyd, T.F. Aged Mouse Hippocampus Exhibits Signs of Chronic Hypoxia and an Impaired HIF-Controlled Response to Acute Hypoxic Exposures. Cells 2022, 11, 423. [Google Scholar] [CrossRef]
- Hirschler, L.; Munting, L.P.; Khmelinskii, A.; Teeuwisse, W.M.; Suidgeest, E.; Warnking, J.M.; Van Der Weerd, L.; Barbier, E.L.; Van Osch, M.J. Transit time mapping in the mouse brain using time-encoded pCASL. NMR Biomed. 2018, 31, e3855. [Google Scholar] [CrossRef]
- Maier, F.C.; Wehrl, H.F.; Schmid, A.M.; Mannheim, J.G.; Wiehr, S.; Lerdkrai, C.; Calaminus, C.; Stahlschmidt, A.; Ye, L.; Burnet, M.; et al. Longitudinal PET-MRI reveals beta-amyloid deposition and rCBF dynamics and connects vascular amyloidosis to quantitative loss of perfusion. Nat. Med. 2014, 20, 1485–1492. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Stoica, B.A.; Sabirzhanov, B.; Burns, M.P.; Faden, A.I.; Loane, D.J. Traumatic brain injury in aged animals increases lesion size and chronically alters microglial/macrophage classical and alternative activation states. Neurobiol. Aging 2013, 34, 1397–1411. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; Sun, X.; Puchowicz, M.A.; LaManna, J.C. Increased Sensitivity to Transient Global Ischemia in Aging Rat Brain. Adv. Exp. Med. Biol. 2007, 599, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.-G.; Han, D.; Wang, R.-M.; Dong, Y.; Yang, F.; Vadlamudi, R.K.; Brann, D.W. C terminus of Hsc70-interacting protein (CHIP)-mediated degradation of hippocampal estrogen receptor-α and the critical period hypothesis of estrogen neuroprotection. Proc. Natl. Acad. Sci. USA 2011, 108, E617–E624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukandala, G.; Tynan, R.; Lanigan, S.; O’Connor, J.J. The Effects of Hypoxia and Inflammation on Synaptic Signaling in the CNS. Brain Sci. 2016, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Bai, N.; Zhang, Q.; Zhang, W.; Liu, B.; Yang, F.; Brann, D.; Wang, R. G-protein-coupled estrogen receptor activation upregulates interleukin-1 receptor antagonist in the hippocampus after global cerebral ischemia: Implications for neuronal self-defense. J. Neuroinflamm. 2020, 17, 45. [Google Scholar] [CrossRef]
- Chan, P.H. Mitochondria and Neuronal Death/Survival Signaling Pathways in Cerebral Ischemia. Neurochem. Res. 2004, 29, 1943–1949. [Google Scholar] [CrossRef]
- Chavez, J.C.; Lamanna, J.C. Activation of Hypoxia-Inducible Factor-1 in the Rat Cerebral Cortex after Transient Global Ischemia: Potential Role of Insulin-Like Growth Factor-1. J. Neurosci. 2002, 22, 8922–8931. [Google Scholar] [CrossRef] [Green Version]
- Raz, L.; Zhang, Q.-G.; Han, D.; Dong, Y.; De Sevilla, L.; Brann, D.W. Acetylation of the Pro-Apoptotic Factor, p53 in the Hippocampus following Cerebral Ischemia and Modulation by Estrogen. PLoS ONE 2011, 6, e27039. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, T.; Fujimura, M.; Noshita, N.; Kim, G.W.; Saito, A.; Hayashi, T.; Narasimhan, P.; Maier, C.M.; Chan, P.H. Neuronal death/survival signaling pathways in cerebral ischemia. NeuroRx 2004, 1, 17–25. [Google Scholar] [CrossRef]
- Helton, R.; Cui, J.; Scheel, J.; Ellison, J.A.; Ames, C.; Gibson, C.; Blouw, B.; Ouyang, L.; Dragatsis, I.; Zeitlin, S.; et al. Brain-Specific Knock-Out of Hypoxia-Inducible Factor-1 Reduces Rather Than Increases Hypoxic-Ischemic Damage. J. Neurosci. 2005, 25, 4099–4107. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, M.; Shirasawa, T. Presenilin-1-deficient neurons are nitric oxide-dependently killed by hydrogen peroxide in vitro. Neuroscience 2004, 125, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.T.; Reeder, B. Oxygen-binding haem proteins. Exp. Physiol. 2008, 93, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Zhang Lab. Available online: https://zlab.bio/guide-design-resources (accessed on 3 July 2018).
- Wang, J.; Zhang, Q.; Lu, Y.; Dong, Y.; Dhandapani, K.M.; Brann, D.W.; Yu, R.K. Ganglioside GD3 is up-regulated in microglia and regulates phagocytosis following global cerebral ischemia. J. Neurochem. 2021, 158, 737–752. [Google Scholar] [CrossRef]
- Wang, J.; Sareddy, G.R.; Lu, Y.; Pratap, U.P.; Tang, F.; Greene, K.M.; Meyre, P.L.; Tekmal, R.R.; Vadlamudi, R.K.; Brann, D.W. Astrocyte-Derived Estrogen Regulates Reactive Astrogliosis and is Neuroprotective following Ischemic Brain Injury. J. Neurosci. 2020, 40, 9751–9771. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Y.; Wang, J.; Tang, F.; Pratap, U.P.; Sareddy, G.R.; Dhandapani, K.M.; Capuano, A.; Arvanitakis, Z.; Vadlamudi, R.K.; Brann, D.W. Regulation and Role of Neuron-Derived Hemoglobin in the Mouse Hippocampus. Int. J. Mol. Sci. 2022, 23, 5360. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105360
Lu Y, Wang J, Tang F, Pratap UP, Sareddy GR, Dhandapani KM, Capuano A, Arvanitakis Z, Vadlamudi RK, Brann DW. Regulation and Role of Neuron-Derived Hemoglobin in the Mouse Hippocampus. International Journal of Molecular Sciences. 2022; 23(10):5360. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105360
Chicago/Turabian StyleLu, Yujiao, Jing Wang, Fulei Tang, Uday P. Pratap, Gangadhara R. Sareddy, Krishnan M. Dhandapani, Ana Capuano, Zoe Arvanitakis, Ratna K. Vadlamudi, and Darrell W. Brann. 2022. "Regulation and Role of Neuron-Derived Hemoglobin in the Mouse Hippocampus" International Journal of Molecular Sciences 23, no. 10: 5360. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105360