
Correction of Beta-Thalassemia IVS-II-654 Mutation in a Mouse Model Using Prime Editing


Abstract
:1. Introduction
2. Results
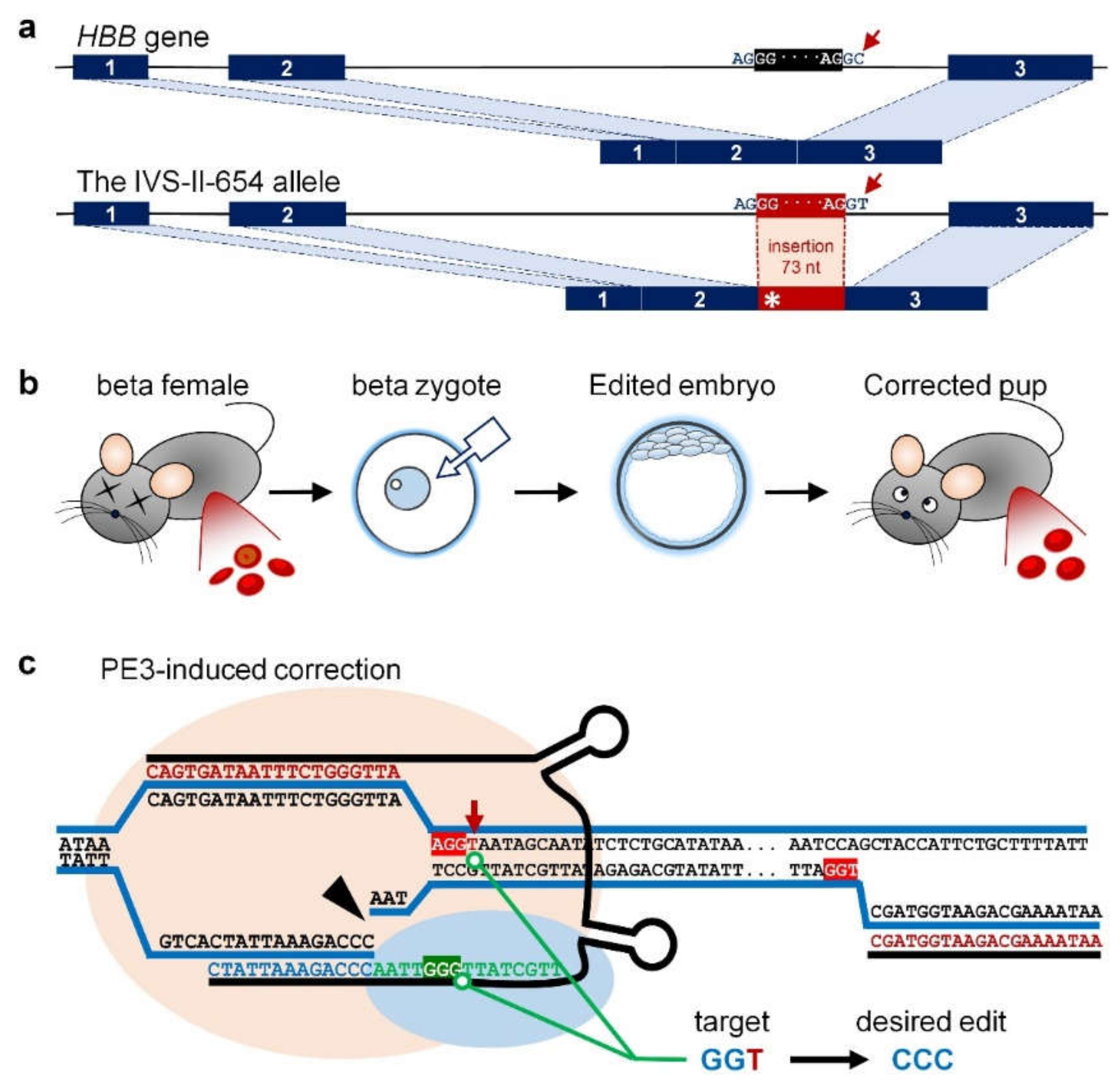
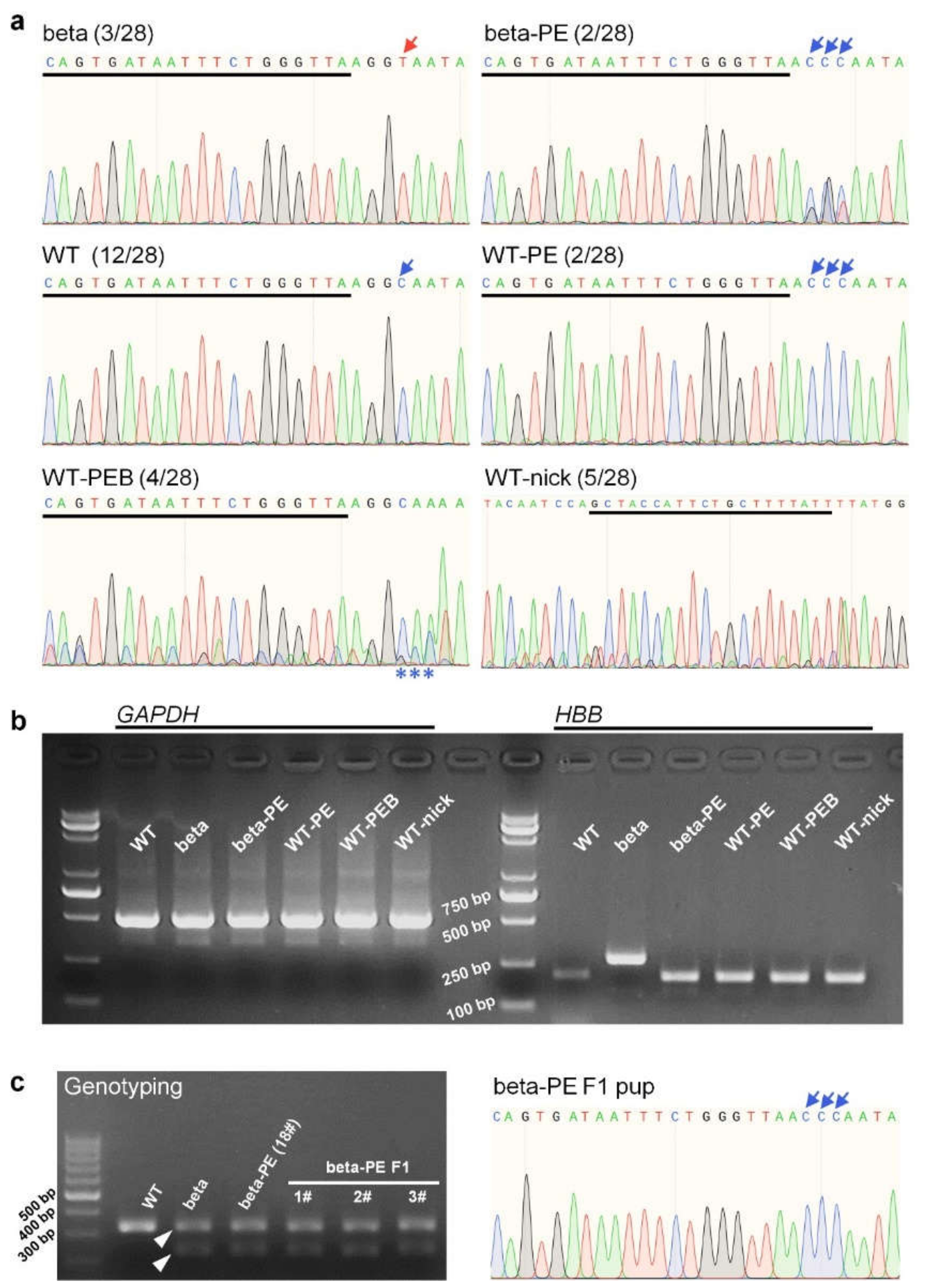
2.1. Genotyping Analysis Showed That PE3 Installed Desired On-Target Edits at the IVS-II-654 Site
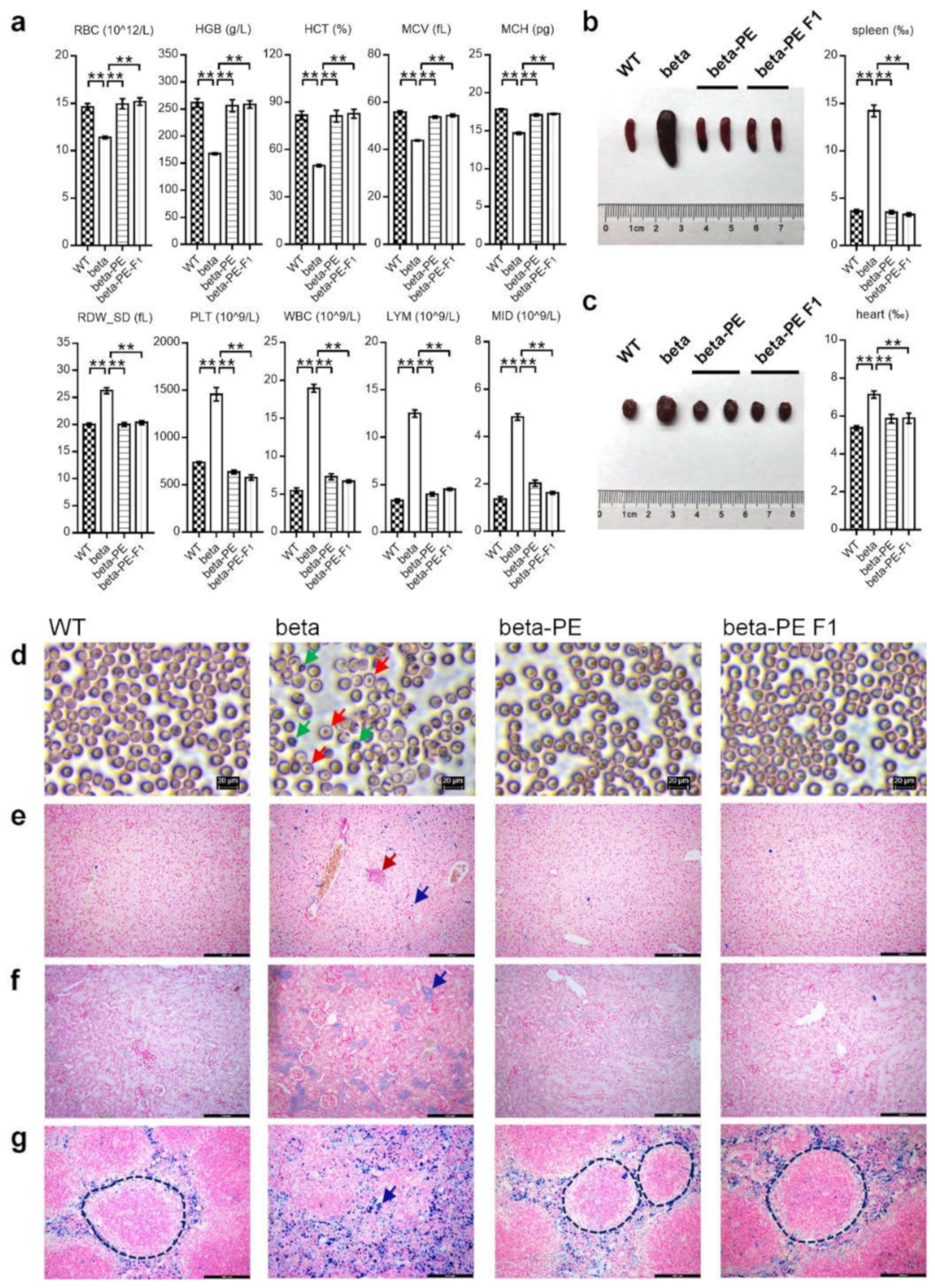
2.2. Phenotypic Analysis Showed That PE3-Induced Treatment Effectively Cured Thalassemia Symptoms
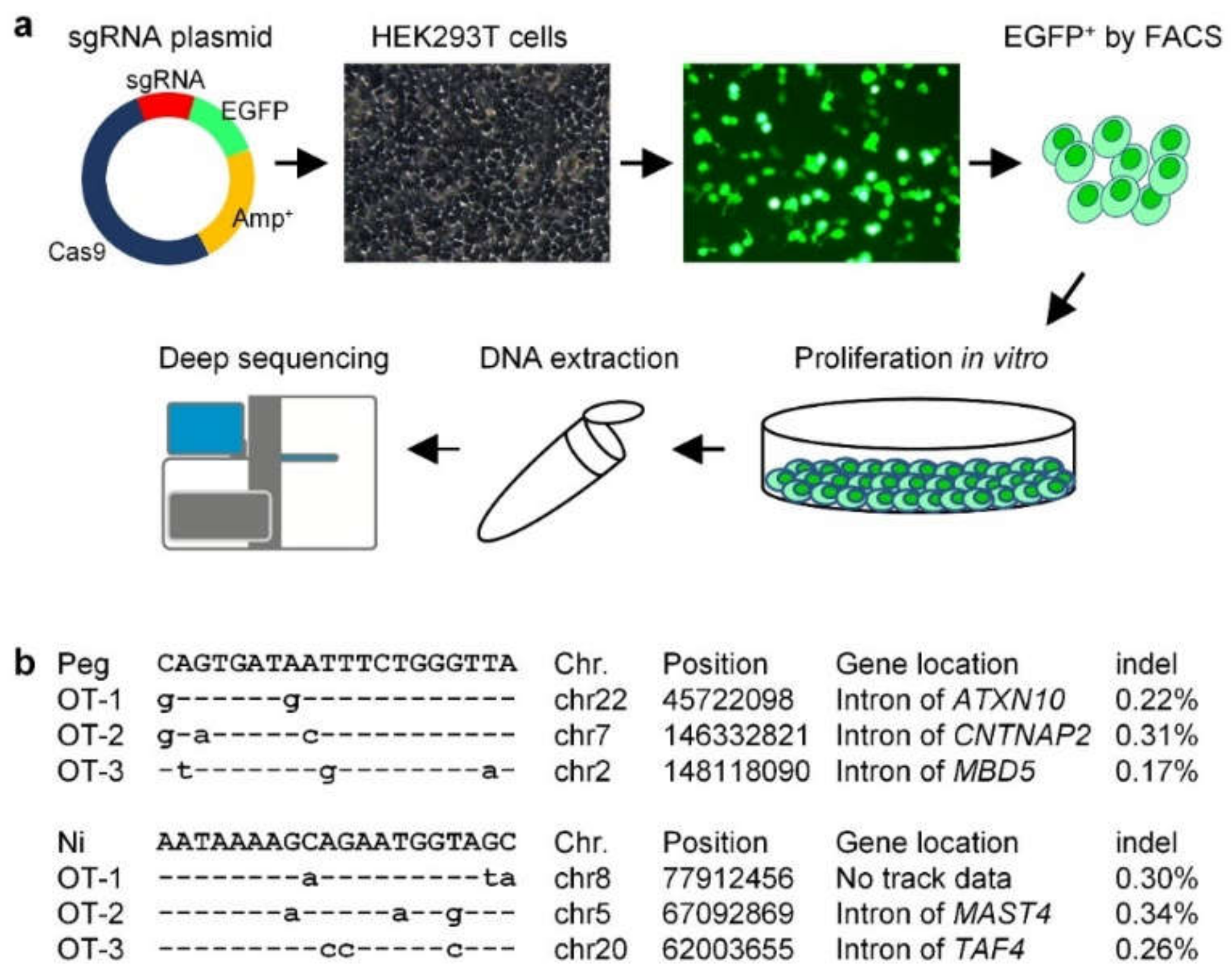
2.3. Off-Target Analysis Showed That PE3-Induced IVS-II-654 Correction Was Safe in the Human Genome
3. Discussion
4. Materials and Methods
4.1. Animal Model and Study Approval
4.2. Plasmid Construction
4.3. In Vitro Transcription
4.4. Embryo Microinjection and Identification of Gene Editing Mice
4.5. RT-PCR Analysis
4.6. Hematological Analysis
4.7. Histopathology
4.8. Statistical Analysis
4.9. Cell Culture and Transfection
4.10. Targeted Deep Sequencing of Off-Targets
4.11. Data Analysis of Deep Sequencing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thein, S.L. Molecular basis of beta thalassemia and potential therapeutic targets. Blood Cells Mol. Dis. 2018, 70, 54–65. [Google Scholar] [PubMed]
- Modell, B.; Darlison, M. Global epidemiology of haemoglobin disorders and derived service indicators. Bull. World Health Organ. 2008, 86, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Hardison, R.C.; Chui, D.H.; Riemer, C.R.; Miller, W.; Carver, M.F.; Molchanova, T.P.; Efremov, G.D.; Huisman, T.H. Access to a syllabus of human hemoglobin variants (1996) via the World Wide Web. Hemoglobin 1998, 22, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.Z.; Zeng, F.Y.; Ren, Z.R.; Lu, Z.H.; Rodgers, G.P.; Schechter, A.N.; Zeng, Y.T. RNA transcripts of the beta-thalassaemia allele IVS-2-654 C-->T: A small amount of normally processed beta-globin mRNA is still produced from the mutant gene. Br. J. Haematol. 1994, 88, 541–546. [Google Scholar] [CrossRef]
- Lewis, J.; Yang, B.; Kim, R.; Sierakowska, H.; Kole, R.; Smithies, O.; Maeda, N. A common human beta globin splicing mutation modeled in mice. Blood 1998, 91, 2152–2156. [Google Scholar] [CrossRef]
- Ali, S.; Mumtaz, S.; Shakir, H.A.; Khan, M.; Tahir, H.M.; Mumtaz, S.; Mughal, T.A.; Hassan, A.; Kazmi, S.A.R.; Sadia; et al. Current status of beta-thalassemia and its treatment strategies. Mol. Genet. Genom. Med. 2021, 9, e1788. [Google Scholar] [CrossRef]
- Zhang, J.; Yan, J.; Zeng, F. Recent Progress on Genetic Diagnosis and Therapy for beta-Thalassemia in China and Around the World. Hum. Gene Ther. 2018, 29, 197–203. [Google Scholar] [CrossRef]
- Xie, S.Y.; Ren, Z.R.; Zhang, J.Z.; Guo, X.B.; Wang, Q.X.; Wang, S.; Lin, D.; Gong, X.L.; Li, W.; Huang, S.Z.; et al. Restoration of the balanced alpha/beta-globin gene expression in beta654-thalassemia mice using combined RNAi and antisense RNA approach. Hum. Mol. Genet. 2007, 16, 2616–2625. [Google Scholar] [CrossRef] [Green Version]
- Xie, S.; Li, W.; Ren, Z.; Zhang, J.; Guo, X.; Wang, S.; Huang, S.; Zeng, F.; Zeng, Y.T. Amelioration of beta654-thalassemia in mouse model with the knockdown of aberrantly spliced beta-globin mRNA. J. Genet. Genom. 2008, 35, 595–601. [Google Scholar] [CrossRef]
- Xie, S.Y.; Li, W.; Ren, Z.R.; Huang, S.Z.; Zeng, F.; Zeng, Y.T. Correction of beta654-thalassaemia mice using direct intravenous injection of siRNA and antisense RNA vectors. Int. J. Hematol. 2011, 93, 301–310. [Google Scholar] [CrossRef]
- Li, W.; Xie, S.; Guo, X.; Gong, X.; Wang, S.; Lin, D.; Zhang, J.; Ren, Z.; Huang, S.; Zeng, F.; et al. A novel transgenic mouse model produced from lentiviral germline integration for the study of beta-thalassemia gene therapy. Haematologica 2008, 93, 356–362. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Shi, W.; Hu, X.; Zhang, J.; Gong, Z.; Guo, X.; Ren, Z.; Zeng, F. Therapeutic effects of induced pluripotent stem cells in chimeric mice with beta-thalassemia. Haematologica 2014, 99, 1304–1311. [Google Scholar] [CrossRef]
- Fang, Y.; Cheng, Y.; Lu, D.; Gong, X.; Yang, G.; Gong, Z.; Zhu, Y.; Sang, X.; Fan, S.; Zhang, J.; et al. Treatment of beta(654)-thalassaemia by TALENs in a mouse model. Cell Prolif. 2018, 51, e12491. [Google Scholar] [CrossRef] [Green Version]
- Lu, D.; Gong, X.; Fang, Y.; Guo, X.; Chen, Y.; Yang, F.; Zhao, G.; Ma, Q.; Zeng, Y.; Zeng, F.; et al. Correction of RNA splicing defect in beta(654)-thalassemia mice using CRISPR/Cas9 gene-editing technology. Haematologica 2021. [Google Scholar] [CrossRef]
- Anzalone, A.V.; Randolph, P.B.; Davis, J.R.; Sousa, A.A.; Koblan, L.W.; Levy, J.M.; Chen, P.J.; Wilson, C.; Newby, G.A.; Raguram, A.; et al. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef]
- Antoniou, P.; Miccio, A.; Brusson, M. Base and Prime Editing Technologies for Blood Disorders. Front. Genome Ed. 2021, 3, 618406. [Google Scholar]
- Petri, K.; Zhang, W.; Ma, J.; Schmidts, A.; Lee, H.; Horng, J.E.; Kim, D.Y.; Kurt, I.C.; Clement, K.; Hsu, J.Y.; et al. CRISPR prime editing with ribonucleoprotein complexes in zebrafish and primary human cells. Nat. Biotechnol. 2022, 40, 189–193. [Google Scholar] [CrossRef]
- Anzalone, A.V.; Koblan, L.W.; Liu, D.R. Genome editing with CRISPR-Cas nucleases, base editors, transposases and prime editors. Nat. Biotechnol. 2020, 38, 824–844. [Google Scholar]
- Yang, L.; Tang, J.; Ma, X.; Lin, Y.; Ma, G.; Shan, M.; Wang, L.; Yang, Y. Progression and application of CRISPR-Cas genomic editors. Methods 2021, 194, 65–74. [Google Scholar] [CrossRef]
- Koniali, L.; Lederer, C.W.; Kleanthous, M. Therapy Development by Genome Editing of Hematopoietic Stem Cells. Cells 2021, 10, 1492. [Google Scholar] [CrossRef]
- Yao, X.; Wang, X.; Hu, X.; Liu, Z.; Liu, J.; Zhou, H.; Shen, X.; Wei, Y.; Huang, Z.; Ying, W.; et al. Homology-mediated end joining-based targeted integration using CRISPR/Cas9. Cell Res. 2017, 27, 801–814. [Google Scholar] [CrossRef] [Green Version]
- Thein, S.L. Genetic association studies in beta-hemoglobinopathies. Hematol. Am. Soc. Hematol. Educ. Program. 2013, 2013, 354–361. [Google Scholar] [CrossRef] [Green Version]
- Bak, R.O.; Dever, D.P.; Porteus, M.H. CRISPR/Cas9 genome editing in human hematopoietic stem cells. Nat. Protoc. 2018, 13, 358–376. [Google Scholar] [CrossRef] [Green Version]
- Malik, P.; Tisdale, J. Gene and Cell Therapies for Beta-Globinopathies. In American Society of Gene Cell Therapy, 1st ed.; Springer: New York, NY, USA, 2017; p. 1. [Google Scholar]
- Komor, A.C.; Kim, Y.B.; Packer, M.S.; Zuris, J.A.; Liu, D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–424. [Google Scholar] [CrossRef] [Green Version]
- Matsoukas, I.G. Commentary: Programmable base editing of A.T to G.C in genomic DNA without DNA cleavage. Front. Genet. 2018, 9, 21. [Google Scholar] [CrossRef] [Green Version]
- Sankaran, V.G.; Menne, T.F.; Xu, J.; Akie, T.E.; Lettre, G.; Van Handel, B.; Mikkola, H.K.; Hirschhorn, J.N.; Cantor, A.B.; Orkin, S.H.; et al. Human fetal hemoglobin expression is regulated by the developmental stage-specific repressor BCL11A. Science 2008, 322, 1839–1842. [Google Scholar] [CrossRef] [Green Version]
- Sankaran, V.G.; Xu, J.; Ragoczy, T.; Ippolito, G.C.; Walkley, C.R.; Maika, S.D.; Fujiwara, Y.; Ito, M.; Groudine, M.; Bender, M.A.; et al. Developmental and species-divergent globin switching are driven by BCL11A. Nature 2009, 460, 1093–1097. [Google Scholar] [CrossRef] [Green Version]
- Jawaid, K.; Wahlberg, K.; Thein, S.L.; Best, S. Binding patterns of BCL11A in the globin and GATA1 loci and characterization of the BCL11A fetal hemoglobin locus. Blood Cells Mol. Dis. 2010, 45, 140–146. [Google Scholar] [CrossRef]
- Zhou, D.; Liu, K.; Sun, C.W.; Pawlik, K.M.; Townes, T.M. KLF1 regulates BCL11A expression and gamma- to beta-globin gene switching. Nat. Genet. 2010, 42, 742–744. [Google Scholar] [CrossRef]
- Xu, J.; Bauer, D.E.; Kerenyi, M.A.; Vo, T.D.; Hou, S.; Hsu, Y.J.; Yao, H.; Trowbridge, J.J.; Mandel, G.; Orkin, S.H.; et al. Corepressor-dependent silencing of fetal hemoglobin expression by BCL11A. Proc. Natl. Acad. Sci. USA 2013, 110, 6518–6523. [Google Scholar] [CrossRef] [Green Version]
- Canver, M.C.; Smith, E.C.; Sher, F.; Pinello, L.; Sanjana, N.E.; Shalem, O.; Chen, D.D.; Schupp, P.G.; Vinjamur, D.S.; Garcia, S.P.; et al. BCL11A enhancer dissection by Cas9-mediated in situ saturating mutagenesis. Nature 2015, 527, 192–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Zeng, J.; Roscoe, B.P.; Liu, P.; Yao, Q.; Lazzarotto, C.R.; Clement, K.; Cole, M.A.; Luk, K.; Baricordi, C.; et al. Highly efficient therapeutic gene editing of human hematopoietic stem cells. Nat. Med. 2019, 25, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Wu, Y.; Ren, C.; Bonanno, J.; Shen, A.H.; Shea, D.; Gehrke, J.M.; Clement, K.; Luk, K.; Yao, Q.; et al. Therapeutic base editing of human hematopoietic stem cells. Nat. Med. 2020, 26, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Demirci, S.; Zeng, J.; Wu, Y.; Uchida, N.; Shen, A.H.; Pellin, D.; Gamer, J.; Yapundich, M.; Drysdale, C.; Bonanno, J.; et al. BCL11A enhancer-edited hematopoietic stem cells persist in rhesus monkeys without toxicity. J. Clin. Invest. 2020, 130, 6677–6687. [Google Scholar] [CrossRef]
- Butt, H.; Rao, G.S.; Sedeek, K.; Aman, R.; Kamel, R.; Mahfouz, M. Engineering herbicide resistance via prime editing in rice. Plant. Biotechnol. J. 2020, 18, 2370–2372. [Google Scholar] [CrossRef]
- Lin, Q.; Zong, Y.; Xue, C.; Wang, S.; Jin, S.; Zhu, Z.; Wang, Y.; Anzalone, A.V.; Raguram, A.; Doman, J.L.; et al. Prime genome editing in rice and wheat. Nat. Biotechnol. 2020, 38, 582–585. [Google Scholar] [CrossRef]
- Tang, X.; Sretenovic, S.; Ren, Q.; Jia, X.; Li, M.; Fan, T.; Yin, D.; Xiang, S.; Guo, Y.; Liu, L.; et al. Plant Prime Editors Enable Precise Gene Editing in Rice Cells. Mol. Plant. 2020, 13, 667–670. [Google Scholar] [CrossRef]
- Kweon, J.; Yoon, J.K.; Jang, A.H.; Shin, H.R.; See, J.E.; Jang, G.; Kim, J.I.; Kim, Y. Engineered prime editors with PAM flexibility. Mol. Ther. 2021, 29, 2001–2007. [Google Scholar] [CrossRef]
- Liu, P.; Liang, S.Q.; Zheng, C.; Mintzer, E.; Zhao, Y.G.; Ponnienselvan, K.; Mir, A.; Sontheimer, E.J.; Gao, G.; Flotte, T.R.; et al. Improved prime editors enable pathogenic allele correction and cancer modelling in adult mice. Nat. Commun. 2021, 12, 2121. [Google Scholar] [CrossRef]
- Concordet, J.P.; Haeussler, M. CRISPOR: Intuitive guide selection for CRISPR/Cas9 genome editing experiments and screens. Nucleic Acids Res. 2018, 46, W242–W245. [Google Scholar] [CrossRef]
- Liang, P.; Ding, C.; Sun, H.; Xie, X.; Xu, Y.; Zhang, X.; Sun, Y.; Xiong, Y.; Ma, W.; Liu, Y.; et al. Correction of beta-thalassemia mutant by base editor in human embryos. Protein Cell 2017, 8, 811–822. [Google Scholar] [CrossRef]
- Antony, J.S.; Latifi, N.; Haque, A.; Lamsfus-Calle, A.; Daniel-Moreno, A.; Graeter, S.; Baskaran, P.; Weinmann, P.; Mezger, M.; Handgretinger, R.; et al. Gene correction of HBB mutations in CD34(+) hematopoietic stem cells using Cas9 mRNA and ssODN donors. Mol. Cell Pediatr. 2018, 5, 9. [Google Scholar] [CrossRef]
- Dahlman, J.E.; Kauffman, K.J.; Xing, Y.; Shaw, T.E.; Mir, F.F.; Dlott, C.C.; Langer, R.; Anderson, D.G.; Wang, E.T. Barcoded nanoparticles for high throughput in vivo discovery of targeted therapeutics. Proc. Natl. Acad. Sci. USA 2017, 114, 2060–2065. [Google Scholar] [CrossRef] [Green Version]
- Paunovska, K.; Sago, C.D.; Monaco, C.M.; Hudson, W.H.; Castro, M.G.; Rudoltz, T.G.; Kalathoor, S.; Vanover, D.A.; Santangelo, P.J.; Ahmed, R.; et al. A Direct Comparison of in Vitro and in Vivo Nucleic Acid Delivery Mediated by Hundreds of Nanoparticles Reveals a Weak Correlation. Nano Lett. 2018, 18, 2148–2157. [Google Scholar] [CrossRef]
- Paunovska, K.; Gil, C.J.; Lokugamage, M.P.; Sago, C.D.; Sato, M.; Lando, G.N.; Gamboa Castro, M.; Bryksin, A.V.; Dahlman, J.E. Analyzing 2000 in Vivo Drug Delivery Data Points Reveals Cholesterol Structure Impacts Nanoparticle Delivery. ACS Nano 2018, 12, 8341–8349. [Google Scholar] [CrossRef]
- Sago, C.D.; Lokugamage, M.P.; Islam, F.Z.; Krupczak, B.R.; Sato, M.; Dahlman, J.E. Nanoparticles That Deliver RNA to Bone Marrow Identified by in Vivo Directed Evolution. J. Am. Chem. Soc. 2018, 140, 17095–17105. [Google Scholar] [CrossRef]
- Sago, C.D.; Kalathoor, S.; Fitzgerald, J.P.; Lando, G.N.; Djeddar, N.; Bryksin, A.V.; Dahlman, J.E. Barcoding chemical modifications into nucleic acids improves drug stability in vivo. J. Mater. Chem. B 2018, 6, 7197–7203. [Google Scholar] [CrossRef] [Green Version]
- Sago, C.D.; Lokugamage, M.P.; Paunovska, K.; Vanover, D.A.; Monaco, C.M.; Shah, N.N.; Gamboa Castro, M.; Anderson, S.E.; Rudoltz, T.G.; Lando, G.N.; et al. High-throughput in vivo screen of functional mRNA delivery identifies nanoparticles for endothelial cell gene editing. Proc. Natl. Acad. Sci. USA 2018, 115, E9944–E9952. [Google Scholar] [CrossRef] [Green Version]
- Paunovska, K.; Loughrey, D.; Sago, C.D.; Langer, R.; Dahlman, J.E. Using Large Datasets to Understand Nanotechnology. Adv. Mater. 2019, 31, e1902798. [Google Scholar] [CrossRef]
- Chow, R.D.; Chen, J.S.; Shen, J.; Chen, S. A web tool for the design of prime-editing guide RNAs. Nat. Biomed. Eng. 2021, 5, 190–194. [Google Scholar] [CrossRef]
- Bae, S.; Park, J.; Kim, J.S. Cas-OFFinder: A fast and versatile algorithm that searches for potential off-target sites of Cas9 RNA-guided endonucleases. Bioinformatics 2014, 30, 1473–1475. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Pup | Identifier | Number | Annotation |
|---|---|---|---|---|
| Desired editing | beta-PE | 17#, 18# | 2 | PE3 installed editing of GGT > CCC in beta-genotype pups |
| WT-PE | 21#, 22# | 2 | PE3 installed editing of GGC > CCC in WT-genotype pups | |
| By-product editing | WT-PEB | 9#, 28#, 30#, 32# | 4 | PE3 induced the CCC conversion near the IVS-II-654 site |
| WT-nick | 7#, 15#, 34#, 35#, 46# | 5 | PE3 generated by-product edits at the nick sgRNA site | |
| Without editing | beta | 20#, 24#, 31# | 3 | |
| WT | 1#, 8#, 12#, 16#, 26#, 27#, 29#, 33#, 37#, 38#, 42#, 44# | 12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Sun, R.; Fei, J.; Chen, H.; Lu, D. Correction of Beta-Thalassemia IVS-II-654 Mutation in a Mouse Model Using Prime Editing. Int. J. Mol. Sci. 2022, 23, 5948. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23115948
Zhang H, Sun R, Fei J, Chen H, Lu D. Correction of Beta-Thalassemia IVS-II-654 Mutation in a Mouse Model Using Prime Editing. International Journal of Molecular Sciences. 2022; 23(11):5948. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23115948
Chicago/Turabian StyleZhang, Haokun, Ruilin Sun, Jian Fei, Hongyan Chen, and Daru Lu. 2022. "Correction of Beta-Thalassemia IVS-II-654 Mutation in a Mouse Model Using Prime Editing" International Journal of Molecular Sciences 23, no. 11: 5948. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23115948