
Betaine Alleviates High-Fat Diet-Induced Disruptionof Hepatic Lipid and Iron Homeostasis in Mice


Abstract
:1. Introduction
2. Results
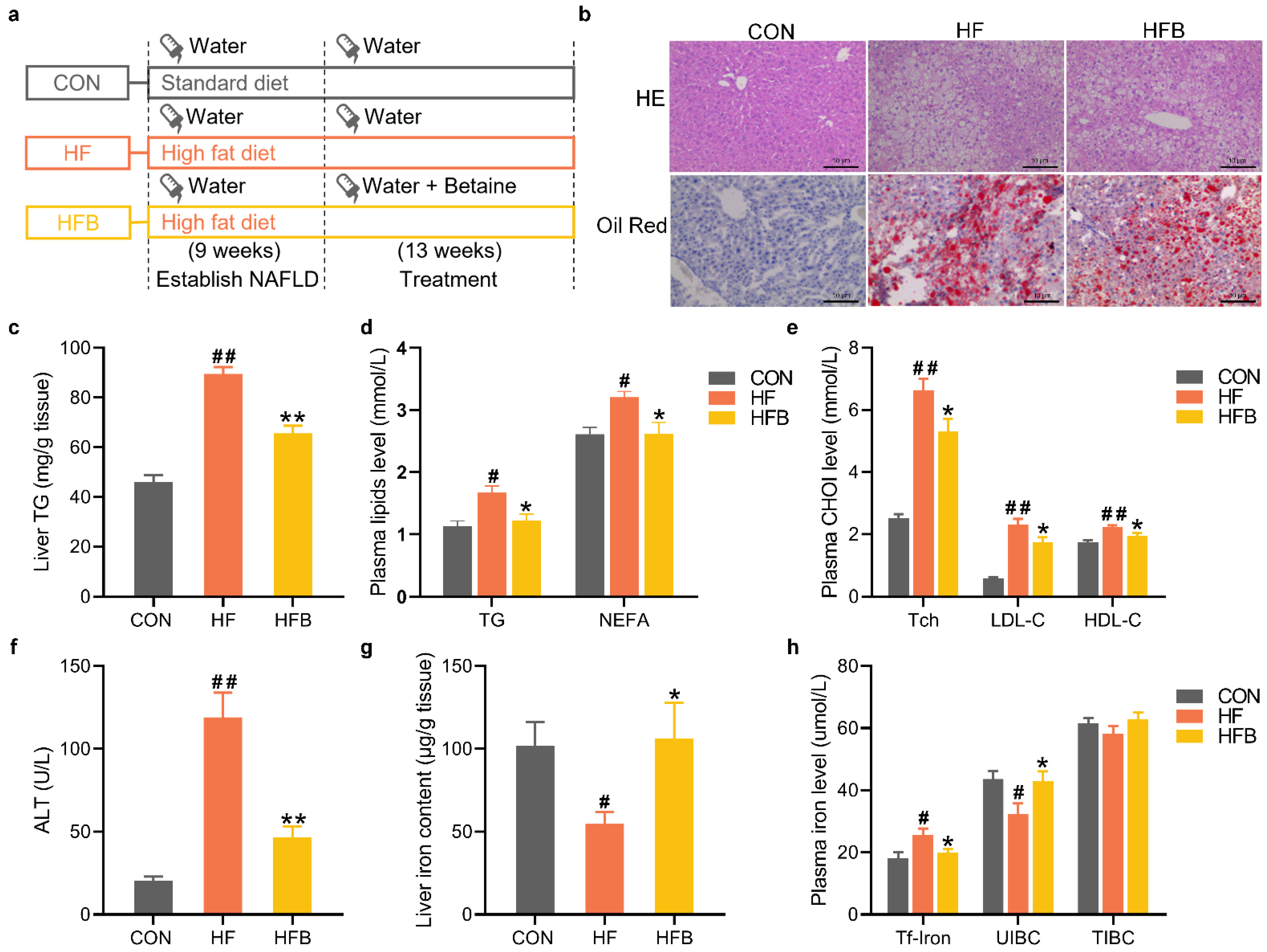
2.1. Betaine Alleviates HFD-Induced Disruption of Lipid and Iron Homeostasis
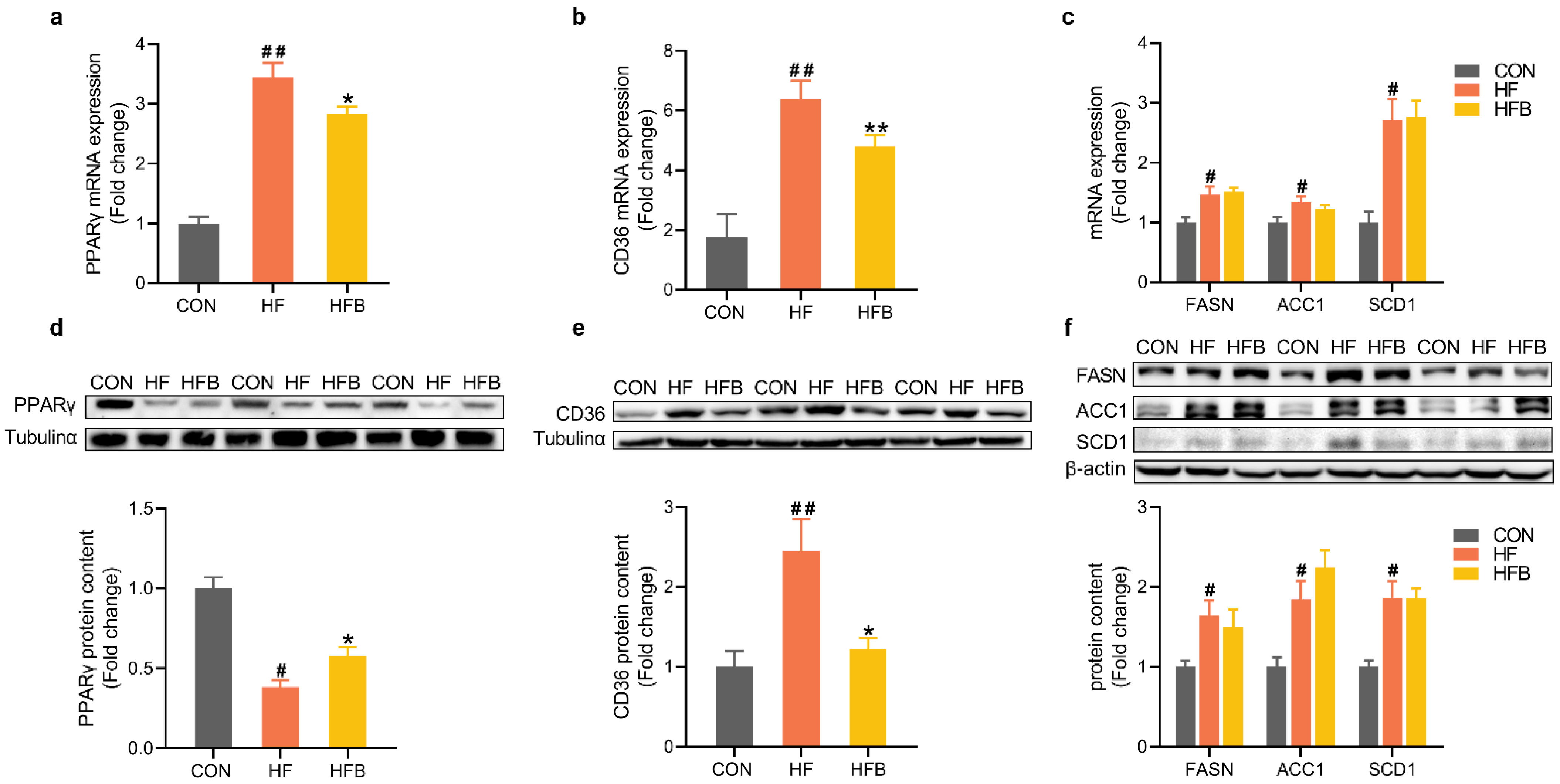
2.2. Betaine Reverses HFD-Induced Dysregulation of Pparγ and CD36 in the Liver
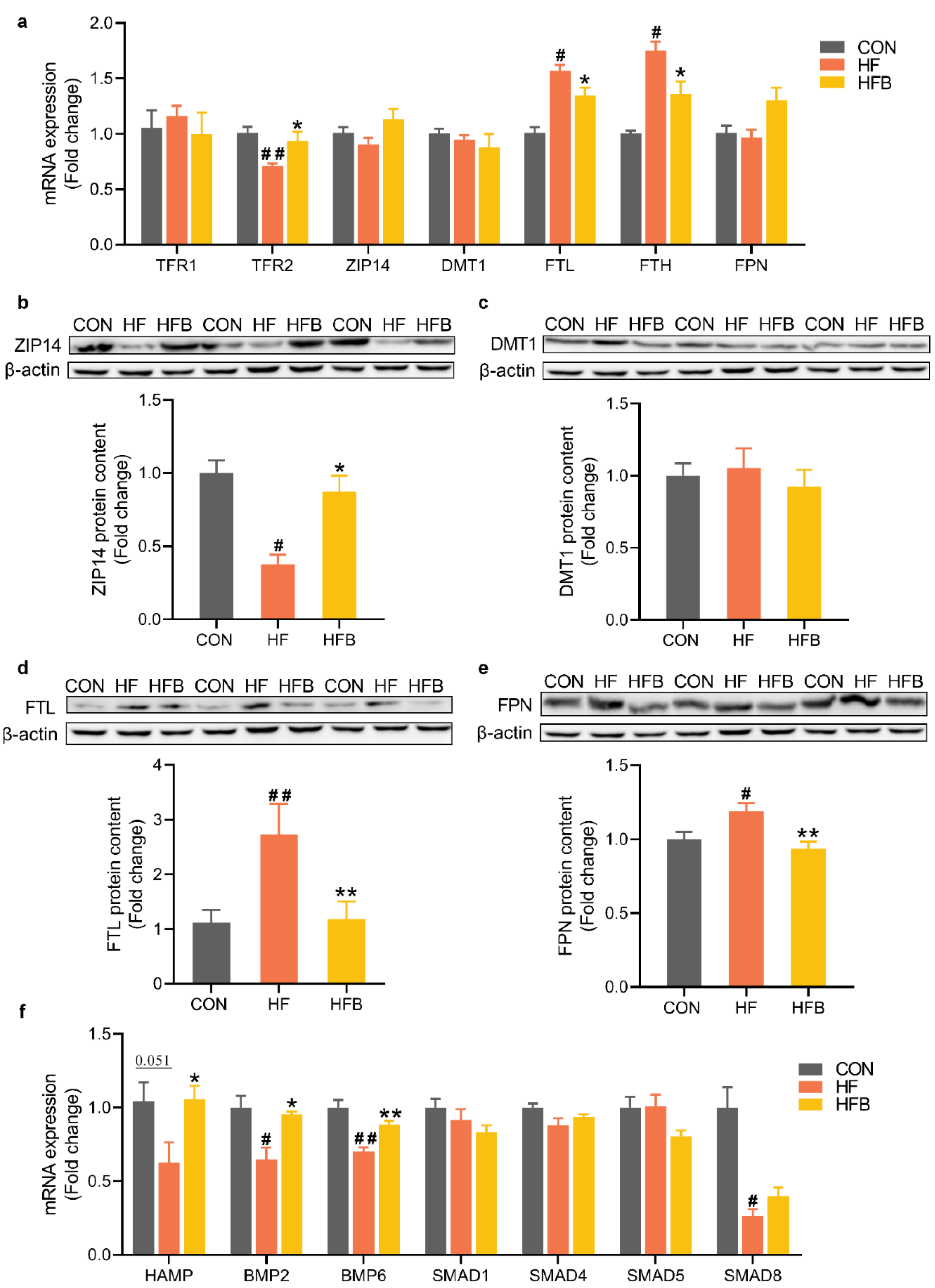
2.3. Betaine Mitigates HFD-Induced Disruption of Iron-Metabolic Gene Expression
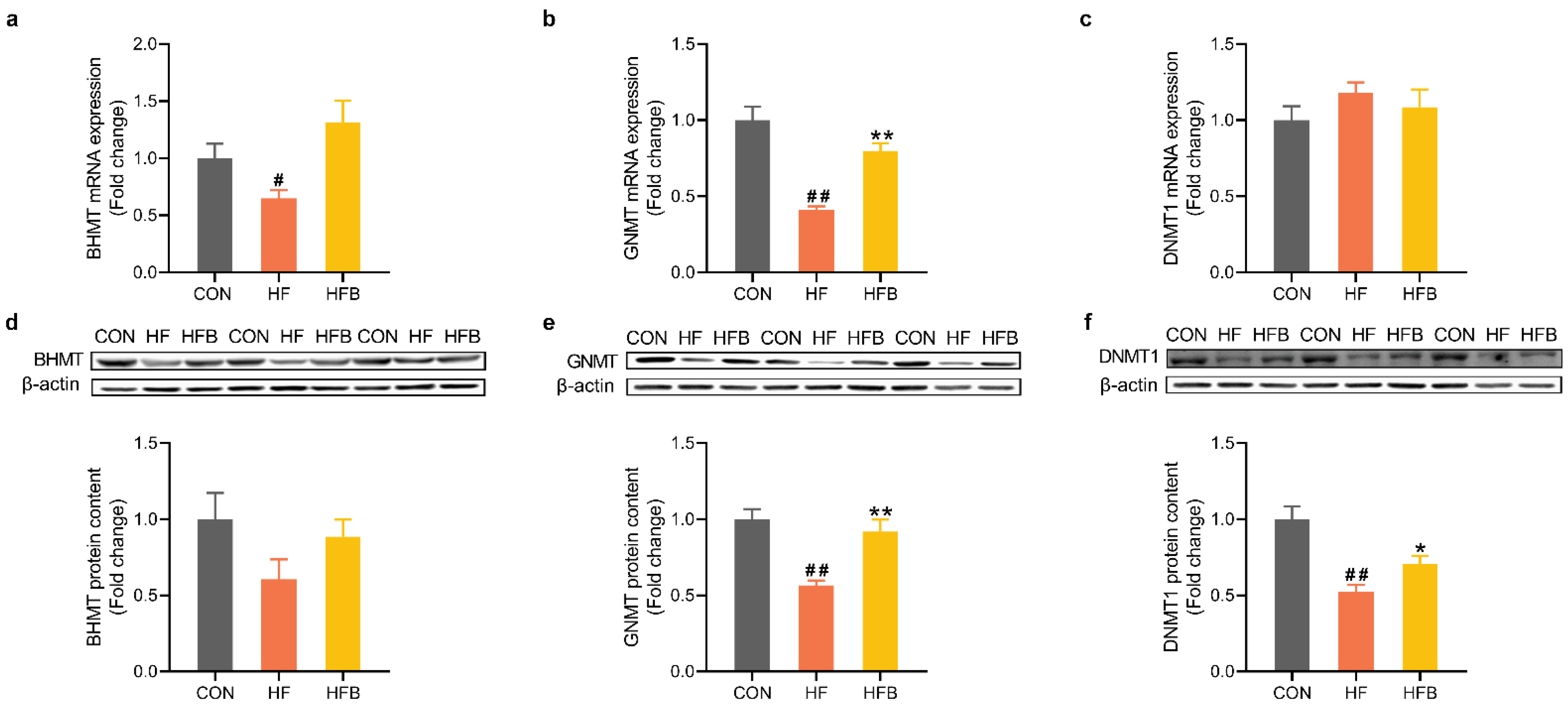
2.4. Betaine Restores HFD-Induced Dysregulation of Methyl-Transfer Genes
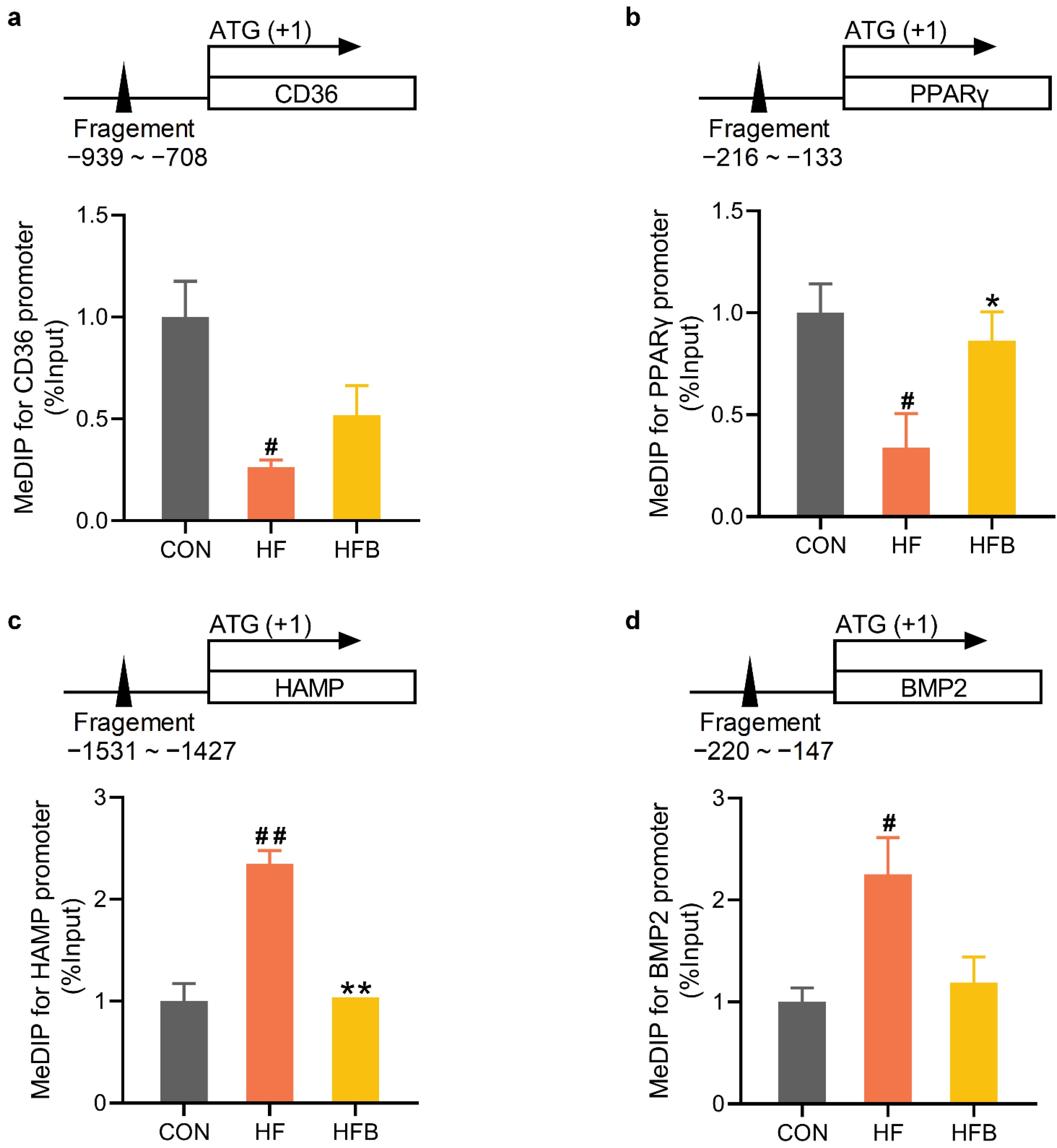
2.5. Betaine Rectifies HFD-Modified DNA Methylation on Promoter of Affected Genes
3. Discussion
4. Materials and Methods
4.1. Animals and Treatment
4.2. Determination of Plasma Biochemical Parameters
4.3. Hepatic Histological Evaluation
4.4. Determination of Hepatic Triglyceride Content
4.5. Total RNA Isolation and Real-Time PCR
4.6. Western Blot Analysis
4.7. Methylated DNA Immunoprecipitation (MeDIP) Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Mundi, M.S.; Velapati, S.; Patel, J.; Kellogg, T.A.; Abu Dayyeh, B.K.; Hurt, R.T. Evolution of NAFLD and Its Management. Nutr. Clin. Pract. 2020, 35, 72–84. [Google Scholar] [CrossRef]
- Ipsen, D.H.; Lykkesfeldt, J.; Tveden-Nyborg, P. Molecular mechanisms of hepatic lipid accumulation in non-alcoholic fatty liver disease. Cell Mol. Life Sci. 2018, 75, 3313–3327. [Google Scholar] [CrossRef] [Green Version]
- Datz, C.; Felder, T.K.; Niederseer, D.; Aigner, E. Iron homeostasis in the metabolic syndrome. Eur. J. Clin. Investig. 2013, 43, 215–224. [Google Scholar] [CrossRef]
- Ahmed, U.; Oates, P.S. Dietary fat level affects tissue iron levels but not the iron regulatory gene HAMP in rats. Nutr. Res. 2013, 33, 126–135. [Google Scholar] [CrossRef]
- Meli, R.; Raso, G.M.; Irace, C.; Simeoli, R.; Di Pascale, A.; Paciello, O.; Pagano, T.B.; Calignano, A.; Colonna, A.; Santamaria, R. High Fat Diet Induces Liver Steatosis and Early Dysregulation of Iron Metabolism in Rats. PLoS ONE 2013, 8, e66570. [Google Scholar]
- Bertinato, J.; Aroche, C.; Plouffe, L.J.; Lee, M.; Murtaza, Z.; Kenney, L.; Lavergne, C.; Aziz, A. Diet-induced obese rats have higher iron requirements and are more vulnerable to iron deficiency. Eur. J. Nutr. 2014, 53, 885–895. [Google Scholar] [CrossRef]
- Sonnweber, T.; Ress, C.; Nairz, M.; Theurl, I.; Schroll, A.; Murphy, A.T.; Wroblewski, V.; Witcher, D.R.; Moser, P.; Ebenbichler, C.F.; et al. High-fat diet causes iron deficiency via hepcidin-independent reduction of duodenal iron absorption. J. Nutr. Biochem. 2012, 23, 1600–1608. [Google Scholar] [CrossRef]
- Chung, H.; Wu, D.; Smith, D.; Meydani, S.N.; Han, S.N. Lower hepatic iron storage associated with obesity in mice can be restored by decreasing body fat mass through feeding a low-fat diet. Nutr. Res. 2016, 36, 955–963. [Google Scholar] [CrossRef] [Green Version]
- Chung, J.; Kim, M.S.; Han, S.N. Diet-induced obesity leads to decreased hepatic iron storage in mice. Nutr. Res. 2011, 31, 915–921. [Google Scholar] [CrossRef]
- Bloomer, S.A.; Brown, K.E. Iron-Induced Liver Injury: A Critical Reappraisal. Int. J. Mol. Sci. 2019, 20, 915–921. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, U.; Latham, P.S.; Oates, P.S. Interactions between hepatic iron and lipid metabolism with possible relevance to steatohepatitis. World J. Gastroenterol. 2012, 18, 4651–4658. [Google Scholar] [CrossRef]
- Koonen, D.P.; Jacobs, R.L.; Febbraio, M.; Young, M.E.; Soltys, C.L.; Ong, H.; Vance, D.E.; Dyck, J.R. Increased hepatic CD36 expression contributes to dyslipidemia associated with diet-induced obesity. Diabetes 2007, 56, 2863–2871. [Google Scholar] [CrossRef] [Green Version]
- Wilson, C.G.; Tran, J.L.; Erion, D.M.; Vera, N.B.; Febbraio, M.; Weiss, E.J. Hepatocyte-Specific Disruption of CD36 Attenuates Fatty Liver and Improves Insulin Sensitivity in HFD-Fed Mice. Endocrinology 2016, 157, 570–585. [Google Scholar] [CrossRef] [Green Version]
- Zeng, H.; Qin, H.; Liao, M.; Zheng, E.; Luo, X.; Xiao, A.; Li, Y.; Chen, L.; Wei, L.; Zhao, L.; et al. CD36 promotes de novo lipogenesis in hepatocytes through INSIG2-dependent SREBP1 processing. Mol. Metab. 2021, 57, 101428. [Google Scholar] [CrossRef]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science 2004, 306, 2090–2093. [Google Scholar] [CrossRef] [Green Version]
- Barisani, D.; Pelucchi, S.; Mariani, R.; Galimberti, S.; Trombini, P.; Fumagalli, D.; Meneveri, R.; Nemeth, E.; Ganz, T.; Piperno, A. Hepcidin and iron-related gene expression in subjects with Dysmetabolic Hepatic Iron Overload. J. Hepatol. 2008, 49, 123–133. [Google Scholar] [CrossRef]
- Aigner, E.; Theurl, I.; Theurl, M.; Lederer, D.; Haufe, H.; Dietze, O.; Strasser, M.; Datz, C.; Weiss, G. Pathways underlying iron accumulation in human nonalcoholic fatty liver disease. Am. J. Clin. Nutr. 2008, 87, 1374–1383. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Wang, X.; Song, G.; Xing, H.; Yang, L.; Han, K.; Chang, Y.Z. A high-fructose diet in rats induces systemic iron deficiency and hepatic iron overload by an inflammation mechanism. J. Food Biochem. 2021, 45, e13578. [Google Scholar] [CrossRef]
- Pogribny, I.P.; Tryndyak, V.P.; Bagnyukova, T.V.; Melnyk, S.; Montgomery, B.; Ross, S.A.; Latendresse, J.R.; Rusyn, I.; Beland, F.A. Hepatic epigenetic phenotype predetermines individual susceptibility to hepatic steatosis in mice fed a lipogenic methyl-deficient diet. J. Hepatol. 2009, 51, 176–186. [Google Scholar] [CrossRef] [Green Version]
- Pirola, C.J.; Gianotti, T.F.; Burgueno, A.L.; Rey-Funes, M.; Loidl, C.F.; Mallardi, P.; Martino, J.S.; Castano, G.O.; Sookoian, S. Epigenetic modification of liver mitochondrial DNA is associated with histological severity of nonalcoholic fatty liver disease. Gut 2013, 62, 1356–1363. [Google Scholar] [CrossRef] [PubMed]
- Hajri, T.; Zaiou, M.; Fungwe, T.V.; Ouguerram, K.; Besong, S. Epigenetic Regulation of Peroxisome Proliferator-Activated Receptor Gamma Mediates High-Fat Diet-Induced Non-Alcoholic Fatty Liver Disease. Cells 2021, 10, 1355. [Google Scholar] [CrossRef] [PubMed]
- Ge, Z.J.; Luo, S.M.; Lin, F.; Liang, Q.X.; Huang, L.; Wei, Y.C.; Hou, Y.; Han, Z.M.; Schatten, H.; Sun, Q.Y. DNA methylation in oocytes and liver of female mice and their offspring: Effects of high-fat-diet-induced obesity. Environ. Health Perspect. 2014, 122, 159–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, G.; He, F.; Wu, C.; Li, P.; Li, N.; Deng, J.; Zhu, G.; Ren, W.; Peng, Y. Betaine in Inflammation: Mechanistic Aspects and Applications. Front. Immunol. 2018, 9, 1070. [Google Scholar] [CrossRef] [Green Version]
- Zhao, N.; Yang, S.; Jia, Y.; Sun, B.; He, B.; Zhao, R. Maternal betaine supplementation attenuates glucocorticoid-induced hepatic lipid accumulation through epigenetic modification in adult offspring rats. J. Nutr. Biochem. 2018, 54, 105–112. [Google Scholar] [CrossRef]
- Zhao, N.; Yang, S.; Sun, B.; Feng, Y.; Zhao, R. Maternal betaine protects rat offspring from glucocorticoid-induced activation of lipolytic genes in adipose tissue through modification of DNA methylation. Eur. J. Nutr. 2020, 59, 1707–1716. [Google Scholar] [CrossRef]
- Hu, Y.; Sun, Q.; Hu, Y.; Hou, Z.; Zong, Y.; Omer, N.A.; Abobaker, H.; Zhao, R. Corticosterone-Induced Lipogenesis Activation and Lipophagy Inhibition in Chicken Liver Are Alleviated by Maternal Betaine Supplementation. J. Nutr. 2018, 148, 316–325. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.M.; Liu, Y.; Zhou, R.F.; Chen, X.L.; Wang, C.; Tan, X.Y.; Wang, L.J.; Zheng, R.D.; Zhang, H.W.; Ling, W.H.; et al. Associations of gut-flora-dependent metabolite trimethylamine-N-oxide, betaine and choline with non-alcoholic fatty liver disease in adults. Sci. Rep. 2016, 6, 19076. [Google Scholar] [CrossRef]
- Huang, T.; Yu, L.; Pan, H.; Ma, Z.; Wu, T.; Zhang, L.; Liu, K.; Qi, Q.; Miao, W.; Song, Z.; et al. Integrated Transcriptomic and Translatomic Inquiry of the Role of Betaine on Lipid Metabolic Dysregulation Induced by a High-Fat Diet. Front. Nutr. 2021, 8, 751436. [Google Scholar] [CrossRef]
- Fan, C.; Hu, H.; Huang, X.; Su, D.; Huang, F.; Zhuo, Z.; Tan, L.; Xu, Y.; Wang, Q.; Hou, K.; et al. Betaine Supplementation Causes an Increase in Fatty Acid Oxidation and Carbohydrate Metabolism in Livers of Mice Fed a High-Fat Diet: A Proteomic Analysis. Foods 2022, 11, 881. [Google Scholar] [CrossRef]
- Szkudelska, K.; Chan, M.H.; Okulicz, M.; Jasaszwili, M.; Lukomska, A.; Malek, E.; Shah, M.; Sunder, S.; Szkudelski, T. Betaine supplementation to rats alleviates disturbances induced by high-fat diet: Pleiotropic effects in model of type 2 diabetes. J. Physiol. Pharmacol. 2021, 72, 763–775. [Google Scholar] [CrossRef]
- Jin, M.; Shen, Y.; Pan, T.; Zhu, T.; Li, X.; Xu, F.; Betancor, M.B.; Jiao, L.; Tocher, D.R.; Zhou, Q. Dietary Betaine Mitigates Hepatic Steatosis and Inflammation Induced by a High-Fat-Diet by Modulating the Sirt1/Srebp-1/Ppara Pathway in Juvenile Black Seabream (Acanthopagrus schlegelii). Front. Immunol. 2021, 12, 694720. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, L.W.; Wang, L.K.; Li, X.; Zhang, H.; Luo, L.P.; Song, J.C.; Gong, Z.J. Betaine protects against high-fat-diet-induced liver injury by inhibition of high-mobility group box 1 and Toll-like receptor 4 expression in rats. Dig. Dis. Sci. 2013, 58, 3198–3206. [Google Scholar] [CrossRef] [PubMed]
- Kathirvel, E.; Morgan, K.; Nandgiri, G.; Sandoval, B.C.; Caudill, M.A.; Bottiglieri, T.; French, S.W.; Morgan, T.R. Betaine improves nonalcoholic fatty liver and associated hepatic insulin resistance: A potential mechanism for hepatoprotection by betaine. Am. J. Physiol. Gastrointest Liver Physiol. 2010, 299, G1068–G1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelmalek, M.F.; Sanderson, S.O.; Angulo, P.; Soldevila-Pico, C.; Liu, C.; Peter, J.; Keach, J.; Cave, M.; Chen, T.; McClain, C.J.; et al. Betaine for nonalcoholic fatty liver disease: Results of a randomized placebo-controlled trial. Hepatology 2009, 50, 1818–1826. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.J.; Zhang, H.W.; Zhou, J.Y.; Liu, Y.; Yang, Y.; Chen, X.L.; Zhu, C.H.; Zheng, R.D.; Ling, W.H.; Zhu, H.L. Betaine attenuates hepatic steatosis by reducing methylation of the MTTP promoter and elevating genomic methylation in mice fed a high-fat diet. J. Nutr. Biochem. 2014, 25, 329–336. [Google Scholar] [CrossRef]
- Kishino, Y.; Tanaka, Y.; Ikeda, T.; Yamamoto, K.; Ogawa, H.; Iwatani, Y.; Kamisako, T. Ezetimibe increases hepatic iron levels in mice fed a high-fat diet. J. Pharmacol. Exp. Ther. 2013, 345, 483–491. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Ma, Y.; Yang, L.Y.; Zhao, D. MicroRNA-20a-5p Ameliorates Non-alcoholic Fatty Liver Disease via Inhibiting the Expression of CD36. Front. Cell Dev. Biol. 2020, 8, 596329. [Google Scholar] [CrossRef]
- Huang, D.W.; Lo, Y.M.; Chang, W.C.; Lin, C.Y.; Chen, J.A.; Wu, J.S.; Huang, W.C.; Shen, S.C. Alleviative effect of Ruellia tuberosa L. on NAFLD and hepatic lipid accumulation via modulating hepatic de novo lipogenesis in high-fat diet plus streptozotocin-induced diabetic rats. Food Sci. Nutr. 2020, 8, 5710–5716. [Google Scholar] [CrossRef]
- Shao, D.; Kolwicz, S.C., Jr.; Wang, P.; Roe, N.D.; Villet, O.; Nishi, K.; Hsu, Y.A.; Flint, G.V.; Caudal, A.; Wang, W.; et al. Increasing Fatty Acid Oxidation Prevents High-Fat Diet-Induced Cardiomyopathy Through Regulating Parkin-Mediated Mitophagy. Circulation 2020, 142, 983–997. [Google Scholar] [CrossRef]
- Chang, X.; Yan, H.; Fei, J.; Jiang, M.; Zhu, H.; Lu, D.; Gao, X. Berberine reduces methylation of the MTTP promoter and alleviates fatty liver induced by a high-fat diet in rats. J. Lipid. Res. 2010, 51, 2504–2515. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Li, Y.; Xiao, L. Berberine ameliorates nonalcoholic fatty liver disease by decreasing the liver lipid content via reversing the abnormal expression of MTTP and LDLR. Exp. Ther. Med. 2021, 22, 1109. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Febbraio, M.; Wada, T.; Zhai, Y.; Kuruba, R.; He, J.; Lee, J.H.; Khadem, S.; Ren, S.; Li, S.; et al. Hepatic fatty acid transporter Cd36 is a common target of LXR, PXR, and PPARgamma in promoting steatosis. Gastroenterology 2008, 134, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F.; Cai, Y. Concurrent exercise improves insulin resistance and nonalcoholic fatty liver disease by upregulating PPAR-gamma and genes involved in the beta-oxidation of fatty acids in ApoE-KO mice fed a high-fat diet. Lipids Health Dis. 2019, 18, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, K.P.; Hao, C.L.; Yen, H.W.; Chen, C.Y.; Wu, B.N.; Lin, H.L. Pre-germinated brown rice prevents high-fat diet induced hyperglycemia through elevated insulin secretion and glucose metabolism pathway in C57BL/6J strain mice. J. Clin. Biochem. Nutr. 2015, 56, 28–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Zhang, Y.; Liu, Y.; Zhu, D.; Yu, J.; Li, G.; Sun, Z.; Wang, W.; Jiang, H.; Hong, Z. MiR-27a promotes insulin resistance and mediates glucose metabolism by targeting PPAR-gamma-mediated PI3K/AKT signaling. Aging 2019, 11, 7510–7524. [Google Scholar] [CrossRef] [PubMed]
- Fougerat, A.; Montagner, A.; Loiseau, N.; Guillou, H.; Wahli, W. Peroxisome Proliferator-Activated Receptors and Their Novel Ligands as Candidates for the Treatment of Non-Alcoholic Fatty Liver Disease. Cells 2020, 9, 1638. [Google Scholar] [CrossRef]
- Wang, F.; Mullican, S.E.; DiSpirito, J.R.; Peed, L.C.; Lazar, M.A. Lipoatrophy and severe metabolic disturbance in mice with fat-specific deletion of PPARgamma. Proc. Natl. Acad. Sci. USA 2013, 110, 18656–18661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Huang, D.; Hu, Q.; Wu, J.; Wang, Y.; Feng, J. Betaine alleviates hepatic lipid accumulation via enhancing hepatic lipid export and fatty acid oxidation in rats fed with a high-fat diet. Br. J. Nutr. 2015, 113, 1835–1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, S.; Yan, K.; Sun, B.; Gao, S.; Yang, X.; Ni, Y.; Ma, W.; Zhao, R. Long-Term High-Fat Diet Decreases Hepatic Iron Storage Associated with Suppressing TFR2 and ZIP14 Expression in Rats. J. Agric. Food Chem. 2018, 66, 11612–11621. [Google Scholar] [CrossRef]
- Camaschella, C.; Nai, A.; Silvestri, L. Iron metabolism and iron disorders revisited in the hepcidin era. Haematologica 2020, 105, 260–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Idriss, A.A.; Hu, Y.; Sun, Q.; Hou, Z.; Yang, S.; Omer, N.A.; Abobaker, H.; Zhao, R. Fetal betaine exposure modulates hypothalamic expression of cholesterol metabolic genes in offspring cockerels with modification of promoter DNA methylation. Poult. Sci. 2020, 99, 2533–2542. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Sun, Q.; Zong, Y.; Liu, J.; Idriss, A.A.; Omer, N.A.; Zhao, R. Prenatal betaine exposure alleviates corticosterone-induced inhibition of CYP27A1 expression in the liver of juvenile chickens associated with its promoter DNA methylation. Gen. Comp. Endocrinol. 2017, 246, 241–248. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CON | HF | |||
|---|---|---|---|---|
| Energizing material | gm% | Kcal% | gm% | Kcal% |
| Protein | 19.2 | 20 | 26.2 | 20 |
| Carbohydrate | 67.3 | 70 | 26.3 | 20 |
| Fat | 4.3 | 10 | 34.9 | 60 |
| Total | 100 | 100 | ||
| 3.85 | 5.24 | |||
| Ingredient | gm | Kcal | gm | Kcal |
| Casein, 80 Mesh | 200 | 800 | 200 | 800 |
| l-cystine | 3 | 12 | 3 | 12 |
| Corn starch | 315 | 1260 | 0 | 0 |
| Maltodextrin 10 | 35 | 140 | 125 | 500 |
| Sucrose | 350 | 1400 | 68.8 | 275.2 |
| Cellulose | 50 | 0 | 50 | 0 |
| Soybean Oil | 25 | 255 | 25 | 255 |
| Lard | 20 | 180 | 245 | 2205 |
| Mineral mix S10026 | 10 | 0 | 10 | 0 |
| Dicalcium Phosphate | 13 | 0 | 13 | 0 |
| Calcium Carbonate | 5.5 | 0 | 5.5 | 0 |
| Potassium Citrate,1 H20 | 16.5 | 0 | 16.5 | 0 |
| Vitamin Mix V10001 | 10 | 40 | 10 | 40 |
| Choline Bitartrate | 2 | 0 | 2 | 0 |
| Pigment | 0.05 | 0 | 0.05 | 0 |
| Total | 1055.05 | 4057 | 773.85 | 4057 |
| Target Genes | Primer Sequences (5′to 3′) | |
|---|---|---|
| qPCR PPARγ | F: CTTGCAGTGGGGATGTCTCA | R: CCTCGCCTTTGCTTTGGT |
| CD36 | F: TTGATGTGCAAAATCCACAGG | R: TGTGTTGTCCTCAGCGTCCT |
| ACC1 | F: GGAGATGTACGCTGACCGAG | R: TACCCGACGCATGGTTTTCA |
| FASN | F: GGCCCCTCTGTTAATTGGCT | R: GGATCTCAGGGTTGGGGTTG |
| SCD1 | F: CCTCCGGAAATGAACGAGAGA | R: ATCCCGAAGAGGCAGGTGTA |
| TFR1 | F: TCGGAGAAACTGGACAGCAC | R: ATCACGCCAGACTTTGCTGA |
| TFR2 | F: GGTCTATTCCAGAGAGCGCA | R: CGACGTAGCCCAGTAGGAAG |
| ZIP14 | F: AGAAGGTCATTGTGGGCTCG | R: AGTGAAGGAAGCACCGATGG |
| DMT1 | F: AGCTGTCATCATGCCACACA | R: AGACTTCAACCACCTGCTCG |
| FTL | F: ATTTCGACCGCGATGATGTG | R: CATGGCGTCTGGGGTTTTAC |
| FTH | F: GCCATCAACCGCCAGATCAA | R: AAGATTCGGCCACCTCGTTG |
| FPN | F: GAGATCACAACCGCCAGAGA | R: CACATCCGATCTCCCCAAGT |
| HAMP | F: CTTTGCACGGGGAAGAAAGC | R: TGCAGATGGGGAAGTTGGTG |
| BMP2 | F: TAGTGTTGCTGCTTCCCCAG | R: CTCCACGGCTTCTTCGTGAT |
| BMP6 | F: TCTCCCCACATCAACGACAC | R: AAACTCCCCACCACACAGTC |
| SMAD1 | F: GCCTCTGGAATGCTGTGAGT | R: GAACTGAGCCAGAAGGCTGT |
| SMAD4 | F: TTCAAGCTGCCCTGTTGTGA | R: GACCTTTATATACGCGCTTGG |
| SMAD5 | F: TGTTGGGCTGGAAACAAGGT | R: GTGACACACTTGCTTGGCTG |
| SMAD8 | F: TCAACACTCAGACTTCCGGC | R: TTGAGAGAAGCCCAAGGCAG |
| PPIA | F: GACTGAGTGGTTGGATGG | R: TGATCTTCTTGCTGGTCTT |
| MeDIP | ||
| PPARγ | F: AGAGAGAGAGATGAAAAGCACATC | R: GTGTCCCTCAGACCGATGTC |
| CD36 | F: GCAGGCACAATGTAAGACCAA | R: TGTCATCAACCTCAGCCATTC |
| HAMP | F: TCCCCAAGAGATTGGCTCAC | R: AGGGGCCAACAGGAGTACTT |
| BMP2 | F: GTGGACTCTGGATTTGCCCTA | R: TGAGTGTGAAGCCGACCCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Jiang, W.; Feng, Y.; Wu, L.; Jia, Y.; Zhao, R. Betaine Alleviates High-Fat Diet-Induced Disruptionof Hepatic Lipid and Iron Homeostasis in Mice. Int. J. Mol. Sci. 2022, 23, 6263. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23116263
Li Y, Jiang W, Feng Y, Wu L, Jia Y, Zhao R. Betaine Alleviates High-Fat Diet-Induced Disruptionof Hepatic Lipid and Iron Homeostasis in Mice. International Journal of Molecular Sciences. 2022; 23(11):6263. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23116263
Chicago/Turabian StyleLi, Yanlin, Wenduo Jiang, Yue Feng, Lei Wu, Yimin Jia, and Ruqian Zhao. 2022. "Betaine Alleviates High-Fat Diet-Induced Disruptionof Hepatic Lipid and Iron Homeostasis in Mice" International Journal of Molecular Sciences 23, no. 11: 6263. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23116263