
Development and Preliminary Testing of Porcine Blood-Derived Endothelial-like Cells for Vascular Tissue Engineering Applications: Protocol Optimisation and Seeding of Decellularised Human Saphenous Veins



Abstract
:1. Introduction
2. Results
2.1. ELC Isolation Success
2.2. Cell Characterisation
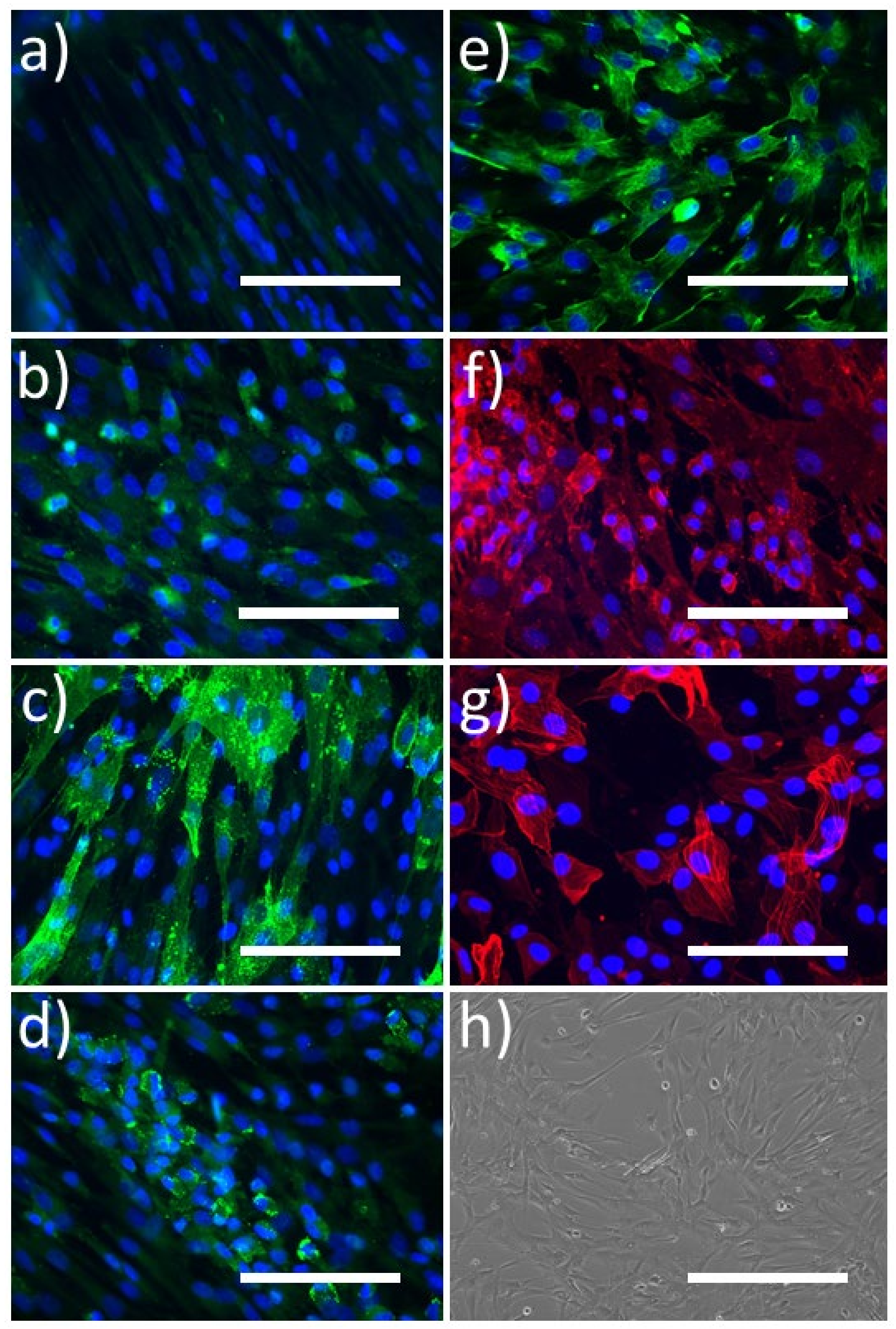
2.2.1. Immunocytochemistry
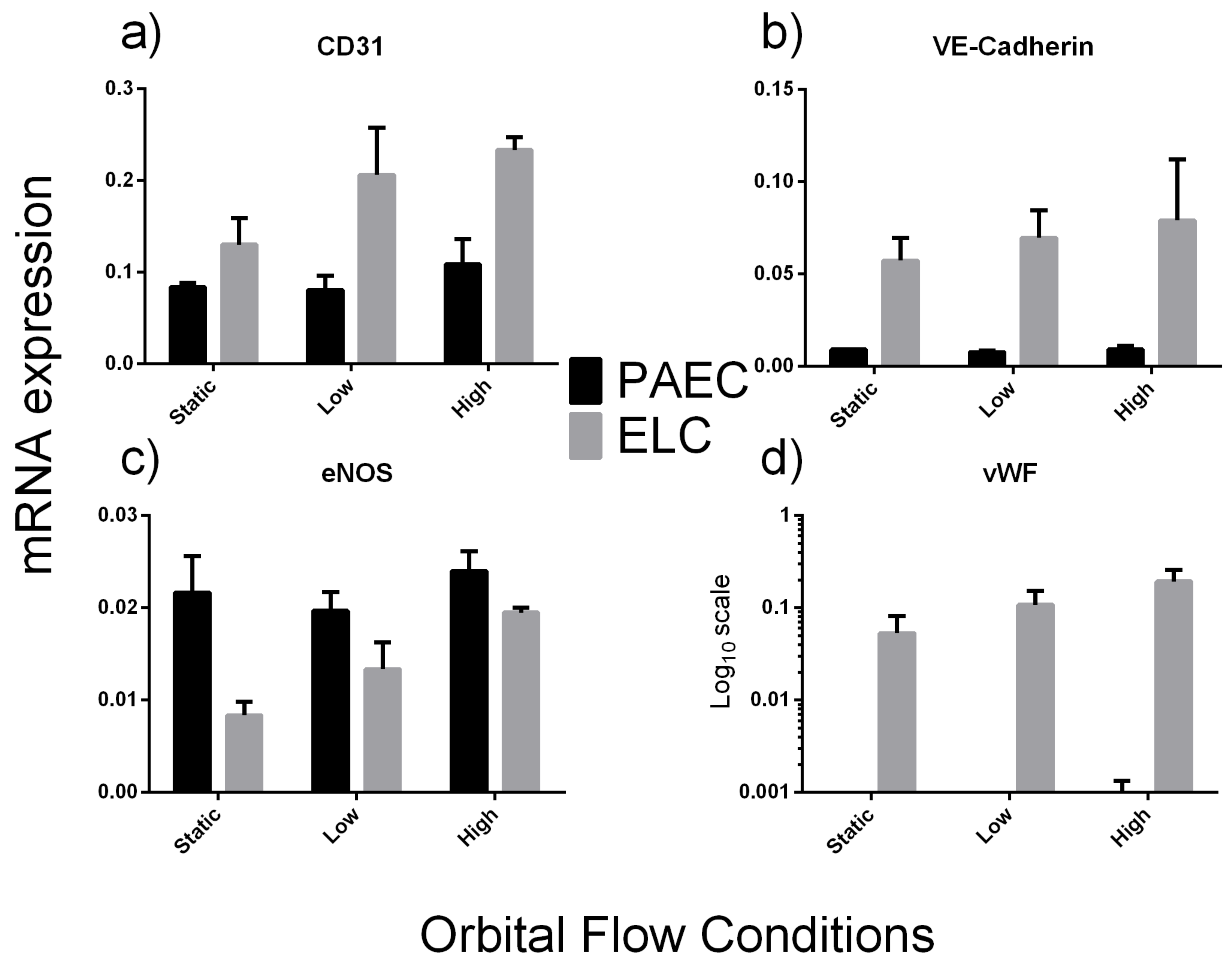
2.2.2. Exposure to Flow Conditions
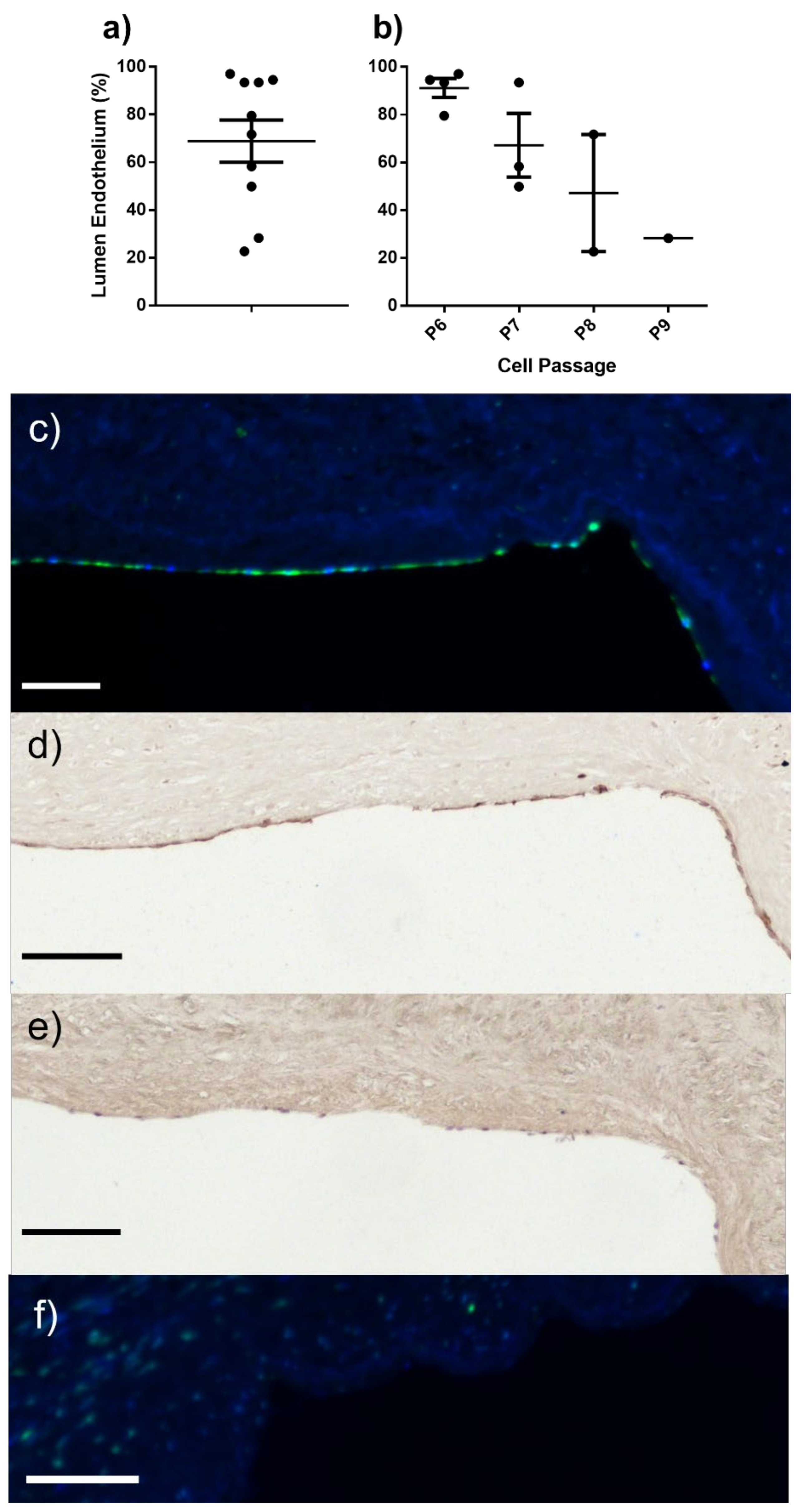
2.3. ELC Seeding/Endothelial Coverage
3. Discussion
Blood Outgrowth Cells (BOCs)
4. Materials and Methods
4.1. Blood Collection
4.2. Cell Isolation
4.3. Red Blood Cell Lysis
4.4. Cell Culture Media
4.5. ELC Characterisation
4.5.1. Immunocytochemistry
4.5.2. Porcine Aortic Endothelial Cell Isolation
4.5.3. Exposure of ELC to Shear Stress
4.5.4. In Vitro Seeding of ELC on Decellularised Human SV (D-hSV)
4.6. mRNA Expression
4.7. Histology
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Type | Orbital Flow Condition | RNA Concentration (ng/µL) | RNA Quality (260/280 Ratio) | RNA Integrity (260/230 Ratio) |
|---|---|---|---|---|
| PAEC #1 | Static | 303.8 | 2.06 | 2.17 |
| Low | 333.3 | 2.04 | 2.07 | |
| High | 305.5 | 2.04 | 2.26 | |
| PAEC #2 | Static | 306.0 | 1.98 | 2.16 |
| Low | 230.9 | 2.00 | 2.24 | |
| High | 201.7 | 2.06 | 2.28 | |
| PAEC #3 | Static | 386.9 | 2.00 | 2.12 |
| Low | 366.7 | 1.99 | 2.20 | |
| High | 396.3 | 2.01 | 2.05 | |
| ELC #1 | Static | 285.6 | 2.01 | 2.15 |
| Low | 246.8 | 1.90 | 1.11 | |
| High | - | - | - | |
| ELC #2 | Static | 276.9 | 2.02 | 2.20 |
| Low | 191.3 | 1.98 | 2.23 | |
| High | 209.5 | 2.03 | 1.39 | |
| ELC #3 | Static | 256.0 | 2.07 | 2.17 |
| Low | 164.7 | 2.02 | 1.52 | |
| High | 232.4 | 2.04 | 1.98 |
| Step | Temperature | Duration | Number of Cycles |
|---|---|---|---|
| Pre-incubation | 95 °C | 5 min | 1 |
| Amplification | Denaturation: 95 °C | 10 s | 45 |
| Annealing: 60 °C | 10 s | ||
| Extension: 72 °C | 10 s | ||
| Melting curve | 95 °C | 5 s | 1 |
| 65 °C | 60 s | ||
| 97 °C | Continuous | ||
| Cooling | 40 °C | 10 s | 1 |
| Primary Antibody | Catalogue No./Supplier | Working Concentration | Secondary Antibody | Catalogue No./Supplier | Working Concentration |
|---|---|---|---|---|---|
| Anti-PECAM-1/CD31 | Ab28364/Abcam | 1.8 μg/mL | Alexa-Fluor 488 Chicken anti-rabbit IgG | A21441/ThermoFisher Scientific | 10 μg/mL |
| Anti-VE-Cadherin (CD144) | Ab33168/Abcam | 3.5 μg/mL | |||
| Anti-vWF * | Ab6994/Abcam | 25 μg/mL | |||
| Anti-SM-MHC ** | Ab53219/Abcam | 2.2 μg/mL | |||
| Anti-rabbit IgG | Ab172730/Abcam | 8.4 μg/mL | |||
| Anti-vimentin | Ab8979/Abcam | 5 μg/mL | AlexaFluor 488 Chicken anti-mouse IgG | A21200/ThermoFisher Scientific | 10 μg/mL |
| Anti-CD45 | MCA1222GA/Bio-rad | 6.7 μg/mL | |||
| Anti-mouse IgG | MAB002/R&D Systems | 5 μg/mL | |||
| Anti-Thy1 (CD90) | AF2067/Biotechne | 4 μg/mL | NorthernLights 557 Donkey anti-sheep IgG | NL010/R&D Systems | 5 μg/mL |
| Anti-sheep IgG | 5–001-A/Biotechne | 4 μg/mL | |||
| DBA-Lectin $$ | B-1035/Vector Laboratories | 25 μg/mL | Alexa-Fluor 488 Streptavidin | S32354/ThermoFisher Scientific | 10 μg/mL |
| Anti-SMA-Cy3 | C6198/Sigma-Aldrich | 2.5 μg/mL | n/a |
| Gene | Description | Primer Sequence (5′ to 3′) |
|---|---|---|
| ACTB | Actin Beta | F: AGATCAAGATCATCGCGCCTCCAGA R: TGGAATGCAACTAACAGTCCGCC |
| PECAM | Platelet and Endothelial Cell Adhesion Molecule 1 (CD31) | F: ACTTCTGAACTCCAACAATG R: GTCAGAGGCTCTTTGTTTTC |
| VWF | Von Willebrand Factor | F: ATCATGAAAATTCCAGGCAC R: TAATGAATGTCCACCTCCTC |
| CDH5 | Vascular Endothelial Cadherin 5 (VE-Cadherin) | F: ATAATCACGATAACACAGCC R: GAGATGAGTATGGGAAGGTG |
| ENOS | Endothelial Nitric Oxide Synthase | F: AGAATGGAGAGAGTTTCGCGGCAG R: AGCAGGAAACGCTATTGAAGCGGA |
| THY1 | Thy-1 Cell Surface Antigen (CD90) | F: CTCTCTTGCTAACAGTCTTG R: AGGTTGGTGGTATTCTCATG |
References
- Harskamp, R.E.; Lopes, R.D.; Baisden, C.E.; de Winter, R.J.; Alexander, J.H. Saphenous vein graft failure after coronary artery bypass surgery: Pathophysiology, management, and future directions. Ann. Surg. 2013, 257, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Watson, H.R.; Buth, J.; Schroeder, T.V.; Simms, M.H.; Horrocks, M. Incidence of stenoses in femorodistal bypass vein grafts in a multicentre study. Eur. J. Vasc. Endovasc. Surg. 2000, 20, 67–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodney, P.P.; Beck, A.W.; Nagle, J.; Welch, H.G.; Zwolak, R.M. National trends in lower extremity bypass surgery, endovascular interventions, and major amputations. J. Vasc. Surg. 2009, 50, 54–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, D. A new no-touch preparation technique. Technical notes. Scand. J. Thorac. Cardiovasc. Surg. 1996, 30, 41–44. [Google Scholar] [CrossRef]
- Samano, N.; Souza, D.; Pinheiro, B.B.; Kopjar, T.; Dashwood, M. Twenty-Five Years of No-Touch Saphenous Vein Harvesting for Coronary Artery Bypass Grafting: Structural Observations and Impact on Graft Performance. Braz. J. Cardiovasc. Surg. 2020, 35, 91–99. [Google Scholar] [CrossRef]
- Tsui, J.C.; Souza, D.S.; Filbey, D.; Bomfim, V.; Dashwood, M.R. Preserved endothelial integrity and nitric oxide synthase in saphenous vein grafts harvested by a ‘no-touch’ technique. Br. J. Surg. 2001, 88, 1209–1215. [Google Scholar] [CrossRef]
- Hinokiyama, K.; Valen, G.; Tokuno, S.; Vedin, J.B.; Vaage, J. Vein graft harvesting induces inflammation and impairs vessel reactivity. Ann. Thorac. Surg. 2006, 82, 1458–1464. [Google Scholar] [CrossRef]
- Alrawi, S.J.; Raju, R.; Alshkaki, G.; Acinapura, A.J.; Cunningham, J.N., Jr. Saphenous vein endothelial cell viability: A comparative study of endoscopic and open saphenectomy for coronary artery bypass grafting. JSLS J. Soc. Laparoendosc. Surg. 2001, 5, 37–45. [Google Scholar]
- Thatte, H.S.; Biswas, K.S.; Najjar, S.F.; Birjiniuk, V.; Crittenden, M.D.; Michel, T.; Khuri, S.F. Multi-photon microscopic evaluation of saphenous vein endothelium and its preservation with a new solution, GALA. Ann. Thorac. Surg. 2003, 75, 1145–1152; discussion 1152. [Google Scholar] [CrossRef]
- Medina, R.J.; Barber, C.L.; Sabatier, F.; Dignat-George, F.; Melero-Martin, J.M.; Khosrotehrani, K.; Ohneda, O.; Randi, A.M.; Chan, J.K.Y.; Yamaguchi, T.; et al. Endothelial Progenitors: A Consensus Statement on Nomenclature. Stem Cells Transl. Med. 2017, 6, 1316–1320. [Google Scholar] [CrossRef]
- Paschalaki, K.E.; Randi, A.M. Recent Advances in Endothelial Colony Forming Cells Toward Their Use in Clinical Translation. Front. Med. 2018, 5, 295. [Google Scholar] [CrossRef] [Green Version]
- Ardila, D.C.; Liou, J.J.; Maestas, D.; Slepian, M.J.; Badowski, M.; Wagner, W.R.; Harris, D.; Vande Geest, J.P. Surface Modification of Electrospun Scaffolds for Endothelialization of Tissue-Engineered Vascular Grafts Using Human Cord Blood-Derived Endothelial Cells. J. Clin. Med. 2019, 8, 185. [Google Scholar] [CrossRef] [Green Version]
- Rotmans, J.I.; Heyligers, J.M.; Stroes, E.S.; Pasterkamp, G. Endothelial progenitor cell-seeded grafts: Rash and risky. Can. J. Cardiol. 2006, 22, 929–932. [Google Scholar] [CrossRef]
- Zhou, M.; Qiao, W.; Liu, Z.; Shang, T.; Qiao, T.; Mao, C.; Liu, C. Development and in vivo evaluation of small-diameter vascular grafts engineered by outgrowth endothelial cells and electrospun chitosan/poly(epsilon-caprolactone) nanofibrous scaffolds. Tissue Eng. Part A 2014, 20, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Glynn, J.J.; Hinds, M.T. Endothelial outgrowth cells: Function and performance in vascular grafts. Tissue Eng. Part B Rev. 2014, 20, 294–303. [Google Scholar] [CrossRef] [Green Version]
- Stroncek, J.D.; Ren, L.C.; Klitzman, B.; Reichert, W.M. Patient-derived endothelial progenitor cells improve vascular graft patency in a rodent model. Acta Biomater. 2012, 8, 201–208. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Weisdorf, D.J.; Solovey, A.; Hebbel, R.P. Origins of circulating endothelial cells and endothelial outgrowth from blood. J. Clin. Investig. 2000, 105, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Hebbel, R.P. Blood endothelial cells: Utility from ambiguity. J. Clin. Investig. 2017, 127, 1613–1615. [Google Scholar] [CrossRef] [Green Version]
- Smadja, D.M.; Melero-Martin, J.M.; Eikenboom, J.; Bowman, M.; Sabatier, F.; Randi, A.M. Standardization of methods to quantify and culture endothelial colony-forming cells derived from peripheral blood: Position paper from the International Society on Thrombosis and Haemostasis SSC. J. Thromb. Haemost. 2019, 17, 1190–1194. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, N.A.; Reinisch, A.; Strunk, D. Isolation and large scale expansion of adult human endothelial colony forming progenitor cells. JoVE J. Vis. Exp. 2009, 32, e1524. [Google Scholar] [CrossRef] [Green Version]
- Reinisch, A.; Hofmann, N.A.; Obenauf, A.C.; Kashofer, K.; Rohde, E.; Schallmoser, K.; Flicker, K.; Lanzer, G.; Linkesch, W.; Speicher, M.R.; et al. Humanized large-scale expanded endothelial colony-forming cells function in vitro and in vivo. Blood 2009, 113, 6716–6725. [Google Scholar] [CrossRef] [Green Version]
- Aper, T.; Teebken, O.E.; Kruger, A.; Heisterkamp, A.; Hilfiker, A.; Haverich, A. Development of implantable autologous small-calibre vascular grafts from peripheral blood samples. Zentralbl. Chir. 2013, 138, 173–179. [Google Scholar] [CrossRef]
- Dubois, C.; Liu, X.; Claus, P.; Marsboom, G.; Pokreisz, P.; Vandenwijngaert, S.; Depelteau, H.; Streb, W.; Chaothawee, L.; Maes, F.; et al. Differential effects of progenitor cell populations on left ventricular remodeling and myocardial neovascularization after myocardial infarction. J. Am. Coll. Cardiol. 2010, 55, 2232–2243. [Google Scholar] [CrossRef]
- Huang, L.; Hou, D.; Thompson, M.A.; Baysden, S.E.; Shelley, W.C.; Ingram, D.A.; March, K.L.; Yoder, M.C. Acute myocardial infarction in swine rapidly and selectively releases highly proliferative endothelial colony forming cells (ECFCs) into circulation. Cell Transplant. 2007, 16, 887–897. [Google Scholar] [CrossRef]
- Kim, J.H.; Oh, B.J.; Lee, H.N.; Park, H.S.; Park, S.G.; Park, K.S. Endothelial colony-forming cell coating of pig islets prevents xenogeneic instant blood-mediated inflammatory reaction. Cell Transplant. 2011, 20, 1805–1815. [Google Scholar] [CrossRef]
- Jung, H.S.; Kim, M.J.; Hong, S.H.; Lee, Y.J.; Kang, S.; Lee, H.; Chung, S.S.; Park, J.S.; Park, K.S. The potential of endothelial colony-forming cells to improve early graft loss after intraportal islet transplantation. Cell Transplant. 2014, 23, 273–283. [Google Scholar] [CrossRef] [Green Version]
- Hughes, D.; Fu, A.A.; Puggioni, A.; Glockner, J.F.; Anwer, B.; McGuire, A.M.; Mukhopadhyay, D.; Misra, S. Adventitial transplantation of blood outgrowth endothelial cells in porcine haemodialysis grafts alleviates hypoxia and decreases neointimal proliferation through a matrix metalloproteinase-9-mediated pathway—A pilot study. Nephrol. Dial. Transplant. 2009, 24, 85–96. [Google Scholar] [CrossRef]
- Wang, W.; Lang, J.K.; Suzuki, G.; Canty, J.M., Jr.; Cimato, T. Statins enhance clonal growth of late outgrowth endothelial progenitors and increase myocardial capillary density in the chronically ischemic heart. PLoS ONE 2011, 6, e24868. [Google Scholar] [CrossRef] [Green Version]
- Kolster, M.; Wilhelmi, M.; Schrimpf, C.; Hilfiker, A.; Haverich, A.; Aper, T. Outgrowing endothelial and smooth muscle cells for tissue engineering approaches. J. Tissue Eng. 2017, 8, 2041731417698852. [Google Scholar] [CrossRef] [Green Version]
- Stroncek, J.D.; Grant, B.S.; Brown, M.A.; Povsic, T.J.; Truskey, G.A.; Reichert, W.M. Comparison of endothelial cell phenotypic markers of late-outgrowth endothelial progenitor cells isolated from patients with coronary artery disease and healthy volunteers. Tissue Eng. Part A 2009, 15, 3473–3486. [Google Scholar] [CrossRef] [Green Version]
- Kolbe, M.; Dohle, E.; Katerla, D.; Kirkpatrick, C.J.; Fuchs, S. Enrichment of outgrowth endothelial cells in high and low colony-forming cultures from peripheral blood progenitors. Tissue Eng. Part C Methods 2010, 16, 877–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solovey, A.; Lin, Y.; Browne, P.; Choong, S.; Wayner, E.; Hebbel, R.P. Circulating activated endothelial cells in sickle cell anemia. N. Engl. J. Med. 1997, 337, 1584–1590. [Google Scholar] [CrossRef] [PubMed]
- Hubert, L.; Darbousset, R.; Panicot-Dubois, L.; Robert, S.; Sabatier, F.; Fallague, K.; Dignat-George, F.; Dubois, C. Neutrophils recruit and activate human endothelial colony-forming cells at the site of vessel injury via P-selectin glycoprotein ligand-1 and L-selectin. J. Thromb. Haemost. 2014, 12, 1170–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, S.S.; Solomon, M.A.; McCoy, J.P., Jr. Detection of circulating endothelial cells and endothelial progenitor cells by flow cytometry. Cytom. B Clin. Cytom. 2005, 64, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Antonelou, M.H.; Kriebardis, A.G.; Papassideri, I.S. Aging and death signalling in mature red cells: From basic science to transfusion practice. Blood Transfus. 2010, 8 (Suppl. S3), s39–s47. [Google Scholar] [CrossRef] [PubMed]
- Foller, M.; Huber, S.M.; Lang, F. Erythrocyte programmed cell death. IUBMB Life 2008, 60, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Fielder, S.E. Hematologic Reference Ranges. In The Merck Veterinary Manual; Merck & Co., Inc.: Whitehouse Station, NJ, USA, 2015. [Google Scholar]
- Dean, L. Blood Groups and Red Cell Antigens; National Center for Biotechnology Information: Bethesda, MD, USA, 2005.
- Wojakowski, W.; Tendera, M.; Michalowska, A.; Majka, M.; Kucia, M.; Maslankiewicz, K.; Wyderka, R.; Ochala, A.; Ratajczak, M.Z. Mobilization of CD34/CXCR4+, CD34/CD117+, c-met+ stem cells, and mononuclear cells expressing early cardiac, muscle, and endothelial markers into peripheral blood in patients with acute myocardial infarction. Circulation 2004, 110, 3213–3220. [Google Scholar] [CrossRef] [Green Version]
- Leone, A.M.; Rutella, S.; Bonanno, G.; Abbate, A.; Rebuzzi, A.G.; Giovannini, S.; Lombardi, M.; Galiuto, L.; Liuzzo, G.; Andreotti, F.; et al. Mobilization of bone marrow-derived stem cells after myocardial infarction and left ventricular function. Eur. Heart J. 2005, 26, 1196–1204. [Google Scholar] [CrossRef] [Green Version]
- Massa, M.; Rosti, V.; Ferrario, M.; Campanelli, R.; Ramajoli, I.; Rosso, R.; De Ferrari, G.M.; Ferlini, M.; Goffredo, L.; Bertoletti, A.; et al. Increased circulating hematopoietic and endothelial progenitor cells in the early phase of acute myocardial infarction. Blood 2005, 105, 199–206. [Google Scholar] [CrossRef] [Green Version]
- Chanimov, M.; Berman, S.; Weissgarten, J.; Averbukh, Z.; Cohen, M.; Grinshpun, Y.; Bahar, M. Substances used for local and general anaesthesia in major surgery suppress proliferative responsiveness of normal rat peripheral blood mononuclear cells in culture. Eur. J. Anaesthesiol. 2000, 17, 248–255. [Google Scholar] [CrossRef]
- Zawadka, M.; Wahome, J.; Oszkiel, H.; Szeto, W.Y.; Cobb, B.; Laudanski, K. Long-term alterations in monocyte function after elective cardiac surgery. Anaesthesia 2017, 72, 879–888. [Google Scholar] [CrossRef] [Green Version]
- Fiechter, M.; Stehli, J.; Fuchs, T.A.; Dougoud, S.; Gaemperli, O.; Kaufmann, P.A. Impact of cardiac magnetic resonance imaging on human lymphocyte DNA integrity. Eur. Heart J. 2013, 34, 2340–2345. [Google Scholar] [CrossRef] [Green Version]
- Touitou, Y.; Djeridane, Y.; Lambrozo, J.; Camus, F.; Selmaoui, B. Long-term (up to 20 years) effects of 50-Hz magnetic field exposure on immune system and hematological parameters in healthy men. Clin. Biochem. 2013, 46, 59–63. [Google Scholar] [CrossRef]
- Onodera, H.; Jin, Z.; Chida, S.; Suzuki, Y.; Tago, H.; Itoyama, Y. Effects of 10-T static magnetic field on human peripheral blood immune cells. Radiat. Res. 2003, 159, 775–779. [Google Scholar] [CrossRef]
- Gavard, J. Endothelial permeability and VE-cadherin: A wacky comradeship. Cell Adhes. Migr. 2014, 8, 158–164. [Google Scholar] [CrossRef] [Green Version]
- Conway, D.E.; Schwartz, M.A. Flow-dependent cellular mechanotransduction in atherosclerosis. J. Cell Sci. 2013, 126, 5101–5109. [Google Scholar] [CrossRef] [Green Version]
- Tzima, E.; Irani-Tehrani, M.; Kiosses, W.B.; Dejana, E.; Schultz, D.A.; Engelhardt, B.; Cao, G.; DeLisser, H.; Schwartz, M.A. A mechanosensory complex that mediates the endothelial cell response to fluid shear stress. Nature 2005, 437, 426–431. [Google Scholar] [CrossRef]
- Conway, D.E.; Breckenridge, M.T.; Hinde, E.; Gratton, E.; Chen, C.S.; Schwartz, M.A. Fluid shear stress on endothelial cells modulates mechanical tension across VE-cadherin and PECAM-1. Curr. Biol. 2013, 23, 1024–1030. [Google Scholar] [CrossRef] [Green Version]
- De Jong, A.; Weijers, E.; Dirven, R.; de Boer, S.; Streur, J.; Eikenboom, J. Variability of von Willebrand factor-related parameters in endothelial colony forming cells. J. Thromb. Haemost. 2019, 17, 1544–1554. [Google Scholar] [CrossRef] [Green Version]
- Luo, G.P.; Ni, B.; Yang, X.; Wu, Y.Z. von Willebrand factor: More than a regulator of hemostasis and thrombosis. Acta Haematol. 2012, 128, 158–169. [Google Scholar] [CrossRef]
- Fleming, I.; Busse, R. Signal transduction of eNOS activation. Cardiovasc. Res. 1999, 43, 532–541. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.E.; Cai, H.; Drummond, G.R.; Harrison, D.G. Shear stress regulates endothelial nitric oxide synthase expression through c-Src by divergent signaling pathways. Circ. Res. 2001, 89, 1073–1080. [Google Scholar] [CrossRef] [Green Version]
- Warboys, C.M.; Ghim, M.; Weinberg, P.D. Understanding mechanobiology in cultured endothelium: A review of the orbital shaker method. Atherosclerosis 2019, 285, 170–177. [Google Scholar] [CrossRef] [Green Version]
- Sulaiman, N.S.; Bond, A.R.; Bruno, V.D.; Joseph, J.; Johnson, J.L.; Suleiman, M.S.; George, S.J.; Ascione, R. Effective decellularisation of human saphenous veins for biocompatible arterial tissue engineering applications: Bench optimisation and feasibility in vivo testing. J. Tissue Eng. 2021, 12, 2041731420987529. [Google Scholar] [CrossRef]
- Hong, X.; Le Bras, A.; Margariti, A.; Xu, Q. Reprogramming towards endothelial cells for vascular regeneration. Genes Dis. 2016, 3, 186–197. [Google Scholar] [CrossRef] [Green Version]
- Margariti, A.; Winkler, B.; Karamariti, E.; Zampetaki, A.; Tsai, T.N.; Baban, D.; Ragoussis, J.; Huang, Y.; Han, J.D.; Zeng, L.; et al. Direct reprogramming of fibroblasts into endothelial cells capable of angiogenesis and reendothelialization in tissue-engineered vessels. Proc. Natl. Acad. Sci. USA 2012, 109, 13793–13798. [Google Scholar] [CrossRef] [Green Version]
- Karamariti, E.; Margariti, A.; Winkler, B.; Wang, X.; Hong, X.; Baban, D.; Ragoussis, J.; Huang, Y.; Han, J.D.; Wong, M.M.; et al. Smooth muscle cells differentiated from reprogrammed embryonic lung fibroblasts through DKK3 signaling are potent for tissue engineering of vascular grafts. Circ. Res. 2013, 112, 1433–1443. [Google Scholar] [CrossRef] [Green Version]
- Ahmetaj-Shala, B.; Marei, I.; Kawai, R.; Rothery, S.; Pericleous, C.; Mohamed, N.A.; Gashaw, H.; Bokea, K.; Samuel, J.; Vandenheste, A.; et al. Activation and Contraction of Human "Vascular" Smooth Muscle Cells Grown from Circulating Blood Progenitors. Front. Cell Dev. Biol. 2021, 9, 681347. [Google Scholar] [CrossRef]
- Dardik, A.; Chen, L.; Frattini, J.; Asada, H.; Aziz, F.; Kudo, F.A.; Sumpio, B.E. Differential effects of orbital and laminar shear stress on endothelial cells. J. Vasc. Surg. 2005, 41, 869–880. [Google Scholar] [CrossRef] [Green Version]




| Isolations | Previous Procedure | Anaesthetic | Transport | Time to Isolation | Centrifuge Time | RBC Lysis | Media | Cell Growth | ||
|---|---|---|---|---|---|---|---|---|---|---|
| ELC | BOC | No Growth | ||||||||
| 28 Pigs = 43 Batches | Yes 12 Pigs 17 Batches | Short 4 Pigs 4 Batches | Yes 4 Pigs 4 Batches | >30 min 4 Pigs 4 Batches | 10–12 min 4 Pigs 4 Batches | Yes 3 Pigs 3 Batches | Lonza 3 Pigs 3 Batches | 1 * | 0 | 2 |
| No 1 Pig 1 Batch | Lonza 1 Pig 1 Batch | 0 | 0 | 1 | ||||||
| Long 8 Pigs 13 Batches | Yes 6 Pigs 10 Batches | >30 min 6 Pigs 10 Batches | 10–12 min 6 Pigs 10 Batches | Yes 6 Pigs 9 Batches | Lonza 2 Pigs 3 Batches | 0 | 0 | 3 | ||
| Promocell 4 Pigs 6 Batches | 0 | 0 | 6 | |||||||
| No 1 Pig 1 Batch | Promocell 1 Pig 1 Batch | 0 | 0 | 1 | ||||||
| No 2 Pigs 3 Batches | Immediate 1 Pig 1 Batch | 20 min 1 Pig 1 Batch | No 1 Pig 1 Batch | Lonza 1 Pig, 1 Batch | 0 | 0 | 1 | |||
| >30 min 1 Pig 2 Batches | 20 min 1 Pig, 2 Batches | Yes 1 Pig, 1 Batch | Lonza 1 Pig 1 Batch | 0 | 0 | 1 | ||||
| No 1 Pig 1 Batch | Lonza 1 Pig 1 Batch | 0 | 0 | 1 | ||||||
| No 16 Pigs 26 Batches | Short 15 Pigs 24 Batches | Yes 4 Pigs 6 Batches | >30 min 4 Pigs 6 Batches | 10–12 min 3 Pigs 5 Batches | Yes 3 Pigs 4 Batches | Lonza 1 Pig 1 Batch | 0 | 1 | 0 | |
| Promocell 2 Pigs 3 Batches | 0 | 0 | 3 | |||||||
| No 1 Pig 1 Batch | Promocell 1 Pig 1 Batch | 0 | 0 | 1 | ||||||
| 20 min 1 Pig 1 Batch | Yes 1 Pig 1 Batch | Lonza 1 Pig 1 Batch | 0 | 0 | 1 | |||||
| No 11 Pigs 18 Batches | Immediate 7 Pigs 9 Batches | 20 min 7 Pigs 9 Batches | Yes 7 Pigs 7 Batches | Lonza 7 Pigs 7 Batches | 4 | 1 | 2 | |||
| No 2 Pigs 2 Batches | Lonza 2 Pigs 2 Batches | 2 | 0 | 0 | ||||||
| >30 min 5 Pigs 9 Batches | 20 min 5 Pigs 9 Batches | Yes 5 pigs 6 Batches | Lonza 5 pigs 6 Batches | 1 * | 4 | 1 | ||||
| No 3 Pigs 3 Batches | Lonza 3 pigs 3 Batches | 0 | 2 | 1 | ||||||
| Long 1 Pig 2 Batches | No 1 Pig 2 Batches | >30 min 1 Pig 2 Batches | 20 min 1 Pig 2 Batches | Yes 1 Pig 1 Batch | Lonza 1 Pig 1 Batch | 1 * | 0 | 0 | ||
| No 1 Pig 1 Batch | Lonza 1 Pig 1 Batch | 1 * | 0 | 0 | ||||||
| Isolations | Anticoagulant | Previous Procedure | Anaesthetic | Transport | Time to Isolation | Centrifuge Time | RBC Lysis | Media | Cell Growth | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ELC | BOC | No Growth | |||||||||
| 5 pigs 10 Batches | EDTA | No | None | Yes | >30 min | 20 min | No | Lonza | 0 | 10 | 0 |
| 2 Pigs 4 Batches | Heparin | No | None | Yes | >30 min | 20 min | No | Lonza | 0 | 4 | 0 |
| CD31 | VE-Cadherin | eNOS | vWF | ||
|---|---|---|---|---|---|
| Aortic Endothelial Cells (PAEC) | Static (n = 3) | 1.0 ± 0.0 | 1.0 ± 0.0 | 1.0 ± 0.2 | 1.3 ± 0.6 |
| Low (n = 3) | 1.0 ± 0.2 | 0.9 ± 0.1 | 0.9 ± 0.1 | 0.5 ± 0.2 | |
| High (n = 3) | 1.3 ± 0.3 | 1.0 ± 0.2 | 1.2 ± 0.1 | 3.6 ± 3.0 | |
| Endothelial-like Cells (ELC) | Static (n = 3) | 1.6 ± 0.3 | 6.5 ± 1.4 | 0.4 ± 0.1 | 300.6 ± 155.1 |
| Low (n = 3) | 2.5 ± 0.6 | 7.8 ± 1.7 | 0.7 ± 0.1 | 609.6 ± 251.0 | |
| High (n = 2) | 2.8 ± 0.2 | 8.8 ± 3.7 | 0.9 ± 0.0 | 1088.1 ± 356.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bond, A.; Bruno, V.; Johnson, J.; George, S.; Ascione, R. Development and Preliminary Testing of Porcine Blood-Derived Endothelial-like Cells for Vascular Tissue Engineering Applications: Protocol Optimisation and Seeding of Decellularised Human Saphenous Veins. Int. J. Mol. Sci. 2022, 23, 6633. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23126633
Bond A, Bruno V, Johnson J, George S, Ascione R. Development and Preliminary Testing of Porcine Blood-Derived Endothelial-like Cells for Vascular Tissue Engineering Applications: Protocol Optimisation and Seeding of Decellularised Human Saphenous Veins. International Journal of Molecular Sciences. 2022; 23(12):6633. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23126633
Chicago/Turabian StyleBond, Andrew, Vito Bruno, Jason Johnson, Sarah George, and Raimondo Ascione. 2022. "Development and Preliminary Testing of Porcine Blood-Derived Endothelial-like Cells for Vascular Tissue Engineering Applications: Protocol Optimisation and Seeding of Decellularised Human Saphenous Veins" International Journal of Molecular Sciences 23, no. 12: 6633. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23126633