
Dysregulation of the Enteric Nervous System in the Mid Colon of Complement Component 3 Knockout Mice with Constipation Phenotypes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Confirmation of the Constipation Phenotypes in C3 KO Mice
2.2. Alteration of the Makers for Neuronal Cells and ICC in the Mid Colon of C3 KO Mice
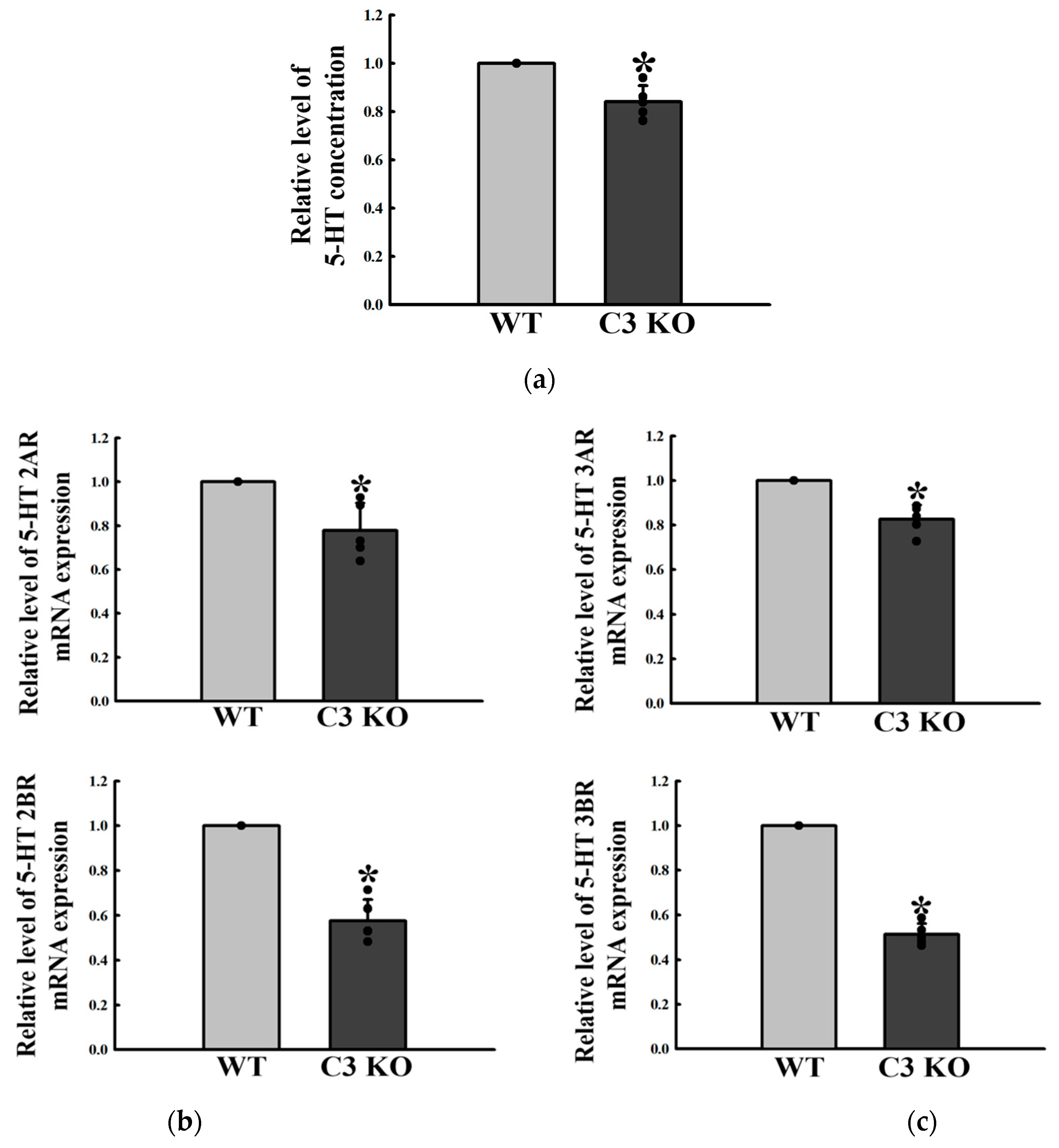
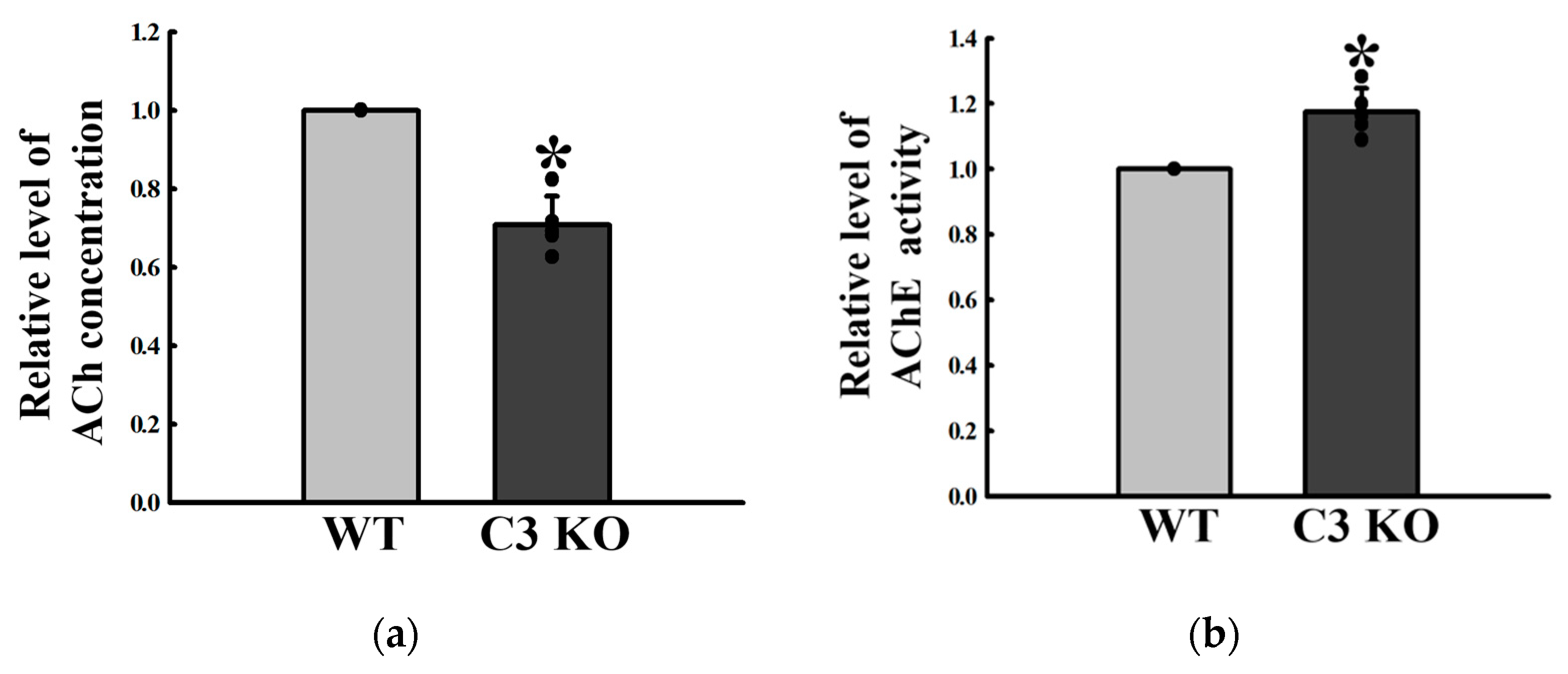
2.3. Changes in the Excitatory Function of ENS in the Mid Colon of C3 KO Mice
2.4. Change in the Inhibitory Function of ENS in the Mid Colon of C3 KO Mice
2.5. Alterations on the Concentration of GI Hormones in the Mid Colon of C3 KO Mice
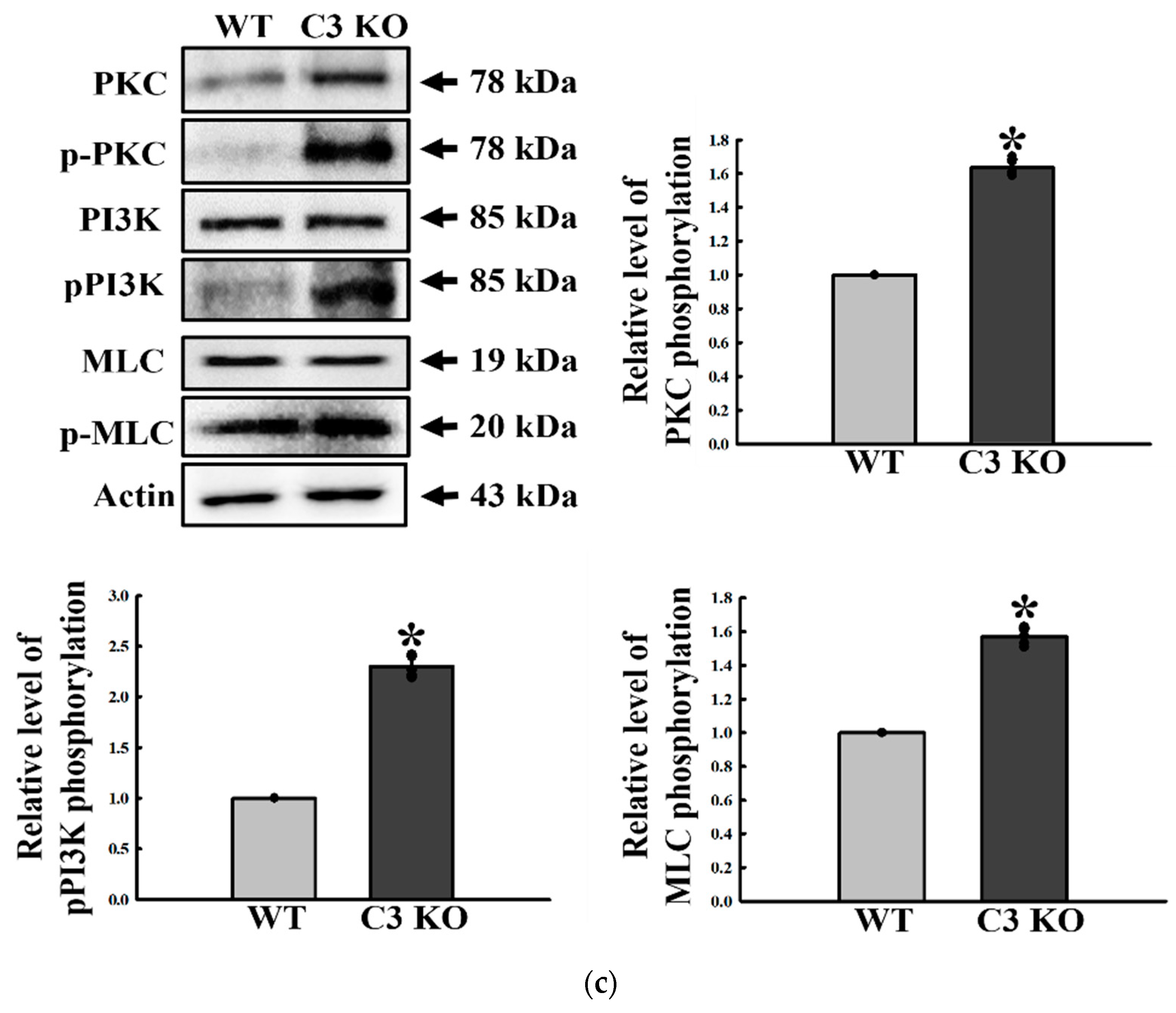
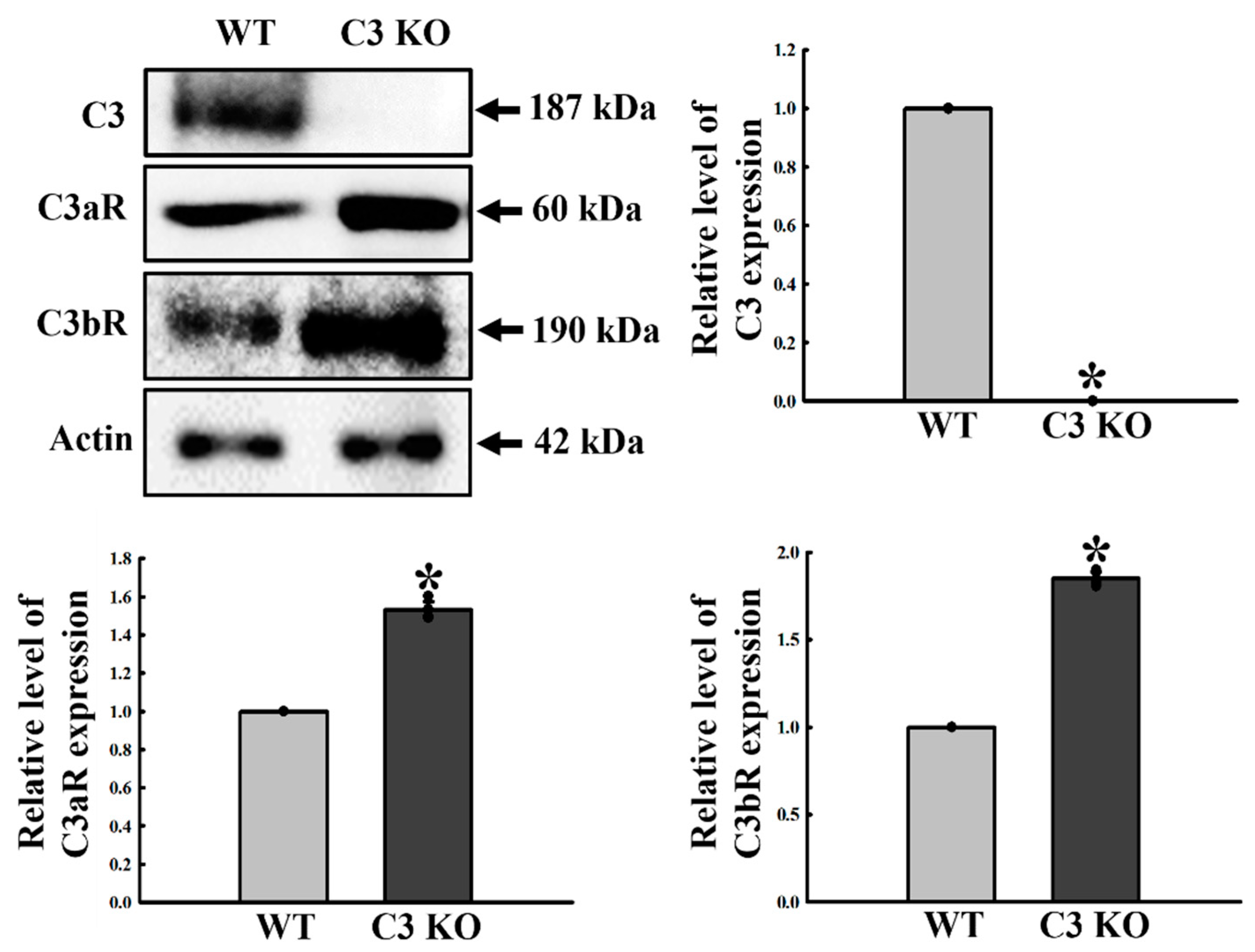
2.6. Verification of the Correlation between ENS Function and C3R
3. Discussion
4. Materials and Methods
4.1. Management and Breeding of C3 KO Mice
4.2. Stool Parameter Analysis
4.3. Histopathological Analysis
4.4. Western Blotting Analysis
4.5. qRT-PCR Analysis
4.6. Measurement of 5-HT Concentrations
4.7. Determination of ACh Concentration
4.8. Determination of AChE Activity
4.9. Determination of the NO Concentration
4.10. Measurement of GI Hormone Concentrations
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Rawal, N.; Pangburn, M.K. Formation of high affinity C5 convertase of the classical pathway of complement. J. Biol. Chem. 2003, 278, 38476–38483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, X.; Liu, L.; Gamallat, Y.; Zhang, B.; Xin, Y. Enteromorpha and polysaccharides from enteromorpha ameliorate loperamide-induced constipation in mice. Biomed. Pharmacother. 2017, 96, 1075–1081. [Google Scholar] [CrossRef] [PubMed]
- Lokki, A.; Teirilä, L.; Triebwasser, M.; Daly, E.; Bhattacharjee, A.; Uotila, L.; Asens, M.L.; Kurki, M.I.; Perola, M.; Auro, K.; et al. Dysfunction of complement receptors CR3 (CD11b/18) and CR4 (CD11c/18) in pre-eclampsia: A genetic and functional study. Int. J. Gynaecol. Obstet. 2021, 128, 1282–1291. [Google Scholar] [CrossRef]
- Riezzo, G.; Chimienti, G.; Orlando, A.; D’Attoma, B.; Clemente, C.; Russo, F. Effects of long-term administration of Lactobacillus reuteri DSM-17938 on circulating levels of 5-HT and BDNF in adults with functional constipation. Benef. Microbes 2019, 10, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Druart, M.; Magueresse, C.L. Emerging roles of complement in psychiatric disorders. Front. Psychiatry 2019, 10, 573. [Google Scholar] [CrossRef] [PubMed]
- Lian, H.; Litvinchuk, A.; Chiang, A.C.; Aithmitti, N.; Jankowsky, J.L.; Zheng, H. Astrocyte-microglia cross talk through complement activation modulates amyloid pathology in mouse models of Alzheimer’s disease. J. Neurosci. 2016, 36, 577–589. [Google Scholar] [CrossRef] [Green Version]
- Gorelik, A.; Sapir, T.; Haffner-Krausz, R.; Olender, T.; Woodruff, T.M.; Reiner, O. Developmental activities of the complement pathway in migrating neurons. Nat. Commun. 2017, 8, 15096. [Google Scholar] [CrossRef] [PubMed]
- Nishida, N.; Walz, T.; Springer, T.A. Structural transitions of complement component C3 and its activation products. Proc. Natl. Acad. Sci. USA 2006, 103, 19737–19742. [Google Scholar] [CrossRef] [Green Version]
- Shah, S.; Hobbs, A.; Singh, R.; Cuevas, J.; Ignarro, L.J.; Chaudhuri, G. Gastrointestinal motility during pregnancy: Role of nitrergic component of NANC nerves. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R1478–R1485. [Google Scholar] [CrossRef] [Green Version]
- Ward, S.M.; Xue, C.; Shuttleworth, C.W.; Bredt, D.S.; Snyder, S.H.; Sanders, K.M. NADPH diaphorase and nitric oxide synthase colocalization in enteric neurons of canine proximal colon. Am. J. Physiol. Gastrointest. Liver. Physiol. 1992, 263, G277–G284. [Google Scholar] [CrossRef]
- Hong, S.; Beja-Glasser, V.F.; Nfonoyim, B.M.; Frouin, A.; Li, S.; Ramakrishnan, S.; Merry, K.M.; Shi, Q.; Rosenthal, A.; Barres, B.A.; et al. Complement and microglia mediate early synapse loss in Alzheimer mouse models. Science 2016, 352, 712–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Litvinchuk, A.; Wan, Y.W.; Swartzlander, D.B.; Chen, F.; Cole, A.; Propson, N.E.; Wang, Q.; Zhang, B.; Liu, Z.; Zheng, H. Complement C3aR inactivation attenuates tau pathology and reverses an immune network deregulated in tauopathy models and Alzheimer’s disease. Neuron 2018, 100, 1337–1353.e5. [Google Scholar] [CrossRef] [PubMed]
- Laufer, J.; Oren, R.; Goldberg, I.; Horwitz, A.; Kopolovic, J.; Chowers, Y.; Passwell, J.H. Cellular localization of complement C3 and C4 transcripts in intestinal specimens from patients with Crohn’s disease. Clin. Exp. Immunol. 2000, 120, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Tomita, R.; Fujisaki, S.; Ikeda, T.; Fukuzawa, M. Role of nitric oxide in the colon of patients with slow-transit constipation. Dis. Colon Rectum 2002, 45, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Ahrenstedt, O.; Knutson, L.; Nilsson, B.; Nilsson-Ekdahl, K.; Odilind, B.; Hallgren, R. Enhanced local production of complement components in the small intestines of patients with Crohn’s disease. N. Engl. J. Med. 1990, 322, 1345–1349. [Google Scholar] [CrossRef]
- Halstensen, T.S.; Mollnes, T.E.; Garred, P.; Fausa, O.; Brandtzaeg, P. Surface epithelium related activation of complement differs in Crohn’s disease and ulcerative colitis. Gut 1992, 33, 902–908. [Google Scholar] [CrossRef]
- Lin, F.; Spencer, D.; Hatala, D.A.; Levine, A.D.; Medof, M.E. Decay-accelerating factor deficiency increases susceptibility to dextran sulfate sodium-induced colitis: Role for complement in inflammatory bowel disease. J. Immunol. 2004, 172, 3836–3841. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Vasek, M.J.; Garber, C.; Dorsey, D.; Durrant, D.M.; Bollman, B.; Soung, A.; Yu, J.; Perez-Torres, C.; Frouin, A.; Wilton, D.K.; et al. A complement-microglial axis drives synapse loss during virus-induced memory impairment. Nature 2016, 534, 538–543. [Google Scholar] [CrossRef] [Green Version]
- Park, J.W.; Kim, J.E.; Choi, Y.J.; Kang, M.J.; Choi, H.J.; Bae, S.J.; Hong, J.T.; Lee, H.; Hwang, D.Y. Deficiency of complement component 3 may be linked to the development of constipation in FVB/N-C3em1Hlee/Korl mice. FASEB J. 2021, 35, e21221. [Google Scholar] [CrossRef]
- Choi, Y.J.; Kim, J.E.; Lee, S.J.; Gong, J.E.; Son, H.J.; Hong, J.T.; Hwang, D.Y. Dysbiosis of fecal microbiota from complement 3 knockout mice with constipation phenotypes contributes to development of defecation delay. Front. Physiol. 2021, 12, 650789. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Kim, J.E.; Lee, S.J.; Gong, J.E.; Jin, Y.J.; Lee, H.; Hwang, D.Y. Promotion of the inflammatory response in mid colon of complement component 3 knockout mice. Sci. Rep. 2022, 12, 1700. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B. The Enteric Nervous System; Blackwell Science Ltd.: Oxford, UK, 2006; p. 271. [Google Scholar]
- Zeng, Q.; Ou, L.; Wang, W.; Guo, D.Y. Gastrin, cholecystokinin, signaling, and biological activities in cellular processes. Front. Endocrinol. 2020, 11, 112. [Google Scholar] [CrossRef] [PubMed]
- Fukudo, S.; Kuwano, H.; Miwa, H. Management and pathophysiology of functional gastrointestinal disorders. Digestion 2012, 85, 85–89. [Google Scholar] [CrossRef] [Green Version]
- Zhan, Y.; Wen, Y.; Zhang, L.L.; Shen, X.L.; Chen, X.H.; Wu, X.H.; Tang, X.G. Paeoniflorin improved constipation in the loperamide-induced rat model via TGR5/TRPA1 signaling-mediated 5-hydroxytryptamine secretion. Evid. Based Complement. Altern. Med. 2021, 2021, 6076293. [Google Scholar] [CrossRef]
- Al-Shboul, O.A. The importance of interstitial cells of Cajal in the gastrointestinal tract. Saudi. J. Gastroenterol. 2013, 19, 3–15. [Google Scholar] [CrossRef]
- Lee, J.I.; Park, H.; Kamm, M.A.; Talbot, I.C. Decreased density of interstitial cells of Cajal and neuronal cells in patients with slow-transit constipation and acquired megacolon. J. Gastroenterol. Hepatol. 2005, 20, 1292–1298. [Google Scholar] [CrossRef]
- Munson, L.G.; Scott, M.E.; Landay, A.L.; Spear, G.T. Decreased levels of complement receptor 1 (CD35) on B lymphocytes in persons with HIV infection. Clin. Immunol. Immunopathol. 1995, 75, 20–25. [Google Scholar] [CrossRef]
- Ueki, T.; Mizuno, M.; Uesu, T.; Kiso, T.; Nasu, J.; Inaba, T.; Kihara, Y.; Matsuoka, Y.; Okada, H.; Fujita, T.; et al. Distribution of activated complement, C3b, and its degraded fragments, iC3b/C3dg, in the colonic mucosa of ulcerative colitis (UC). Clin. Exp. Immunol. 1996, 104, 286–292. [Google Scholar] [CrossRef]
- Bassotti, G.; Villanacci, V.; Nascimbeni, R.; Asteria, C.R.; Fisogni, S.; Nesi, G.; Legrenzi, L.; Mariano, M.; Tonelli, F.; Morelli, A.; et al. Colonic neuropathological aspects in patients with intractable constipation due to obstructed defecation. Mod. Pathol. 2007, 20, 367–374. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.E.; Choi, Y.J.; Lee, S.J.; Gong, J.E.; Lim, Y.; Hong, J.T.; Hwang, D.Y. Molecular characterization of constipation disease as novel phenotypes in CRISPR-Cas9-generated leptin knockout mice with obesity. Int. J. Mol. Sci. 2020, 21, 9464. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Go, J.; Koh, E.K.; Song, S.H.; Sung, J.E.; Lee, H.A.; Lee, Y.H.; Hong, J.T.; Hwang, D.Y. Gallotannin-enriched extract isolated from galla rhois may be a functional candidate with laxative effects for treatment of loperamide-induced constipation of SD rats. PLoS ONE 2016, 11, e0161144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeung, A.K.; Lorenzo, C.D. Primary gastrointestinal motility disorders in childhood. Minerva. Pediatr. 2012, 64, 567–584. [Google Scholar]
- Atkinson, W.; Lockhart, S.; Whorwell, P.J.; Keevil, B.; Houghton, L.A. Altered 5-hydroxytryptamine signaling in patients with constipation- and diarrhea-predominant irritable bowel syndrome. Gastroenterology 2006, 130, 34–43. [Google Scholar] [CrossRef]
- Riordan, S.M.; McIver, C.J.; Thomas, M.C.; Wakefield, D.; Andreopoulos, P.C.; Duncombe, V.M.; Bolin, T.D. The expression of complement protein 4 and IgG3 in luminal secretions. Scand. J. Gastroenterol. 1996, 31, 1098–1102. [Google Scholar] [CrossRef] [PubMed]
- Schafer, D.P.; Lehrman, E.K.; Kautzman, A.G.; Koyama, R.; Mardinly, A.R.; Yamasaki, R.; Ransohoff, R.M.; Greenberg, M.E.; Barres, B.A.; Stevens, B. Microglia sculpt postnatal neural circuits in an activity and complement-dependent manner. Neuron 2012, 74, 691–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, R.; Li, Z.; Zhou, R.; Li, C.; Shao, S.; Li, J. The mechanism of intestinal flora dysregulation mediated by intestinal bacterial biofilm to induce constipation. Bioengineered 2021, 12, 6484–6498. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, L.; Bi, C.; Tang, K.; Zhang, B. Nostoc sphaeroides kütz polysaccharide improved constipation and promoted intestinal motility in rats. Evid. Based Complement. Altern. Med. 2021, 2021, 5596531. [Google Scholar] [CrossRef]
- Ricklin, D.; Reis, E.S.; Mastellos, D.C.; Gros, P.; Lambris, J.D. Complement component C3-The “Swiss Army Knife” of innate immunity and host defense. Immunol. Rev. 2016, 274, 33–58. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Suo, H.Y.; Qian, Y.; Li, G.J.; Liu, Z.H.; Li, J. Therapeutic effects of Lactobacillus casei Qian treatment in activated carbon-induced constipated mice. Mol. Med. Rep. 2015, 12, 3191–3199. [Google Scholar] [CrossRef] [Green Version]
- Miwa, J.; Echizen, H.; Matsueda, K.; Umeda, N. Patients with constipation-predominant irritable bowel syndrome (IBS) may have elevated serotonin concentrations in colonic mucosa as compared with diarrhea-predominant patients and subjects with normal bowel habits. Digestion 2001, 63, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Go, J.; Sung, J.E.; Lee, H.A.; Yun, W.B.; Hong, J.T.; Hwang, D.Y. Uridine stimulate laxative effect in the loperamide-induced constipation of SD rats through regulation of the mAChRs signaling pathway and mucin secretion. BMC Gastroenterol. 2017, 17, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.E.; Park, J.W.; Kang, M.J.; Choi, H.J.; Bae, S.J.; Choi, Y.; Lee, Y.J.; Seo, S.; Hong, J.T.; Hwang, D.Y. Laxative effect of spicatoside A by cholinergic regulation of enteric nerve in loperamide-induced constipation: ICR mice model. Molecules 2019, 24, 896. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Wang, Q.; Qian, Y.; Zhou, Y.; Wang, R.; Zhao, X. Component analysis of Pu-erh and its anti-constipation effects. Mol. Med. Rep. 2014, 9, 2003–2009. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, R.A.; Smits, G.J.; Timmermans, J.P. Study of NO and VIP as non-adrenergic non-cholinergic neurotransmitters in the pig gastric fundus. Br. J. Pharmacol. 1995, 116, 2017–2026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugihara, T.; Kobori, A.; Imaeda, H.; Tsujikawa, T.; Amagase, K.; Takeuchi, K.; Fujiyama, Y.; Andoh, A. The increased mucosal mRNA expressions of complement C3 and interleukin-17 in inflammatory bowel disease. Clin. Exp. Immunol. 2010, 160, 386–393. [Google Scholar] [CrossRef]
- Xia, Y.; Ross, G.D. Roles of integrins CR3 and CR4 in disease and therapeutic strategies. In Therapeutic Interventions in the Complement System; Humana Press: Totowa, NJ, USA, 2000; pp. 171–204. [Google Scholar]
- Huang, P.L.; Dawson, T.M.; Bredt, D.S.; Snyder, S.H.; Fishman, M.C. Targeted disruption of the neuronal nitric oxide synthase gene. Cell 1993, 75, 1273–1286. [Google Scholar] [CrossRef]
- Rawal, N.; Pangburn, M. Formation of high-affinity C5 convertases of the alternative pathway of complement. J. Immunol. 2001, 166, 2635–2642. [Google Scholar] [CrossRef] [Green Version]
- Tong, W.D.; Liu, B.H.; Zhang, L.Y.; Zhang, S.B.; Lei, Y. Decreased interstitial cells of Cajal in the sigmoid colon of patients with slow transit constipation. Int. J. Colorectal. Dis. 2004, 19, 467–473. [Google Scholar] [CrossRef]
- Fan, Y.H.; Xu, G.P.; Feng, W. Effects of zhizhu tongbian decoction on the colon ink propelling rate, GDNF, and NOS mRNA expression in rats with slow transit constipation. Chin. J. Integr. Med. 2012, 32, 486–489. [Google Scholar]
- Zhu, F.; Xu, S.; Zhang, Y.; Chen, F.; Ji, J.; Xie, G. Total glucosides of paeony promote intestinal motility in slow transit constipation rats through amelioration of interstitial cells of Cajal. PLoS ONE 2016, 11, e0160398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcorlo, M.; Martinez-Barricarte, R.; Fernandez, F.J.; Rodriguez-Gallego, C.; Round, A.; Vega, M.C.; Harris, C.L.; Cordoba, S.R.; Llorca, O. Unique structure of iC3b resolved at a resolution of 24 A by 3D-electron microscopy. Proc. Natl. Acad. Sci. USA 2011, 108, 13236–13240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kieslich, C.A.; Morikis, D. The two sides of complement C3d: Evolution of electrostatics in a link between innate and adaptive immunity. PLoS Comput. Biol. 2012, 8, e1002840. [Google Scholar] [CrossRef] [PubMed]
- Kremlitzka, M.; Mácsik-Valent, B.; Polgár, A.; Kiss, E.; Poór, G.; Erdei, A. Complement receptor type 1 suppresses human B cell functions in SLE patients. J. Immunol. Res. 2016, 2016, 5758192. [Google Scholar] [CrossRef] [Green Version]
- Moriyama, M.; Fukuhara, T.; Britschgi, M.; He, Y.; Narasimhan, R.; Villeda, S.; Molina, H.; Huber, B.T.; Holers, M.; Wyss-Coray, T. Complement receptor 2 is expressed in neural progenitor cells and regulates adult hippocampal neurogenesis. J. Neurosci. 2011, 31, 3981–3989. [Google Scholar] [CrossRef] [Green Version]
- Lyford, G.L.; He, C.L.; Soffer, E.; Hull, T.L.; Strong, S.A.; Senagore, A.J.; Burgart, L.J.; Young-Fadok, T.; Szurszewski, J.H.; Farrugia, G. Pan-colonic decrease in interstitial cells of Cajal in patients with slow transit constipation. Gut 2002, 51, 496–501. [Google Scholar] [CrossRef]
- Yin, J.; Liang, Y.; Wang, D.; Yan, Z.; Yin, H.; Wu, D.; Su, Q. Naringenin induces laxative effects by upregulating the expression levels of c-Kit and SCF, as well as those of aquaporin 3 in mice with loperamide-induced constipation. Int. J. Mol. Med. 2018, 41, 649–658. [Google Scholar] [CrossRef] [Green Version]
- Park, J.W.; Kim, J.E.; Kang, M.J.; Choi, H.J.; Bae, S.J.; Hwang, D.Y. Compensatory role of C3 convertase on the strain difference for C3 protein expression in FVB/N, C3H/HeN and C57BL/6N mice. Lab. Anim. Res. 2020, 36, 4. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.J.; Kim, J.E.; Lee, S.J.; Gong, J.E.; Jang, M.; Hong, J.T.; Hwang, D.Y. Loperamide-induced constipation activates inflammatory signaling pathways in the mid colon of SD rats via complement C3 and its receptors. Curr. Mol. Med. 2021, 21, 458–469. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, Y.J.; Song, H.J.; Kim, J.E.; Lee, S.J.; Jin, Y.J.; Roh, Y.J.; Seol, A.; Kim, H.S.; Hwang, D.Y. Dysregulation of the Enteric Nervous System in the Mid Colon of Complement Component 3 Knockout Mice with Constipation Phenotypes. Int. J. Mol. Sci. 2022, 23, 6862. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23126862
Choi YJ, Song HJ, Kim JE, Lee SJ, Jin YJ, Roh YJ, Seol A, Kim HS, Hwang DY. Dysregulation of the Enteric Nervous System in the Mid Colon of Complement Component 3 Knockout Mice with Constipation Phenotypes. International Journal of Molecular Sciences. 2022; 23(12):6862. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23126862
Chicago/Turabian StyleChoi, Yun Ju, Hee Jin Song, Ji Eun Kim, Su Jin Lee, You Jeong Jin, Yu Jeong Roh, Ayun Seol, Hye Sung Kim, and Dae Youn Hwang. 2022. "Dysregulation of the Enteric Nervous System in the Mid Colon of Complement Component 3 Knockout Mice with Constipation Phenotypes" International Journal of Molecular Sciences 23, no. 12: 6862. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23126862