
Mechanistic Effects of Baicalein on Aqueous Humor Drainage and Intraocular Pressure

 , , and
, , and
Abstract
:1. Introduction
2. Results
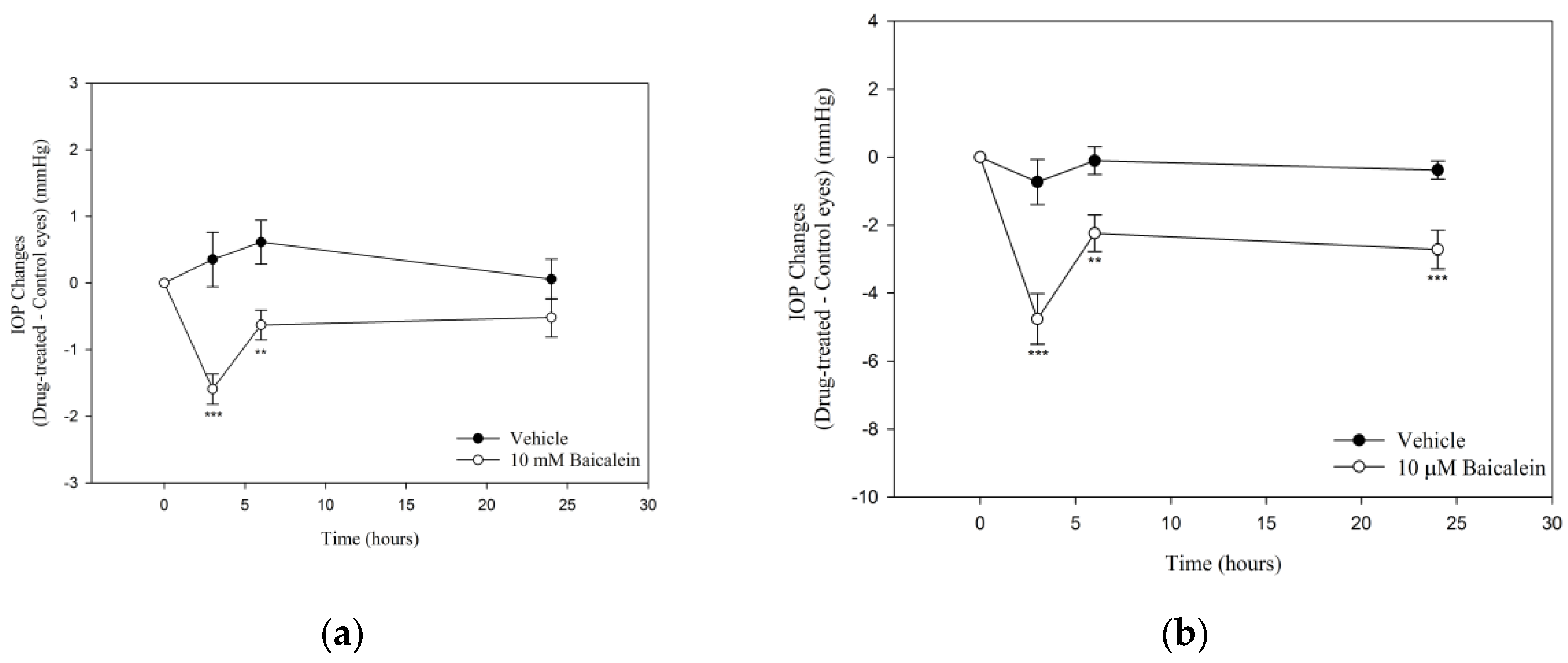
2.1. Ba Lowers IOP in Mouse Eyes
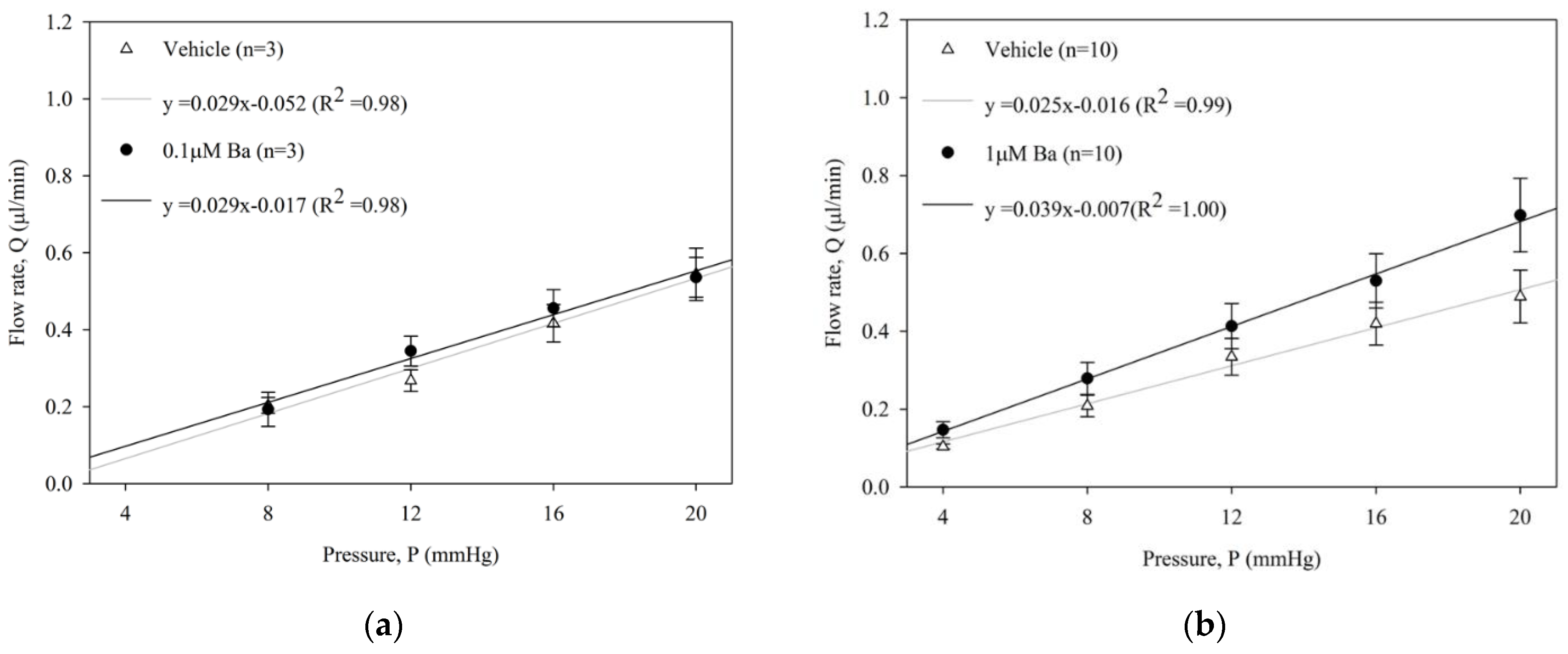
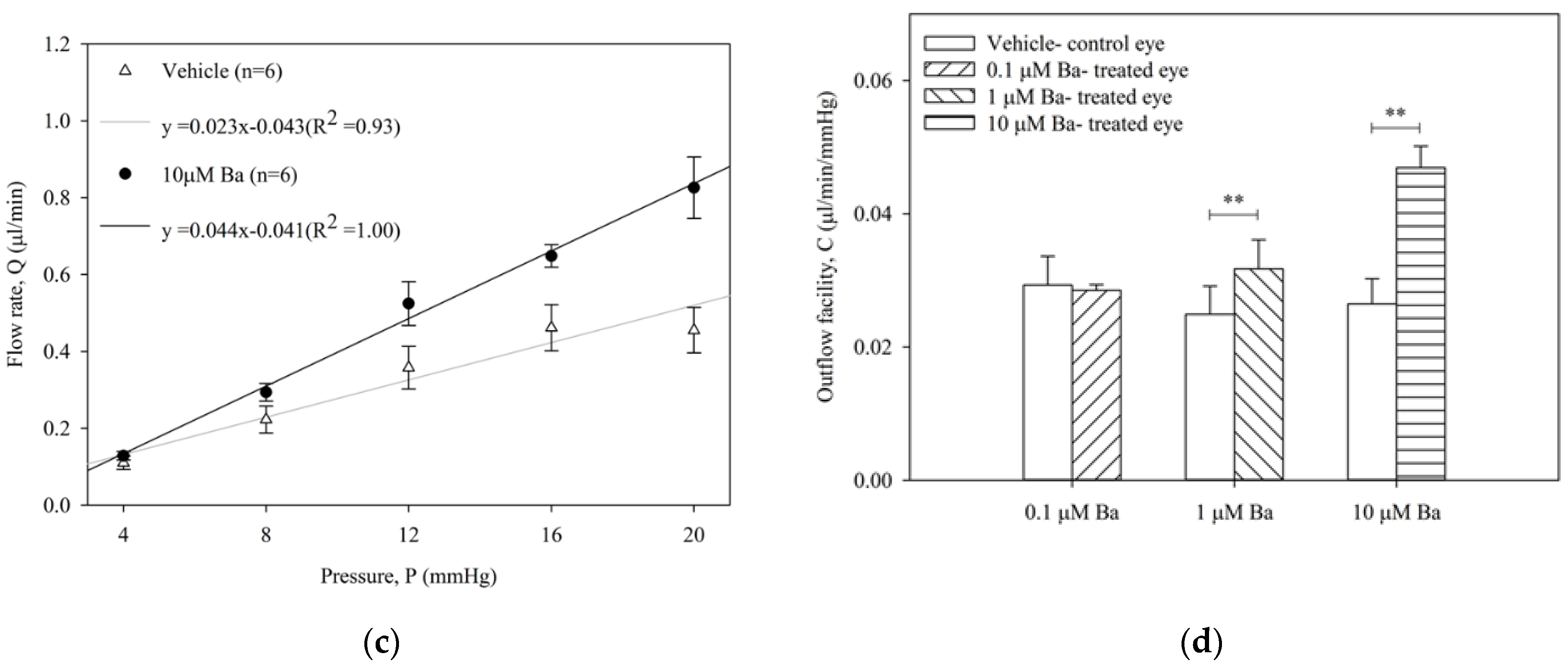
2.2. Ba Increases Conventional Outflow Facility in Ex Vivo Mouse Eyes
2.3. Mechanistic Studies of Ba in hTM Cells
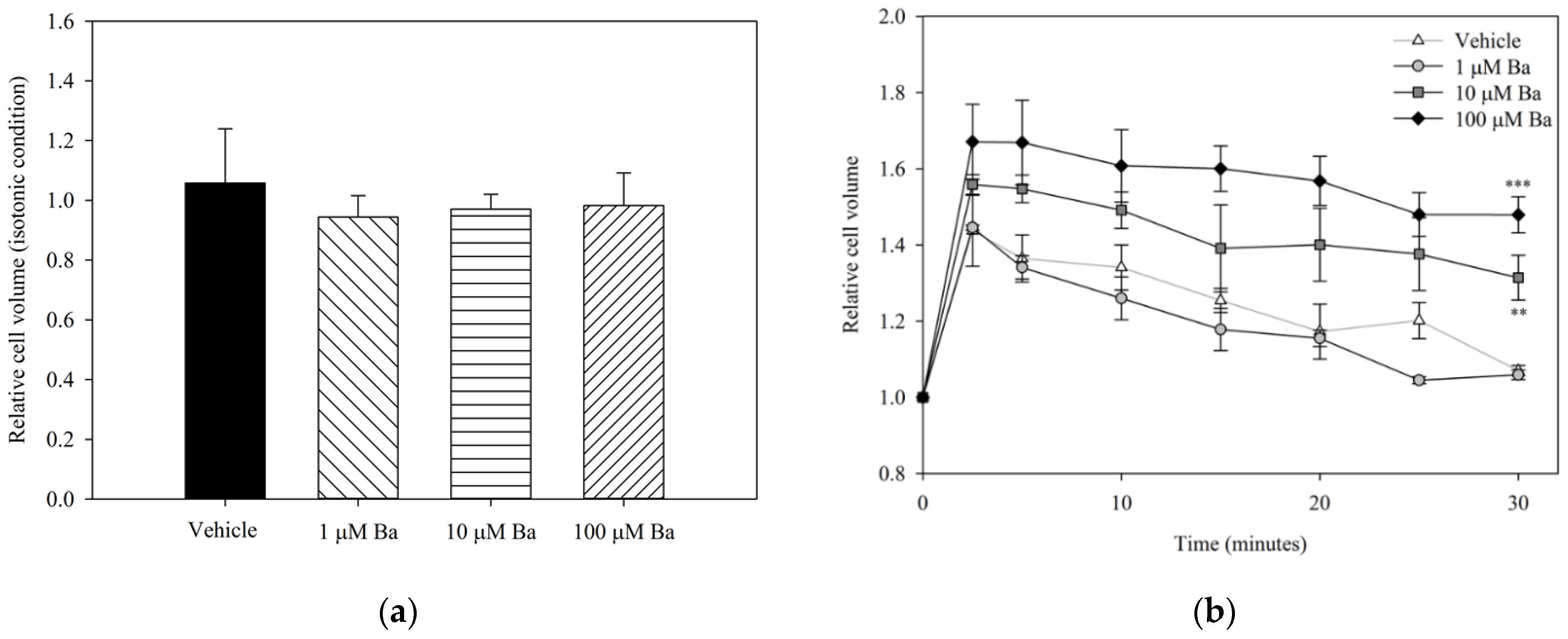
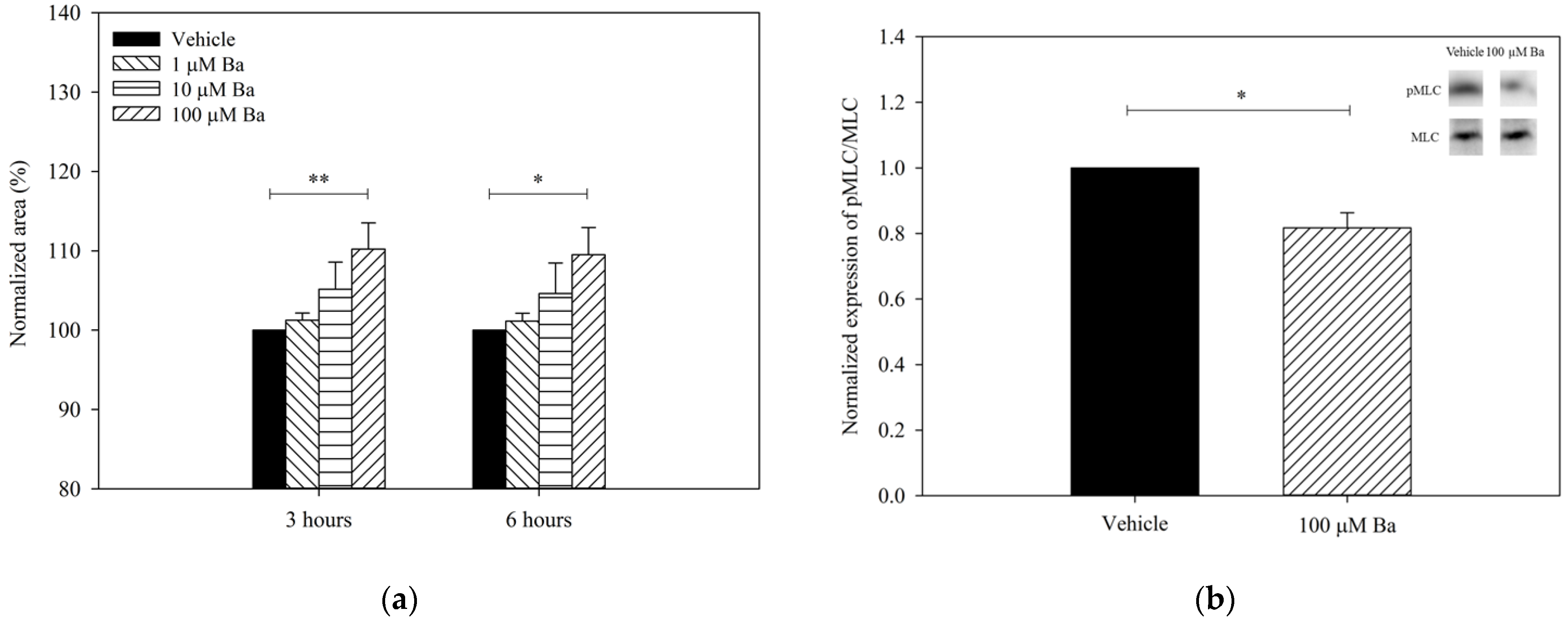
2.3.1. Ba Inhibits Cell Volume Regulation in hTM Cells
2.3.2. Ba Triggers hTM Cell Relaxation via Inhibition of MLC Phosphorylation
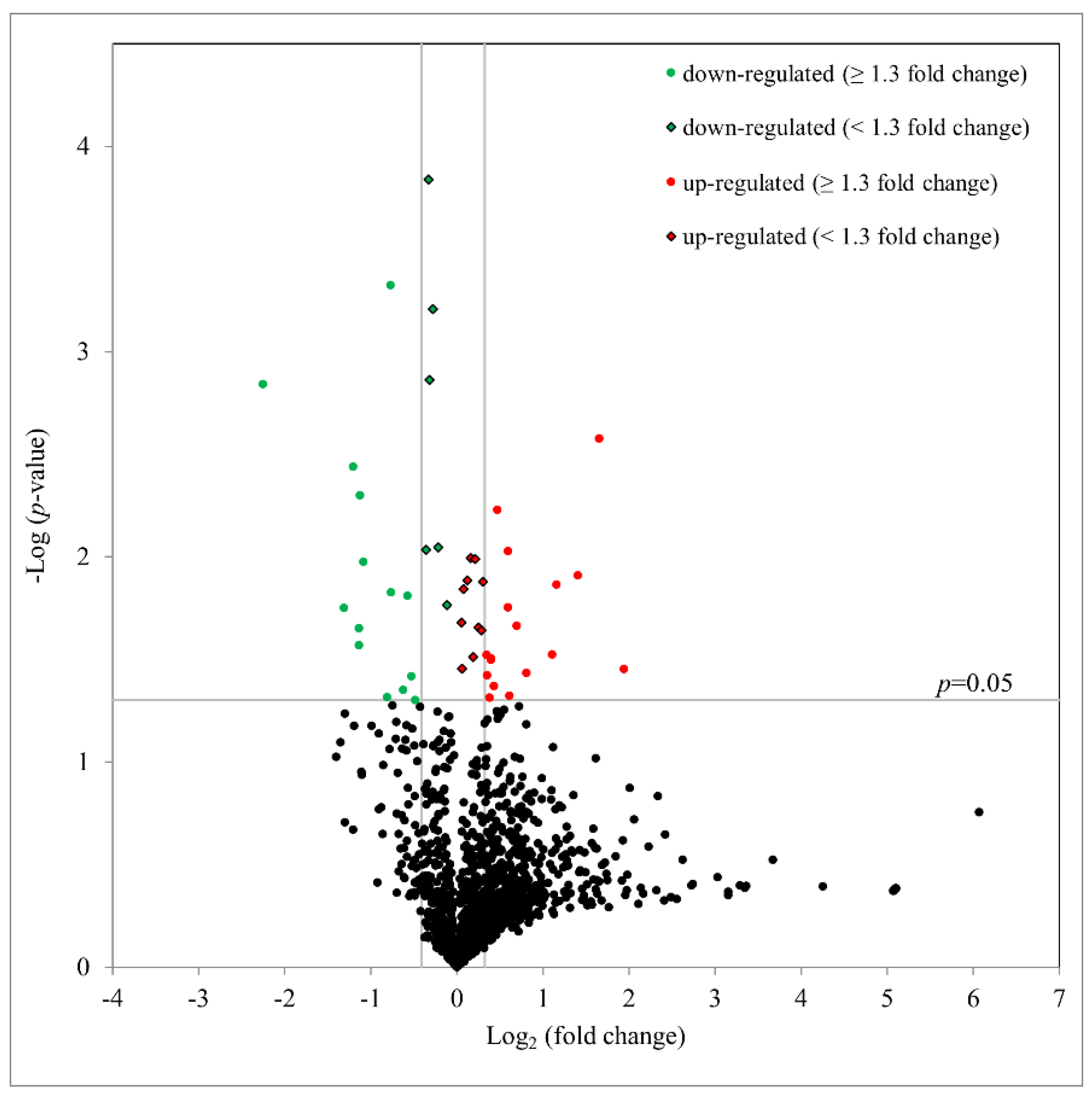
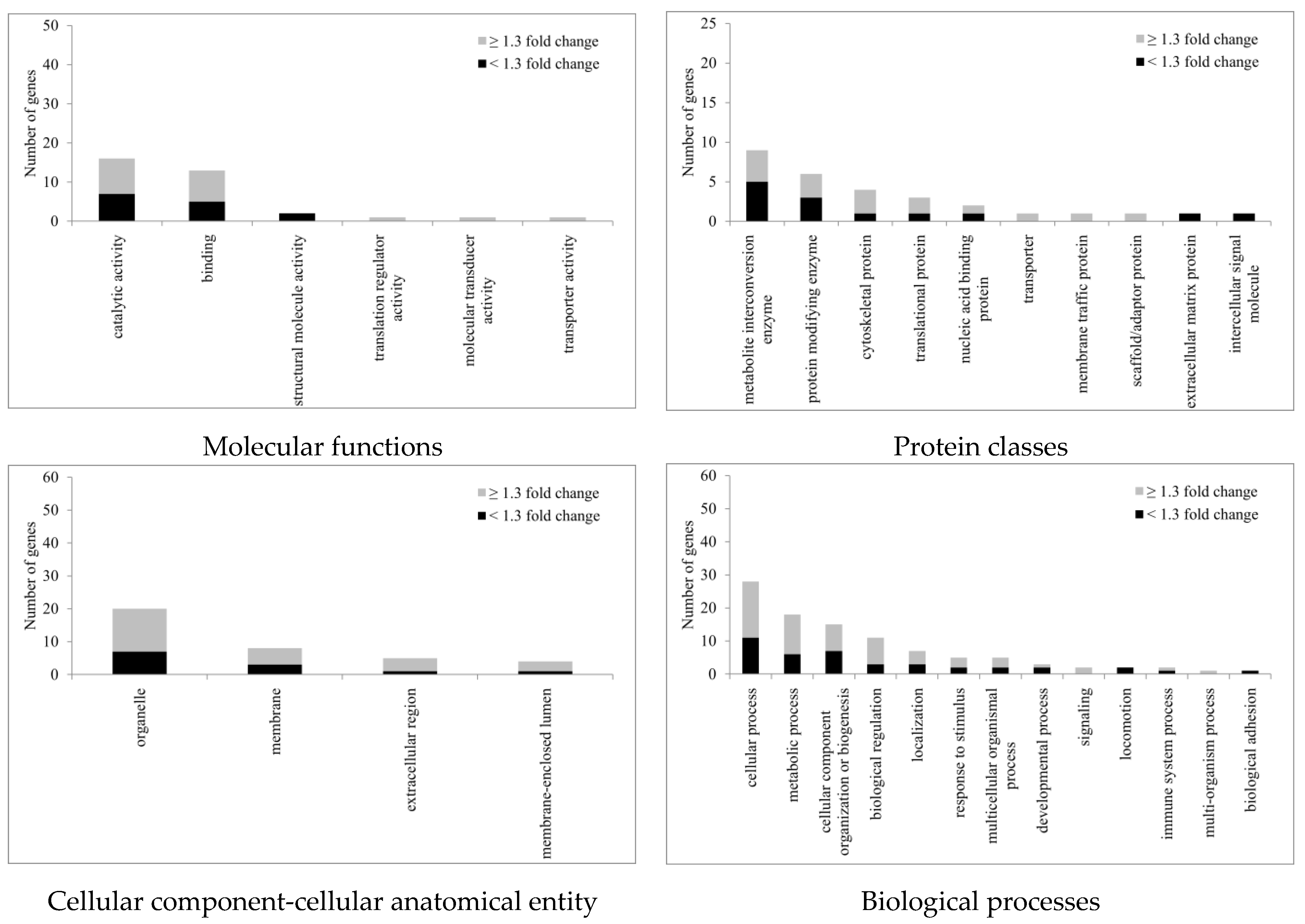
2.4. Proteomic Study of hTM Cells following Ba Treatment
3. Discussion
3.1. Ba Reduces IOP and Outflow Resistance through Modulating TM Cell Functions
3.2. Potential Protein Candidates Regulating Conventional Outflow Facility
3.3. Other Properties of Ba in Potentially Regulating TM Cell Functions
4. Materials and Methods
4.1. Non-Invasive IOP Measurements
4.2. Measurement of Outflow Facility in Mouse Eyes
4.3. Preparation of hTM Cells
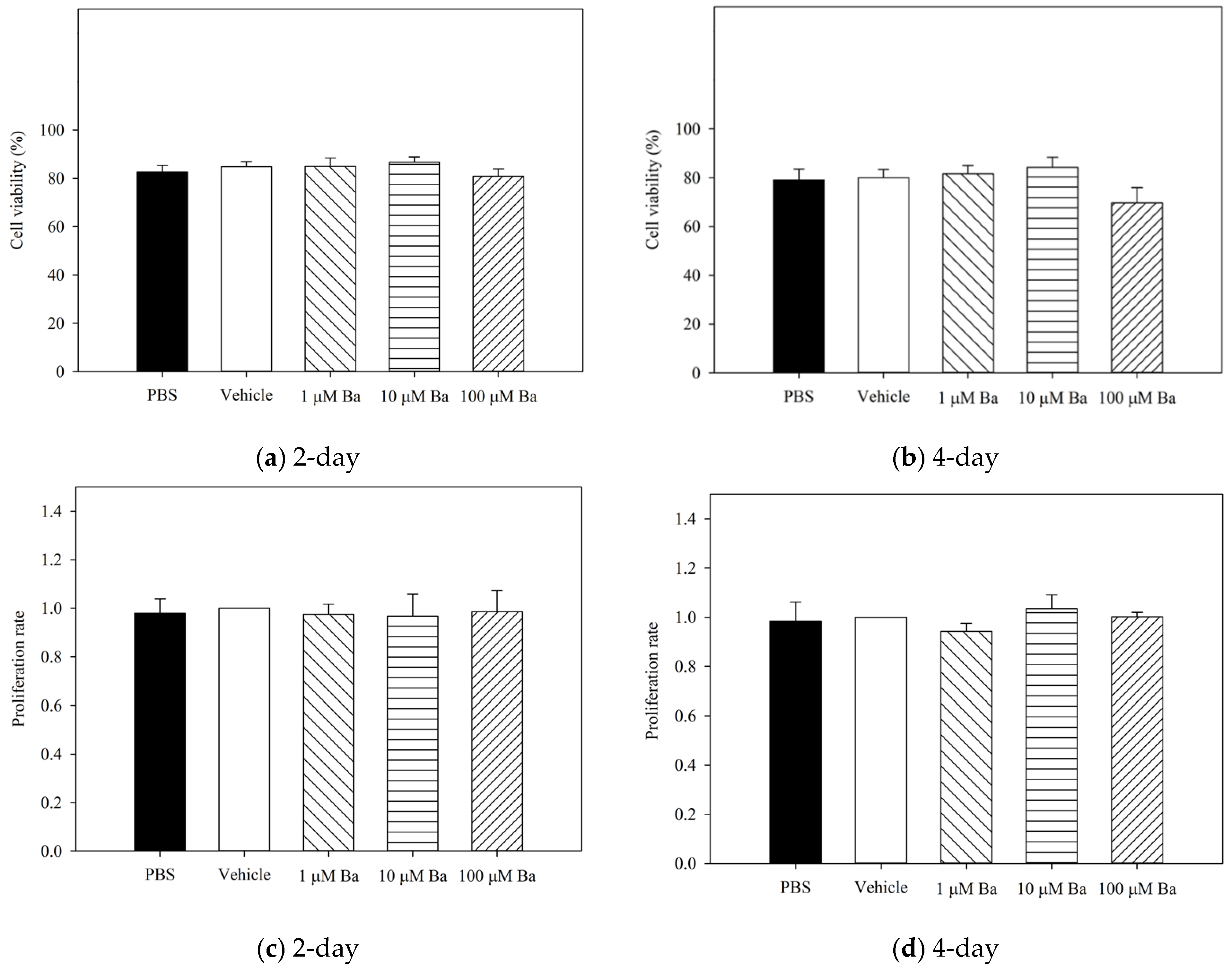
4.3.1. Measurement of Cell Viability and Proliferation
4.3.2. Measurements of Real-Time Cell Volume Changes
4.3.3. Measurement of Cell Contractility and Phosphorylated Myosin Light Chain (pMLC) Expression
4.4. Proteomics Study of Ba in hTM Cells
4.5. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Tham, Y.C.; Li, X.; Wong, T.Y.; Quigley, H.A.; Aung, T.; Cheng, C.Y. Global prevalence of glaucoma and projections of glaucoma burden through 2040: A systematic review and meta-analysis. Ophthalmology 2014, 121, 2081–2090. [Google Scholar] [CrossRef]
- To, C.H.; Kong, C.W.; Chan, C.Y.; Shahidullah, M.; Do, C.W. The mechanism of aqueous humour formation. Clin. Exp. Optom. 2002, 85, 335–349. [Google Scholar] [PubMed]
- The AGIS Investigators. The advanced glaucoma intervention study (AGIS): 7. The relationship between control of intraocular pressure and visual field deterioration. Am. J. Ophthalmol. 2000, 130, 429–440. [Google Scholar] [CrossRef]
- Dey, A.; Manthey, A.L.; Chiu, K.; Do, C.W. Methods to Induce Chronic Ocular Hypertension: Reliable Rodent Models as a Platform for Cell Transplantation and Other Therapies. Cell Transplant. 2018, 27, 213–229. [Google Scholar] [CrossRef] [Green Version]
- Pang, I.H.; Clark, A.F. Outflow signaling mechanisms and new therapeutic strategies for the control of intraocular pressure. In The Eye’s Aqeuous Humor, 2nd ed.; Civan, M.M., Ed.; Academic Press: San Diego, CA, USA, 2008; pp. 428–469. [Google Scholar]
- Overby, D.R.; Stamer, W.D.; Johnson, M. The changing paradigm of outflow resistance generation: Towards synergistic models of the JCT and inner wall endothelium. Exp. Eye Res. 2009, 88, 656–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamer, W.D. The cell and molecular biology of glaucoma: Mechanisms in the conventional outflow pathway. Investig. Ophthalmol. Vis. Sci. 2012, 53, 2470–2472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutjen-Drecoll, E.; Futa, R.; Rohen, J.W. Ultrahistochemical studies on tangential sections of the trabecular meshwork in normal and glaucomatous eyes. Invest. Ophthalmol. Vis. Sci. 1981, 21, 563–573. [Google Scholar]
- Ueda, J.; Yue, B.Y. Distribution of myocilin and extracellular matrix components in the corneoscleral meshwork of human eyes. Invest. Ophthalmol. Vis. Sci 2003, 44, 4772–4779. [Google Scholar] [CrossRef] [Green Version]
- Gabelt, B.T.; Kaufman, P.L. Changes in aqueous humor dynamics with age and glaucoma. Prog. Retin. Eye Res. 2005, 24, 612–637. [Google Scholar] [CrossRef]
- Dinda, B.; Dinda, S.; DasSharma, S.; Banik, R.; Chakraborty, A.; Dinda, M. Therapeutic potentials of baicalin and its aglycone, baicalein against inflammatory disorders. Eur. J. Med. Chem. 2017, 131, 68–80. [Google Scholar] [CrossRef]
- Li-Weber, M. New therapeutic aspects of flavones: The anticancer properties of Scutellaria and its main active constituents Wogonin, Baicalein and Baicalin. Cancer Treat. Rev. 2009, 35, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Tsang, S.Y.; Yao, X.; Chen, Z.Y. Biological properties of baicalein in cardiovascular system. Curr. Drug Targets Cardiovasc. Haematol. Disord. 2005, 5, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Cho, K.S.; Yi, I.; To, C.H.; Chen, D.F.; Do, C.W. Baicalein, Baicalin, and Wogonin: Protective Effects against Ischemia-Induced Neurodegeneration in the Brain and Retina. Oxid. Med. Cell Longev. 2021, 2021, 8377362. [Google Scholar] [CrossRef]
- Xiao, J.-R.; Do, C.-W.; To, C.-H. Potential Therapeutic Effects of Baicalein, Baicalin, and Wogonin in Ocular Disorders. Ocul. Pharmacol. Ther. 2014, 30, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.J.; Huang, T.S.; Cheng, W.F.; Lu, F.J. Baicalein and baicalin are potent inhibitors of angiogenesis: Inhibition of endothelial cell proliferation, migration and differentiation. Int. J. Cancer 2003, 106, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Ko, W.H.; Law, V.W.; Yip, W.C.; Yue, G.G.; Lau, C.W.; Chen, Z.Y.; Huang, Y. Stimulation of chloride secretion by baicalein in isolated rat distal colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 282, G508–G518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dismuke, W.M.; Liang, J.; Overby, D.R.; Stamer, W.D. Concentration-related effects of nitric oxide and endothelin-1 on human trabecular meshwork cell contractility. Exp. Eye Res. 2014, 120, 28–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenthal, R.; Choritz, L.; Schlott, S.; Bechrakis, N.E.; Jaroszewski, J.; Wiederholt, M.; Thieme, H. Effects of ML-7 and Y-27632 on carbachol- and endothelin-1-induced contraction of bovine trabecular meshwork. Exp. Eye Res. 2005, 80, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.V.; Deng, P.; Sasaki, Y.; Epstein, D.L. Regulation of myosin light chain phosphorylation in the trabecular meshwork: Role in aqueous humour outflow facility. Exp. Eye Res. 2005, 80, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Ho, L.C.; Conner, I.P.; Do, C.W.; Kim, S.G.; Wu, E.X.; Wollstein, G.; Schuman, J.S.; Chan, K.C. In vivo assessment of aqueous humor dynamics upon chronic ocular hypertension and hypotensive drug treatment using gadolinium-enhanced MRI. Investig. Ophthalmol. Vis. Sci. 2014, 55, 3747–3757. [Google Scholar] [CrossRef] [Green Version]
- Vranka, J.A.; Kelley, M.J.; Acott, T.S.; Keller, K.E. Extracellular matrix in the trabecular meshwork: Intraocular pressure regulation and dysregulation in glaucoma. Exp. Eye Res. 2015, 133, 112–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tektas, O.Y.; Lütjen-Drecoll, E. Structural changes of the trabecular meshwork in different kinds of glaucoma. Exp. Eye Res. 2009, 88, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Gasiorowski, K.; Lamer-Zarawska, E.; Leszek, J.; Parvathaneni, K.; Yendluri, B.B.; Blach-Olszewska, Z.; Aliev, G. Flavones from root of Scutellaria baicalensis Georgi: Drugs of the future in neurodegeneration? CNS Neurol. Disord. Drug Targets 2011, 10, 184–191. [Google Scholar] [CrossRef]
- Oh, K.S.; Oh, B.K.; Park, C.H.; Mun, J.; Won, S.H.; Lee, B.H. Baicalein potently inhibits Rho kinase activity and suppresses actin stress fiber formation in angiotensin II-stimulated H9c2 cells. Biol. Pharm. Bull. 2012, 35, 1281–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.H.; Kim, C.; Zhang, L.; Lee, Y.J. Role of p53, PUMA, and Bax in wogonin-induced apoptosis in human cancer cells. Biochem. Pharmacol. 2008, 75, 2020–2033. [Google Scholar] [CrossRef] [Green Version]
- Maher, P.; Hanneken, A. Flavonoids protect retinal ganglion cells from oxidative stress-induced death. Investig. Ophthalmol. Vis. Sci. 2005, 46, 4796–4803. [Google Scholar] [CrossRef] [PubMed]
- Chao, H.M.; Chuang, M.J.; Liu, J.H.; Liu, X.Q.; Ho, L.K.; Pan, W.H.; Zhang, X.M.; Liu, C.M.; Tsai, S.K.; Kong, C.W. Baicalein protects against retinal ischemia by antioxidation, antiapoptosis, downregulation of HIF-1α, VEGF, and MMP-9 and upregulation of HO-1. J. Ocul. Pharmacol. Ther. 2013, 29, 539–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanneken, A.; Lin, F.F.; Johnson, J.; Maher, P. Flavonoids protect human retinal pigment epithelial cells from oxidative-stress-induced death. Investig. Ophthalmol. Vis. Sci. 2006, 47, 3164–3177. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Su, Y.L.; Lau, C.W.; Law, W.I.; Huang, Y. Endothelium-dependent contraction and direct relaxation induced by baicalein in rat mesenteric artery. Eur. J. Pharmacol. 1999, 374, 41–47. [Google Scholar] [CrossRef]
- Huang, Y.; Wong, C.M.; Lau, C.-W.; Yao, X.; Tsang, S.Y.; Su, Y.L.; Chen, Z.Y. Inhibition of nitric oxide/cyclic GMP-mediated relaxation by purified flavonoids, baicalin and baicalein, in rat aortic rings. Biochem. Pharmacol. 2004, 67, 787–794. [Google Scholar] [CrossRef]
- Xue, Y.; Li, H.; Zhang, Y.; Han, X.; Zhang, G.; Li, W.; Zhang, H.; Lin, Y.; Chen, P.; Sun, X. Natural and synthetic flavonoids, novel blockers of the volume-regulated anion channels, inhibit endothelial cell proliferation. Pflügers Arch. Eur. J. Physiol. 2018, 470, 1473–1483. [Google Scholar] [CrossRef]
- Sun, H.; Che, Q.M.; Zhao, X.; Pu, X.P. Antifibrotic effects of chronic baicalein administration in a CCl4 liver fibrosis model in rats. Eur. J. Pharmacol. 2010, 631, 53–60. [Google Scholar] [CrossRef]
- Gao, Y.; Lu, J.; Zhang, Y.; Chen, Y.; Gu, Z.; Jiang, X. Baicalein attenuates bleomycin-induced pulmonary fibrosis in rats through inhibition of miR-21. Pulm. Pharmacol. Ther. 2013, 26, 649–654. [Google Scholar] [CrossRef]
- Shan, S.W.; Do, C.W.; Lam, T.C.; Li, H.L.; Stamer, W.D.; To, C.H. Thrombospondin-1 mediates Rho-kinase inhibitor-induced increase in outflow-facility. J. Cell. Physiol. 2021, 236, 8226–8238. [Google Scholar] [CrossRef]
- Rao, P.V.; Deng, P.-F.; Kumar, J.; Epstein, D.L. Modulation of Aqueous Humor Outflow Facility by the Rho Kinase-Specific Inhibitor Y-27632. Investig. Ophthalmol. Vis. Sci. 2001, 42, 1029–1037. [Google Scholar]
- Lu, Z.; Overby, D.R.; Scott, P.A.; Freddo, T.F.; Gong, H. The mechanism of increasing outflow facility by rho-kinase inhibition with Y-27632 in bovine eyes. Exp. Eye Res. 2008, 86, 271–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gual, A.; Llobet, A.; Gilabert, R.; Borras, M.; Pales, J.; Bergamini, M.; Belmonte, C. Effects of time of storage, albumin, and osmolality changes on outflow facility (C) of bovine anterior segment in vitro. Investig. Ophthalmol. Vis. Sci. 1997, 38, 2165–2171. [Google Scholar]
- Al-Aswad, L.A.; Gong, H.; Lee, D.; O’Donnell, M.E.; Brandt, J.D.; Ryan, W.J.; Schroeder, A.; Erickson, K.A. Effects of Na-K-2Cl cotransport regulators on outflow facility in calf and human eyes in Vitro. Investig. Ophthalmol. Vis. Sci. 1999, 40, 1695–1701. [Google Scholar]
- McLaughlin, C.W.; Karl, M.O.; Zellhuber-McMillan, S.; Wang, Z.; Do, C.W.; Leung, C.T.; Li, A.; Stone, R.A.; Macknight, A.D.; Civan, M.M. Electron probe X-ray microanalysis of intact pathway for human aqueous humor outflow. Am. J. Physiol. Cell Physiol. 2008, 295, C1083–C1091. [Google Scholar] [CrossRef]
- Dismuke, W.M.; Ellis, D.Z. Activation of the BKCa channel increases outflow facility and decreases trabecular meshwork cell volume. J. Ocul. Pharmacol. Ther. 2009, 25, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Comes, N.; Abad, E.; Morales, M.; Borrás, T.; Gual, A.; Gasull, X. Identification and functional characterization of ClC-2 chloride channels in trabecular meshwork cells. Exp. Eye Res. 2006, 83, 877–889. [Google Scholar] [CrossRef]
- O’Donnell, M.E.; Brandt, J.D.; Curry, F. Na-K-Cl cotransport regulates intracellular volume and monolayer permeability of trabecular meshwork cells. Am. J. Physiol. Cell Physiol. 1995, 268, C1067–C1074. [Google Scholar] [CrossRef]
- Mitchell, C.H.; Fleischhauer, J.C.; Stamer, W.D.; Peterson-Yantorno, K.; Civan, M.M. Human trabecular meshwork cell volume regulation. Am. J. Physiol. Cell Physiol. 2002, 283, C315–C326. [Google Scholar] [CrossRef] [Green Version]
- Gasull, X.; Ferrer, E.; Llobet, A.; Castellano, A.; Nicolás, J.M.; Palés, J.; Gual, A. Cell Membrane Stretch Modulates the High-Conductance Ca2+-Activated K+ Channel in Bovine Trabecular Meshwork Cells. Investig. Ophthalmol. Vis. Sci. 2003, 44, 706–714. [Google Scholar] [CrossRef] [Green Version]
- Carreon, T.A.; Castellanos, A.; Gasull, X.; Bhattacharya, S.K. Interaction of cochlin and mechanosensitive channel TREK-1 in trabecular meshwork cells influences the regulation of intraocular pressure. Sci. Rep. 2017, 7, 452. [Google Scholar] [CrossRef] [Green Version]
- Soto, D.; Comes, N.; Ferrer, E.; Morales, M.; Escalada, A.; Palés, J.; Solsona, C.; Gual, A.; Gasull, X. Modulation of aqueous humor outflow by ionic mechanisms involved in trabecular meshwork cell volume regulation. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3650–3661. [Google Scholar] [CrossRef] [Green Version]
- Dismuke, W.M.; Mbadugha, C.C.; Ellis, D.Z. NO-induced regulation of human trabecular meshwork cell volume and aqueous humor outflow facility involve the BKCa ion channel. Am. J. Physiol. Cell Physiol. 2008, 294, C1378–C1386. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, J.; Leung, C.T.; Li, A.; Peterson-Yantorno, K.; Ouyang, H.; Stamer, W.D.; Civan, M.M. Regulatory Roles of Anoctamin-6 in Human Trabecular Meshwork Cells. Invest. Ophthalmol. Vis. Sci. 2017, 58, 492–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleischhauer, J.C.; Mitchell, C.H.; Stamer, W.D.; Karl, M.O.; Peterson-Yantorno, K.; Civan, M.M. Common actions of adenosine receptor agonists in modulating human trabecular meshwork cell transport. J. Membr. Biol. 2003, 193, 121–136. [Google Scholar] [CrossRef]
- Li, A.; Leung, C.T.; Peterson-Yantorno, K.; Stamer, W.D.; Mitchell, C.H.; Civan, M.M. Mechanisms of ATP release by human trabecular meshwork cells, the enabling step in purinergic regulation of aqueous humor outflow. J. Cell. Physiol. 2012, 227, 172–182. [Google Scholar] [CrossRef] [Green Version]
- Li, A.; Leung, C.T.; Peterson-Yantorno, K.; Stamer, W.D.; Civan, M.M. Cytoskeletal dependence of adenosine triphosphate release by human trabecular meshwork cells. Investig. Ophthalmol. Vis. Sci. 2011, 52, 7996–8005. [Google Scholar] [CrossRef] [PubMed]
- Crosson, C.E.; Sloan, C.F.; Yates, P.W. Modulation of conventional outflow facility by the adenosine A1 agonist N6-cyclohexyladenosine. Investig. Ophthalmol. Vis. Sci. 2005, 46, 3795–3799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crosson, C.E. Intraocular pressure responses to the adenosine agonist cyclohexyladenosine: Evidence for a dual mechanism of action. Investig. Ophthalmol. Vis. Sci. 2001, 42, 1837–1840. [Google Scholar]
- Koga, T.; Koga, T.; Awai, M.; Tsutsui, J.I.; Yue, B.Y.J.T.; Tanihara, H. Rho-associated protein kinase inhibitor, Y-27632, induces alterations in adhesion, contraction and motility in cultured human trabecular meshwork cells. Exp. Eye Res. 2006, 82, 362–370. [Google Scholar] [CrossRef]
- Ramachandran, C.; Patil, R.V.; Combrink, K.; Sharif, N.A.; Srinivas, S.P. Rho-Rho kinase pathway in the actomyosin contraction and cell-matrix adhesion in immortalized human trabecular meshwork cells. Mol. Vis. 2011, 17, 1877–1890. [Google Scholar]
- Obermajer, N.; Jevnikar, Z.; Doljak, B.; Kos, J. Role of cysteine cathepsins in matrix degradation and cell signalling. Connect. Tissue Res. 2008, 49, 193–196. [Google Scholar] [CrossRef]
- Cavallo Medved, D.; Rudy, D.; Blum, G.; Bogyo, M.; Caglic, D.; Sloane, B.F. Live-cell imaging demonstrates extracellular matrix degradation in association with active cathepsin B in caveolae of endothelial cells during tube formation. Exp. Cell Res. 2009, 315, 1234–1246. [Google Scholar] [CrossRef] [Green Version]
- Porter, K.; Hirt, J.; Stamer, W.D.; Liton, P.B. Autophagic dysregulation in glaucomatous trabecular meshwork cells. Biochim. Biophys. Acta Mol. Basis Dis. 2015, 1852, 379–385. [Google Scholar] [CrossRef] [Green Version]
- Nettesheim, A.; Shim, M.S.; Dixon, A.; Raychaudhuri, U.; Gong, H.; Liton, P.B. Cathepsin B Localizes in the Caveolae and Participates in the Proteolytic Cascade in Trabecular Meshwork Cells. Potential New Drug Target for the Treatment of Glaucoma. J. Clin. Med. 2021, 10, 78. [Google Scholar] [CrossRef]
- Sonesson, B.; Rosengren, E.; Hansson, A.S.; Hansson, C. UVB-induced inflammation gives increased d-dopachrome tautomerase activity in blister fluid which correlates with macrophage migration inhibitory factor. Exp. Dermatol. 2003, 12, 278–282. [Google Scholar] [CrossRef] [Green Version]
- Manavathi, B.; Lo, D.; Bugide, S.; Dey, O.; Imren, S.; Weiss, M.J.; Humphries, R.K. Functional regulation of pre-B-cell leukemia homeobox interacting protein 1 (PBXIP1/HPIP) in erythroid differentiation. J. Biol. Chem. 2012, 287, 5600–5614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abramovich, C.; Chavez, E.A.; Lansdorp, P.M.; Humphries, R.K. Functional characterization of multiple domains involved in the subcellular localization of the hematopoietic Pbx interacting protein (HPIP). Oncogene 2002, 21, 6766–6771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Li, L.; Zhang, X.; Zhang, Y.; Liang, Y.; Lv, J.; Fan, Z.; Guo, J.; Hong, T.; Ji, B. Hematopoietic pre-B cell leukemia transcription factor interacting protein is overexpressed in gastric cancer and promotes gastric cancer cell proliferation, migration, and invasion. Cancer Sci. 2015, 106, 1313–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.C.; Chai, D.S.; Chen, C.B.; Wang, Z.Y.; Wang, L. HPIP promotes thyroid cancer cell growth, migration and EMT through activating PI3K/AKT signaling pathway. Biomed. Pharmacother. 2015, 75, 33–39. [Google Scholar] [CrossRef]
- Pan, J.; Qin, Y.; Zhang, M. HPIP promotes non-small cell lung cancer cell proliferation, migration and invasion through regulation of the Sonic hedgehog signaling pathway. Biomed. Pharmacother. 2016, 77, 176–181. [Google Scholar] [CrossRef]
- Machesky, L.M.; Gould, K.L. The Arp2/3 complex: A multifunctional actin organizer. Curr. Opin. Cell Biol. 1999, 11, 117–121. [Google Scholar] [CrossRef]
- Kinoshita, T.; Nohata, N.; Watanabe-Takano, H.; Yoshino, H.; Hidaka, H.; Fujimura, L.; Fuse, M.; Yamasaki, T.; Enokida, H.; Nakagawa, M. Actin-related protein 2/3 complex subunit 5 (ARPC5) contributes to cell migration and invasion and is directly regulated by tumor-suppressive microRNA-133a in head and neck squamous cell carcinoma. Int. J. Oncol. 2012, 40, 1770–1778. [Google Scholar]
- De Groef, L.; Van Hove, I.; Dekeyster, E.; Stalmans, I.; Moons, L. MMPs in the Trabecular Meshwork: Promising Targets for Future Glaucoma Therapies? Investig. Ophthalmol. Vis. Sci. 2013, 54, 7756–7763. [Google Scholar] [CrossRef] [Green Version]
- Bradley, J.M.B.; Kelley, M.J.; Rose, A.; Acott, T.S. Signaling Pathways Used in Trabecular Matrix Metalloproteinase Response to Mechanical Stretch. Investig. Ophthalmol. Vis. Sci. 2003, 44, 5174–5181. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.F.; Sun, Y.Y.; Acott, T.S.; Keller, K.E. Effects of induction and inhibition of matrix cross-linking on remodeling of the aqueous outflow resistance by ocular trabecular meshwork cells. Sci. Rep. 2016, 6, 30505. [Google Scholar] [CrossRef]
- Li, G.; Torrejon, K.Y.; Unser, A.M.; Ahmed, F.; Navarro, I.D.; Baumgartner, R.A.; Albers, D.S.; Stamer, W.D. Trabodenoson, an Adenosine Mimetic with A1 Receptor Selectivity Lowers Intraocular Pressure by Increasing Conventional Outflow Facility in Mice. Invest. Ophthalmol. Vis. Sci 2018, 59, 383–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimoto, T.; Inoue, T.; Maki, K.; Inoue-Mochita, M.; Tanihara, H. Vascular Endothelial Growth Factor-A Increases the Aqueous Humor Outflow Facility. PLoS ONE 2016, 11, e0161332. [Google Scholar] [CrossRef] [PubMed]
- Saccà, S.C.; Pascotto, A.; Camicione, P.; Capris, P.; Izzotti, A. Oxidative DNA damage in the human trabecular meshwork: Clinical correlation in patients with primary open-angle glaucoma. Arch. Ophthalmol. 2005, 123, 458–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghanem, A.A.; Arafa, L.F.; El-Baz, A. Oxidative stress markers in patients with primary open-angle glaucoma. Curr. Eye Res. 2010, 35, 295–301. [Google Scholar] [CrossRef]
- Wax, M.B.; Tezel, G. Immunoregulation of retinal ganglion cell fate in glaucoma. Exp. Eye Res. 2009, 88, 825–830. [Google Scholar] [CrossRef]
- Martin, E.; Rosenthal, R.E.; Fiskum, G. Pyruvate dehydrogenase complex: Metabolic link to ischemic brain injury and target of oxidative stress. J. Neurosci. Res. 2005, 79, 240–247. [Google Scholar] [CrossRef] [Green Version]
- Balakrishnan, A.; Nemeria, N.S.; Chakraborty, S.; Kakalis, L.; Jordan, F. Determination of pre-steady-state rate constants on the Escherichia coli pyruvate dehydrogenase complex reveals that loop movement controls the rate-limiting step. J. Am. Chem. Soc. 2012, 134, 18644–18655. [Google Scholar] [CrossRef] [Green Version]
- Aslan, M.; Cort, A.; Yucel, I. Oxidative and nitrative stress markers in glaucoma. Free Radic. Biol. Med. 2008, 45, 367–376. [Google Scholar] [CrossRef]
- He, Y.; Leung, K.W.; Zhang, Y.H.; Duan, S.; Zhong, X.F.; Jiang, R.Z.; Peng, Z.; Tombran-Tink, J.; Ge, J. Mitochondrial complex I defect induces ROS release and degeneration in trabecular meshwork cells of POAG patients: Protection by antioxidants. Investig. Ophthalmol. Vis. Sci. 2008, 49, 1447–1458. [Google Scholar] [CrossRef]
- Zhang, S.; Ye, J.; Dong, G. Neuroprotective effect of baicalein on hydrogen peroxide-mediated oxidative stress and mitochondrial dysfunction in PC12 cells. J. Mol. Neurosci. 2010, 40, 311–320. [Google Scholar] [CrossRef]
- Wang, Y.H.; Yu, H.T.; Pu, X.P.; Du, G.H. Baicalein prevents 6-hydroxydopamine-induced mitochondrial dysfunction in SH-SY5Y cells via inhibition of mitochondrial oxidation and up-regulation of DJ-1 protein expression. Molecules 2013, 18, 14726–14738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarado, J.; Murphy, C.; Juster, R. Trabecular meshwork mellularity in primary open-angle glaucoma and nonglaucomatous normals. Ophthalmology 1984, 91, 564–579. [Google Scholar] [CrossRef]
- Sacca, S.C.; Gandolfi, S.; Bagnis, A.; Manni, G.; Damonte, G.; Traverso, C.E.; Izzotti, A. From DNA damage to functional changes of the trabecular meshwork in aging and glaucoma. Ageing Res. Rev. 2016, 29, 26–41. [Google Scholar] [CrossRef]
- Anders, F.; Mann, C.; Liu, A.; Teister, J.; Funke, S.; Thanos, S.; Grus, F.; Pfeiffer, N.; Prokosch, V. Correlation of Crystallin Expression and RGC Susceptibility in Experimental Glaucoma Rats of Different Ages. Curr. Eye Res. 2018, 43, 1267–1273. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Sze, Y.H.; Yang, M.; Tang, J.; Zhao, S.; Yi, I.; To, C.H.; Lam, C.; Chen, D.F.; Cho, K.S.; et al. Baicalein-A Potent Pro-Homeostatic Regulator of Microglia in Retinal Ischemic Injury. Front. Immunol. 2022, 13, 837497. [Google Scholar] [CrossRef]
- Taurone, S.; Ripandelli, G.; Pacella, E.; Bianchi, E.; Plateroti, A.M.; De Vito, S.; Plateroti, P.; Grippaudo, F.R.; Cavallotti, C.; Artico, M. Potential regulatory molecules in the human trabecular meshwork of patients with glaucoma: Immunohistochemical profile of a number of inflammatory cytokines. Mol. Med. Rep. 2015, 11, 1384–1390. [Google Scholar] [CrossRef] [Green Version]
- Sumida, G.M.; Stamer, W.D. Sphingosine-1-phosphate enhancement of cortical actomyosin organization in cultured human Schlemm’s canal endothelial cell monolayers. Investig. Ophthalmol. Vis. Sci. 2010, 51, 6633–6638. [Google Scholar] [CrossRef] [Green Version]
- Lei, Y.; Overby, D.R.; Boussommier-Calleja, A.; Stamer, W.D.; Ethier, C.R. Outflow physiology of the mouse eye: Pressure dependence and washout. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1865–1871. [Google Scholar] [CrossRef] [Green Version]
- Stamer, W.D.; Seftor, R.E.; Williams, S.K.; Samaha, H.A.; Snyder, R.W. Isolation and culture of human trabecular meshwork cells by extracellular matrix digestion. Curr. Eye Res. 1995, 14, 611–617. [Google Scholar] [CrossRef]
- Stamer, W.D.; Roberts, B.C.; Epstein, D.L.; Allingham, R.R. Isolation of primary open-angle glaucomatous trabecular meshwork cells from whole eye tissue. Curr. Eye Res. 2000, 20, 347–350. [Google Scholar] [CrossRef]
- Keller, K.E.; Bhattacharya, S.K.; Borras, T.; Brunner, T.M.; Chansangpetch, S.; Clark, A.F.; Dismuke, W.M.; Du, Y.; Elliott, M.H.; Ethier, C.R.; et al. Consensus recommendations for trabecular meshwork cell isolation, characterization and culture. Exp. Eye Res. 2018, 171, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Strober, W. Trypan blue exclusion test of cell viability. Curr. Protoc. Immunol. 2015, 111, A3.B.1–A3.B.2. [Google Scholar] [CrossRef] [PubMed]
- Honjo, M.; Tanihara, H.; Kameda, T.; Kawaji, T.; Yoshimura, N.; Araie, M. Potential role of Rho-associated protein kinase inhibitor Y-27632 in glaucoma filtration surgery. Investig. Ophthalmol. Vis. Sci. 2007, 48, 5549–5557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yantorno, R.E.; Coca-Prados, M.; Krupin, T.; Civan, M.M. Volume regulation of cultured, transformed, non-pigmented epithelial cells from human ciliary body. Exp. Eye Res. 1989, 49, 423–437. [Google Scholar] [CrossRef]
- Luna, C.; Li, G.; Huang, J.; Qiu, J.; Wu, J.; Yuan, F.; Epstein, D.L.; Gonzalez, P. Regulation of trabecular meshwork cell contraction and intraocular pressure by miR-200c. PLoS ONE 2012, 7, e51688. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Protein Name | Accession No | Fold Change | Log2 (Fold Change) | p-Value |
|---|---|---|---|---|---|
| NAGA | Alpha-N-acetylgalactosaminidase | P17050 | 0.2 | −2.3 | ** |
| PBXIP1 | Pre-B-cell leukemia transcription factor-interacting protein 1 | Q96AQ6 | 0.4 | −1.3 | * |
| SAE1 | Isoform 3 of SUMO-activating enzyme subunit 1 | Q9UBE0-3 | 0.4 | −1.2 | ** |
| BCAP29 | Isoform 2 of B-cell receptor-associated protein 29 | Q9UHQ4-2 | 0.5 | −1.2 | * |
| AKR1B10 | Aldo-keto reductase family 1 member B10 | O60218 | 0.5 | −1.2 | * |
| PSMD10 | 26S proteasome non-ATPase regulatory subunit 10 | O75832 | 0.5 | −1.1 | ** |
| PDCD6 | Programmed cell death protein 6 | O75340 | 0.5 | −1.1 | * |
| XRCC6 | X-ray repair cross-complementing protein 6 | P12956 | 0.6 | −0.8 | * |
| DDT | D-dopachrome decarboxylase | P30046 | 0.6 | −0.8 | *** |
| MANF | Mesencephalic astrocyte-derived neurotrophic factor | P55145 | 0.6 | −0.8 | * |
| ARPC1A | Actin-related protein 2/3 complex subunit 1A | Q92747 | 0.7 | −0.6 | * |
| MIF | Macrophage migration inhibitory factor | P14174 | 0.7 | −0.6 | * |
| KLC1 | Isoform I of Kinesin light chain 1 | Q07866-9 | 0.7 | −0.5 | * |
| DAB2 | Disabled homolog 2 | P98082 | 0.7 | −0.5 | * |
| AHCYL1 | S-adenosylhomocysteine hydrolase-like protein 1 | O43865 | 1.3 | 0.3 | * |
| TUFM | Elongation factor Tu, mitochondrial | P49411 | 1.3 | 0.3 | * |
| ATP5PO | ATP synthase subunit O, mitochondrial | P48047 | 1.3 | 0.4 | * |
| EML4 | Echinoderm microtubule-associated protein-like 4 | Q9HC35 | 1.3 | 0.4 | * |
| RUVBL2 | RuvB-like 2 | Q9Y230 | 1.3 | 0.4 | * |
| RBMX | RNA-binding motif protein, X chromosome | P38159 | 1.4 | 0.4 | * |
| ACTA2 | Actin, aortic smooth muscle | P62736 | 1.4 | 0.5 | ** |
| CTSB | Cathepsin B | P07858 | 1.5 | 0.6 | ** |
| PA2G4 | Proliferation-associated protein 2G4 | Q9UQ80 | 1.5 | 0.6 | * |
| CARS | Isoform 3 of Cysteine--tRNA ligase, cytoplasmic | P49589-3 | 1.5 | 0.6 | * |
| PDHB | Pyruvate dehydrogenase E1 component subunit beta, mitochondrial | P11177 | 1.6 | 0.7 | * |
| SLC25A3 | Isoform B of Phosphate carrier protein, mitochondrial | Q00325-2 | 1.8 | 0.8 | * |
| XPNPEP1 | Xaa-Pro aminopeptidase 1 | Q9NQW7 | 2.2 | 1.1 | * |
| TLR7 | Toll-like receptor 7 | Q9NYK1 | 2.2 | 1.2 | * |
| PFDN6 | Prefoldin subunit 6 | O15212 | 2.7 | 1.4 | * |
| CPA4 | Carboxypeptidase A4 | Q9UI42 | 3.1 | 1.7 | ** |
| MGARP | Protein MGARP | Q8TDB4 | 3.8 | 1.9 | * |
| Upstream Regulator | Description | Molecule Type | p-Value | Activation z-Score |
|---|---|---|---|---|
| TP53 | Tumor protein p53 | transcription regulator | ** | 0.7 |
| NFKBIA | NF-kappa-B inhibitor alpha | transcription regulator | ** | 0.8 |
| MAPK14 | Mitogen-activated protein kinase 14 | kinase | ** | 1.2 |
| GATA6 | GATA binding protein 6 | transcription regulator | ** | −1.1 |
| CDKN1A | Cyclin dependent kinase inhibitor 1A | kinase | ** | 0.5 |
| OGA | O-GlcNAcase | enzyme | ** | 1.3 |
| HIF1A | hypoxia inducible factor 1 subunit alpha | transcription regulator | ** | 0.2 |
| AGT | Angiotensinogen | growth factor | ** | −0.4 |
| TGFB1 | transforming growth factor beta 1 | growth factor | ** | 0.6 |
| AKT1 | AKT serine/threonine kinase 1 | kinase | ** | 0.2 |
| MYC | MYC Proto-Oncogene, BHLH Transcription Factor | transcription regulator | ** | −1.6 |
| VEGFA | vascular endothelial growth factor A | growth factor | * | 2 |
| MAPK1 | mitogen-activated protein kinase 1 | kinase | ** | 0.2 |
| PTEN | phosphatase and tensin homolog | phosphatase | ** | 0.4 |
| EGFR | epidermal growth factor receptor | kinase | ** | 0.6 |
| FGF2 | fibroblast growth factor 2 | growth factor | ** | −2 |
| CEBPB | CCAAT Enhancer Binding Protein Beta | transcription regulator | ** | −0.8 |
| IL1B | interleukin 1 beta | cytokine | ** | −0.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.-l.; Shan, S.W.; Stamer, W.D.; Li, K.-k.; Chan, H.H.-l.; Civan, M.M.; To, C.-h.; Lam, T.C.; Do, C.-w. Mechanistic Effects of Baicalein on Aqueous Humor Drainage and Intraocular Pressure. Int. J. Mol. Sci. 2022, 23, 7372. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137372
Li H-l, Shan SW, Stamer WD, Li K-k, Chan HH-l, Civan MM, To C-h, Lam TC, Do C-w. Mechanistic Effects of Baicalein on Aqueous Humor Drainage and Intraocular Pressure. International Journal of Molecular Sciences. 2022; 23(13):7372. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137372
Chicago/Turabian StyleLi, Hoi-lam, Sze Wan Shan, W. Daniel Stamer, King-kit Li, Henry Ho-lung Chan, Mortimer M. Civan, Chi-ho To, Thomas Chuen Lam, and Chi-wai Do. 2022. "Mechanistic Effects of Baicalein on Aqueous Humor Drainage and Intraocular Pressure" International Journal of Molecular Sciences 23, no. 13: 7372. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137372