
Bile Duct Ligation Impairs Function and Expression of Mrp1 at Rat Blood–Retinal Barrier via Bilirubin-Induced P38 MAPK Pathway Activations

 , , , and
, , , and
Abstract
:1. Introduction
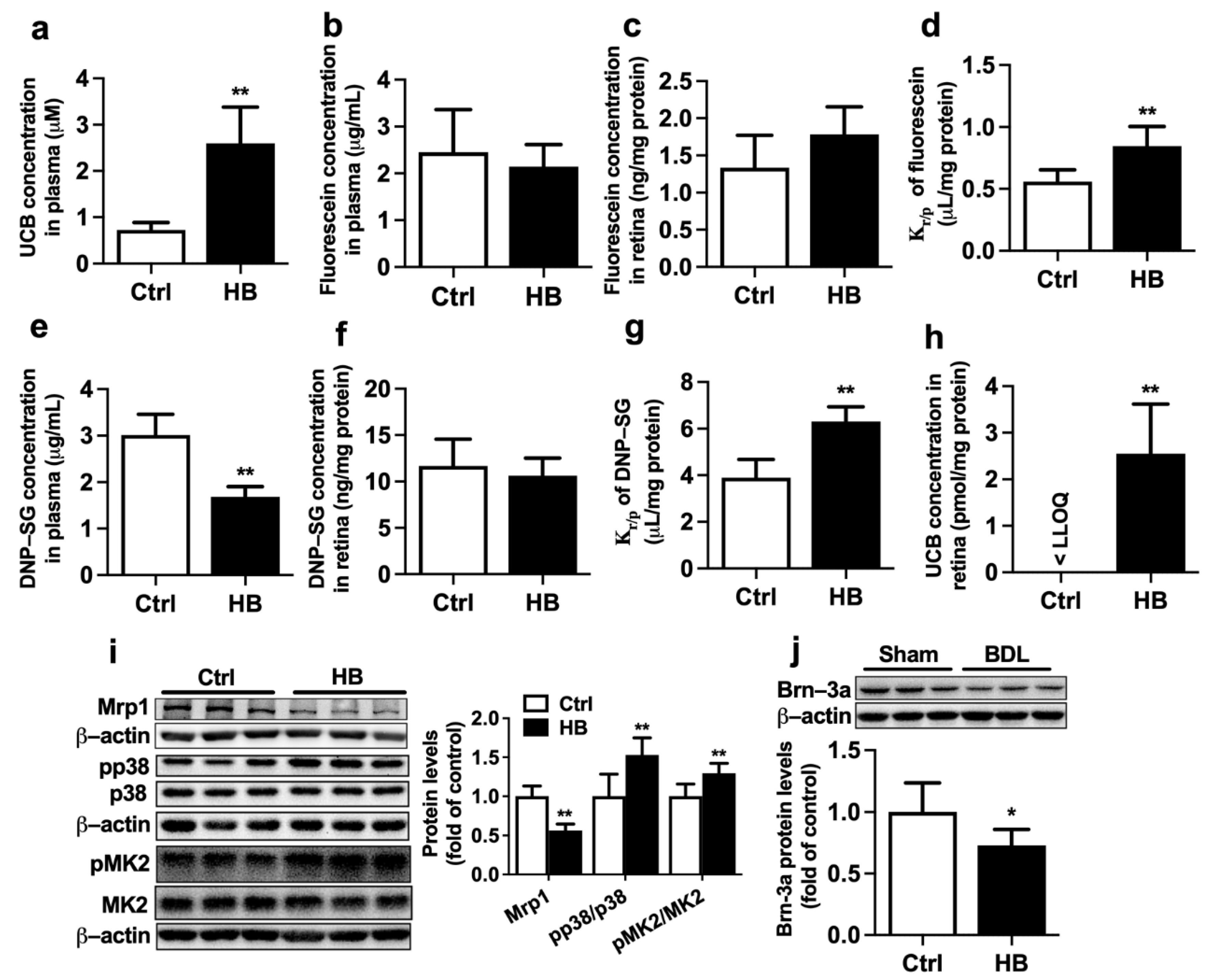
2. Results
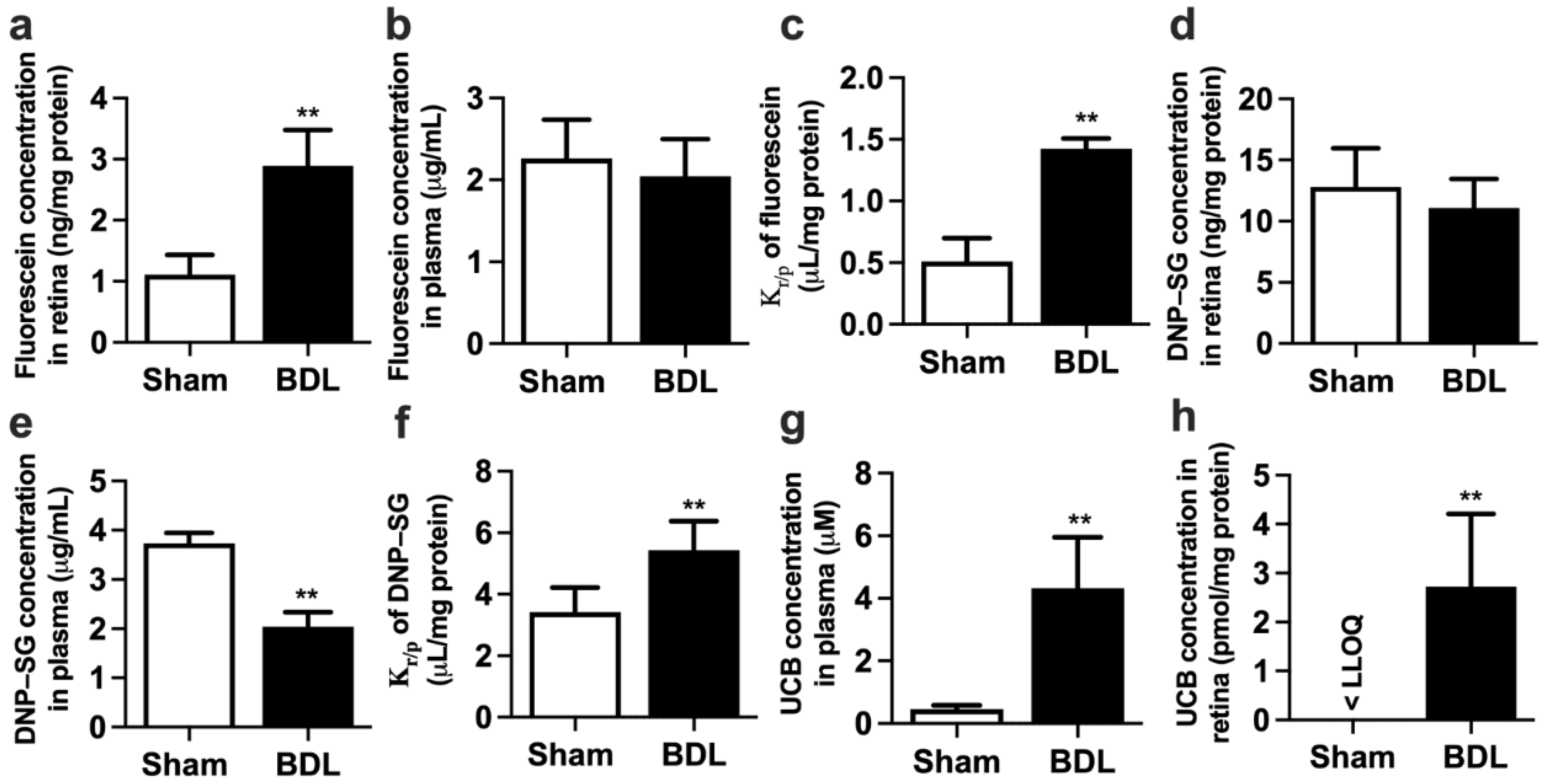
2.1. Physiological and Biochemical Parameters in Sham and BDL Rats
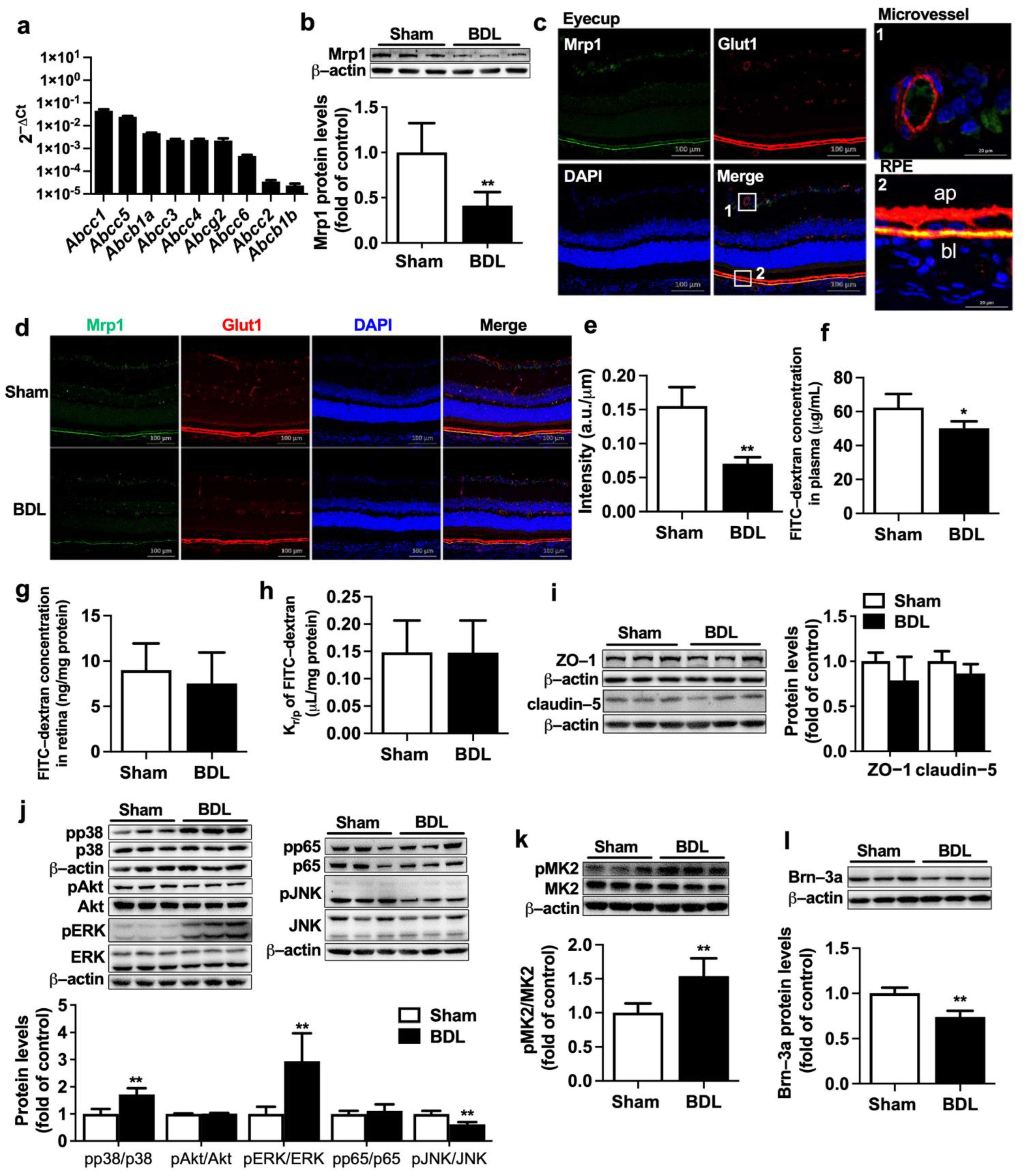
2.2. Effect of BDL on Function and Expression of Mrp1 at Rat BRB
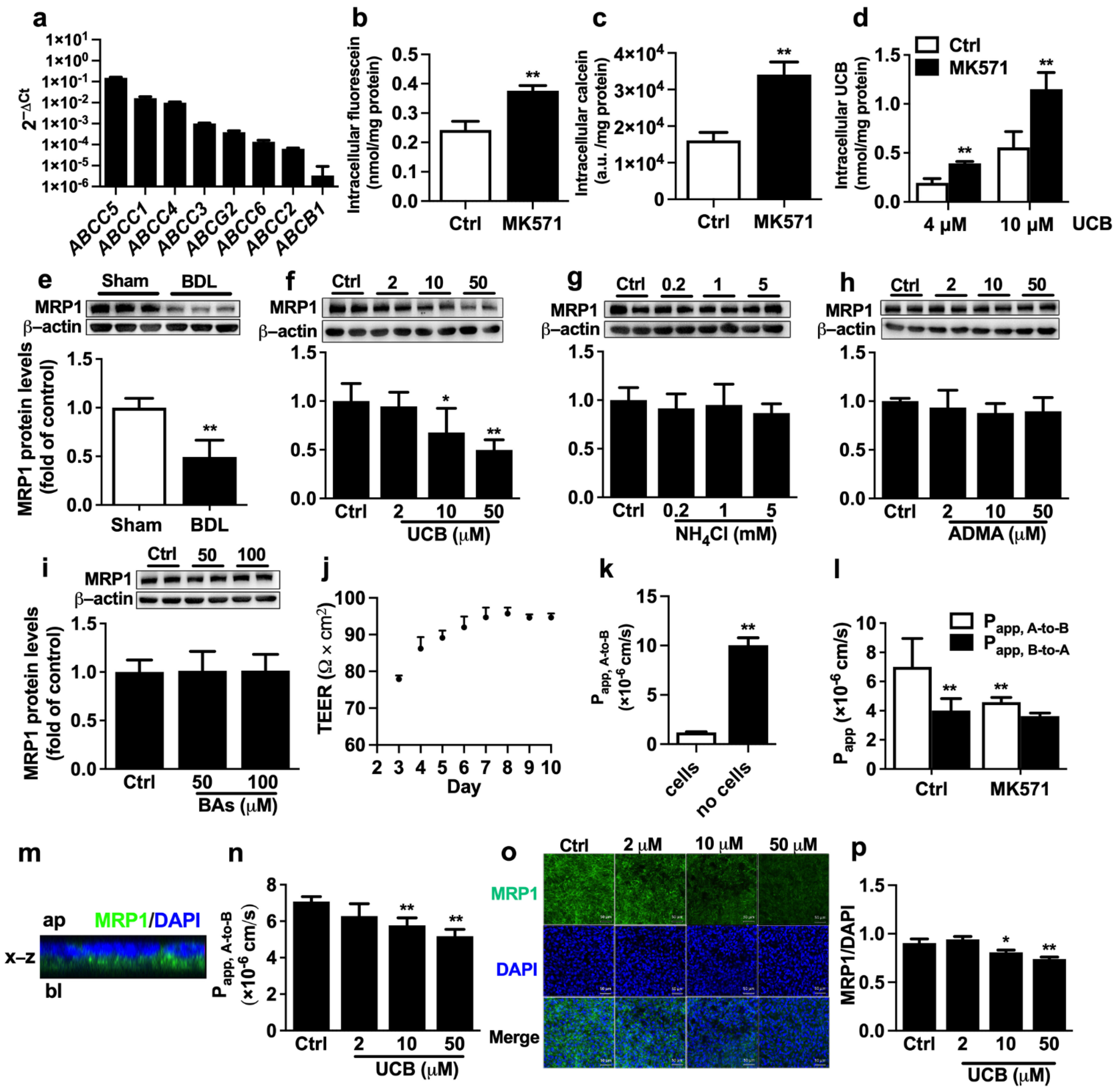
2.3. Effects of Abnormally Altered Components in BDL Rat Serum on Function and Expression of MRP1 in ARPE-19 Cells
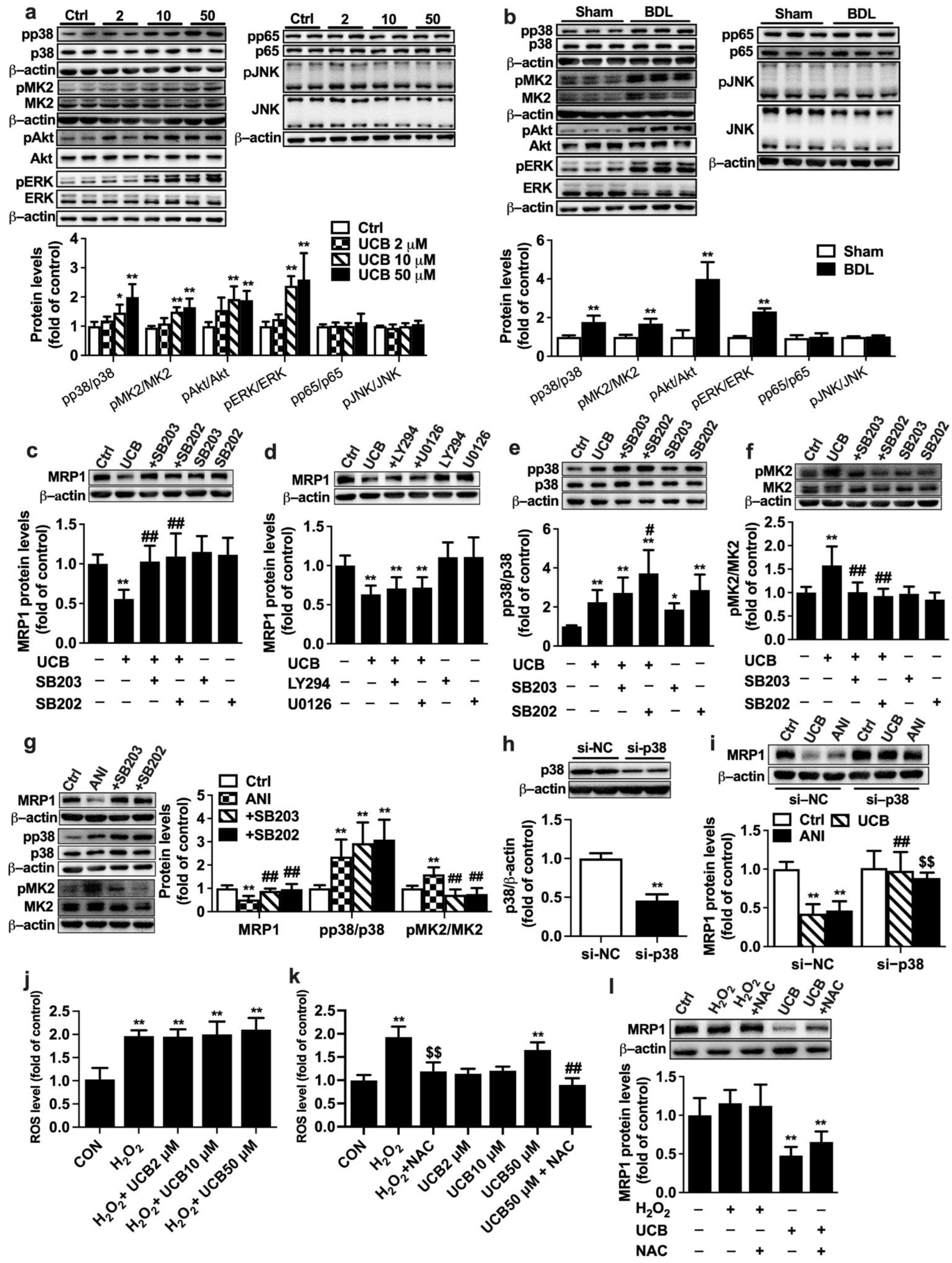
2.4. Involvement of P38 MAPK Pathway in UCB-Induced Downregulation of MRP1 Expression in APRE-19 Cells
2.5. Effect of Bilirubin on the Function and Expression of Mrp1 at Rat BRB
3. Discussion
4. Methods and Materials
4.1. Reagents
4.2. Animals
4.3. Development of BDL Rats
4.4. Distributions of Fluorescein and DNP-SG in Rat Retina
4.5. Drug Assays
4.6. Immunofluorescence Staining
4.7. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.8. Cell Culture and Drug Treatment
4.9. P38 Knockdown with siRNA
4.10. Uptake of MRP1 Substrates by ARPE-19 Cells
4.11. Transport of Fluorescein across ARPE-19 Monolayer
4.12. Western Blot
4.13. Development of HB Rats
4.14. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cabrera DeBuc, D.; Somfai, G.M.; Koller, A. Retinal microvascular network alterations: Potential biomarkers of cerebrovascular and neural diseases. Am. J. Physiol. Heart Circ. Physiol. 2017, 312, H201–H212. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhang, D.; Ji, J.; Wang, Y.; Zhang, R. Central retina changes in Parkinson’s disease: A systematic review and meta-analysis. J. Neurol. 2021, 268, 4646–4654. [Google Scholar] [CrossRef] [PubMed]
- Reichenbach, A.; Fuchs, U.; Kasper, M.; el-Hifnawi, E.; Eckstein, A.K. Hepatic retinopathy: Morphological features of retinal glial (Müller) cells accompanying hepatic failure. Acta Neuropathol. 1995, 90, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Hampel, H.; Toschi, N.; Babiloni, C.; Baldacci, F.; Black, K.L.; Bokde, A.L.W.; Bun, R.S.; Cacciola, F.; Cavedo, E.; Chiesa, P.A.; et al. Revolution of Alzheimer precision neurology. Passageway of systems biology and neurophysiology. J. Alzheimers Dis. 2018, 64 (Suppl. S1), S47–S105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckstein, A.K.; Reichenbach, A.; Jacobi, P.; Weber, P.; Gregor, M.; Zrenner, E. Hepatic retinopathia. Changes in retinal function. Vision Res. 1997, 37, 1699–1706. [Google Scholar] [CrossRef] [Green Version]
- Uhlmann, S.; Uhlmann, D.; Hauss, J.; Reichenbach, A.; Wiedemann, P.; Faude, F. Recovery from hepatic retinopathy after liver transplantation. Graefes Arch. Clin. Exp. Ophthalmol. 2003, 241, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Liccardo, D.; Mosca, A.; Petroni, S.; Valente, P.; Giordano, U.; Mico, A.G.; Pescosolido, S.; Buzzonetti, L.; Nobili, V. The association between retinal microvascular changes, metabolic risk factors, and liver histology in pediatric patients with non-alcoholic fatty liver disease (NAFLD). J. Gastroenterol. 2015, 50, 903–912. [Google Scholar] [CrossRef]
- Jung, E.; Kim, J. Aloin inhibits Müller cells swelling in a rat model of thioacetamide-induced hepatic retinopathy. Molecules 2018, 23, 2806. [Google Scholar] [CrossRef] [Green Version]
- Algazo, M.A.; Amiri-Ghashlaghi, S.; Delfan, B.; Hassanzadeh, G.; Sabbagh-Ziarani, F.; Jazaeri, F.; Dehpour, A.R. Cirrhosis-induced morphological changes in the retina: Possible role of endogenous opioid. Int. J. Ophthalmol. 2015, 8, 681–684. [Google Scholar] [CrossRef]
- Albrecht, J.; Gadamski, R.; Kuhrt, H.; Walski, M.; Reichenbach, A. Retinal gliopathy accompanying thioacetamide-induced liver insufficiency: Light and electron microscopic observations. Acta Neuropathol. 1998, 96, 57–66. [Google Scholar] [CrossRef]
- Chen, P.; Chen, H.; Zang, X.; Chen, M.; Jiang, H.; Han, S.; Wu, X. Expression of efflux transporters in human ocular tissues. Drug Metab. Dispos. 2013, 41, 1934–1948. [Google Scholar] [CrossRef] [PubMed]
- Hosoya, K.; Yamamoto, A.; Akanuma, S.; Tachikawa, M. Lipophilicity and transporter influence on blood-retinal barrier permeability: A comparison with blood-brain barrier permeability. Pharm. Res. 2010, 27, 2715–2724. [Google Scholar] [CrossRef]
- Pelkonen, L.; Sato, K.; Reinisalo, M.; Kidron, H.; Tachikawa, M.; Watanabe, M.; Uchida, Y.; Urtti, A.; Terasaki, T. LC-MS/MS based quantitation of ABC and SLC transporter proteins in plasma membranes of cultured primary human retinal pigment epithelium cells and immortalized ARPE19 cell line. Mol. Pharm. 2017, 14, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Liu, X. Roles of drug transporters in blood-retinal barrier. Adv. Exp. Med. Biol. 2019, 1141, 467–504. [Google Scholar] [CrossRef] [PubMed]
- Hosoya, K.; Tomi, M. Advances in the cell biology of transport via the inner blood-retinal barrier: Establishment of cell lines and transport functions. Biol. Pharm. Bull. 2005, 28, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Mannermaa, E.; Vellonen, K.S.; Ryhanen, T.; Kokkonen, K.; Ranta, V.P.; Kaarniranta, K.; Urtti, A. Efflux protein expression in human retinal pigment epithelium cell lines. Pharm. Res. 2009, 26, 1785–1791. [Google Scholar] [CrossRef]
- Aukunuru, J.V.; Sunkara, G.; Bandi, N.; Thoreson, W.B.; Kompella, U.B. Expression of multidrug resistance-associated protein (MRP) in human retinal pigment epithelial cells and its interaction with BAPSG, a novel aldose reductase inhibitor. Pharm. Res. 2001, 18, 565–572. [Google Scholar] [CrossRef]
- Xu, P.; Ling, Z.L.; Zhang, J.; Li, Y.; Shu, N.; Zhong, Z.Y.; Chen, Y.; Di, X.Y.; Wang, Z.J.; Liu, L.; et al. Unconjugated bilirubin elevation impairs the function and expression of breast cancer resistance protein (BCRP) at the blood-brain barrier in bile duct-ligated rats. Acta Pharmacol. Sin. 2016, 37, 1129–1140. [Google Scholar] [CrossRef] [Green Version]
- Qin, Y.Y.; Xu, P.; Wu, T.; Qian, C.Q.; Fan, Y.L.; Gen, D.H.; Zhu, L.; Kong, W.M.; Yang, H.Y.; Xu, F.; et al. Bile duct ligation enhances AZT CNS toxicity partly by impairing the expression and function of BCRP in rat brain. Acta Pharmacol. Sin. 2020, 41, 181–191. [Google Scholar] [CrossRef]
- Wu, T.; Sheng, Y.; Qin, Y.Y.; Kong, W.M.; Jin, M.M.; Yang, H.Y.; Zheng, X.K.; Dai, C.; Liu, M.; Liu, X.D.; et al. Bile duct ligation causes opposite impacts on the expression and function of BCRP and P-gp in rat brain partly via affecting membrane expression of ezrin/radixin/moesin proteins. Acta Pharmacol. Sin. 2021, 42, 1942–1950. [Google Scholar] [CrossRef]
- Jin, S.; Wang, X.T.; Liu, L.; Yao, D.; Liu, C.; Zhang, M.; Guo, H.F.; Liu, X.D. P-glycoprotein and multidrug resistance-associated protein 2 are oppositely altered in brain of rats with thioacetamide-induced acute liver failure. Liver Int. 2013, 33, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, J.; Xu, P.; Sun, B.; Zhong, Z.; Liu, C.; Ling, Z.; Chen, Y.; Shu, N.; Zhao, K.; et al. Acute liver failure impairs function and expression of breast cancer-resistant protein (BCRP) at rat blood-brain barrier partly via ammonia-ROS-ERK1/2 activation. J. Neurochem. 2016, 138, 282–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, S.; Setoguchi, C.; Kawazu, K.; Hosoya, K. Impact of P-glycoprotein on blood-retinal barrier permeability: Comparison of blood-aqueous humor and blood-brain barrier using mdr1a knockout rats. Investig. Ophthalmol. Vis. Sci. 2014, 55, 4650–4658. [Google Scholar] [CrossRef] [Green Version]
- Chapy, H.; Saubamea, B.; Tournier, N.; Bourasset, F.; Behar-Cohen, F.; Decleves, X.; Scherrmann, J.M.; Cisternino, S. Blood-brain and retinal barriers show dissimilar ABC transporter impacts and concealed effect of P-glycoprotein on a novel verapamil influx carrier. Br. J. Pharmacol. 2016, 173, 497–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Xiang, C.D.; Gale, D.; Carreiro, S.; Wu, E.Y.; Zhang, E.Y. Drug transporter and cytochrome P450 mRNA expression in human ocular barriers: Implications for ocular drug disposition. Drug Metab. Dispos. 2008, 36, 1300–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sreekumar, P.G.; Spee, C.; Ryan, S.J.; Cole, S.P.; Kannan, R.; Hinton, D.R. Mechanism of RPE cell death in alpha-crystallin deficient mice: A novel and critical role for MRP1-mediated GSH efflux. PLoS ONE 2012, 7, e33420. [Google Scholar] [CrossRef] [Green Version]
- Gerhart, D.Z.; Leino, R.L.; Drewes, L.R. Distribution of monocarboxylate transporters MCT1 and MCT2 in rat retina. Neuroscience 1999, 92, 367–375. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, J.; Li, P.; Xie, Q.; Sun, B.; Li, Y.; Chen, Y.; Zhao, K.; Yang, T.; Zhu, L.; et al. Increase in P-glycoprotein levels in the blood-brain barrier of partial portal vein ligation /chronic hyperammonemia rats is medicated by ammonia/reactive oxygen species/ERK1/2 activation: In vitro and in vivo studies. Eur. J. Pharmacol. 2019, 846, 119–127. [Google Scholar] [CrossRef]
- Xie, Q.S.; Zhang, J.X.; Liu, M.; Liu, P.H.; Wang, Z.J.; Zhu, L.; Jiang, L.; Jin, M.M.; Liu, X.N.; Liu, L.; et al. Short-chain fatty acids exert opposite effects on the expression and function of p-glycoprotein and breast cancer resistance protein in rat intestine. Acta Pharmacol. Sin. 2021, 42, 470–481. [Google Scholar] [CrossRef]
- Chen, J.R.; Jia, X.H.; Wang, H.; Yi, Y.J.; Wang, J.Y.; Li, Y.J. Timosaponin A-III reverses multi-drug resistance in human chronic myelogenous leukemia K562/ADM cells via downregulation of MDR1 and MRP1 expression by inhibiting PI3K/Akt signaling pathway. Int. J. Oncol. 2016, 48, 2063–2070. [Google Scholar] [CrossRef] [Green Version]
- Beamer, E.; Correa, S.A.L. The p38(MAPK)-MK2 signaling axis as a critical link between inflammation and synaptic transmission. Front. Cell Dev. Biol. 2021, 9, 635636. [Google Scholar] [CrossRef] [PubMed]
- Moyer, A.L.; Ramadan, R.T.; Thurman, J.; Burroughs, A.; Callegan, M.C. Bacillus cereus induces permeability of an in vitro blood-retina barrier. Infect. Immun. 2008, 76, 1358–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Jiang, M.S.; Adams, J.L.; Lee, J.C. Pyridinylimidazole compound SB 203580 inhibits the activity but not the activation of p38 mitogen-activated protein kinase. Biochem. Biophys. Res. Commun. 1999, 263, 825–831. [Google Scholar] [CrossRef]
- Xu, L.; Chen, S.; Bergan, R.C. MAPKAPK2 and HSP27 are downstream effectors of p38 MAP kinase-mediated matrix metalloproteinase type 2 activation and cell invasion in human prostate cancer. Oncogene 2006, 25, 2987–2998. [Google Scholar] [CrossRef] [Green Version]
- Buffington, C.A.; Woodworth, B.E. Excretion of fluorescein in the urine of women with interstitial cystitis. J. Urol. 1997, 158 Pt 1, 786–789. [Google Scholar] [CrossRef]
- Hong, S.; Li, S.; Meng, X.; Li, P.; Wang, X.; Su, M.; Liu, X.; Liu, L. Bile duct ligation differently regulates protein expressions of organic cation transporters in intestine, liver and kidney of rats through activation of farnesoid X receptor by cholate and bilirubin. Acta Pharm. Sin. B 2022, in press. [Google Scholar] [CrossRef]
- Purucker, E.; Wernze, W.; Krandik, G. Glutathione in plasma, liver, and kidney in the development of CCl4-induced cirrhosis of the rat. Res. Exp. Med. 1995, 195, 193–199. [Google Scholar] [CrossRef]
- Villanueva, J.A.; Esfandiari, F.; Wong, D.H.; Ahmad, I.; Melnyk, S.; James, S.J.; Halsted, C.H. Abnormal transsulfuration and glutathione metabolism in the micropig model of alcoholic liver disease. Alcohol. Clin. Exp. Res. 2006, 30, 1262–1270. [Google Scholar] [CrossRef]
- Ideo, G.; Morganti, A.; Dioguardi, N. Gamma-glutamyl transpeptidase: A clinical and experimental study. Digestion 1972, 5, 326–336. [Google Scholar] [CrossRef]
- Olynyk, J.K.; Yeoh, G.C.; Ramm, G.A.; Clarke, S.L.; Hall, P.M.; Britton, R.S.; Bacon, B.R.; Tracy, T.F. Gadolinium chloride suppresses hepatic oval cell proliferation in rats with biliary obstruction. Am. J. Pathol. 1998, 152, 347–352. [Google Scholar]
- Villanueva, S.S.; Ruiz, M.L.; Soroka, C.J.; Cai, S.Y.; Luquita, M.G.; Torres, A.M.; Sanchez Pozzi, E.J.; Pellegrino, J.M.; Boyer, J.L.; Catania, V.A.; et al. Hepatic and extrahepatic synthesis and disposition of dinitrophenyl-S-glutathione in bile duct-ligated rats. Drug Metab. Dispos. 2006, 34, 1301–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgunder, J.M.; Lauterburg, B.H. Decreased production of glutathione in patients with cirrhosis. Eur. J. Clin. Investig. 1987, 17, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Gazzin, S.; Berengeno, A.L.; Strazielle, N.; Fazzari, F.; Raseni, A.; Ostrow, J.D.; Wennberg, R.; Ghersi-Egea, J.F.; Tiribelli, C. Modulation of Mrp1 (ABCc1) and Pgp (ABCb1) by bilirubin at the blood-CSF and blood-brain barriers in the Gunn rat. PLoS ONE 2011, 6, e16165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cekic, D.; Bellarosa, C.; Garcia-Mediavilla, M.V.; Rigato, I.; Pascolo, L.; Ostrow, J.D.; Tiribelli, C. Upregulation in the expression of multidrug resistance protein Mrp1 mRNA and protein by increased bilirubin production in rat. Biochem. Biophys. Res. Commun. 2003, 311, 891–896. [Google Scholar] [CrossRef]
- Gennuso, F.; Fernetti, C.; Tirolo, C.; Testa, N.; L’Episcopo, F.; Caniglia, S.; Morale, M.C.; Ostrow, J.D.; Pascolo, L.; Tiribelli, C.; et al. Bilirubin protects astrocytes from its own toxicity by inducing up-regulation and translocation of multidrug resistance-associated protein 1 (Mrp1). Proc. Natl. Acad. Sci. USA 2004, 101, 2470–2475. [Google Scholar] [CrossRef] [Green Version]
- Falcao, A.S.; Bellarosa, C.; Fernandes, A.; Brito, M.A.; Silva, R.F.; Tiribelli, C.; Brites, D. Role of multidrug resistance-associated protein 1 expression in the in vitro susceptibility of rat nerve cell to unconjugated bilirubin. Neuroscience 2007, 144, 878–888. [Google Scholar] [CrossRef]
- Kapitulnik, J. Bilirubin: An endogenous product of heme degradation with both cytotoxic and cytoprotective properties. Mol. Pharmacol. 2004, 66, 773–779. [Google Scholar] [CrossRef] [Green Version]
- Kapitulnik, J.; Benaim, C.; Sasson, S. Endothelial cells derived from the blood-brain barrier and islets of langerhans differ in their response to the effects of bilirubin on oxidative stress under hyperglycemic conditions. Front. Pharmacol. 2012, 3, 131. [Google Scholar] [CrossRef] [Green Version]
- Palmela, I.; Cardoso, F.L.; Bernas, M.; Correia, L.; Vaz, A.R.; Silva, R.F.; Fernandes, A.; Kim, K.S.; Brites, D.; Brito, M.A. Elevated levels of bilirubin and long-term exposure impair human brain microvascular endothelial cell integrity. Curr. Neurovasc. Res. 2011, 8, 153–169. [Google Scholar] [CrossRef]
- Gou, P.; Qi, X.; Yuan, R.; Li, H.; Gao, X.; Wang, J.; Zhang, B. Tet1-mediated DNA demethylation involves in neuron damage induced by bilirubin in vitro. Toxicol. Mech. Methods 2018, 28, 55–61. [Google Scholar] [CrossRef]
- Bianco, A.; Dvorak, A.; Capkova, N.; Gironde, C.; Tiribelli, C.; Furger, C.; Vitek, L.; Bellarosa, C. The extent of intracellular accumulation of bilirubin determines its anti- or pro-oxidant effect. Int. J. Mol. Sci. 2020, 21, 8101. [Google Scholar] [CrossRef] [PubMed]
- Idelman, G.; Smith, D.L.H.; Zucker, S.D. Bilirubin inhibits the up-regulation of inducible nitric oxide synthase by scavenging reactive oxygen species generated by the toll-like receptor 4-dependent activation of NADPH oxidase. Redox Biol. 2015, 5, 398–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghaffar, T.; Marwat, Z.I.; Ullah, F.; Khan, S.; Hassan Aamir, A.U. Association of serum total bilirubin level with diabetic retinopathy in type 2 diabetes mellitus. J. Ayub Med. Coll. Abbottabad 2016, 28, 537–541. [Google Scholar]
- Zhu, B.; Wu, X.; Bi, Y.; Yang, Y. Effect of bilirubin concentration on the risk of diabetic complications: A meta-analysis of epidemiologic studies. Sci. Rep. 2017, 7, 41681. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Wang, J.; He, Y. The U-shaped association between bilirubin and diabetic retinopathy risk: A five-year cohort based on 5323 male diabetic patients. J. Diabetes Res. 2018, 2018, 4603087. [Google Scholar] [CrossRef] [PubMed]
- Rigato, I.; Pascolo, L.; Fernetti, C.; Ostrow, J.D.; Tiribelli, C. The human multidrug-resistance-associated protein MRP1 mediates ATP-dependent transport of unconjugated bilirubin. Biochem. J. 2004, 383 Pt 2, 335–341. [Google Scholar] [CrossRef] [Green Version]
- Corich, L.; Aranda, A.; Carrassa, L.; Bellarosa, C.; Ostrow, J.D.; Tiribelli, C. The cytotoxic effect of unconjugated bilirubin in human neuroblastoma SH-SY5Y cells is modulated by the expression level of MRP1 but not MDR1. Biochem. J. 2009, 417, 305–312. [Google Scholar] [CrossRef] [Green Version]
- Albanna, W.; Luke, J.N.; Schubert, G.A.; Dibue-Adjei, M.; Kotliar, K.; Hescheler, J.; Clusmann, H.; Steiger, H.J.; Hanggi, D.; Kamp, M.A.; et al. Modulation of Cav2.3 channels by unconjugated bilirubin (UCB)—Candidate mechanism for UCB-induced neuromodulation and neurotoxicity. Mol. Cell. Neurosci. 2019, 96, 35–46. [Google Scholar] [CrossRef]
- Vaidya, S.S.; Walsh, S.W.; Gerk, P.M. Formation and efflux of ATP-binding cassette transporter substrate 2,4-dinitrophenyl-S-glutathione from cultured human term placental villous tissue fragments. Mol. Pharm. 2009, 6, 1689–1702. [Google Scholar] [CrossRef]
- Yang, J.J.; Ann, D.K.; Kannan, R.; Lee, V.H. Multidrug resistance protein 1 (MRP1) in rabbit conjunctival epithelial cells: Its effect on drug efflux and its regulation by adenoviral infection. Pharm. Res. 2007, 24, 1490–1500. [Google Scholar] [CrossRef]
- Noailles, A.; Maneu, V.; Campello, L.; Gomez-Vicente, V.; Lax, P.; Cuenca, N. Persistent inflammatory state after photoreceptor loss in an animal model of retinal degeneration. Sci. Rep. 2016, 6, 33356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musashi, K.; Kiryu, J.; Miyamoto, K.; Miyahara, S.; Katsuta, H.; Tamura, H.; Hirose, F.; Yoshimura, N. Thrombin inhibitor reduces leukocyte-endothelial cell interactions and vascular leakage after scatter laser photocoagulation. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2561–2566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schou-Pedersen, A.M.V.; Lykkesfeldt, J. Comparison of three sample preparation procedures for the quantification of L-arginine, asymmetric dimethylarginine, and symmetric dimethylarginine in human plasma using HPLC-FLD. J. Anal. Methods Chem. 2018, 2018, 6148515. [Google Scholar] [CrossRef] [Green Version]
- Zelenka, J.; Lenicek, M.; Muchova, L.; Jirsa, M.; Kudla, M.; Balaz, P.; Zadinova, M.; Ostrow, J.D.; Wong, R.J.; Vitek, L. Highly sensitive method for quantitative determination of bilirubin in biological fluids and tissues. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2008, 867, 37–42. [Google Scholar] [CrossRef]
- Shao, A.; Xu, L.; Wu, X.; Liu, S.; Lu, Y.; Fan, C. Gal epitope expression and immunological properties in iGb3S deficient mice. Sci. Rep. 2018, 8, 15433. [Google Scholar] [CrossRef]
- Thakare, R.; Alamoudi, J.A.; Gautam, N.; Rodrigues, A.D.; Alnouti, Y. Species differences in bile acids II. Bile acid metabolism. J. Appl. Toxicol. 2018, 38, 1336–1352. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, L.; Jin, J.; Luan, Y.; Chen, C.; Li, Y.; Chu, H.; Wang, X.; Liao, G.; Yu, Y.; et al. p38 inhibition provides anti-DNA virus immunity by regulation of USP21 phosphorylation and STING activation. J. Exp. Med. 2017, 214, 991–1010. [Google Scholar] [CrossRef]
- Szabo, M.; Veres, Z.; Batai-Konczos, A.; Kekesi, O.; Kis, E.; Szabo, K.; Jemnitz, K. Statins alter the hepatobiliary transport of unconjugated and conjugated bilirubin in sandwich-cultured rat hepatocytes. Toxicol. In Vitro 2014, 28, 1136–1143. [Google Scholar] [CrossRef] [Green Version]
- Praveschotinunt, P.; Duraj-Thatte, A.M.; Gelfat, I.; Bahl, F.; Chou, D.B.; Joshi, N.S. Engineered E. coli Nissle 1917 for the delivery of matrix-tethered therapeutic domains to the gut. Nat. Commun. 2019, 10, 5580. [Google Scholar] [CrossRef] [Green Version]
- Artursson, P. Epithelial transport of drugs in cell culture. I: A model for studying the passive diffusion of drugs over intestinal absorptive (Caco-2) cells. J. Pharm. Sci. 1990, 79, 476–482. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Sham Rats | BDL Rats |
|---|---|---|
| Body weight (BW) (g) | 225.0 ± 19.9 | 231.8 ± 15.5 |
| Liver weight (%BW) | 2.84 ± 0.15 | 6.84 ± 0.96 ** |
| Spleen weight (%BW) | 0.23 ± 0.04 | 0.55 ± 0.17 ** |
| ALT (IU/L) | 6.45 ± 4.55 | 22.47 ± 11.37 ** |
| AST (IU/L) | 20.22 ± 6.00 | 90.99 ± 31.51 ** |
| ALP (IU/L) | 114.90 ± 25.80 | 254.80 ± 47.67 ** |
| Total bilirubin (µmol/L) | ND a | 91.50 ± 15.50 ** |
| Total bile acids (µmol/L) | 29.50 ± 7.48 | 133.84 ± 23.95 ** |
| Serum ammonia (µmol/L) | 210.41 ± 27.41 | 276.89 ± 45.35 * |
| ADMA (ng/mL) | 174.49 ± 32.84 | 277.29 ± 52.74 ** |
| SDMA (ng/mL) | 25.14 ± 11.45 | 37.71 ± 16.28 |
| Parameters | Control Rats | HB Rats |
|---|---|---|
| Body weight (BW) (g) | 241.2 ± 8.5 | 237.3 ± 10.7 |
| Liver weight (%BW) | 2.70 ± 0.48 | 3.31 ± 0.43 * |
| Spleen weight (%BW) | 0.23 ± 0.05 | 0.39 ± 0.06 ** |
| ALT (IU/L) | 8.70 ± 4.70 | 10.08 ± 4.58 |
| AST (IU/L) | 16.69 ± 4.42 | 17.69 ± 3.31 |
| ALP (IU/L) | 116.93 ± 24.89 | 111.47 ± 44.67 |
| Total bilirubin (µmol/L) | ND a | 12.32 ± 1.23 ** |
| Total bile acids (µmol/L) | 43.63 ± 10.82 | 37.29 ± 8.04 |
| Genes | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| rAbcb1a | CAACCAGCATTCTCCATAATA | CCCAAGGATCAGGAACAATA |
| rAbcb1b | CTCGCTGCTATCATCCACGGAAC | CGCTGACGGTCTGTGTACTGTTG |
| rAbcg2 | GTACTTTGCATCAGCAGGTTACCACT | ATTACAGCCGAAGAATCTCCGTTG |
| rAbcc1 | GGCCTACTGAAGAGCAAGAC | GGATGATGATGACAGCTCC |
| rAbcc2 | GTGTTTCCACAGAGCGGCTAG | GCTAGGCTGATATCAAGGAG |
| rAbcc3 | ACACCGAGCCAGCCATATAC | ACATTGGCTCCGATAGCAAC |
| rAbcc4 | TCTGGGTGGAAATCGGAATC | GCAGAATAACCAGAATGGCCA |
| rAbcc5 | TGGAGAACGGGGACAACTT | AAAGGCGAGGTTTCAGCAG |
| rAbcc6 | GCCACTACACCTGCCTTGACTCACCA | TCATGGGTGCTATTAGGGCGGATCAA |
| rActb | GCTATGTTGCCCTAGACTTCG | GCCACAGGATTCCATACCCAG |
| hABCB1 | ACAGAGGGGATGGTCAGTGT | TCACGGCCATAGCGAATGTT |
| hABCG2 | AACCTGGTCTCAACGCCATC | GTCGCGGTGCTCCATTTATC |
| hABCC1 | CTACCTCCTGTGGCTGAATCTG | CATCAGCTTGATCCGATTGTCT |
| hABCC2 | ATTCAGACGACCATCCAAAACGAGTT | GCCATAAAGTAAAAGGGTCCAGGGAT |
| hABCC3 | CGCACACCGGCTTAACACTATCATGG | AAACCAGGAAAGGCCAGGAGGAAATC |
| hABCC4 | GGATCCAAGAACTGATGAGTTAAT | TCACAGTGCTGTCTCGAAAATAG |
| hABCC5 | ACCGCAGTCGTCGCACAGTCTCTCTC | GCGGGAACACACCAAACCACACAGCA |
| hABCC6 | GCCTGCAGTGCTGGAGATGGAAGTGA | GTCCTTCCGGCTCTGATGCTCTGTGA |
| hACTB | AAGAGCTACGAGCTGCCTGAC | TCCTGCTTGCTGATCCACAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.; Yang, Y.; Lin, Z.; Hong, S.; Jiang, L.; Zhou, H.; Yang, L.; Zhu, L.; Liu, X.; Liu, L. Bile Duct Ligation Impairs Function and Expression of Mrp1 at Rat Blood–Retinal Barrier via Bilirubin-Induced P38 MAPK Pathway Activations. Int. J. Mol. Sci. 2022, 23, 7666. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23147666
Li P, Yang Y, Lin Z, Hong S, Jiang L, Zhou H, Yang L, Zhu L, Liu X, Liu L. Bile Duct Ligation Impairs Function and Expression of Mrp1 at Rat Blood–Retinal Barrier via Bilirubin-Induced P38 MAPK Pathway Activations. International Journal of Molecular Sciences. 2022; 23(14):7666. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23147666
Chicago/Turabian StyleLi, Ping, Yiting Yang, Zijin Lin, Shijin Hong, Ling Jiang, Han Zhou, Lu Yang, Liang Zhu, Xiaodong Liu, and Li Liu. 2022. "Bile Duct Ligation Impairs Function and Expression of Mrp1 at Rat Blood–Retinal Barrier via Bilirubin-Induced P38 MAPK Pathway Activations" International Journal of Molecular Sciences 23, no. 14: 7666. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23147666