
Upregulation of the Long Noncoding RNA CASC10 Promotes Cisplatin Resistance in High-Grade Serous Ovarian Cancer

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Identification of Differentially Expressed Genes in Cisplatin-Sensitive versus Cisplatin-Resistant HGSOC Cells
2.2. CASC10 Is Upregulated in Ovarian Cancer Patients and Cisplatin-Resistant Ovarian Cancer Cells
2.3. CASC10 siRNA-Mediated Knockdown Reduced Cell Growth, Invasion, and Viability in Ovarian Cancer Cells
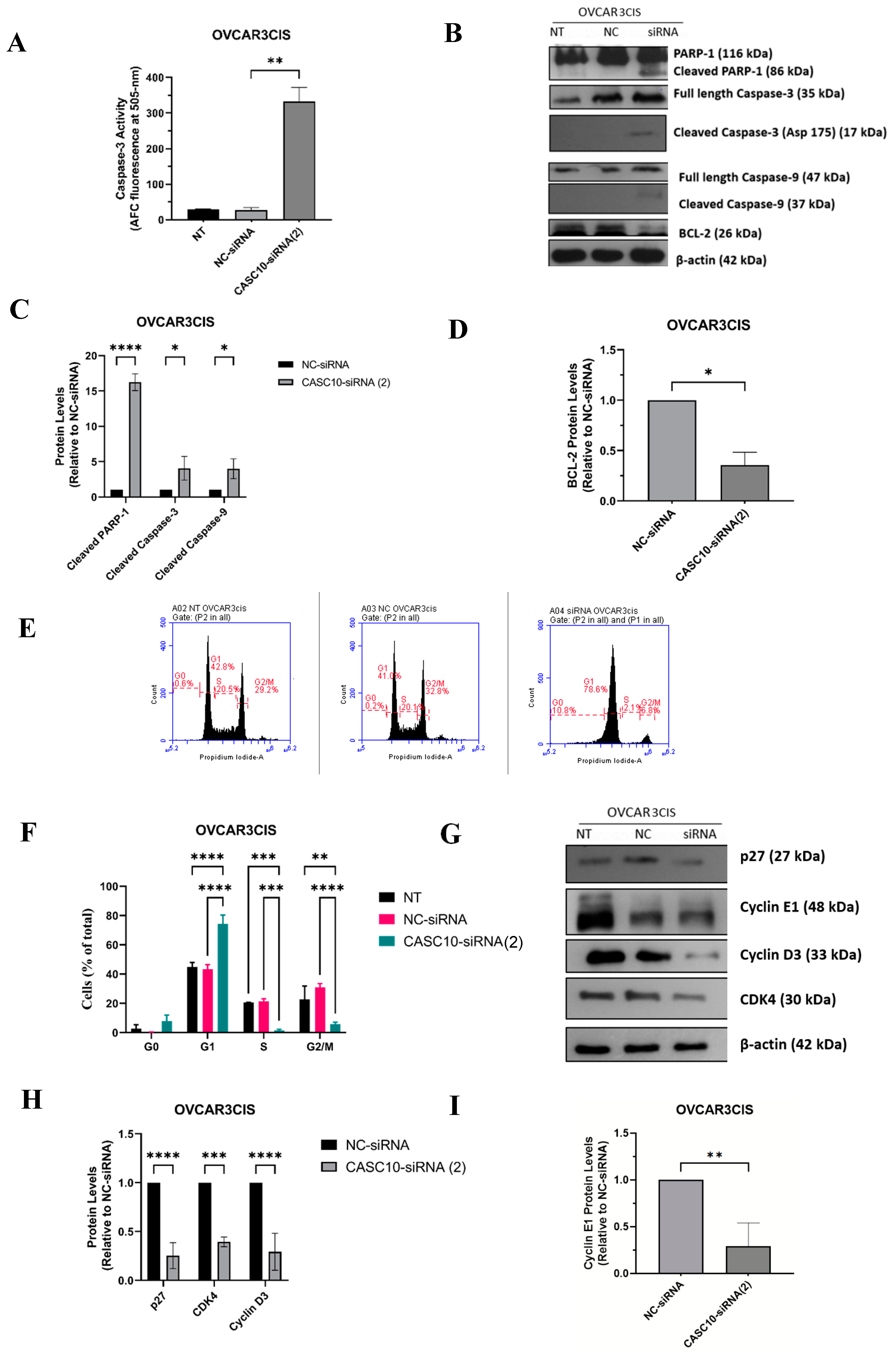
2.4. CASC10 siRNA-Mediated Knockdown Induced Apoptosis and Cell Cycle Arrest
2.5. In Vivo Targeting of CASC10 with Liposome-Encapsulated siRNAs
2.6. Downstream Effectors of CASC10 in HGSOC Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. RNA-Seq and Data Analysis of HGSOC Cells
4.3. Western Blot Analysis
4.4. Small-Interfering RNA (siRNA) and In-Vitro Transfection
4.5. Cell Growth and Cell Viability
4.6. Cell Invasion
4.7. Caspase-3 Activity
4.8. Kaplan-Meier Survival Analysis
4.9. KEGG Pathway Enrichment, Gene Ontology, and Network Analysis
4.10. SYBR-Green Based qRT-PCR
4.11. Tumor Implantation and Drug Treatment
4.12. RNA-Seq Analysis of CASC10-siRNA Transfected Cells
4.13. Statistical Analysis
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef]
- Lisio, M.A.; Fu, L.; Goyeneche, A.; Gao, Z.H.; Telleria, C. High-Grade Serous Ovarian Cancer: Basic Sciences, Clinical and Therapeutic Standpoints. Int. J. Mol. Sci. 2019, 20, 952. [Google Scholar] [CrossRef] [Green Version]
- Vasey, P.A. Resistance to Chemotherapy in Advanced Ovarian Cancer: Mechanisms and Current Strategies. Br. J. Cancer 2003, 89, 23–28. [Google Scholar] [CrossRef]
- Dasari, S.; Bernard Tchounwou, P. Cisplatin in Cancer Therapy: Molecular Mechanisms of Action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef] [Green Version]
- Beck, D.J.; Brubaker, R.R. Effect of Cis Platinum (II) Diamminodichloride on Wild Type and Deoxyribonucleic Acid Repair Deficient Mutants of Escherichia Coli. J. Bacteriol. 1973, 116, 1247–1252. [Google Scholar] [CrossRef] [Green Version]
- Fumarola, S.; Cecati, M.; Sartini, D.; Ferretti, G.; Milanese, G.; Galosi, A.B.; Pozzi, V.; Campagna, R.; Morresi, C.; Emanuelli, M.; et al. Bladder Cancer Chemosensitivity Is Affected by Paraoxonase-2 Expression. Antioxidants 2020, 9, 175. [Google Scholar] [CrossRef] [Green Version]
- Saad, S.Y.; Najjar, T.A.O.; Alashari, M. Role of Non-Selective Adenosine Receptor Blockade and Phosphodiesterase Inhibition in Cisplatin-Induced Nephrogonadal Toxicity in Rats. Clin. Exp. Pharmacol. Physiol. 2004, 31, 862–867. [Google Scholar] [CrossRef]
- Van Zyl, B.; Tang, D.; Bowden, N.A. Biomarkers of Platinum Resistance in Ovarian Cancer: What Can We Use to Improve Treatment. Endocr. Relat. Cancer 2018, 25, R303–R318. [Google Scholar] [CrossRef]
- Surowiak, P.; Materna, V.; Kaplenko, I.; Spaczyński, M.; Dietel, M.; Lage, H.; Zabel, M. Augmented Expression of Metallothionein and Glutathione S-Transferase Pi as Unfavourable Prognostic Factors in Cisplatin-Treated Ovarian Cancer Patients. Virchows Arch. 2005, 447, 626–633. [Google Scholar] [CrossRef]
- Vaezi, A.; Wang, X.Z.; Buch, S.; Gooding, W.; Wang, L.; Seethala, R.R.; Weaver, D.T.; D’Andrea, A.D.; Argiris, A.; Romkes, M.; et al. XPF Expression Correlates with Clinical Outcome in Squamous Cell Carcinoma of the Head and Neck. Clin. Cancer Res. 2011, 17, 5513–5522. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Qing, Y.; Guan, W.; Li, M.; Peng, Y.; Zhang, S.; Xiong, Y.; Wang, D. Predictive Value of APE1, BRCA1, ERCC1 and TUBB3 Expression in Patients with Advanced Non-Small Cell Lung Cancer (NSCLC) Receiving First-Line Platinum-Paclitaxel Chemotherapy. Cancer Chemother. Pharmacol. 2014, 74, 777–786. [Google Scholar] [CrossRef]
- Cocetta, V.; Ragazzi, E.; Montopoli, M. Links between Cancer Metabolism and Cisplatin Resistance, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2020; Volume 354. [Google Scholar]
- Wu, H.J.; Cao, Y.; Weng, D.; Xing, H.; Song, X.; Zhou, J.; Xu, G.; Lu, Y.; Wang, S.; Ma, D. Effect of Tumor Suppressor Gene PTEN on the Resistance to Cisplatin in Human Ovarian Cancer Cell Lines and Related Mechanisms. Cancer Lett. 2008, 271, 260–271. [Google Scholar] [CrossRef]
- Moghbeli, M. MicroRNAs as the Critical Regulators of Cisplatin Resistance in Ovarian Cancer Cells. J. Ovarian Res. 2021, 14, 127. [Google Scholar] [CrossRef]
- Abildgaard, C.; Do Canto, L.M.; Steffensen, K.D.; Rogatto, S.R. Long Non-Coding RNAs Involved in Resistance to Chemotherapy in Ovarian Cancer. Front. Oncol. 2020, 9, 1549. [Google Scholar] [CrossRef] [Green Version]
- Santana-Rivera, Y.; Rabelo-Fernández, R.J.; Quiñones-Díaz, B.I.; Grafals-Ruíz, N.; Santiago-Sánchez, G.; Lozada-Delgado, E.L.; Echevarría-Vargas, I.M.; Apiz, J.; Soto, D.; Rosado, A.; et al. Reduced Expression of Enolase-1 Correlates with High Intracellular Glucose Levels and Increased Senescence in Cisplatin-Resistant Ovarian Cancer Cells. Am. J. Transl. Res. 2020, 12, 1275–1292. [Google Scholar]
- Parker, R.J.; Eastman, A.; BostickBruton, F.; Reed, E. Acquired Cisplatin Resistance in Human Ovarian Cancer Cells Is Associated with Enhanced Repair of Cisplatin-DNA Lesions and Reduced Drug Accumulation. J. Clin. Investig. 1991, 87, 772–777. [Google Scholar] [CrossRef] [Green Version]
- Barnes, B.M.; Nelson, L.; Tighe, A.; Burghel, G.J.; Lin, I.H.; Desai, S.; McGrail, J.C.; Morgan, R.D.; Taylor, S.S. Distinct Transcriptional Programs Stratify Ovarian Cancer Cell Lines into the Five Major Histological Subtypes. Genome Med. 2021, 13, 140. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Reyes-gonzález, J.M.; Armaiz-peña, G.N.; Mangala, L.S.; Ivan, C.; Pradeep, S.; Echevarría-vargas, I.M.; Rivera-, A.; Sood, A.K.; Vivas-mejía, P.E. Targeting C-MYC in Platinum-Resistant Ovarian Cancer. Mol. Cancer Ther. 2016, 14, 2260–2269. [Google Scholar] [CrossRef] [Green Version]
- Dobbins, S.E.; Broderick, P.; Melin, B.; Feychting, M.; Andersson, U.; Brännström, T.; Schramm, J.; Olver, B.; Lloyd, A.; Ma, Y.P.; et al. Europe PMC Funders Group Common Variation at 10p12.31 near MLLT10 Influences Meningioma Risk. Nat. Gen. 2016, 43, 825–827. [Google Scholar] [CrossRef]
- Mas-Ponte, D.; Carlevaro-Fita, J.; Palumbo, E.; Pulido, T.H.; Guigo, R.; Johnson, R. LncATLAS Database for Subcellular Localization of Long Noncoding RNAs. RNA 2017, 23, 1080–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape Provides a Biologist-Oriented Resource for the Analysis of Systems-Level Datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, G.; Taylor, J.A.; Woolbright, B.L. Natural Products as a Means of Overcoming Cisplatin Chemoresistance in Bladder Cancer. Cancer Drug Resist. 2021, 4, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Smith, M.; Halder, J.B.; Meltzer, P.S.; Gonda, T.A.; Mangala, L.S.; Rupaimoole, R.; Lu, C.; Nagaraja, A.S.; Gharpure, K.M.; Kang, Y.; et al. ATP11B Mediates Platinum Resistance in Ovarian Cancer. J. Clin. Investig. 2018, 128, 3199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Ding, Y.; Nie, Y.; Yang, M. EMP1 Promotes the Proliferation and Invasion of Ovarian Cancer Cells through Activating the MAPK Pathway. OncoTargets Ther. 2020, 13, 2047–2055. [Google Scholar] [CrossRef] [Green Version]
- Bignotti, E.; Tassi, R.A.; Calza, S.; Ravaggi, A.; Bandiera, E.; Rossi, E.; Donzelli, C.; Pasinetti, B.; Pecorelli, S.; Santin, A.D. Gene Expression Profile of Ovarian Serous Papillary Carcinomas: Identification of Metastasis-Associated Genes. Am. J. Obstet. Gynecol. 2007, 196, 245.e1–245.e11. [Google Scholar] [CrossRef]
- Necsulea, A.; Soumillon, M.; Warnefors, M.; Liechti, A.; Daish, T.; Zeller, U.; Baker, J.C.; Grützner, F.; Kaessmann, H. The Evolution of LncRNA Repertoires and Expression Patterns in Tetrapods. Nature 2014, 505, 635–640. [Google Scholar] [CrossRef]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. The GENCODE v7 Catalog of Human Long Noncoding RNAs: Analysis of Their Gene Structure, Evolution, and Expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [Green Version]
- Katayama, S.; Tomaru, Y.; Kasukawa, T.; Waki, K.; Nakanishi, M.; Nakamura, M.; Nishida, H.; Yap, C.C.; Suzuki, M.; Kawai, J.; et al. Molecular Biology: Antisense Transcription in the Mammalian Transcriptome. Science 2005, 309, 1564–1566. [Google Scholar] [CrossRef]
- Ulitsky, I.; Bartel, D.P. XLincRNAs: Genomics, Evolution, and Mechanisms. Cell 2013, 154, 26. [Google Scholar] [CrossRef] [Green Version]
- Kelemen, E.; Danis, J.; Göblös, A.; Bata-Csörgő, Z.; Széll, M. Exosomal Long Non-Coding RNAs as Biomarkers in Human Diseases. Electron. J. Int. Fed. Clin. Chem. Lab. Med. 2019, 30, 224–236. [Google Scholar]
- Gutschner, T.; Diederichs, S. The Hallmarks of Cancer: A Long Non-Coding RNA Point of View. RNA Biol. 2012, 9, 703–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Han, L.; Zhou, L.; Wang, L.; Zhang, L.M. Prediction of Candidate RNA Signatures for Recurrent Ovarian Cancer Prognosis by the Construction of an Integrated Competing Endogenous RNA Network. Oncol. Rep. 2018, 40, 2659–2673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Gao, S.; Zheng, Y.; Yao, M.; Ruan, F. LncRNA CASC15 Functions as an Unfavorable Predictor of Ovarian Cancer Prognosis and Inhibits Tumor Progression through Regulation of Mir-221/Arid1a Axis. Onco. Targets. Ther. 2019, 12, 8725–8736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, S.; Li, H.F.; Yang, X.Y.; Guo, S.; Guo, Y.; Chu, L.; Xu, M.J.; Xin, D.M. Higher LncRNA CASC15 Expression Predicts Poor Prognosis and Associates with Tumor Growth in Cervical Cancer. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 507–512. [Google Scholar] [CrossRef]
- Hsu, W.; Liu, L.; Chen, X.; Zhang, Y.; Zhu, W. LncRNA CASC11 Promotes the Cervical Cancer Progression by Activating Wnt/Beta-Catenin Signaling Pathway. Biol. Res. 2019, 52, 33. [Google Scholar] [CrossRef]
- Luo, K.; Geng, J.; Zhang, Q.; Xu, Y.; Zhou, X.; Huang, Z.; Shi, K.Q.; Pan, C.; Wu, J. LncRNA CASC9 Interacts with CPSF3 to Regulate TGF-β Signaling in Colorectal Cancer. J. Exp. Clin. Cancer Res. 2019, 38, 249. [Google Scholar] [CrossRef] [Green Version]
- Dai, W.; Mu, L.; Cui, Y.; Li, Y.; Chen, P.; Xie, H.; Wang, X. Berberine Promotes Apoptosis of Colorectal Cancer via Regulation of the Long Non-Coding RNA (LncRNA) Cancer Susceptibility Candidate 2 (CASC2)/AU-Binding Factor 1 (AUF1)/B-Cell CLL/ Lymphoma 2 (Bcl-2) Axis. Med. Sci. Monit. 2019, 25, 730–738. [Google Scholar] [CrossRef]
- Cuadrado, M.; Gutierrez-Martinez, P.; Swat, A.; Nebreda, A.R.; Fernandez-Capetillo, O. P27Kip1 Stabilization Is Essential for the Maintenance of Cell Cycle Arrest in Response to DNA Damage. Cancer Res. 2009, 69, 8726–8732. [Google Scholar] [CrossRef] [Green Version]
- Hiromura, K.; Pippin, J.W.; Fero, M.L.; Roberts, J.M.; Shankland, S.J. Modulation of Apoptosis by the Cyclin-Dependent Kinase Inhibitor P27(Kip1). J. Clin. Investig. 1999, 103, 597–604. [Google Scholar] [CrossRef] [Green Version]
- Levkau, B.; Koyama, H.; Raines, E.W.; Clurman, B.E.; Herren, B.; Orth, K.; Roberts, J.M.; Ross, R. Cleavage of P21Cip1/Waf1 and P27Kip1 Mediates Apoptosis in Endothelial Cells through Activation of Cdk2: Role of a Caspase Cascade. Cell 1998, 1, 553–563. [Google Scholar] [CrossRef]
- Reyes-González, J.M.; Vivas-Mejía, P.E. C-MYC and Epithelial Ovarian Cancer. Front. Oncol. 2021, 11, 601512. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.W.; Wang, Y.; Chen, L.L. Cellular Functions of Long Noncoding RNAs. Nat. Cell Biol. 2019, 21, 542–551. [Google Scholar] [CrossRef]
- Tian, C.; Zhou, S.; Yi, C. High NUP43 Expression Might Independently Predict Poor Overall Survival in Luminal A and in HER2+ Breast Cancer. Futur. Oncol. 2018, 14, 1431–1442. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Tang, D. Patient-Specific Carotid Plaque Progression Simulation. C. Model. Eng. Sci. 2000, 1, 119–131. [Google Scholar]
- Xu, X.; Nie, J.; Lu, L.; Du, C.; Meng, F.; Song, D. LINC00337 Promotes Tumor Angiogenesis in Colorectal Cancer by Recruiting DNMT1, Which Suppresses the Expression of CNN1. Cancer Gene Ther. 2021, 28, 1285–1297. [Google Scholar] [CrossRef]
- Wang, Z.; Li, T.E.; Chen, M.; Pan, J.J.; Shen, K.W. MiR-106b-5p Contributes to the Lung Metastasis of Breast Cancer via Targeting CNN1 and Regulating Rho/ROCK1 Pathway. Aging 2020, 12, 1867–1887. [Google Scholar] [CrossRef]
- Echevarría-Vargas, I.M.; Valiyeva, F.; Vivas-Mejía, P.E. Upregulation of MiR-21 in Cisplatin Resistant Ovarian Cancer via JNK-1/c-Jun Pathway. PLoS ONE 2014, 9, e970194. [Google Scholar] [CrossRef]
- Báez-Vega, P.M.; Vargas, I.M.E.; Valiyeva, F.; Rosado, J.E.; Roman, A.; Flores, J.; Marcos-Martínez, M.J.; Vivas-Mejía, P.E. Targeting MiR-21-3p Inhibits Proliferation and Invasion of Ovarian Cancer Cells. Oncotarget 2016, 7, 36321–36337. [Google Scholar] [CrossRef]
- Gyorffy, B.; Lánczky, A.; Szállási, Z. Implementing an Online Tool for Genomewide Validation of Survival-Associated Biomarkers in Ovarian-Cancer Using Microarray Data from 1287 Patients. Endocr. Relat. Cancer 2012, 19, 197–208. [Google Scholar] [CrossRef] [Green Version]







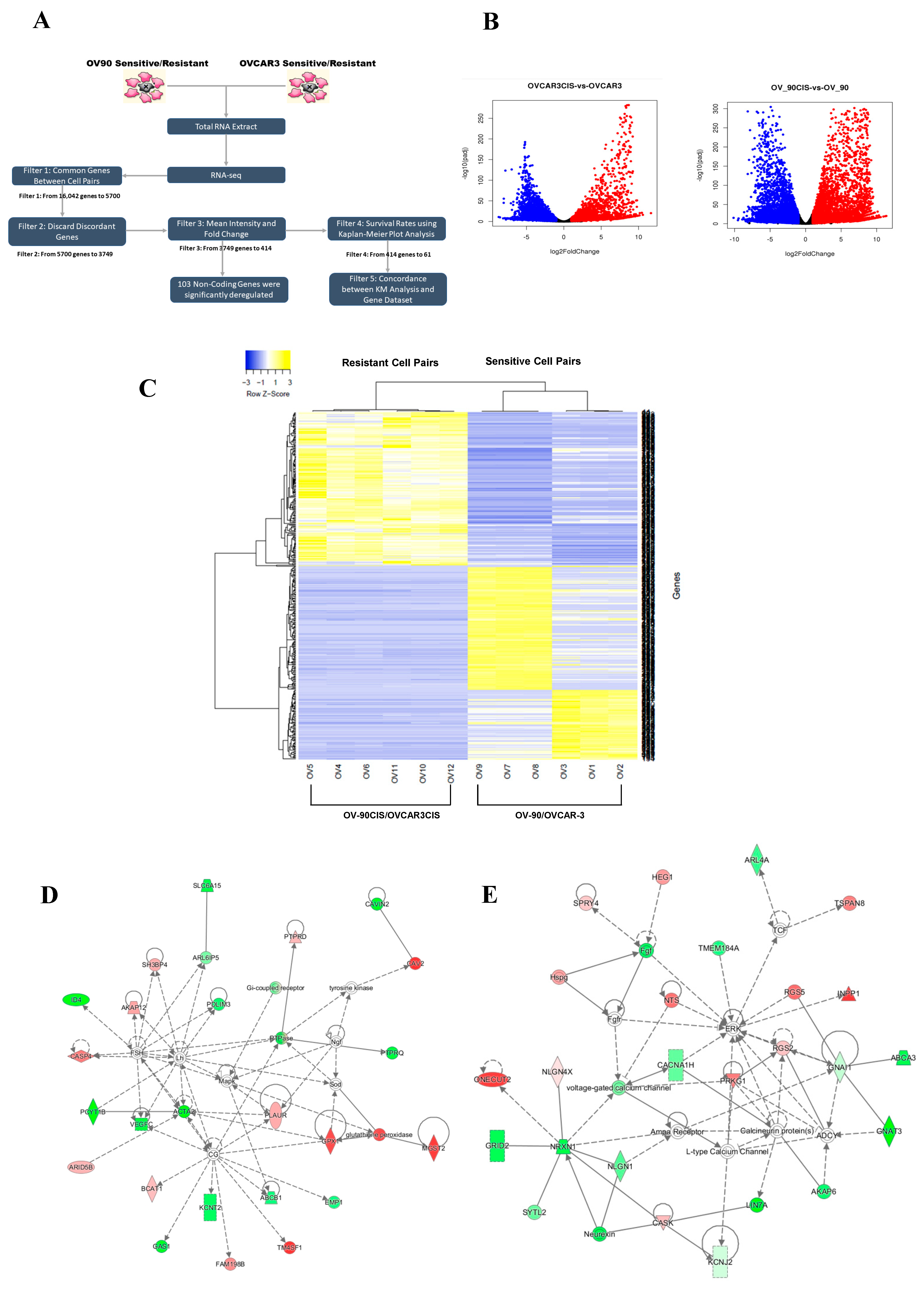{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | qPCR Log2FC | RNA-Seq Log2FC |
|---|---|---|
| PDLIM3 | 13.16 | 5.56 |
| TPH2 | 11.53 | 8.04 |
| TRHDE | 10.67 | 7.82 |
| KCNT2 | 10.47 | 5.09 |
| GAS1 | 10.24 | 5.01 |
| CLDN16 | 9.33 | 5.29 |
| PRRX1 | 9.09 | 4.31 |
| SESN3 | 9.07 | 3.41 |
| GALNT13 | 8.50 | 4.78 |
| SYTL2 | 7.63 | 4.55 |
| SULF1 | 7.43 | 3.78 |
| BCHE | 6.50 | 1.31 |
| ATP11B | 6.44 | 3.24 |
| SLIT2 | 5.33 | 2.09 |
| SLC6A15 | 5.29 | 5.45 |
| SH2D5 | 5.19 | 2.51 |
| VEGFC | 4.85 | 4.84 |
| CFL2 | 4.42 | 1.98 |
| ARRDC3 | 4.17 | 2.24 |
| CRABP2 | 3.43 | 1.62 |
| NUDT11 | 3.34 | 2.13 |
| EYA4 | 2.74 | 0.58 |
| SACS | 2.55 | 1.03 |
| KIF1A | 2.36 | 1.43 |
| ARHGAP28 | 2.19 | 1.24 |
| EMP1 | 2.11 | 1.20 |
| DDAH1 | 2.09 | 0.96 |
| CASC10 | 2.06 | 0.50 |
| LRRC17 | −0.94 | −1.42 |
| LRG1 | −0.97 | −2.72 |
| TMCC3 | −1.03 | −2.25 |
| SPRY4 | −1.67 | −1.91 |
| F2R | −2.09 | −3.80 |
| SULT1A1 | −2.67 | −3.43 |
| PBX1 | −2.78 | −2.29 |
| SLC7A2 | −2.85 | −5.47 |
| MXRA8 | −3.16 | −4.14 |
| NID1 | −3.36 | −4.50 |
| RUNX1 | −3.59 | −1.64 |
| PROCR | −4.28 | −5.77 |
| LIPG | −5.20 | −5.68 |
| PDE1A | −5.76 | −7.53 |
| PTPRD | −6.38 | −7.88 |
| VCAN | −6.60 | −5.64 |
| THBS1 | −9.79 | −4.06 |
| Gene Symbol | Gene Name | Biological Role | p-Value |
|---|---|---|---|
| Upregulated | |||
| RTN4R | Reticulon 4 receptor | Receptor-mediated axonal growth inhibition | 4.59 × 10−21 |
| MACF1 | Microtubule actin crosslinking factor 1 | Actin-microtubule interactions | 2.83 × 10−12 |
| PYM1 | PYM homolog 1 | Nuclear-transcribed mRNA catabolic process and regulation of translation | 5.24 × 10−20 |
| CNN1 | Cellular communication network factor 1 | Regulation of vascular-associated smooth muscle cell proliferation | 7.38 × 10−6 |
| TGFBRAP1 | Transforming growth factor beta receptor associated protein 1 | TGF-beta signaling and association to SMAD4 | 3.32 × 10−39 |
| Downregulated | |||
| NUP43 | Nucleoporin 43 | Transport of macromolecules between cytoplasm and nucleus | 1.69 × 10−86 |
| FHL1 | Four and a half LIM domains 1 | Assembly of sarcomeres and muscle contraction regulation | 2.49 × 10−11 |
| DHFR2 | Dihydrofolate reductase 2 | Tetrahydrofolate metabolic process and thymidine biosynthesis | 1.21 × 10−10 |
| MIR1915HG | microRNA1915 host gene | Long Noncoding RNA | 2.76 × 10−59 |
| NDUFA7 | NADH:ubiquinone oxidoreductase subunit A7 | Member of complex I in the electron transport chain | 7.76 × 10−29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noriega-Rivera, R.; Rivera-Serrano, M.; Rabelo-Fernandez, R.J.; Pérez-Santiago, J.; Valiyeva, F.; Vivas-Mejía, P.E. Upregulation of the Long Noncoding RNA CASC10 Promotes Cisplatin Resistance in High-Grade Serous Ovarian Cancer. Int. J. Mol. Sci. 2022, 23, 7737. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23147737
Noriega-Rivera R, Rivera-Serrano M, Rabelo-Fernandez RJ, Pérez-Santiago J, Valiyeva F, Vivas-Mejía PE. Upregulation of the Long Noncoding RNA CASC10 Promotes Cisplatin Resistance in High-Grade Serous Ovarian Cancer. International Journal of Molecular Sciences. 2022; 23(14):7737. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23147737
Chicago/Turabian StyleNoriega-Rivera, Ricardo, Mariela Rivera-Serrano, Robert J. Rabelo-Fernandez, Josué Pérez-Santiago, Fatima Valiyeva, and Pablo E. Vivas-Mejía. 2022. "Upregulation of the Long Noncoding RNA CASC10 Promotes Cisplatin Resistance in High-Grade Serous Ovarian Cancer" International Journal of Molecular Sciences 23, no. 14: 7737. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23147737