
Natural Products of Marine Origin for the Treatment of Colorectal and Pancreatic Cancers: Mechanisms and Potential


Abstract
:1. Introduction
2. Marine Natural Products (MNPs) with the Potential to Treat Cancer
3. Mechanisms of Action of MNPs in CRC and PC
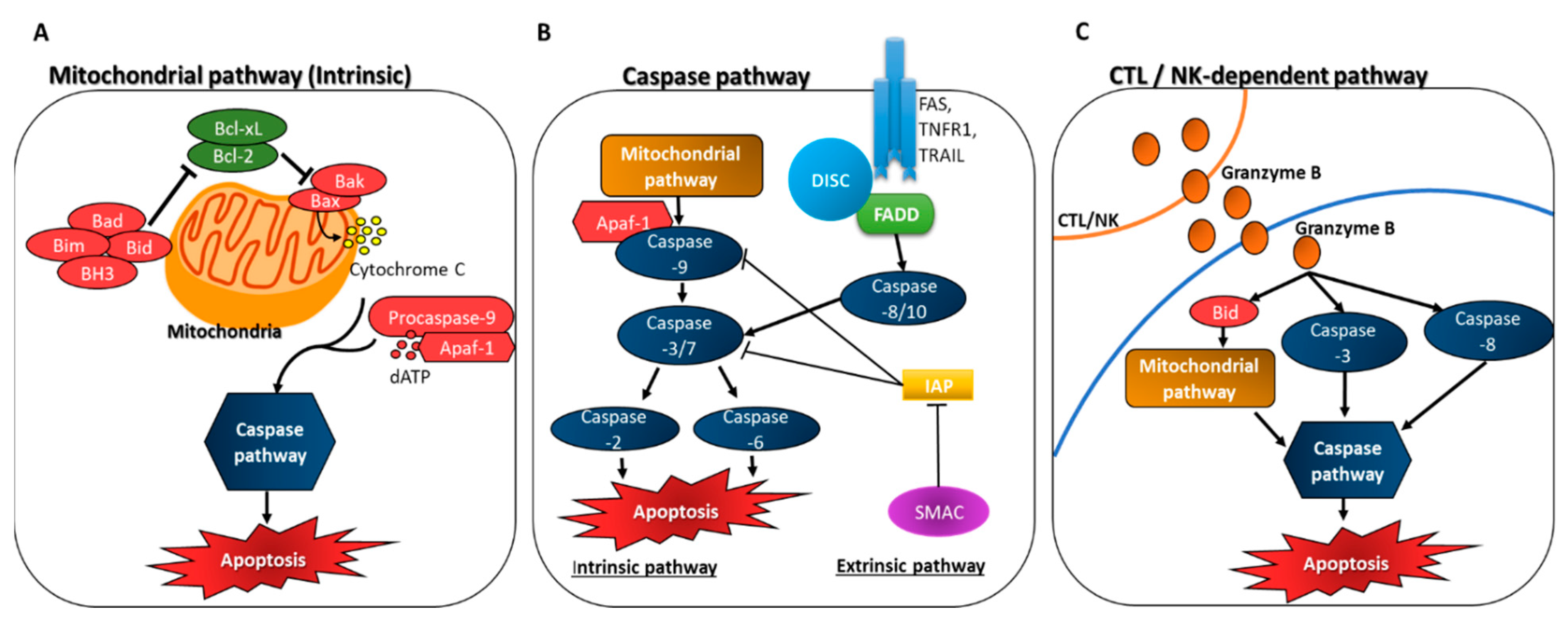
3.1. Induction of Apoptosis through Caspase Activation
3.1.1. Mitochondrial Apoptosome-Driven Pathway of Caspase Activation (Intrinsic)
3.1.2. Death Receptor Pathway of Caspase Activation (Extrinsic)
3.1.3. CTL/NK-Dependent Pathway
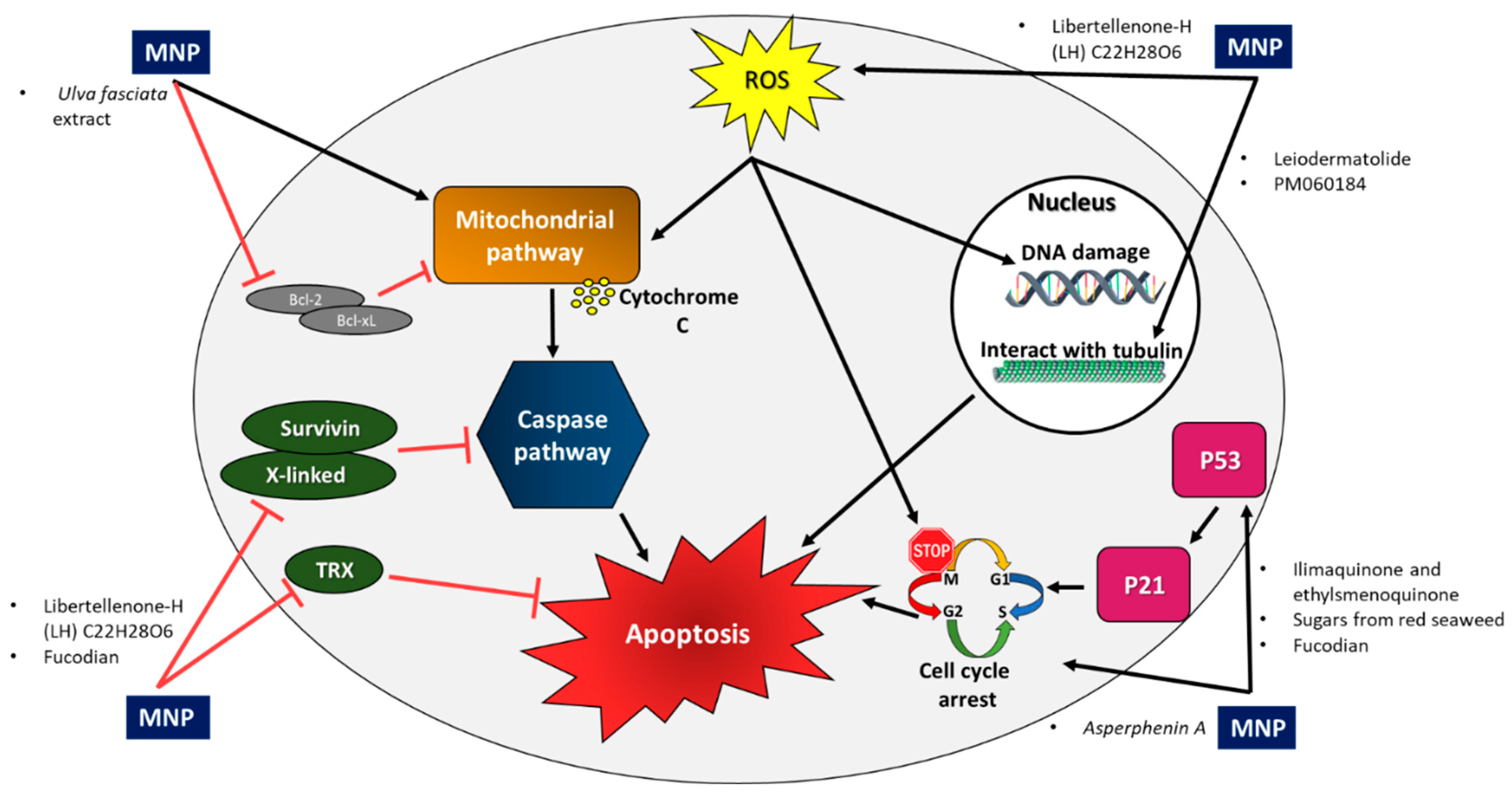
3.2. Inhibition of Anti-Apoptotic Factors
3.3. Interaction of MNPs with Tubulin to Cause Anti-Mitotic Activity
3.4. Suppression of Cell Cycle Progression
3.5. The Role of NFκB and p53 in Apoptosis
3.6. Increased Intracellular ROS Accumulation and Induction of Apoptosis
4. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA. Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, Y.; Xu, P. Global Colorectal Cancer Burden in 2020 and Projections to 2040. Transl. Oncol. 2021, 14, 101174. [Google Scholar] [CrossRef] [PubMed]
- NIH National Cancer Institute. Pancreatic Cancer-Cancer Stat Facts. Available online: https://seer.cancer.gov/statfacts/html/pancreas.html (accessed on 13 March 2022).
- Rawla, P.; Sunkara, T.; Gaduputi, V. Epidemiology of Pancreatic Cancer: Global Trends, Etiology and Risk Factors. World J. Oncol. 2019, 10, 10. [Google Scholar] [CrossRef]
- Klein, A.P. Pancreatic Cancer Epidemiology: Understanding the Role of Lifestyle and Inherited Risk Factors. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 493–502. [Google Scholar] [CrossRef]
- Cancer.Net. Colorectal Cancer: Types of Treatment. Available online: https://www.cancer.net/cancer-types/colorectal-cancer/types-treatment (accessed on 13 March 2022).
- Pancreatic Cancer: Types of Treatment. Available online: https://www.cancer.net/cancer-types/pancreatic-cancer/types-treatment (accessed on 26 April 2022).
- Aslam, M.S.; Naveed, S.; Ahmed, A.; Abbas, Z.; Gull, I.; Athar, M.A. Side Effects of Chemotherapy in Cancer Patients and Evaluation of Patients Opinion about Starvation Based Differential Chemotherapy. J. Cancer Ther. 2014, 2014, 817–822. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.B.; Gupta, R. Drug Development from Natural Resource: A Systematic Approach. Mini Rev. Med. Chem. 2015, 15, 52–57. [Google Scholar] [CrossRef]
- Blunt, J.W.; Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine Natural Products. Nat. Prod. Rep. 2018, 35, 8–53. [Google Scholar] [CrossRef] [Green Version]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine Natural Products. Nat. Prod. Rep. 2016, 33, 382–431. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Ye, K.; Jiang, S.; Zhou, G. Marine Power on Cancer: Drugs, Lead Compounds, and Mechanisms. Mar. Drugs 2021, 19, 488. [Google Scholar] [CrossRef]
- Parate, S.; Kumar, V.; Lee, G.; Rampogu, S.; Hong, J.C.; Lee, K.W. Marine-Derived Natural Products as ATP-Competitive MTOR Kinase Inhibitors for Cancer Therapeutics. Pharmaceuticals 2021, 14, 282. [Google Scholar] [CrossRef] [PubMed]
- Mbaoji, F.N.; Nweze, J.A.; Yang, L.; Huang, Y.; Huang, S.; Onwuka, A.M.; Peter, I.E.; Mbaoji, C.C.; Jiang, M.; Zhang, Y.; et al. Novel Marine Secondary Metabolites Worthy of Development as Anticancer Agents: A Review. Molecules 2021, 26, 5769. [Google Scholar] [CrossRef] [PubMed]
- Zuo, W.; Kwok, H.F. Development of Marine-Derived Compounds for Cancer Therapy. Mar. Drugs 2021, 19, 342. [Google Scholar] [CrossRef] [PubMed]
- Gogineni, V.; Hamann, M.T. Marine Natural Product Peptides with Therapeutic Potential: Chemistry, Biosynthesis, and Pharmacology. Biochim. Biophys. Acta-Gen. Subj. 2018, 1862, 81–196. [Google Scholar] [CrossRef]
- Lee, H.; Selvaraj, B.; Lee, J.W. Anticancer Effects of Seaweed-Derived Bioactive Compounds. Appl. Sci. 2021, 11, 11261. [Google Scholar] [CrossRef]
- Wang, S.C.; Li, R.N.; Lin, L.C.; Tang, J.Y.; Su, J.H.; Sheu, J.H.; Chang, H.W. Comparison of Antioxidant and Anticancer Properties of Soft Coral-Derived Sinularin and Dihydrosinularin. Molecules 2021, 26, 3853. [Google Scholar] [CrossRef]
- Guzmán, E.A.; Pitts, T.P.; Diaz, M.C.; Wright, A.E. The Marine Natural Product Scalarin Inhibits the Receptor for Advanced Glycation End Products (RAGE) and Autophagy in the PANC-1 and MIA PaCa-2 Pancreatic Cancer Cell Lines. Investig. New Drugs 2019, 37, 262–270. [Google Scholar] [CrossRef]
- Crews, C.M.; Collins, J.L.; Lane, W.S.; Snapper, M.L.; Schreiber, S.L. GTP-Dependent Binding of the Antiproliferative Agent Didemnin to Elongation Factor 1 Alpha. J. Biol. Chem. 1994, 269, 15411–15414. [Google Scholar] [CrossRef]
- Chun, H.G.; Davies, B.; Hoth, D.; Suffness, M.; Plowman, J.; Flora, K.; Grieshaber, C.; Leyland-Jones, B.; Didemnin, B. The First Marine Compound Entering Clinical Trials as an Antineoplastic Agent. Investig. New Drugs 1986, 4, 279–284. [Google Scholar] [CrossRef]
- Simmons, T.L.; Andrianasolo, E.; McPhail, K.; Flatt, P.; Gerwick, W.H. Marine Natural Products as Anticancer Drugs. Mol. Cancer Ther. 2005, 4, 333–342. [Google Scholar]
- Schyschka, L.; Rudy, A.; Jeremias, I.; Barth, N.; Pettit, G.R.; Vollmar, A.M. Spongistatin 1: A New Chemosensitizing Marine Compound That Degrades XIAP. Leukemia 2008, 22, 1737–1745. [Google Scholar] [CrossRef] [PubMed]
- Fenical, W.; Jensen, P.R.; Palladino, M.A.; Lam, K.S.; Lloyd, G.K.; Potts, B.C. Discovery and Development of the Anticancer Agent Salinosporamide A (NPI-0052). Bioorg. Med. Chem. 2009, 17, 2175–2180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bross, P.F.; Kane, R.; Farrell, A.T.; Abraham, S.; Benson, K.; Brower, M.E.; Bradley, S.; Gobburu, J.V.; Goheer, A.; Lee, S.L.; et al. Approval Summary for Bortezomib for Injection in the Treatment of Multiple Myeloma. Clin. Cancer Res. 2004, 10, 3954–3964. [Google Scholar] [CrossRef] [Green Version]
- Richardson, P.G.; Barlogie, B.; Berenson, J.; Singhal, S.; Jagannath, S.; Irwin, D.; Rajkumar, S.V.; Srkalovic, G.; Alsina, M.; Alexanian, R.; et al. A Phase 2 Study of Bortezomib in Relapsed, Refractory Myeloma. N. Engl. J. Med. 2003, 348, 2609–2617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, E.; Sorolla, M.A.; Krishnan, P.D.G.; Sorolla, A. From Seabed to Bedside: A Review on Promising Marine Anticancer Compounds. Biomolecules 2020, 10, 248. [Google Scholar] [CrossRef] [Green Version]
- Dyshlovoy, S.A.; Honecker, F. Marine Compounds and Cancer: Updates 2020. Mar. Drugs 2020, 18, 643. [Google Scholar] [CrossRef]
- Wu, A.C.; Jelielek, K.K.; Le, H.Q.; Butt, M.; Newman, D.J.; Glaser, K.B.; Pierce, M.L.; Mayer, A.M. The 2021 Marine Pharmacology and Pharmaceuticals Pipeline. FASEB J. 2022, 36, L7586. [Google Scholar] [CrossRef]
- Li, T.; Wang, N.; Zhang, T.; Zhang, B.; Sajeevan, T.P.; Joseph, V.; Armstrong, L.; He, S.; Yan, X.; Benjamin Naman, C. A Systematic Review of Recently Reported Marine Derived Natural Product Kinase Inhibitors. Mar. Drugs 2019, 17, 493. [Google Scholar] [CrossRef] [Green Version]
- Khalifa, S.A.M.; Elias, N.; Farag, M.A.; Chen, L.; Saeed, A.; Hegazy, M.E.F.; Moustafa, M.S.; El-Wahed, A.A.; Al-Mousawi, S.M.; Musharraf, S.G.; et al. Marine Natural Products: A Source of Novel Anticancer Drugs. Mar. Drugs 2019, 17, 491. [Google Scholar] [CrossRef] [Green Version]
- Sarabia, F.; Han, N.; Li, J.; Li, X. Natural Marine Products: Anti-Colorectal Cancer In Vitro and In Vivo. Mar. Drugs 2022, 20, 349. [Google Scholar] [CrossRef]
- Gao, D.; Wang, Y.; Xie, W.; Yang, T.; Jiang, Y.; Guo, Y.; Guan, J.; Liu, H. Metabolomics Study on the Antitumor Effect of Marine Natural Compound Flexibilide in HCT-116 Colon Cancer Cell Line. J. Chromatogr. B. Analyt. Technol. Biomed. Life Sci. 2016, 1014, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Taira, J.; Miyazato, H.; Ueda, K. Marine Peroxy Sesquiterpenoids Induce Apoptosis by Modulation of Nrf2-ARE Signaling in HCT116 Colon Cancer Cells. Mar. Drugs 2018, 16, 347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Torres, V.; Rodríguez-Pérez, C.; Herranz-López, M.; Martín-García, B.; Gómez-Caravaca, A.-M.; Arráez-Román, D.; Segura-Carretero, A.; Barrajón-Catalán, E.; Micol, V. Marine Invertebrate Extracts Induce Colon Cancer Cell Death via ROS-Mediated DNA Oxidative Damage and Mitochondrial Impairment. Biomolecules 2019, 9, 771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esmaeelian, B.; Benkendorff, K.; Le Leu, R.K.; Abbott, C.A. Simultaneous Assessment of the Efficacy and Toxicity of Marine Mollusc–Derived Brominated Indoles in an in Vivo Model for Early Stage Colon Cancer. Integr. Cancer Ther. 2018, 17, 248–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Díez, M.; Guillén-Navarro, M.; Pera, B.; Bouchet, B.; Martínez-Leal, J.; Barasoain, I.; Cuevas, C.; Andreu, J.; García-Fernández, L.; Díaz, J.; et al. PM060184, a New Tubulin Binding Agent with Potent Antitumor Activity Including P-Glycoprotein over-Expressing Tumors. Biochem. Pharmacol. 2014, 88, 291–302. [Google Scholar] [CrossRef]
- Aravindan, S.; Somasundaram, D.B.; Somasundaram, S.T.; Natarajan, M.; Herman, T.S.; Aravindan, N. Targeting Acquired Oncogenic Burden in Resilient Pancreatic Cancer: A Novel Benefit from Marine Polyphenols. Mol. Cell. Biochem. 2019, 460, 175–193. [Google Scholar] [CrossRef]
- Lee, H.; Chung, K.; Hwang, I.; Gwak, J.; Park, S.; Ju, B.; Yun, E.; Kim, D.; Chung, Y.; Na, M.; et al. Activation of P53 with Ilimaquinone and Ethylsmenoquinone, Marine Sponge Metabolites, Induces Apoptosis and Autophagy in Colon Cancer Cells. Mar. Drugs 2015, 13, 543–557. [Google Scholar] [CrossRef] [Green Version]
- Song, M.; Ou, F.-S.; Zemla, T.J.; Hull, M.A.; Shi, Q.; Limburg, P.J.; Alberts, S.R.; Sinicrope, F.A.; Giovannucci, E.L.; Van Blarigan, E.L.; et al. Marine Omega-3 Fatty Acid Intake and Survival of Stage III Colon Cancer According to Tumor Molecular Markers in NCCTG Phase III Trial N0147 (Alliance). Wiley Online Libr. 2019, 145, 380–389. [Google Scholar] [CrossRef]
- Yun, E.; Yu, S.; Kim, Y.; Liu, J.; Kang, N.; Jin, Y.; Kim, K. In Vitro Prebiotic and Anti-Colon Cancer Activities of Agar-Derived Sugars from Red Seaweeds. Mar. Drugs 2021, 19, 213. [Google Scholar] [CrossRef]
- Jimi, K.; Jeonghee, L.; Oh, J.H.; Chang, H.J.; Dae, K.S.; Aesun, S.; Jeongseon, K. Associations among Dietary Seaweed Intake, c-MYC Rs6983267 Polymorphism, and Risk of Colorectal Cancer in a Korean Population: A Case–Control Study. Eur. J. Nutr. 2020, 59, 1963–1974. [Google Scholar] [CrossRef]
- Ryu, M.J.; Kim, A.D.; Kang, K.A.; Chung, H.S.; Kim, H.S.; Suh, I.S.; Chang, W.Y.; Hyun, J.W. The Green Algae Ulva Fasciata Delile Extract Induces Apoptotic Cell Death in Human Colon Cancer Cells. Vitr. Cell. Dev. Biol.-Anim. 2013, 49, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J.; Park, S.Y.; Lee, J.Y.; Park, J.H.Y. Fucoidan Present in Brown Algae Induces Apoptosis of Human Colon Cancer Cells. BMC Gastroenterol. 2010, 10, 96. [Google Scholar] [CrossRef] [Green Version]
- LaCasse, E.C.; Baird, S.; Korneluk, R.G.; MacKenzie, A.E. The Inhibitors of Apoptosis (IAPs) and Their Emerging Role in Cancer. Oncogene 1998, 17, 3247–3259. [Google Scholar]
- Alves, C.; Pinteus, S.; Rodrigues, A.; Horta, A.; Pedrosa, R. Algae from Portuguese Coast Presented High Cytotoxicity and Antiproliferative Effects on an In Vitro Model of Human Colorectal Cancer. Pharmacogn. Res. 2018, 10, 24. [Google Scholar] [CrossRef]
- Bae, S.; Liao, L.; Park, S.; Kim, W.; Shin, J.; Lee, S. Antitumor Activity of Asperphenin A, a Lipopeptidyl Benzophenone from Marine-Derived Aspergillus sp. Fungus, by Inhibiting Tubulin Polymerization in Colon Cancer Cells. Mar. Drugs 2020, 18, 110. [Google Scholar]
- Choi, E.; Park, J.; Kim, Y.; Jung, J.; Lee, J.; Kwon, H.; Yang, H. Apoptosis-inducing Effect of Diketopiperazine Disulfides Produced by Aspergillus sp. KMD 901 Isolated from Marine Sediment on HCT116 Colon Cancer Cell Lines. J. Appl. Microbiol. 2011, 110, 304–313. [Google Scholar] [CrossRef]
- Ding, L.; Bar-Shalom, R.; Aharonovich, D.; Kurisawa, N.; Patial, G.; Li, S.; He, S.; Yan, X.; Iwasaki, A.; Suenaga, K.; et al. Metabolomic Characterization of a Cf. Neolyngbya Cyanobacterium from the South China Sea Reveals Wenchangamide A, a Lipopeptide with In Vitro. Mar. Drugs 2021, 19, 397. [Google Scholar] [CrossRef]
- Freitas, S.; Martins, R.; Campos, A.; Azevedo, J.; Osório, H.; Costa, M.; Barros, P.; Vasconcelos, V.; Urbatzka, R. Insights into the Potential of Picoplanktonic Marine Cyanobacteria Strains for Cancer Therapies–Cytotoxic Mechanisms against the RKO Colon Cancer Cell Line. Toxicon 2016, 119, 140–151. [Google Scholar] [CrossRef]
- Salvador-Reyes, L.A.; Engene, N.; Paul, V.J.; Luesch, H. Targeted Natural Products Discovery from Marine Cyanobacteria Using Combined Phylogenetic and Mass Spectrometric Evaluation. J. Nat. Prod. 2015, 78, 486–492. [Google Scholar] [CrossRef] [Green Version]
- Ratnayake, R.; Gunasekera, S.P.; Ma, J.J.; Dang, L.H.; Carney, T.J.; Paul, V.J.; Luesch, H. Dolastatin 15 from a Marine Cyanobacterium Suppresses HIF-1α Mediated Cancer Cell Viability and Vascularization. ChemBioChem 2020, 21, 2356–2366. [Google Scholar] [CrossRef]
- Guzmán, E.A.; Maers, K.; Roberts, J.; Kemami-Wangun, H.V.; Harmody, D.; Wright, A.E. The Marine Natural Product Microsclerodermin A Is a Novel Inhibitor of the Nuclear Factor Kappa B and Induces Apoptosis in Pancreatic Cancer Cells. Investig. New Drugs 2015, 33, 86–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kallifatidis, G.; Hoepfner, D.; Jaeg, T.; Guzmán, E.A.; Wright, A.E. The Marine Natural Product Manzamine A Targets Vacuolar ATPases and Inhibits Autophagy in Pancreatic Cancer Cells. Mar. Drugs 2013, 11, 3500–3516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzmán, E.A.; Johnson, J.D.; Carrier, M.K.; Meyer, C.I.; Pitts, T.P.; Gunasekera, S.P.; Wright, A.E. Selective Cytotoxic Activity of the Marine Derived Batzelline Compounds against Pancreatic Cancer Cell Lines. Anticancer. Drugs 2009, 20, 149. [Google Scholar] [CrossRef]
- Guzman, E.A.; Xu, Q.; Pitts, T.P.; Mitsuhashi, K.O.; Baker, C.; Linley, P.A.; Oestreicher, J.; Tendyke, K.; Winder, P.L.; Suh, E.M.; et al. Leiodermatolide, a Novel Marine Natural Product, Has Potent Cytotoxic and Antimitotic Activity against Cancer Cells, Appears to Affect Microtubule Dynamics, and Exhibits. Int. J. Cancer 2016, 139, 2116–2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.W.; Yan, Y.; Wang, L.; Wu, D.; Nai, K.Y.; Shi, H.C.; Li, F. Marine Bioactive Compound Dieckol Induces Apoptosis and Inhibits the Growth of Human Pancreatic Cancer Cells PANC-1. J. Biochem. Mol. Toxicol. 2021, 35, e22648. [Google Scholar] [CrossRef]
- Green, D.R. Apoptotic Pathways: Paper Wraps Stone Blunts Scissors. Cell 2000, 102, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Delma, C.R.; Thirugnanasambandan, S.; Srinivasan, G.P.; Raviprakash, N.; Manna, S.K.; Natarajan, M.; Aravindan, N. Fucoidan from Marine Brown Algae Attenuates Pancreatic Cancer Progression by Regulating P53–NFκB Crosstalk. Phytochemistry 2019, 167, 112078. [Google Scholar]
- Zhang, W.; Zhu, Y.; Yu, H.; Liu, X.; Jiao, B.; Lu, X. Libertellenone H, a Natural Pimarane Diterpenoid, Inhibits Thioredoxin System and Induces ROS-Mediated Apoptosis in Human Pancreatic Cancer Cells. Molecules 2021, 26, 315. [Google Scholar] [CrossRef]
- Denicourt, C.; Dowdy, S.F. Targeting Apoptotic Pathways in Cancer Cells. Science 2004, 305, 1411–1413. [Google Scholar] [CrossRef]
- Creagh, E.M.; Conroy, H.; Martin, S.J. Caspase-Activation Pathways in Apoptosis and Immunity. Immunol. Rev. 2003, 193, 10–21. [Google Scholar] [CrossRef]
- Slee, E.A.; Harte, M.T.; Kluck, R.M.; Wolf, B.B.; Casiano, C.A.; Newmeyer, D.D.; Wang, H.-G.; Reed, J.C.; Nicholson, D.W.; Alnemri, E.S.; et al. Ordering the Cytochrome c–Initiated Caspase Cascade: Hierarchical Activation of Caspases-2,-3,-6,-7,-8, and-10 in a Caspase-9–Dependent Manner. J. Cell Biol. 1999, 144, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Adrain, C.; Martin, S. The Mitochondrial Apoptosome: A Killer Unleashed by the Cytochrome Seas. Trends Biochem. Sci. 2001, 26, 390–397. [Google Scholar] [CrossRef]
- Acehan, D.; Jiang, X.; Morgan, D.G.; Heuser, J.E.; Wang, X.; WAkey, C. Three-Dimensional Structure of the Apoptosome: Implications for Assembly, Procaspase-9 Binding, and Activation. Mol. Cell 2002, 9, 423–432. [Google Scholar] [CrossRef]
- Green, D.R.; Reed, J.C. Mitochondria and Apoptosis. Science 1998, 281, 1309–1312. [Google Scholar] [CrossRef]
- van Loo, G.; Saelens, X.; Van Gurp, M.; MacFarlane, M.; Martin, S.J.; Vandenabeele, P. The Role of Mitochondrial Factors in Apoptosis: A Russian Roulette with More than One Bullet. Cell Death Differ. 2002, 9, 1031–1042. [Google Scholar] [CrossRef]
- Froelich, C.J.; Dixit, V.M.; Yang, X. Lymphocyte Granule-Mediated Apoptosis: Matters of Viral Mimicry and Deadly Proteases. Immunol. Today 1998, 19, 30–36. [Google Scholar] [CrossRef]
- Medema, J.P.; Toes, R.E.M.; Scaffidi, C.; Zheng, T.S.; Flavell, R.A.; Melief, C.J.M.; Peter, M.E.; Offringa, R.; Krammer, P.H. Cleavage of FLICE (Caspase-8) by Granzyme B during Cytotoxic T Lymphocyte-Induced Apoptosis. Eur. J. Immunol. 1997, 27, 3492–3498. [Google Scholar] [CrossRef]
- Heibein, J.A.; Goping, I.S.; Barry, M.; Pinkoski, M.J.; Shore, G.C.; Green, D.R.; Bleackley, R.C. Granzyme B–Mediated Cytochrome C Release Is Regulated by the Bcl-2 Family Members Bid and Bax. J. Exp. Med. 2000, 192, 1391–1402. [Google Scholar] [CrossRef] [Green Version]
- Boice, A.; Bouchier-Hayes, L. Targeting Apoptotic Caspases in Cancer. Biochim. Biophys. Acta-Mol. Cell Res. 2020, 1867, 118688. [Google Scholar] [CrossRef]
- Joung, H.Y.; Son, E.; Pyo, S.; Hong, K.L. Novel Sulfated Polysaccharide Derived from Red-Tide Microalga Gyrodinium Impudicum Strain KG03 with Immunostimulating Activity In Vivo. Mar. Biotechnol. 2005, 7, 331–338. [Google Scholar] [CrossRef]
- Deveraux, Q.L.; Reed, J.C. IAP Family Proteins—Suppressors of Apoptosis. Genes Dev. 1999, 13, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Holmgren, A. The Thioredoxin Antioxidant System. Free Radic. Biol. Med. 2014, 66, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Meuillet, E.J.; Mahadevan, D.; Berggren, M.; Coon, A.; Powis, G. Thioredoxin-1 Binds to the C2 Domain of PTEN Inhibiting PTEN’s Lipid Phosphatase Activity and Membrane Binding: A Mechanism for the Functional Loss of PTEN’s Tumor Suppressor Activity. Arch. Biochem. Biophys. 2004, 429, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, M.; Nishitoh, H.; Fujii, M.; Takeda, K.; Tobiume, K.; Sawada, Y.; Kawabata, M.; Miyazono, K.; Ichijo, H. Mammalian Thioredoxin Is a Direct Inhibitor of Apoptosis Signal-Regulating Kinase (ASK) 1. EMBO J. 1998, 17, 2596–2606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogales, E. Structural Insights into Microtubule Function. Annu. Rev. Biochem. 2000, 69, 277–302. [Google Scholar] [CrossRef]
- Valiron, O.; Caudron, N.; Job, D. Microtubule Dynamics. Cell. Mol. Life Sci. CMLS 2001, 58, 2069–2084. [Google Scholar] [CrossRef]
- Checchi, P.M.; Nettles, J.H.; Zhou, J.; Snyder, J.P.; Joshi, H.C. Microtubule-Interacting Drugs for Cancer Treatment. Rends Pharmacol. Sci. 2003, 24, 361–365. [Google Scholar] [CrossRef]
- Lim, S.; Kaldis, P. Cdks, Cyclins and CKIs: Roles beyond Cell Cycle Regulation. Development 2013, 140, 3079–3093. [Google Scholar] [CrossRef] [Green Version]
- Tsai, W.-C.; Wang, W.-H.; Huang, B.-C.; Huang, C.-Y.; Sheu, J.-H. 5-Epi-Sinuleptolide from Soft Corals of the Genus Sinularia Exerts Cytotoxic Effects on Pancreatic Cancer Cell Lines via the Inhibition of JAK2/STAT3, AKT, and ERK Activity. Molecules 2021, 26, 6932. [Google Scholar] [CrossRef]
- Ono, M.; Takeshima, M.; Nishi, A.; Higuchi, T.; Nakano, S. Genistein Suppresses V-Src-Driven Proliferative Activity by Arresting the Cell-Cycle at G2/M through Increasing P21 Level in Src-Activated Human Gallbladder Carcinoma Cells. Nutr. Cancer 2021, 73, 1471–1479. [Google Scholar] [CrossRef]
- Wang, W.; Abbruzzese, J.L.; Evans, D.B.; Larry, L.; Cleary, K.R.; Chiao, P.J. The Nuclear Factor-ΚB RelA Transcription Factor Is Constitutively Activated in Human Pancreatic Adenocarcinoma Cells. Clin. Cancer Res. 1999, 5, 119–127. [Google Scholar] [PubMed]
- Nakanishi, C.; Toi, M. Nuclear Factor-ΚB Inhibitors as Sensitizers to Anticancer Drugs. Nat. Rev. Cancer 2005, 5, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Arlt, A.; Vorndamm, J.; Breitenbroich, M.; FoÈlsch, U.; Kalthoff, H.; Schmidt, W.E.; SchaÈfer, H. Inhibition of NF-ΚB Sensitizes Human Pancreatic Carcinoma Cells to Apoptosis Induced by Etoposide (VP16) or Doxorubicin. Oncogene 2001, 20, 859–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Verma, U.N.; Gaynor, R.B.; Frenkel, E.P.; Becerra, C.R. Enhanced Chemosensitivity to Irinotecan by RNA Interference-Mediated down-Regulation of the Nuclear Factor-ΚB P65 Subunit. Clin. Cancer Res. 2004, 10, 3333–3341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mabuchi, S.; Ohmichi, M.; Nishio, Y.; Hayasaka, T.; Kimura, A.; Ohta, T.; Saito, M.; Kawagoe, J.; Takahashi, K.; Yada-Hashimoto, N.; et al. Inhibition of NFκB Increases the Efficacy of Cisplatin in In Vitro and In Vivo Ovarian Cancer Models. J. Biol. Chem. 2004, 279, 23477–23485. [Google Scholar] [CrossRef] [Green Version]
- Ko, L.J.; Prives, C. P53: Puzzle and Paradigm. Genes Dev. 1996, 10, 1054–1072. [Google Scholar] [CrossRef] [Green Version]
- Levine, A.J. P53, the Cellular Gatekeeper for Growth and Division. Cell 1997, 88, 323–331. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Sun, J.; Su, X.; Yu, Q.; Yu, Q.; Zhang, P. A Review about the Development of Fucoidan in Antitumor Activity: Progress and Challenges. Carbohydr. Polym. 2016, 154, 96–111. [Google Scholar] [CrossRef]
- Park, H.Y.; Kim, G.Y.; Moon, S.K.; Kim, W.J.; Yoo, Y.H.; Choi, Y.H. Fucoidan Inhibits the Proliferation of Human Urinary Bladder Cancer T24 Cells by Blocking Cell Cycle Progression and Inducing Apoptosis. Molecules 2014, 19, 5981–5998. [Google Scholar] [CrossRef] [Green Version]
- Teng, H.; Yang, Y.; Wei, H.; Liu, Z.; Liu, Z.; Ma, Y.; Gao, Z.; Hou, L.; Zou, X. Fucoidan Suppresses Hypoxia-Induced Lymphangiogenesis and Lymphatic Metastasis in Mouse Hepatocarcinoma. Mar. Drugs 2015, 13, 3514–3530. [Google Scholar] [CrossRef] [Green Version]
- Kuznetsova, T.A.; Besednova, N.N.; Mamaev, A.N.; Momot, A.P.; Shevchenko, N.M.; Zvyagintseva, T.N. Anticoagulant Activity of Fucoidan from Brown Algae Fucus Evanescens of the Okhotsk Sea. Bull. Exp. Biol. Med. 2003, 136, 471–473. [Google Scholar] [CrossRef]
- Min, E.Y.; Kim, I.H.; Lee, J.; Kim, E.Y.; Choi, Y.H.; Nam, T.J. The Effects of Fucodian on Senescence Are Controlled by the P16 INK4a-PRb and P14Arf-P53 Pathways in Hepatocellular Carcinoma and Hepatic Cell Lines. Int. J. Oncol. 2014, 45, 47–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, M.D.; Yao, C.J.; Chow, J.M.; Chang, C.L.; Hwang, P.A.; Chuang, S.E.; Whang-Peng, J.; Lai, G.M. Fucoidan Elevates MicroRNA-29b to Regulate DNMT3B-MTSS1 Axis and Inhibit EMT in Human Hepatocellular Carcinoma Cells. Mar. Drugs 2015, 13, 6099–6116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Zhang, Q.; Song, J. Toxicological Evaluation of Fucoidan Extracted from Laminaria Japonica in Wistar Rats. Food Chem. Toxicol. 2005, 43, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Alekseyenko, T.V.; Zhanayeva, S.Y.; Venediktova, A.A.; Zvyagintseva, T.N.; Kuznetsova, T.A.; Besednova, N.N.; Korolenko, T.A. Antitumor and Antimetastatic Activity of Fucoidan, a Sulfated Polysaccharide Isolated from the Okhotsk Sea Fucus Evanescens Brown Alga. Bull. Exp. Biol. Med. 2007, 143, 730–732. [Google Scholar] [CrossRef]
- Etman, S.M.; Mehanna, R.A.; Bary, A.A.; Elnaggar, Y.S.R.; Abdallah, O.Y. Undaria Pinnatifida Fucoidan Nanoparticles Loaded with Quinacrine Attenuate Growth and Metastasis of Pancreatic Cancer. Int. J. Biol. Macromol. 2021, 170, 284–297. [Google Scholar] [CrossRef]
- Han, Y.S.; Lee, J.H.; Lee, S.H. Antitumor Effects of Fucoidan on Human Colon Cancer Cells via Activation of Akt Signaling. Biomol. Ther. 2015, 23, 232. [Google Scholar] [CrossRef] [Green Version]
- Cross, C.E.; Halliwell, B.; Borish, E.T.; Pryor, W.A.; Ames, B.N.; Saul, R.L.; McCORD, J.M.; Harman, D. Oxygen Radicals and Human Disease. Ann. Intern. Med. 1987, 107, 526–545. [Google Scholar] [CrossRef]
- Kim, A.; Ha, J.; Kim, J.; Cho, Y.; Ahn, J.; Cheon, C.; Kim, S.H.; Ko, S.G.; Kim, B. Natural Products for Pancreatic Cancer Treatment: From Traditional Medicine to Modern Drug Discovery. Nutrients 2021, 13, 3801. [Google Scholar] [CrossRef]
- Tripathi, S.K.; Biswal, B.K. Pterospermum acerifolium (L.) Wild Bark Extract Induces Anticarcinogenic Effect in Human Cancer Cells through Mitochondrial-Mediated ROS Generation. Mol. Biol. Rep. 2018, 45, 2283–2294. [Google Scholar] [CrossRef]
- Dyshlovoy, S.A.; Honecker, F. Marine Compounds and Cancer: 2017 Updates. Mar. Drugs 2018, 16, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DrugBank Online|Database for Drug and Drug Target Info. Available online: https://go.drugbank.com/ (accessed on 28 April 2022).
- Stonik, V.A.; Fedorov, S.N. Marine Low Molecular Weight Natural Products as Potential Cancer Preventive Compounds. Mar. Drugs 2014, 12, 636–671. [Google Scholar] [CrossRef] [PubMed]
- Dyshlovoy, S.A. Recent Updates on Marine Cancer-Preventive Compounds. Mar. Drugs 2021, 19, 558. [Google Scholar] [CrossRef] [PubMed]
- Candela, C.G.; López, L.B.; Kohen, V.L. Importance of a Balanced Omega 6/Omega 3 Ratio for the Maintenance of Health. Nutritional Recommendations. Nutr. Hosp. 2011, 26, 323–329. [Google Scholar] [CrossRef]
- Wendel, M.; Axel, R. Heller Anticancer Actions of Omega-3 Fatty Acids-Current State and Future Perspectives. Anti-Cancer Agents Med. Chem. 2009, 9, 457–470. [Google Scholar] [CrossRef]
- Nobili, S.; Lippi, D.; Witort, E.; Donnini, M.; Bausi, L.; Mini, E.; Capaccioli, S. Natural Compounds for Cancer Treatment and Prevention. Pharmacol. Res. 2009, 59, 365–378. [Google Scholar] [CrossRef]
- Choudhary, A.; Naughton, L.M.; Montánchez, I.; Dobson, A.D.W.; Rai, D.K. Current Status and Future Prospects of Marine Natural Products (MNPs) as Antimicrobials. Mar. Drugs 2017, 15, 272. [Google Scholar] [CrossRef]
- Ashburn, T.T.; Thor, K.B. Drug Repositioning: Identifying and Developing New Uses for Existing Drugs. Nat. Rev. Drug Discov. 2004, 3, 673–683. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Kingdom | Species | Extraction/Chemical Group | Key Findings | Cell Line/Animal Model/Epidemiology | Reference |
|---|---|---|---|---|---|
| Animalia | Soft coral: Sinularia flexibilis | Flexibilide C20H30O4 | Regulates several metabolic pathways, including glycerophospholipid and sphingolipid, leading to activation of caspases and induction of apoptosis | HCT-116 cells | [34] |
| Downregulates the tricarboxylic acid cycle (TCA), which leads to the loss of mitochondrial transmembrane potential and cell apoptosis | |||||
| Upregulates sphingosine-1-phosphate | |||||
| Soft coral: Sinularia sp. | Peroxy sesquiterpenoids | Increases production of H2O2 | HCT-116 cells | [35] | |
| Inhibits anti-apoptosis proteins, such as Bcl-xL and pAkt, leading to apoptosis | |||||
| Suppresses hemeoxygenase-1 (HO-1), nuclear factor-erythroid-2-related factor (Nrf2), and phosphoNrf2 (pNrf2) | |||||
| Soft coral: Carotalcyon sp. | DCM/MeOH extract | Causes G2/M transition and induction of apoptosis | HGUE-C-1, HT-29, and SW-480 cells | [36] | |
| Nudibranchs: Phyllidia varicosa and Dolabella auricularia | Increase intracellular reactive oxygen species (ROS) accumulation, mitochondrial depolarization, caspase activation, and DNA damage that leads to apoptosis | ||||
| Holothurian: Pseudocol ochirus violaceus | Extract causes necrotic cell death | ||||
| Mollusc: Dicathais orbita | Chloroform/MeOH extract | Apoptotic response to a genotoxic carcinogen without any apparent toxic side effects | In vivo model: mice | [37] | |
| Contains 6-bromoisatin, which significantly enhances apoptosis and reduces cell proliferation | |||||
| Sponge: Lithoplocamia lithistoides | PM060184 C31H45N3O7 | Inhibits tubulin polymerization, which reduces microtubule dynamicity, and inhibits cell migration | HCT-116 cells | [38] | |
| Sponges: Smenospongia aurea, S. cerebriformis, and Verongula rigida | Monohydroxy-1,4-benzoquinones | Induce expression of tumor suppressor gene P53 | HCT-116 and RKO cells | [39] | |
| Stabilize the tumor suppressor gene p53 through phosphorylation at Ser15 | |||||
| 1,4-naphthoquinone | Upregulates the expression of p21WAF1/CIP1, a p53-dependent gene, and suppresses proliferation of cancer cells | ||||
| Causes G2/M cell cycle arrest and increases caspase-3 cleavage; induces apoptosis | |||||
| Sponge: Hippospongia metachromia | Ilimaquinone and ethylsmenoquinone | Activat p53 pathway and upregulate the expression of p21 | HCT116 and RKO cells | [40] | |
| Causes G2/M cell cycle arrest | |||||
| Increases caspase-3 cleavage | |||||
| Fish | Fatty acids | Improve survival among stage III CRC patients with wild-type KRAS (Kirsten rat sarcoma virus) | Epidemiology research | [41] | |
| Algae | Red seaweed | Carbohydrate | Inhibits proliferation of colon cancer cells and induces apoptosis | HCT-116 cells | [42] |
| Activates caspases -3 and -9, which leads to apoptosis | |||||
| Reduces the expression levels of anti-apoptotic proteins Bcl-2 and Bcl-xL and enhances the expression of the pro-apoptotic protein Bax | |||||
| Induces P5, which is involved in apoptosis | |||||
| Seaweeds: Miyeok and Dashima | Dietary intake | Lower the risk of CRC associated with the cellular Myelocytomatosis (c-MYC) | Epidemiology | [43] | |
| Inhibit proliferation and induce apoptosis via regulation of the WNT/β-catenin signaling pathway | |||||
| Green algae: Ulva fasciata | EtOH 80% extract | Causes morphological changes indicative of apoptosis (apoptotic bodies, DNA fragmentation, and mitochondrial membrane depolarization) | HCT-116 cells | [44] | |
| Induces the mitochondrial intrinsic pathway by lowering Bcl-2 regulation and raising Bax regulation, subsequently upregulating caspases -9 and -3 and increasing mitochondrial membrane permeability | |||||
| Brown algae: Fucus vesiculosus | Sulfated polysaccharide Fucoidan | Increases levels of cleaved caspases -8, -9, -7, and -3 and cleaved poly (ADP-ribose) polymerase, leading to induction of apoptosis in HT-29 cells | HT-29 and HCT-116 cells | [45] | |
| Attenuates the levels of the X-linked inhibitor of apoptosis protein and survivin | |||||
| Enhances mitochondrial membrane permeability as well as cytochrome c and Smac/Diablo release from the mitochondria, leading to apoptosis | |||||
| Increases the levels of the pro-apoptotic proteins Bak and truncated Bid and reduces the levels of the anti-apoptotic protein Mcl-1 | |||||
| Increases the levels of the tumor necrosis factor-related apoptosis-inducing ligand, Fas, and death receptor proteins | |||||
| Activates the AKT pathway | [46] | ||||
| Red algae: Asparagopsis armata and Sphaerococcus coronopifolius | Crude methanol and dichloromethane extracts | Decreases Caco-2 cell proliferation | Caco-2 cells | [47] | |
| Fungus | Aspergillus sp. | Ketone aryl | Inhibits the growth of cells through G2/M cell cycle arrest followed by apoptosis | RKO cells | [48] |
| Triggers microtubule disassembly and induces ROS | |||||
| Diketopiperazine disulfides + a new aranotin derivative deoxyapoaranotin | Strain KMD 901 shows potent cytotoxic activity towards five cancer cell lines (HCT-116, AGS, A549, MCF-7 and HepG2) | HCT-116 cells | [49] | ||
| Has a direct cytotoxic and apoptosis-inducing impact towards HCT-116 cells | |||||
| Bacteria | Cyanobacteria: cf. Neolyngbya sp. | Lipopeptides | Signifcantly reduces cell viability by an unknown mechanism | HCT-116 cells | [50] |
| Cyanobacteria: Cyanobium sp. | Acetonitrile fractions extracts | Affects Bcl-2 expression and alters protein networks from endoplasmic reticulum stress to proteasome degradation and apoptosis | RKO cells | [51] | |
| Cyanobacteria: Synechocystis salina | Affects progression of the cell cycle at the G2/M transition | ||||
| Cyanobacteria: Symploca sp. | Peptides Dolastatins | The combination of largazole and dolastatin 10 curbs the growth of HCT-116 cancer cells | HCT-116 cells | [52,53] |
| Kingdom | Species | Extraction/Chemical Group | Key Findings | Cell Line/Animal Model/Epidemiology | Reference |
|---|---|---|---|---|---|
| Animalia | Sponge: Amphibleptula | Acetonitrile fractions extracts | Induces apoptosis in the AsPC-1, BxPC-3, and PANC-1 cell lines but not in the MIA PaCa-2 cell line | AsPC-1, BxPC-3, MIA PaCa-2, and PANC-1 cells | [54] |
| Sponge: Batzella sp. | Inhibits NFκB transcriptional activity | ||||
| Sponge: Haliclona | Alkaloid | Affects vacuolar ATPase activity and significantly increases the level of the autophagosome markers LC3-II and p62/SQSTM1 | AsPC-1 and PANC-1 cells | [55] | |
| Sponge: Batzella sp. | Causes cell cycle arrest by intercalating into DNA and/or inhibiting topoisomerase II activity | AsPC-1, Panc-1, BxPC-3, and MIA PaCa2 cells | [56] | ||
| Has low toxicity against normal cells | |||||
| Sponge: Leiodermatium | Polyketide macrolide Leiodermatolide | Is a potent antimitotic agent | AsPC-1, BxPC-3 and MIA PaCa-2 cells | [57] | |
| Reduces cell viability and causes cell cycle arrest at the G2/M phase | |||||
| Interacts with tubulin, causing antimitotic activity | |||||
| Sponge: Amphibleptula | Methanolic extraction | induced apoptosis in the AsPC-1, BxPC-3 and PANC-1 cell lines and not to MIA PaCa-2 cell line | AsPC-1, BxPC-3, PANC-1 and MIA PaCa-2 cells | [54] | |
| inhibit NFκB transcriptional activity | |||||
| Brown algae: Ecklonia cava | Polyphenols | Increases ROS levels | PANC-1 cells | [58] | |
| Induces apoptosis | |||||
| Decreases the expression of cell progression inducers PCNA and Cyclin D1 as well as anti-apoptotic protein Bcl2 and increases the expression of the pro-apoptotic protein Bax | |||||
| Reduces the antioxidant defense system in cancer cells without increasing inflammatory cytokine levels | |||||
| Suppresses microtubules, appearance of multipolar mitosis, and lagging chromosomes at the metaphase plate | |||||
| Algae | Brown algae: Turbinaria conoides | Sulfated polysaccharide Fucoidan | Fucoidan nanoparticles loaded with quinacrine drug can reduce growth and metastasis of pancreatic cancer | In vivo model–mice and PANC-1 cells | [59] |
| Induce apoptosis, activates caspases -3, -8, and -9, and cleaves Poly ADP ribose polymerase (PARP) | PANC-1 cells | [60] | |||
| Fucoidan fraction–F5 | Inhibits 57 and 38 nuclear factor κB (NFκB) pathway molecules | ||||
| Increases cellular p53 and revert NFκB expression | |||||
| Brown algae: Hormophysa triquetra, Spatoglossum asperum, and Padina tetrastromatica | Polyphenols | Cause death of cancer cells | Panc-1, MiaPaCa-2, Panc-3.27, and BxPC-3 cells | [39] | |
| Inhibit tumor growth in xenograft mice after radiation therapy | MiaPaCa-2 cell line based on xenograft mice | ||||
| Fungus | Arctic fungi: Eutypella sp. | Libertellenone-H (LH) C22H28O6 | Increases the ROS level | PANC-1, SW1990, AsPC-1, and BxPC-3 cells | [61] |
| Inhibits the thioredoxin system (TRX) |
| Colon Cancer | Pancreatic Cancer |
|---|---|
| Upregulate sphingosine-1-phosphate expression | Inhibit NFκB transcriptional activity |
| Increase DNA fragmentation and damage | Inhibit the JAK/STAT signaling pathway |
| Induce reactive oxygen species (ROS) | |
| Downregulate the tricarboxylic acid cycle, leading to the loss of mitochondrial transmembrane potential | |
| Affect the mitochondrial apoptotic pathway through depolarization and membrane permeability and release of cytochrome c and SMAC/Diablo | Increase the levels of auto-phagosome marker LC3-II and p62/SQSTM2 |
| Increase the levels of the tumor necrosis factor-related apoptosis-inducing ligand, Fas, and death receptor 5 protein | |
| Suppress proteins related to the cell survival regulation signal of Nrf2-ARE (an antioxidant response element) | Affect vacuolar ATPase activity |
| Induce apoptosis through the caspase independent pathway | Reduce the antioxidant defense system |
| Activate the AKT pathway | |
| Cleave poly (ADP-ribose) polymerase (PARP) | Decrease the expression of tumor cell progression inducers (proliferating cell nuclear antigen and cyclin D1) |
| Regulate the WNT/β-catenin signaling pathway | |
| Alter protein networks, from endoplasmic reticulum stress to proteasome degradation, leading to induction of apoptosis | |
| Common pathways | |
| Induce apoptosis by caspase activation through the extrinsic and the intrinsic pathways | |
| Inhibit anti-apoptotic factors thioredoxin (TRX), X-linked (XIAP), and survivin | |
| Interact with tubulin to cause anti-mitotic activity | |
| Suppress cell cycle progression; cause cell cycle arrest at the G2/M phase | |
| Activate the p53 pathway | |
| Increase intracellular ROS accumulation and induction of apoptosis | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fares Amer, N.; Luzzatto Knaan, T. Natural Products of Marine Origin for the Treatment of Colorectal and Pancreatic Cancers: Mechanisms and Potential. Int. J. Mol. Sci. 2022, 23, 8048. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23148048
Fares Amer N, Luzzatto Knaan T. Natural Products of Marine Origin for the Treatment of Colorectal and Pancreatic Cancers: Mechanisms and Potential. International Journal of Molecular Sciences. 2022; 23(14):8048. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23148048
Chicago/Turabian StyleFares Amer, Nasrin, and Tal Luzzatto Knaan. 2022. "Natural Products of Marine Origin for the Treatment of Colorectal and Pancreatic Cancers: Mechanisms and Potential" International Journal of Molecular Sciences 23, no. 14: 8048. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23148048