
Social Fear Affects Limbic System Neuronal Activity and Gene Expression

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Behavioral Analyses Confirm Successful Social Fear Conditioning
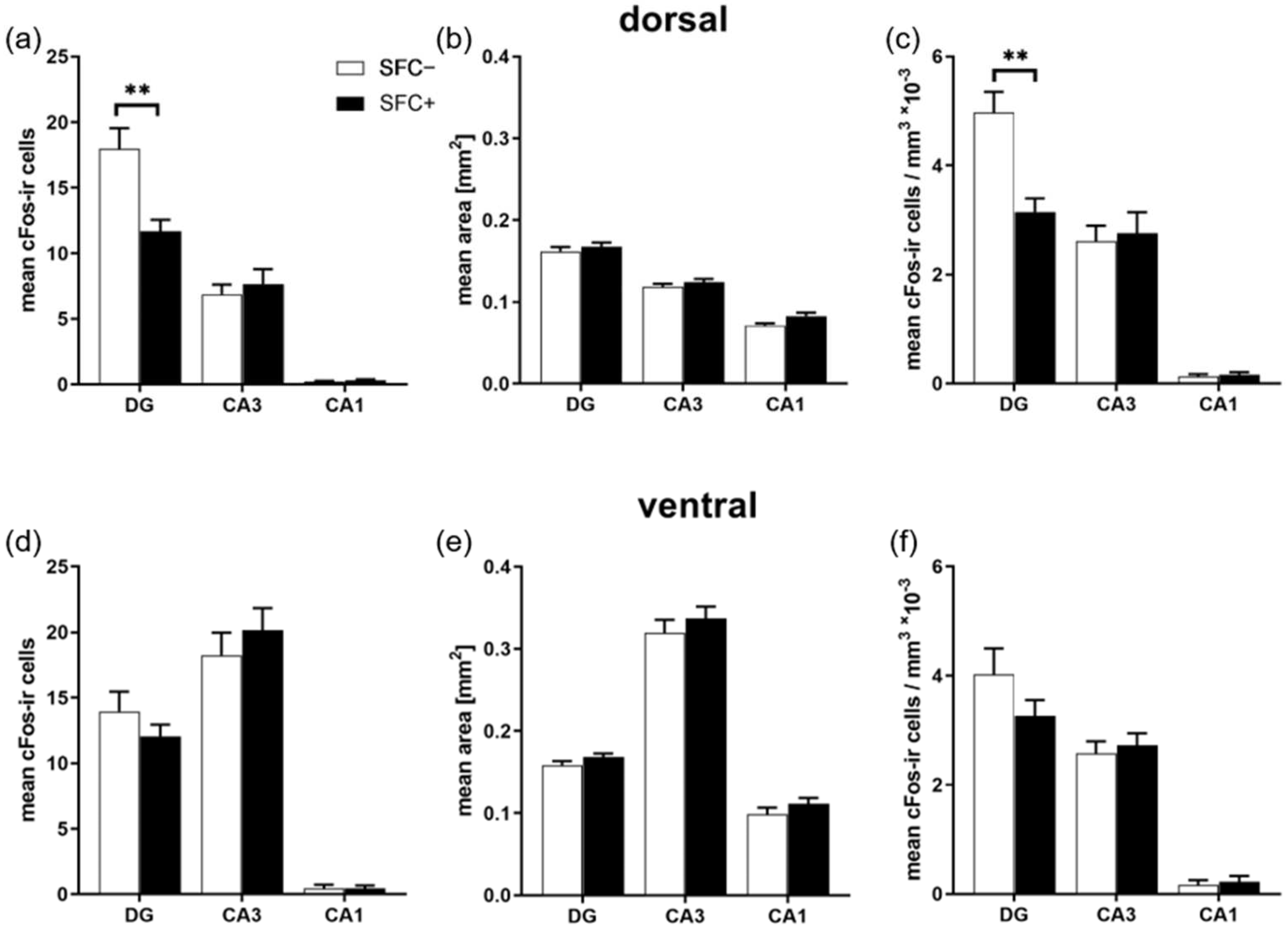
2.2. Social Fear Decreases Neuronal Activity in the Dorsal Dentate Gyrus, but Not in Other Dorsal Hippocampus Regions or in the Ventral Hippocampus
2.3. Social Fear Does Not Affect Neuronal Activity in the PVN
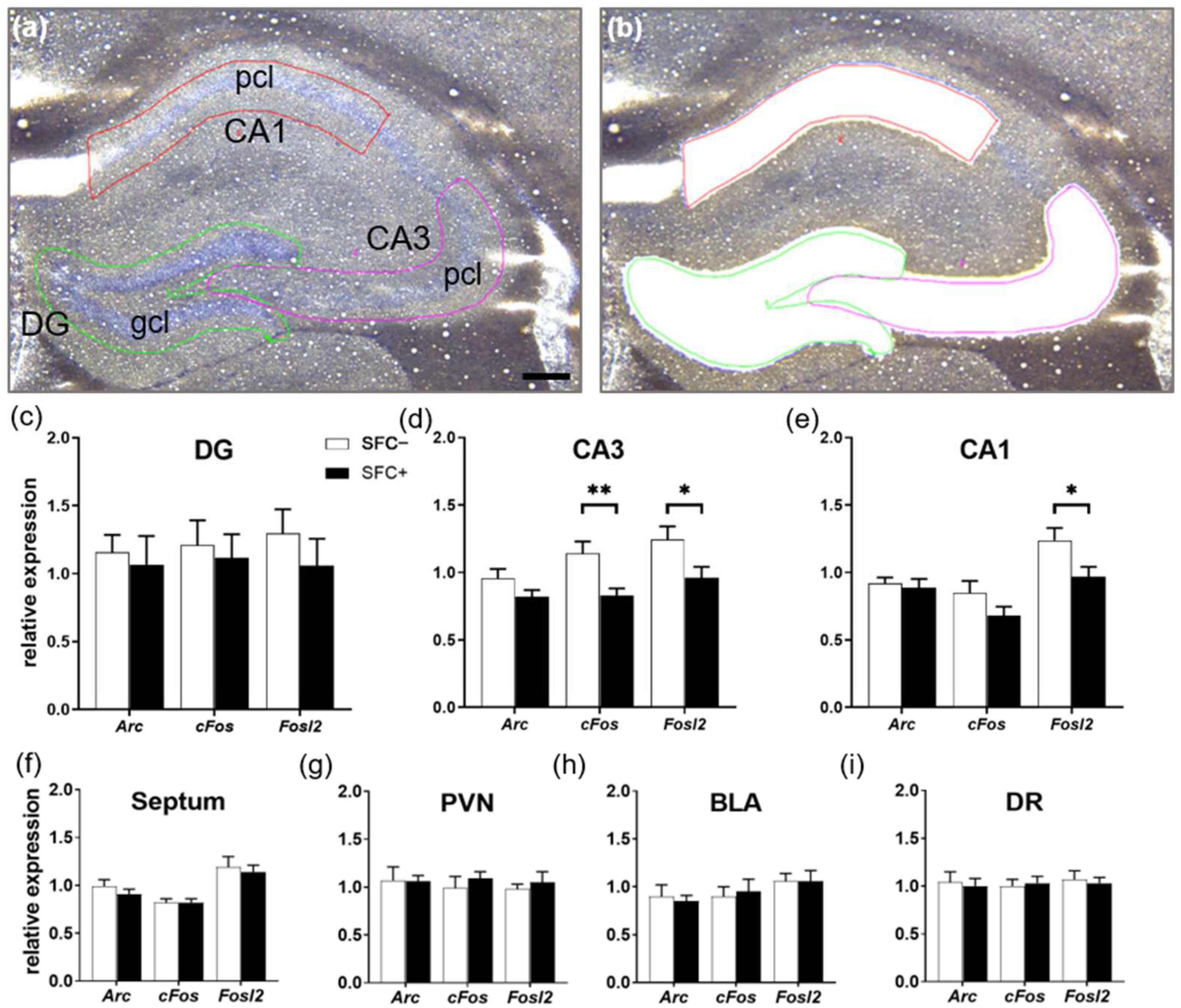
2.4. Social Fear Reduces the Expression Levels of Immediate Early Genes in the Dorsal Hippocampus
2.5. Social Fear Alters the Expression of Neuropeptide Y and Serotonin System-Related Genes
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Behavioral Experiment
4.2.1. Social Fear Conditioning (SFC) Paradigm
4.2.2. Social Fear Assessment
4.3. Tissue Preparation
4.3.1. Immunohistochemistry
Single Immunohistochemistry Staining of cFos
Immunofluorescence Double Staining of cFos and Vasopressin
Quantification of Immunolabeled Cells
4.3.2. Gene Expression Analysis
Laser Capture Microdissection
Quantitative Real-Time PCR (qPCR)
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kessler, R.C.; Petukhova, M.; Sampson, N.A.; Zaslavsky, A.M.; Wittchen, H.U. Twelve-month and lifetime prevalence and lifetime morbid risk of anxiety and mood disorders in the United States. Int. J. Methods Psychiatr. Res. 2012, 21, 169–184. [Google Scholar] [CrossRef]
- Wittchen, H.U.; Jacobi, F. Size and burden of mental disorders in Europe—A critical review and appraisal of 27 studies. Eur. Neuropsychopharmacol. 2005, 15, 357–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craske, M.G.; Stein, M.B. Anxiety. Lancet 2016, 388, 3048–3059. [Google Scholar] [CrossRef]
- Stein, M.B.; Stein, D.J. Social anxiety disorder. Lancet 2008, 371, 1115–1125. [Google Scholar] [CrossRef]
- Ruscio, A.M.; Brown, T.A.; Chiu, W.T.; Sareen, J.; Stein, M.B.; Kessler, R.C. Social fears and social phobia in the USA: Results from the National Comorbidity Survey Replication. Psychol. Med. 2008, 38, 15–28. [Google Scholar] [CrossRef]
- Koyuncu, A.; İnce, E.; Ertekin, E.; Tükel, R. Comorbidity in social anxiety disorder: Diagnostic and therapeutic challenges. Drugs Context 2019, 8, 212573. [Google Scholar] [CrossRef]
- Chartier, M.J.; Walker, J.R.; Stein, M.B. Considering comorbidity in social phobia. Soc. Psychiatry Psychiatr. Epidemiol. 2003, 38, 728–734. [Google Scholar] [CrossRef]
- Pelissolo, A.; Abou Kassm, S.; Delhay, L. Therapeutic strategies for social anxiety disorder: Where are we now? Expert Rev. Neurother. 2019, 19, 1179–1189. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.; Hattingh, C.J.; Kariuki, C.M.; Tromp, S.A.; van Balkom, A.J.; Ipser, J.C.; Stein, D.J. Pharmacotherapy for social anxiety disorder (SAnD). Cochrane Database Syst. Rev. 2017, 10, Cd001206. [Google Scholar] [CrossRef]
- Blanco, C.; Bragdon, L.B.; Schneier, F.R.; Liebowitz, M.R. The evidence-based pharmacotherapy of social anxiety disorder. Int. J. Neuropsychopharmacol. 2013, 16, 235–249. [Google Scholar] [CrossRef]
- Tanimizu, T.; Kono, K.; Kida, S. Brain networks activated to form object recognition memory. Brain Res. Bull. 2018, 141, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Tillfors, M.; Furmark, T.; Marteinsdottir, I.; Fischer, H.; Pissiota, A.; Långström, B.; Fredrikson, M. Cerebral blood flow in subjects with social phobia during stressful speaking tasks: A PET study. Am. J. Psychiatry 2001, 158, 1220–1226. [Google Scholar] [CrossRef] [PubMed]
- Lorberbaum, J.P.; Kose, S.; Johnson, M.R.; Arana, G.W.; Sullivan, L.K.; Hamner, M.B.; Ballenger, J.C.; Lydiard, R.B.; Brodrick, P.S.; Bohning, D.E.; et al. Neural correlates of speech anticipatory anxiety in generalized social phobia. Neuroreport 2004, 15, 2701–2705. [Google Scholar]
- Phan, K.L.; Fitzgerald, D.A.; Nathan, P.J.; Tancer, M.E. Association between amygdala hyperactivity to harsh faces and severity of social anxiety in generalized social phobia. Biol. Psychiatry 2006, 59, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Guyer, A.E.; Monk, C.S.; McClure-Tone, E.B.; Nelson, E.E.; Roberson-Nay, R.; Adler, A.D.; Fromm, S.J.; Leibenluft, E.; Pine, D.S.; Ernst, M. A developmental examination of amygdala response to facial expressions. J. Cogn. Neurosci. 2008, 20, 1565–1582. [Google Scholar] [CrossRef] [Green Version]
- Fanselow, M.S.; LeDoux, J.E. Why we think plasticity underlying Pavlovian fear conditioning occurs in the basolateral amygdala. Neuron 1999, 23, 229–232. [Google Scholar] [CrossRef] [Green Version]
- Kilts, C.D.; Kelsey, J.E.; Knight, B.; Ely, T.D.; Bowman, F.D.; Gross, R.E.; Selvig, A.; Gordon, A.; Newport, D.J.; Nemeroff, C.B. The neural correlates of social anxiety disorder and response to pharmacotherapy. Neuropsychopharmacology 2006, 31, 2243–2253. [Google Scholar] [CrossRef]
- Schneider, F.; Weiss, U.; Kessler, C.; Müller-Gärtner, H.W.; Posse, S.; Salloum, J.B.; Grodd, W.; Himmelmann, F.; Gaebel, W.; Birbaumer, N. Subcortical correlates of differential classical conditioning of aversive emotional reactions in social phobia. Biol. Psychiatry 1999, 45, 863–871. [Google Scholar] [CrossRef]
- Squire, L.R. Memory and the hippocampus: A synthesis from findings with rats, monkeys, and humans. Psychol. Rev. 1992, 99, 195–231. [Google Scholar] [CrossRef]
- Kim, J.J.; Rison, R.A.; Fanselow, M.S. Effects of amygdala, hippocampus, and periaqueductal gray lesions on short- and long-term contextual fear. Behav. Neurosci. 1993, 107, 1093–1098. [Google Scholar] [CrossRef]
- Toth, I.; Neumann, I.D. Animal models of social avoidance and social fear. Cell Tissue Res. 2013, 354, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Réus, G.Z.; Dos Santos, M.A.; Abelaira, H.M.; Quevedo, J. Animal models of social anxiety disorder and their validity criteria. Life Sci. 2014, 114, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Toth, I.; Neumann, I.D.; Slattery, D.A. Social fear conditioning: A novel and specific animal model to study social anxiety disorder. Neuropsychopharmacology 2012, 37, 1433–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornhuber, J.; Huber, S.E.; Zoicas, I. Effects of conditioned social fear on ethanol drinking and vice-versa in male mice. Psychopharmacology 2019, 236, 2059–2067. [Google Scholar] [CrossRef] [PubMed]
- Kornhuber, J.; Zoicas, I. Neuropeptide Y as Alternative Pharmacotherapy for Antidepressant-Resistant Social Fear. Int. J. Mol. Sci. 2020, 21, 8220. [Google Scholar] [CrossRef]
- Zorn, J.V.; Schür, R.R.; Boks, M.P.; Kahn, R.S.; Joëls, M.; Vinkers, C.H. Cortisol stress reactivity across psychiatric disorders: A systematic review and meta-analysis. Psychoneuroendocrinology 2017, 77, 25–36. [Google Scholar] [CrossRef]
- Kische, H.; Ollmann, T.M.; Voss, C.; Hoyer, J.; Rückert, F.; Pieper, L.; Kirschbaum, C.; Beesdo-Baum, K. Associations of saliva cortisol and hair cortisol with generalized anxiety, social anxiety, and major depressive disorder: An epidemiological cohort study in adolescents and young adults. Psychoneuroendocrinology 2021, 126, 105167. [Google Scholar] [CrossRef]
- Ledoux, J.E.; Muller, J. Emotional memory and psychopathology. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1997, 352, 1719–1726. [Google Scholar] [CrossRef] [Green Version]
- Lim, M.M.; Bielsky, I.F.; Young, L.J. Neuropeptides and the social brain: Potential rodent models of autism. Int. J. Dev. Neurosci. 2005, 23, 235–243. [Google Scholar] [CrossRef]
- Viviani, D.; Stoop, R. Opposite effects of oxytocin and vasopressin on the emotional expression of the fear response. Prog. Brain Res. 2008, 170, 207–218. [Google Scholar] [CrossRef]
- Bocchio, M.; McHugh, S.B.; Bannerman, D.M.; Sharp, T.; Capogna, M. Serotonin, Amygdala and Fear: Assembling the Puzzle. Front. Neural Circuits 2016, 10, 24. [Google Scholar] [CrossRef] [PubMed]
- Sagar, S.M.; Sharp, F.R.; Curran, T. Expression of c-fos protein in brain: Metabolic mapping at the cellular level. Science 1988, 240, 1328–1331. [Google Scholar] [CrossRef] [PubMed]
- Dragunow, M.; Faull, R. The use of c-fos as a metabolic marker in neuronal pathway tracing. J. Neurosci. Methods 1989, 29, 261–265. [Google Scholar] [CrossRef]
- Kovács, K.J. Measurement of immediate-early gene activation-c-fos and beyond. J. Neuroendocrinol. 2008, 20, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Zoicas, I.; Slattery, D.A.; Neumann, I.D. Brain oxytocin in social fear conditioning and its extinction: Involvement of the lateral septum. Neuropsychopharmacology 2014, 39, 3027–3035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornhuber, J.; Zoicas, I. Neuropeptide Y reduces expression of social fear via simultaneous activation of Y1 and Y2 receptors. J. Psychopharmacol. 2019, 33, 1533–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornhuber, J.; Zoicas, I. Neuropeptide Y Reduces Social Fear in Male Mice: Involvement of Y1 and Y2 Receptors in the Dorsolateral Septum and Central Amygdala. Int. J. Mol. Sci. 2021, 22, 10142. [Google Scholar] [CrossRef]
- Chen, C.C.; Huang, C.C.; Hsu, K.S. Chronic Social Stress Affects Synaptic Maturation of Newly Generated Neurons in the Adult Mouse Dentate Gyrus. Int. J. Neuropsychopharmacol. 2015, 19, pyv097. [Google Scholar] [CrossRef] [Green Version]
- Yohn, C.N.; Ashamalla, S.A.; Bokka, L.; Gergues, M.M.; Garino, A.; Samuels, B.A. Social instability is an effective chronic stress paradigm for both male and female mice. Neuropharmacology 2019, 160, 107780. [Google Scholar] [CrossRef]
- Stephenson, D.T.; O’Neill, S.M.; Narayan, S.; Tiwari, A.; Arnold, E.; Samaroo, H.D.; Du, F.; Ring, R.H.; Campbell, B.; Pletcher, M.; et al. Histopathologic characterization of the BTBR mouse model of autistic-like behavior reveals selective changes in neurodevelopmental prote说ins and adult hippocampal neurogenesis. Mol. Autism. 2011, 2, 7. [Google Scholar] [CrossRef] [Green Version]
- Aranarochana, A.; Chaisawang, P.; Sirichoat, A.; Pannangrong, W.; Wigmore, P.; Welbat, J.U. Protective effects of melatonin against valproic acid-induced memory impairments and reductions in adult rat hippocampal neurogenesis. Neuroscience 2019, 406, 580–593. [Google Scholar] [CrossRef] [PubMed]
- Toth, I.; Neumann, I.D.; Slattery, D.A. Social fear conditioning as an animal model of social anxiety disorder. Curr. Protoc. Neurosci. 2013, 63, 9–42. [Google Scholar] [CrossRef]
- Zoicas, I.; Menon, R.; Neumann, I.D. Neuropeptide S reduces fear and avoidance of con-specifics induced by social fear conditioning and social defeat, respectively. Neuropharmacology 2016, 108, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Zoicas, I.; Neumann, I.D. Maternal separation facilitates extinction of social fear in adult male mice. Behav. Brain Res. 2016, 297, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Menon, R.; Grund, T.; Zoicas, I.; Althammer, F.; Fiedler, D.; Biermeier, V.; Bosch, O.J.; Hiraoka, Y.; Nishimori, K.; Eliava, M.; et al. Oxytocin Signaling in the Lateral Septum Prevents Social Fear during Lactation. Curr. Biol. 2018, 28, 1066–1078.e1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slattery, D.A.; Neumann, I.D.; Flor, P.J.; Zoicas, I. Pharmacological modulation of metabotropic glutamate receptor subtype 5 and 7 impairs extinction of social fear in a time-point-dependent manner. Behav. Brain Res. 2017, 328, 57–61. [Google Scholar] [CrossRef]
- Schwartz, W.J.; Smith, C.B.; Davidsen, L.; Savaki, H.; Sokoloff, L.; Mata, M.; Fink, D.J.; Gainer, H. Metabolic mapping of functional activity in the hypothalamo-neurohypophysial system of the rat. Science 1979, 205, 723–725. [Google Scholar] [CrossRef]
- Honkaniemi, J.; Kononen, J.; Kainu, T.; Pyykönen, I.; Pelto-Huikko, M. Induction of multiple immediate early genes in rat hypothalamic paraventricular nucleus after stress. Brain Res. Mol. Brain Res. 1994, 25, 234–241. [Google Scholar] [CrossRef]
- Kovács, K.J.; Sawchenko, P.E. Sequence of stress-induced alterations in indices of synaptic and transcriptional activation in parvocellular neurosecretory neurons. J. Neurosci. 1996, 16, 262–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ons, S.; Martí, O.; Armario, A. Stress-induced activation of the immediate early gene Arc (activity-regulated cytoskeleton-associated protein) is restricted to telencephalic areas in the rat brain: Relationship to c-fos mRNA. J. Neurochem. 2004, 89, 1111–1118. [Google Scholar] [CrossRef]
- Giustino, T.F.; Maren, S. The Role of the Medial Prefrontal Cortex in the Conditioning and Extinction of Fear. Front. Behav. Neurosci. 2015, 9, 298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maren, S.; Phan, K.L.; Liberzon, I. The contextual brain: Implications for fear conditioning, extinction and psychopathology. Nat. Rev. Neurosci. 2013, 14, 417–428. [Google Scholar] [CrossRef]
- Mikics, E.; Tóth, M.; Varjú, P.; Gereben, B.; Liposits, Z.; Ashaber, M.; Halász, J.; Barna, I.; Farkas, I.; Haller, J. Lasting changes in social behavior and amygdala function following traumatic experience induced by a single series of foot-shocks. Psychoneuroendocrinology 2008, 33, 1198–1210. [Google Scholar] [CrossRef] [PubMed]
- Navarro, J.F.; Rivera, A.; Maldonado, E.; Cavas, M.; de la Calle, A. Anxiogenic-like activity of 3,4-methylenedioxy-methamphetamine (“Ecstasy”) in the social interaction test is accompanied by an increase of c-fos expression in mice amygdala. Prog. Neuropsychopharmacol. Biol. Psychiatry 2004, 28, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, G.M.; Apergis, J.; Gorman, J.M.; LeDoux, J.E. Rodent doxapram model of panic: Behavioral effects and c-Fos immunoreactivity in the amygdala. Biol. Psychiatry 2003, 53, 863–870. [Google Scholar] [CrossRef]
- Asgari, P.; McKinney, G.; Hodges, T.E.; McCormick, C.M. Social Instability Stress in Adolescence and Social Interaction in Female Rats. Neuroscience 2021, 477, 1–13. [Google Scholar] [CrossRef]
- Tanimizu, T.; Kenney, J.W.; Okano, E.; Kadoma, K.; Frankland, P.W.; Kida, S. Functional Connectivity of Multiple Brain Regions Required for the Consolidation of Social Recognition Memory. J. Neurosci. 2017, 37, 4103–4116. [Google Scholar] [CrossRef] [PubMed]
- Lüscher Dias, T.; Fernandes Golino, H.; Oliveira, V.E.M.; Dutra Moraes, M.F.; Schenatto Pereira, G. c-Fos expression predicts long-term social memory retrieval in mice. Behav. Brain Res. 2016, 313, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, J.N.; Aldag, J.M.; Insel, T.R.; Young, L.J. Oxytocin in the medial amygdala is essential for social recognition in the mouse. J. Neurosci. 2001, 21, 8278–8285. [Google Scholar] [CrossRef]
- von Heimendahl, M.; Rao, R.P.; Brecht, M. Weak and nondiscriminative responses to conspecifics in the rat hippocampus. J. Neurosci. 2012, 32, 2129–2141. [Google Scholar] [CrossRef]
- Moretto, J.N.; Duffy, Á.M.; Scharfman, H.E. Acute restraint stress decreases c-fos immunoreactivity in hilar mossy cells of the adult dentate gyrus. Brain Struct. Funct. 2017, 222, 2405–2419. [Google Scholar] [CrossRef] [PubMed]
- Chung, L. A Brief Introduction to the Transduction of Neural Activity into Fos Signal. Dev. Reprod. 2015, 19, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.H.; Rendall, S.D.; Gray, J.M. Brain-wide maps of Fos expression during fear learning and recall. Learn. Mem. 2017, 24, 169–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishina, H.; Sato, H.; Suzuki, T.; Sato, M.; Iba, H. Isolation and characterization of fra-2, an additional member of the fos gene family. Proc. Natl. Acad. Sci. USA 1990, 87, 3619–3623. [Google Scholar] [CrossRef] [Green Version]
- Hughes, E.M.; Calcagno, P.; Sanchez, C.; Smith, K.; Kelly, J.P.; Finn, D.P.; Roche, M. Mu-opioid receptor agonism differentially alters social behaviour and immediate early gene expression in male adolescent rats prenatally exposed to valproic acid versus controls. Brain Res. Bull. 2021, 174, 260–267. [Google Scholar] [CrossRef]
- Hara, Y.; Ago, Y.; Higuchi, M.; Hasebe, S.; Nakazawa, T.; Hashimoto, H.; Matsuda, T.; Takuma, K. Oxytocin attenuates deficits in social interaction but not recognition memory in a prenatal valproic acid-induced mouse model of autism. Horm. Behav. 2017, 96, 130–136. [Google Scholar] [CrossRef]
- Meyza, K.; Nikolaev, T.; Kondrakiewicz, K.; Blanchard, D.C.; Blanchard, R.J.; Knapska, E. Neuronal correlates of asocial behavior in a BTBR T (+) Itpr3(tf)/J mouse model of autism. Front. Behav. Neurosci. 2015, 9, 199. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.B.; Cho, J.H. Synaptic Targeting of Double-Projecting Ventral CA1 Hippocampal Neurons to the Medial Prefrontal Cortex and Basal Amygdala. J. Neurosci. 2017, 37, 4868–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orsini, C.A.; Kim, J.H.; Knapska, E.; Maren, S. Hippocampal and prefrontal projections to the basal amygdala mediate contextual regulation of fear after extinction. J. Neurosci. 2011, 31, 17269–17277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radulovic, J.; Kammermeier, J.; Spiess, J. Generalization of fear responses in C57BL/6N mice subjected to one-trial foreground contextual fear conditioning. Behav. Brain Res. 1998, 95, 179–189. [Google Scholar] [CrossRef]
- Ploski, J.E.; Park, K.W.; Ping, J.; Monsey, M.S.; Schafe, G.E. Identification of plasticity-associated genes regulated by Pavlovian fear conditioning in the lateral amygdala. J. Neurochem. 2010, 112, 636–650. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J.; Horovitz, O.; Pellman, B.A.; Tan, L.M.; Li, Q.; Richter-Levin, G.; Kim, J.J. Dorsal periaqueductal gray-amygdala pathway conveys both innate and learned fear responses in rats. Proc. Natl. Acad. Sci. USA 2013, 110, 14795–14800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, T.C.; Cerminara, N.L.; Lumb, B.M.; Apps, R. Neural Correlates of Fear in the Periaqueductal Gray. J. Neurosci. 2016, 36, 12707–12719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichmann, F.; Holzer, P. Neuropeptide Y: A stressful review. Neuropeptides 2016, 55, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Sheehan, T.P.; Chambers, R.A.; Russell, D.S. Regulation of affect by the lateral septum: Implications for neuropsychiatry. Brain Res. Brain Res. Rev. 2004, 46, 71–117. [Google Scholar] [CrossRef]
- Harro, J. CCK and NPY as anti-anxiety treatment targets: Promises, pitfalls, and strategies. Amino Acids 2006, 31, 215–230. [Google Scholar] [CrossRef] [PubMed]
- Kornhuber, J.; Zoicas, I. Brain Region-Dependent Effects of Neuropeptide Y on Conditioned Social Fear and Anxiety-Like Behavior in Male Mice. Int. J. Mol. Sci. 2021, 22, 3695. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Damjanoska, K.J.; Carrasco, G.A.; Dudas, B.; D’Souza, D.N.; Tetzlaff, J.; Garcia, F.; Hanley, N.R.; Scripathirathan, K.; Petersen, B.R.; et al. Evidence that 5-HT2A receptors in the hypothalamic paraventricular nucleus mediate neuroendocrine responses to (-)DOI. J. Neurosci. 2002, 22, 9635–9642. [Google Scholar] [CrossRef] [Green Version]
- Van de Kar, L.D.; Blair, M.L. Forebrain pathways mediating stress-induced hormone secretion. Front. Neuroendocrinol. 1999, 20, 1–48. [Google Scholar] [CrossRef]
- Carrasco, G.A.; Van de Kar, L.D. Neuroendocrine pharmacology of stress. Eur. J. Pharmacol. 2003, 463, 235–272. [Google Scholar] [CrossRef]
- Alhaider, A.A.; Ageel, A.M.; Ginawi, O.T. The quipazine- and TFMPP-increased conditioned avoidance response in rats: Role of 5HT1C/5-HT2 receptors. Neuropharmacology 1993, 32, 1427–1432. [Google Scholar] [CrossRef]
- Carhart-Harris, R.L.; Nutt, D.J. Serotonin and brain function: A tale of two receptors. J. Psychopharmacol. 2017, 31, 1091–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asher, M.; Asnaani, A.; Aderka, I.M. Gender differences in social anxiety disorder: A review. Clin. Psychol. Rev. 2017, 56, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Karabeg, M.M.; Grauthoff, S.; Kollert, S.Y.; Weidner, M.; Heiming, R.S.; Jansen, F.; Popp, S.; Kaiser, S.; Lesch, K.P.; Sachser, N.; et al. 5-HTT deficiency affects neuroplasticity and increases stress sensitivity resulting in altered spatial learning performance in the Morris water maze but not in the Barnes maze. PLoS ONE 2013, 8, e78238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, K.; Paxinos, G. The Mouse Brain in Stereotaxic Coordinates; Elsevier: Amsterdam, The Netherlands, 1997. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruijter, J.M.; Ramakers, C.; Hoogaars, W.M.; Karlen, Y.; Bakker, O.; van den Hoff, M.J.; Moorman, A.F. Amplification efficiency: Linking baseline and bias in the analysis of quantitative PCR data. Nucleic Acids Res. 2009, 37, e45. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamann, C.S.; Bankmann, J.; Mora Maza, H.; Kornhuber, J.; Zoicas, I.; Schmitt-Böhrer, A. Social Fear Affects Limbic System Neuronal Activity and Gene Expression. Int. J. Mol. Sci. 2022, 23, 8228. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23158228
Hamann CS, Bankmann J, Mora Maza H, Kornhuber J, Zoicas I, Schmitt-Böhrer A. Social Fear Affects Limbic System Neuronal Activity and Gene Expression. International Journal of Molecular Sciences. 2022; 23(15):8228. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23158228
Chicago/Turabian StyleHamann, Catharina S., Julian Bankmann, Hanna Mora Maza, Johannes Kornhuber, Iulia Zoicas, and Angelika Schmitt-Böhrer. 2022. "Social Fear Affects Limbic System Neuronal Activity and Gene Expression" International Journal of Molecular Sciences 23, no. 15: 8228. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23158228