
AMPK Activation Is Important for the Preservation of Insulin Sensitivity in Visceral, but Not in Subcutaneous Adipose Tissue of Postnatally Overfed Rat Model of Polycystic Ovary Syndrome

 ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Results
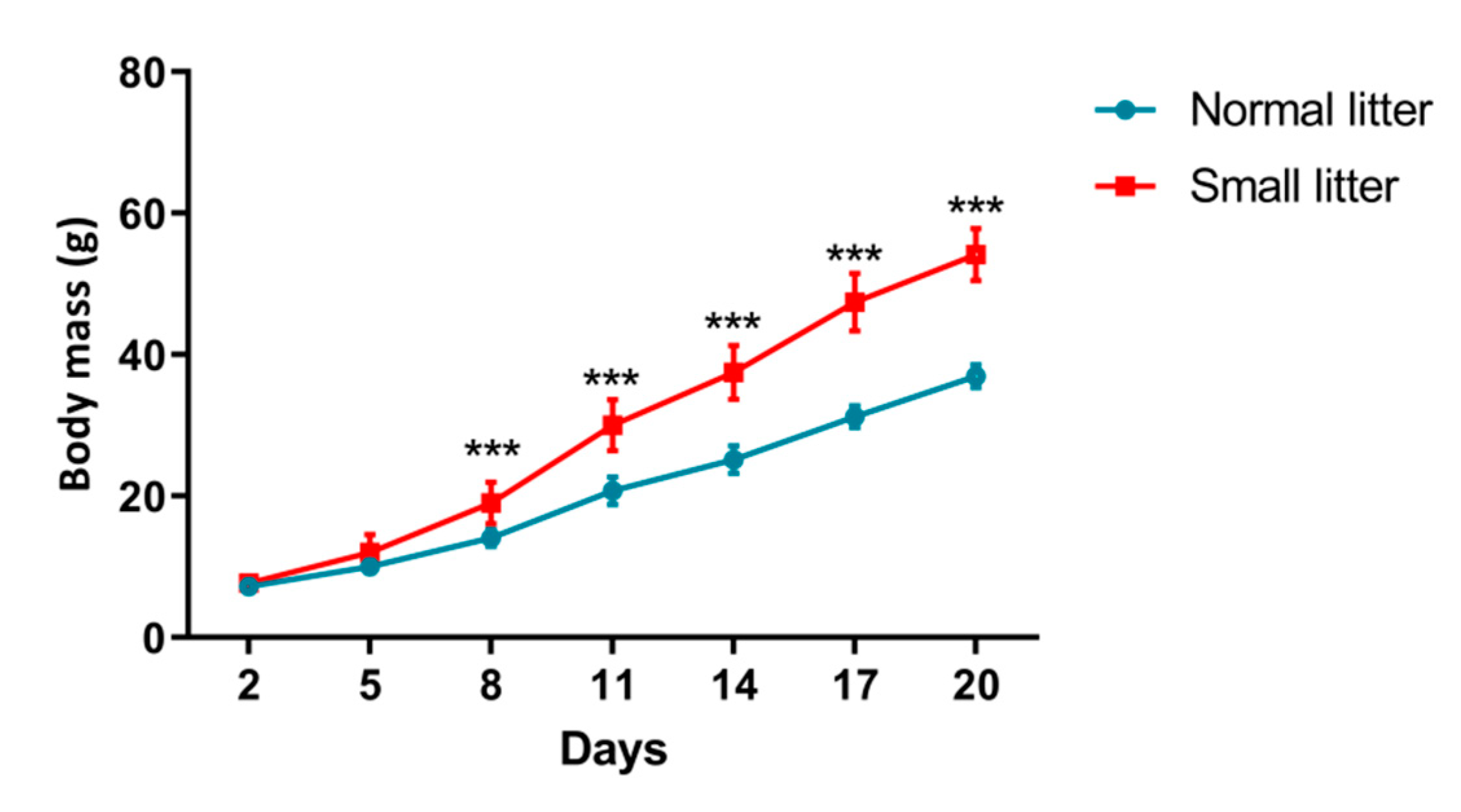
2.1. Physiological and Biochemical Parameters
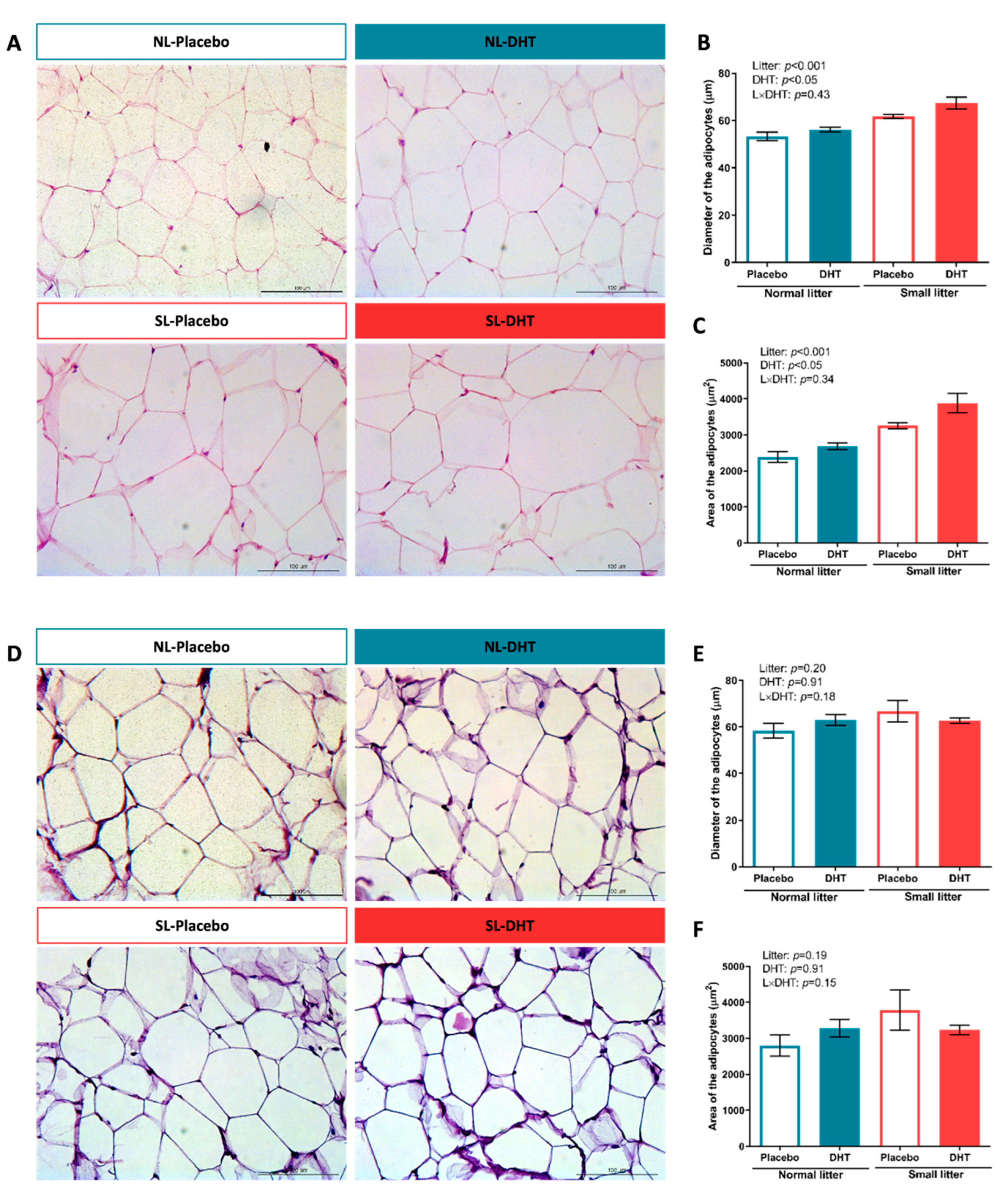
2.2. Histological and Morphometric Analysis of VAT and SAT
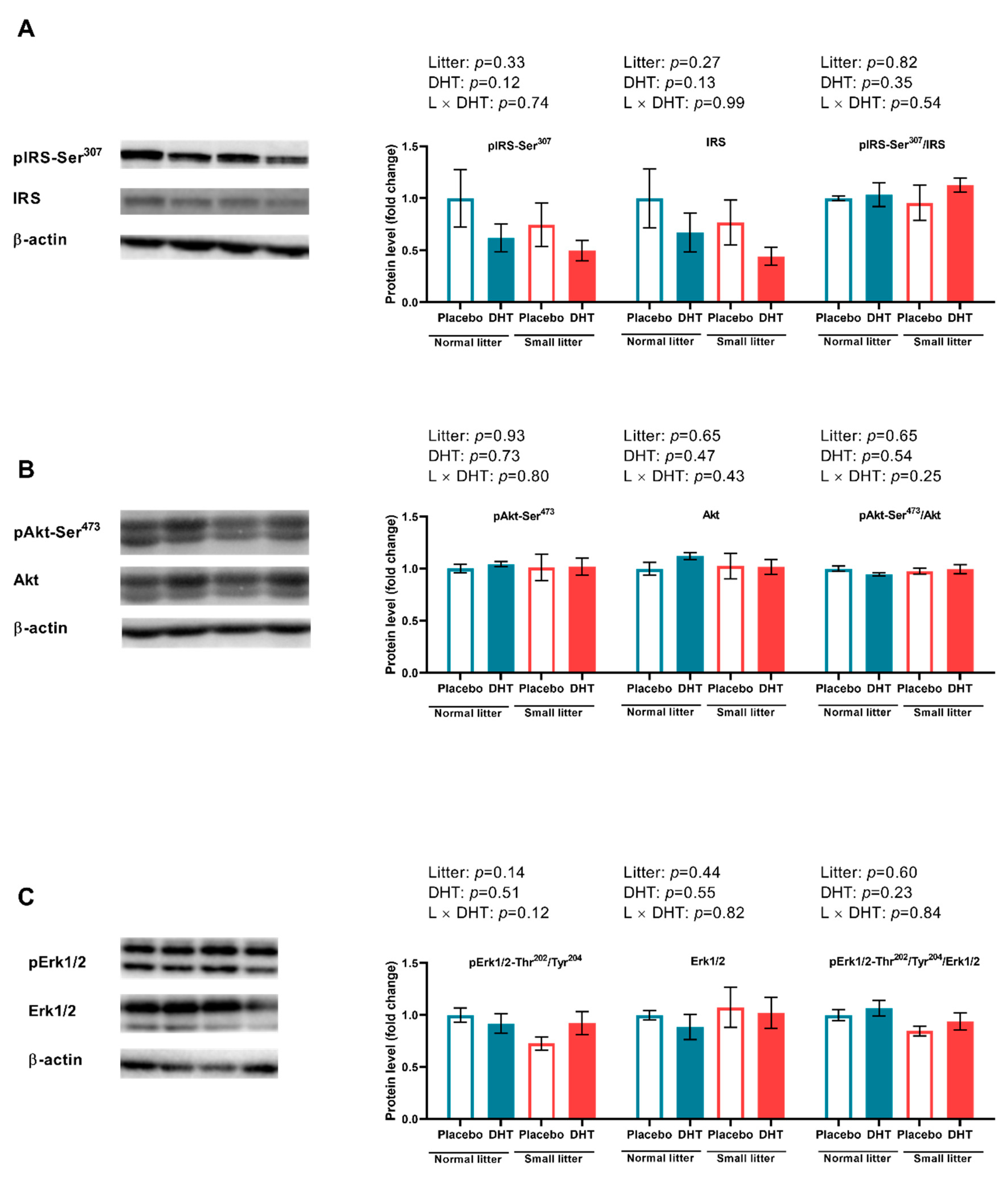
2.3. Insulin Signaling in the VAT
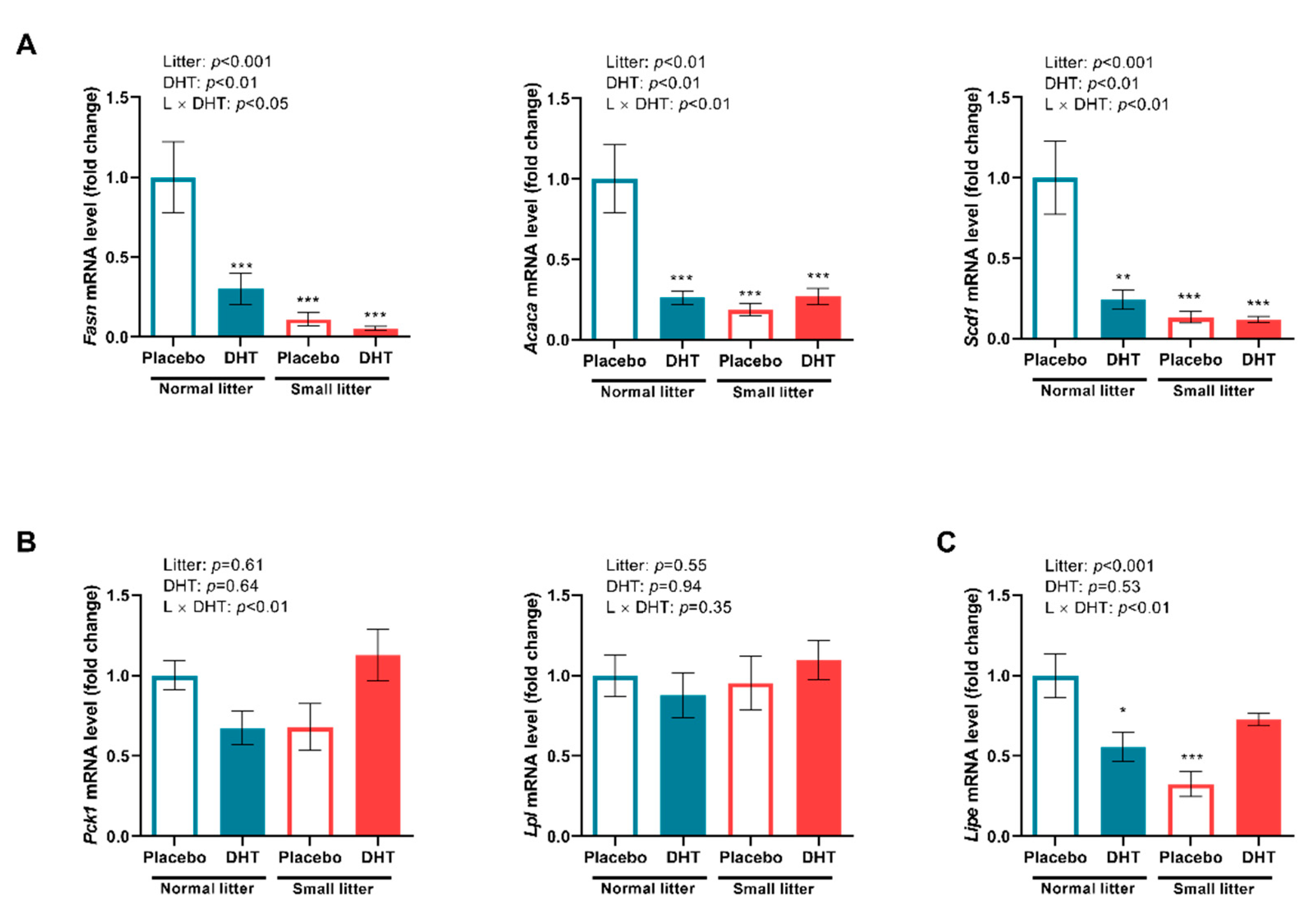
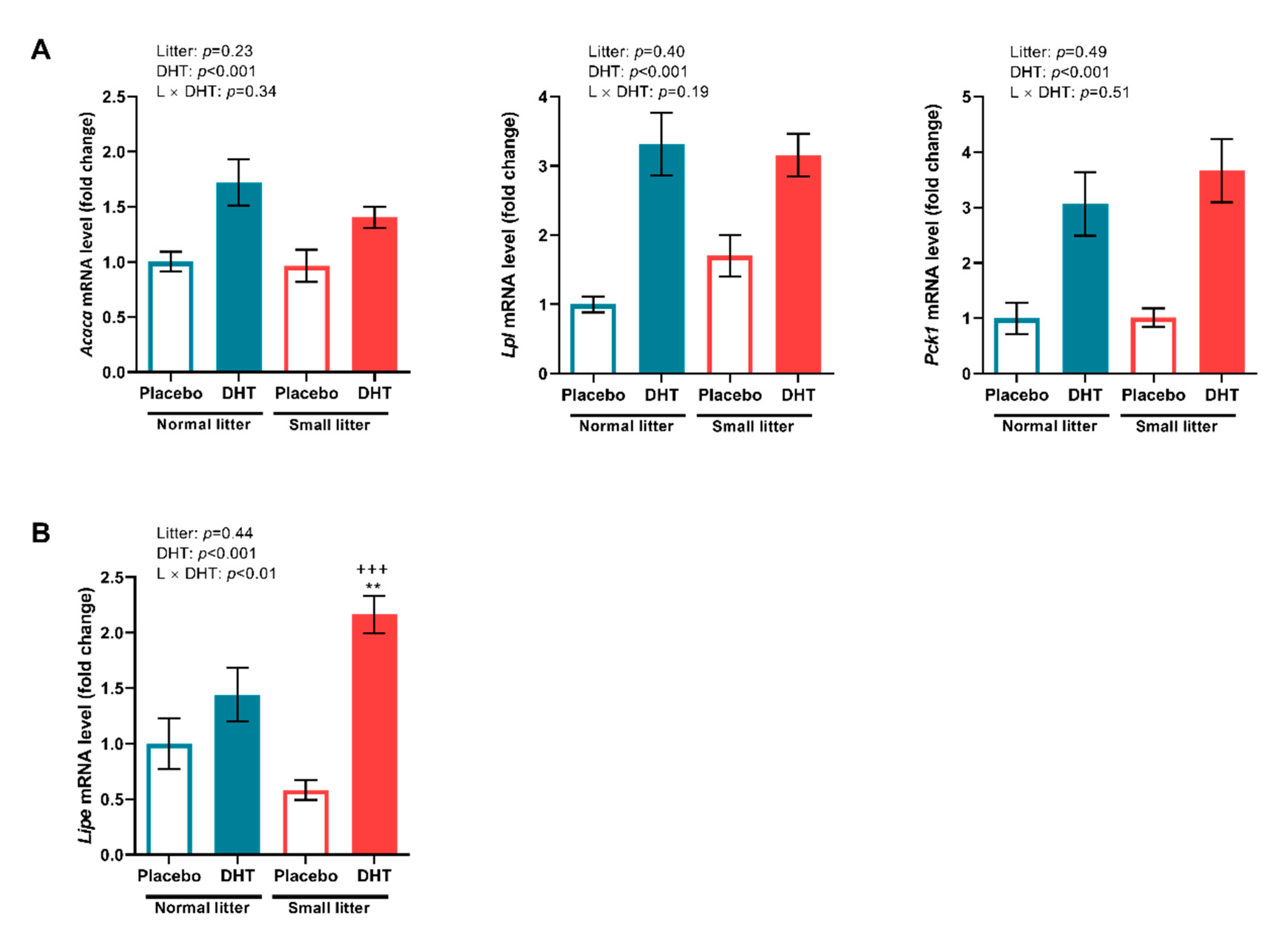
2.4. Markers of Lipid Metabolism in the VAT and SAT
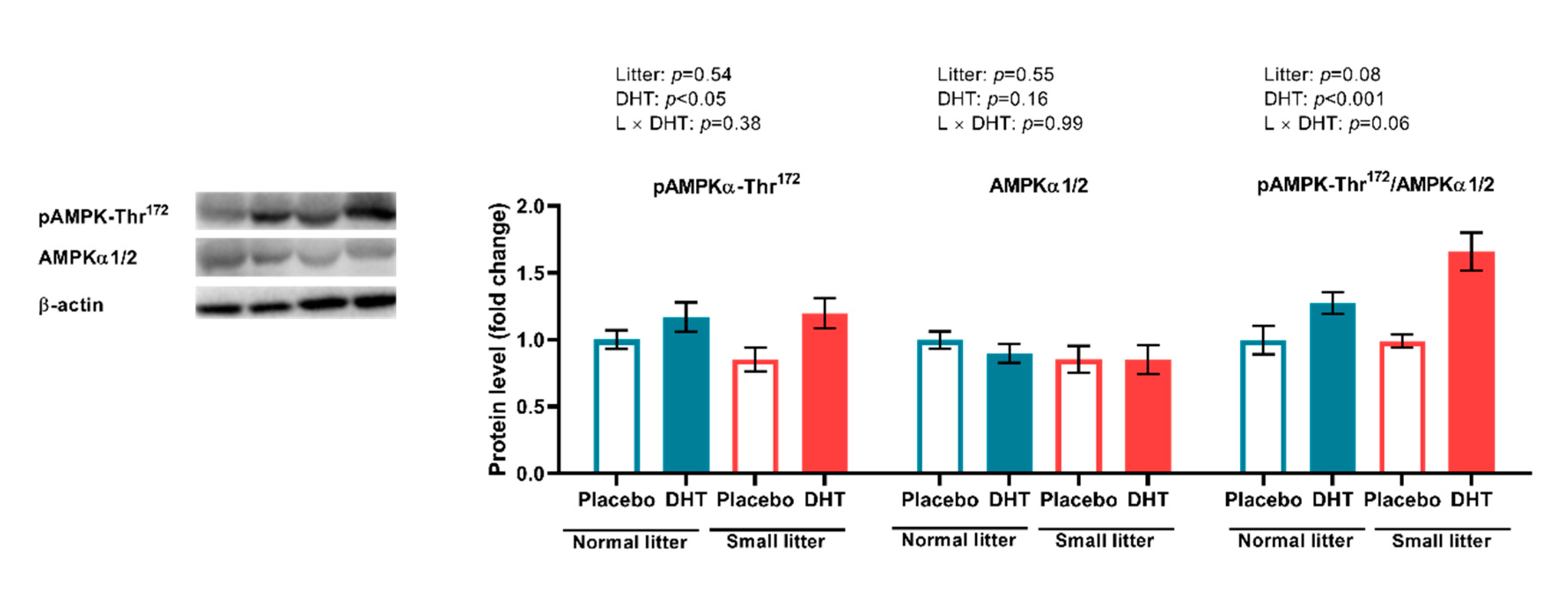
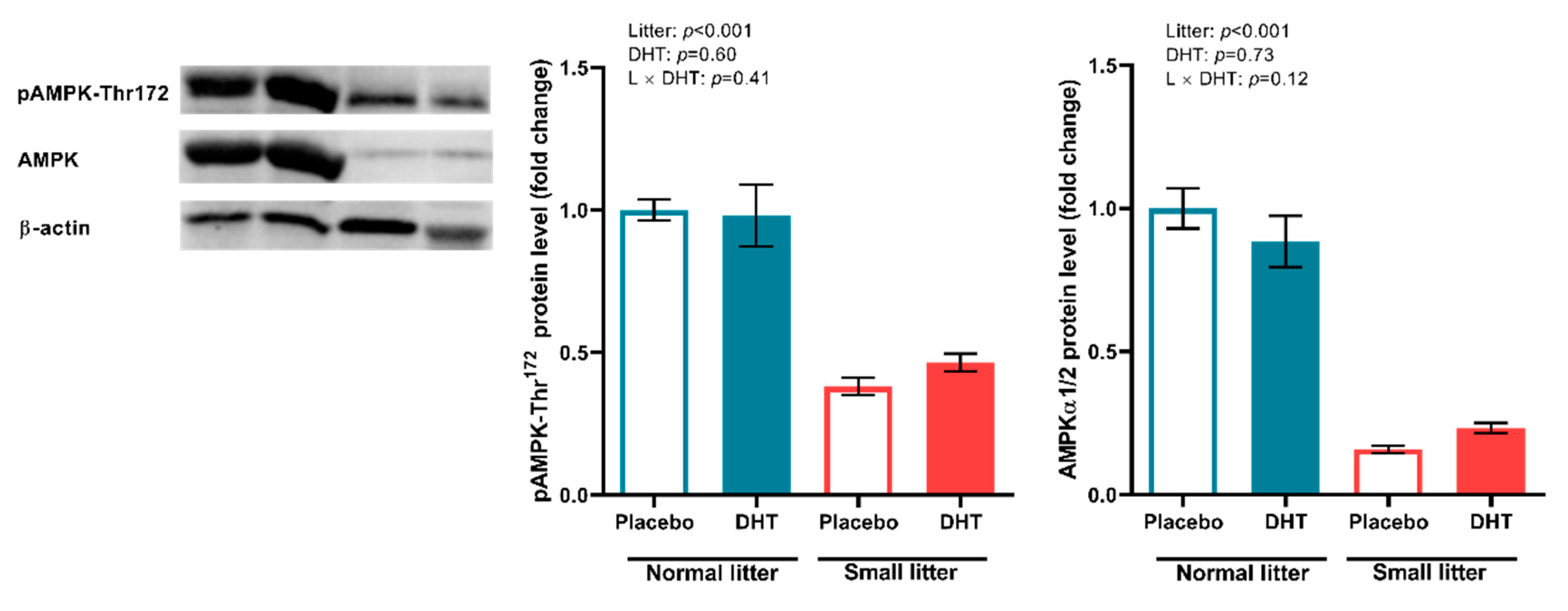
2.5. Energy Sensing in the VAT and SAT
3. Discussion
4. Materials and Methods
4.1. Experimental Design and Animals
4.2. Serum and Tissue Preparation
4.3. Determination of Blood Parameters
4.4. Assessment of Insulin Sensitivity
4.5. Histological and Morphometric Analysis
4.6. RNA Isolation, Reverse Transcription and Real-Time PCR
4.7. Preparation of Cytoplasmic, Nuclear and Mitochondrial Extracts from the VAT
4.8. Preparation of Total Protein from the SAT
4.9. Western Blot Analysis
4.10. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Conway, G.; Dewailly, D.; Diamanti-Kandarakis, E.; Escobar-Morreale, H.F.; Franks, S.; Gambineri, A.; Kelestimur, F.; Macut, D.; Micic, D.; Pasquali, R.; et al. The Polycystic Ovary Syndrome: A Position Statement from the European Society of Endocrinology. Eur. J. Endocrinol. 2014, 171, P1–P29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azziz, R.; Carmina, E.; Chen, Z.; Dunaif, A.; Laven, J.S.E.; Legro, R.S.; Lizneva, D.; Natterson-Horowtiz, B.; Teede, H.J.; Yildiz, B.O. Polycystic Ovary Syndrome. Nat. Rev. Dis. Prim. 2016, 2, 16057. [Google Scholar] [CrossRef] [PubMed]
- Walters, K.A. Androgens in Polycystic Ovary Syndrome: Lessons from Experimental Models. Curr. Opin. Endocrinol. Diabetes Obes. 2016, 23, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Kamada, S.; Yamamoto, Y.; Aoki, H.; Tamura, K.; Takeda, A.; Minato, S.; Masaki, R.; Yanagihara, R.; Hayashi, N.; Yano, Y.; et al. A Novel PCOS Rat Model and an Evaluation of Its Reproductive, Metabolic, and Behavioral Phenotypes. Reprod. Med. Biol. 2021, 21, e12416. [Google Scholar] [CrossRef] [PubMed]
- Sartor, B.M.; Dickey, R.P. Polycystic Ovarian Syndrome and the Metabolic Syndrome. Am. J. Med. Sci. 2005, 330, 336–342. [Google Scholar] [CrossRef]
- Glueck, C.J.; Goldenberg, N. Characteristics of Obesity in Polycystic Ovary Syndrome: Etiology, Treatment, and Genetics. Metabolism 2019, 92, 108–120. [Google Scholar] [CrossRef]
- Habbout, A.; Li, N.; Rochette, L.; Vergely, C. Postnatal Overfeeding in Rodents by Litter Size Reduction Induces Major Short- and Long-Term Pathophysiological Consequences. J. Nutr. 2013, 143, 553–562. [Google Scholar] [CrossRef]
- Morrison, J.A.; Friedman, L.A.; Wang, P.; Glueck, C.J. Metabolic Syndrome in Childhood Predicts Adult Metabolic Syndrome and Type 2 Diabetes Mellitus 25 to 30 Years Later. J. Pediatr. 2008, 152, 201–206. [Google Scholar] [CrossRef]
- McCartney, C.R.; Prendergast, K.A.; Chhabra, S.; Eagleson, C.A.; Yoo, R.; Chang, R.J.; Foster, C.M.; Marshall, J.C. The Association of Obesity and Hyperandrogenemia during the Pubertal Transition in Girls: Obesity as a Potential Factor in the Genesis of Postpubertal Hyperandrogenism. J. Clin. Endocrinol. Metab. 2006, 91, 1714–1722. [Google Scholar] [CrossRef] [Green Version]
- Franks, S. Polycystic Ovary Syndrome in Adolescents. Int. J. Obes. 2008, 32, 1035–1041. [Google Scholar] [CrossRef] [Green Version]
- Ogden, C.L.; Carrol, M.D.; Kit, B.K.; Flegal, K.M. Prevalence of Obesity and Trends in Body Mass Index Among US Children and Adolescents, 1999–2010. J. Am. Med. Assoc. 2012, 307, 483–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambineri, A.; Pelusi, C. Sex Hormones, Obesity and Type 2 Diabetes: Is There a Link? Endocr. Connect. 2019, 8, R1–R9. [Google Scholar] [CrossRef] [PubMed]
- Moran, L.J.; Misso, M.L.; Wild, R.A.; Norman, R.J. Impaired Glucose Tolerance, Type 2 Diabetes and Metabolic Syndrome in Polycystic Ovary Syndrome: A Systematic Review and Meta-Analysis. Hum. Reprod. Update 2010, 16, 347–363. [Google Scholar] [CrossRef]
- Marshall, J.C.; Dunaif, A. All Women With PCOS Should Be Treated For Insulin Resistance. Fertil. Steril. 2012, 97, 18–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choe, S.S.; Huh, J.Y.; Hwang, I.J.; Kim, J.I.; Kim, J.B. Adipose Tissue Remodeling: Its Role in Energy Metabolism and Metabolic Disorders. Front. Endocrinol. 2016, 7, 30. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Lee, M.; Oh, K. Adipose Tissue-Derived Signatures for Obesity and Type 2 Diabetes: Adipokines, Batokines and MicroRNAs. J. Clin. Med. 2019, 8, 854. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Garrido, M.A.; Tena-Sempere, M. Metabolic Dysfunction in Polycystic Ovary Syndrome: Pathogenic Role of Androgen Excess and Potential Therapeutic Strategies. Mol. Metab. 2020, 35, 100937. [Google Scholar] [CrossRef]
- Chazenbalk, G.; Singh, P.; Irge, D.; Shah, A.; Abbott, D.H.; Dumesic, D.A. Androgens Inhibit Adipogenesis during Human Adipose Stem Cell Commitment to Preadipocyte Formation. Steroids 2013, 78, 920–926. [Google Scholar] [CrossRef] [Green Version]
- Bijland, S.; Mancini, S.J.; Salt, I.P. Role of AMP-Activated Protein Kinase in Adipose Tissue Metabolism and Inflammation. Clin. Sci. 2013, 124, 491–507. [Google Scholar] [CrossRef] [Green Version]
- Yokokawa, T.; Sato, K.; Narusawa, R.; Kido, K.; Mori, R.; Iwanaka, N.; Hayashi, T.; Hashimoto, T. Dehydroepiandrosterone Activates 5′-Adenosine Monophosphate-Activated Protein Kinase and Suppresses Lipid Accumulation and Adipocyte Differentiation in 3T3-L1 Cells. Biochem. Biophys. Res. Commun. 2020, 528, 612–619. [Google Scholar] [CrossRef]
- Daval, M.; Foufelle, F.; Ferré, P. Functions of AMP-Activated Protein Kinase in Adipose Tissue. J. Physiol. 2006, 574, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.M. Regulation and Function of AMPK in Physiology and Diseases. Exp. Mol. Med. 2016, 48, e245. [Google Scholar] [CrossRef] [PubMed]
- Mannerås, L.; Cajander, S.; Holmäng, A.; Seleskovic, Z.; Lystig, T.; Lönn, M.; Stener-Victorin, E. A New Rat Model Exhibiting Both Ovarian and Metabolic Characteristics of Polycystic Ovary Syndrome. Endocrinology 2007, 148, 3781–3791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walters, K.A.; Bertoldo, M.J.; Handelsman, D.J. Evidence from Animal Models on the Pathogenesis of PCOS. Best Pract. Res. Clin. Endocrinol. Metab. 2018, 32, 271–281. [Google Scholar] [CrossRef]
- Milutinović, D.V.; Nikolić, M.; Veličković, N.; Djordjevic, A.; Bursać, B.; Nestorov, J.; Teofilović, A.; Antić, I.B.; Macut, J.B.; Zidane, A.S.; et al. Enhanced Inflammation without Impairment of Insulin Signaling in the Visceral Adipose Tissue of 5α-Dihydrotestosterone-Induced Animal Model of Polycystic Ovary Syndrome. Exp. Clin. Endocrinol. Diabetes 2017, 125, 522–529. [Google Scholar] [CrossRef]
- Nikolić, M.; Velicković, N.; Djordjevic, A.; Bursać, B.; Macut, D.; Antić, I.B.; Macut, J.B.; Matić, G.; Milutinović, D.V. 5α-Dihydrotestosterone Treatment Induces Metabolic Changes Associated with Polycystic Ovary Syndrome without Interfering with Hypothalamic Leptin and Glucocorticoid Signaling. Arch. Biol. Sci. 2016, 68, 473–481. [Google Scholar] [CrossRef]
- Šefčíková, Z.; Bujňáková, D.; Raček, L.; Kmet, V.; Mozeš, Š. Developmental Changes in Gut Microbiota and Enzyme Activity Predict Obesity Risk in Rats Arising From Reduced Nests. Physiol. Res. 2011, 8408, 337–346. [Google Scholar] [CrossRef]
- Caldwell, A.S.L.; Middleton, L.J.; Jimenez, M.; Desai, R.; McMahon, A.C.; Allan, C.M.; Handelsman, D.J.; Walters, K.A. Characterization of Reproductive, Metabolic and Endocrine Features of Polycystic Ovary Syndrome in Female Hyperandrogenic Mouse Models. Endocrinology 2014, 155, 3146. [Google Scholar] [CrossRef] [Green Version]
- Plagemann, A.; Harder, T.; Rake, A.; Melchior, K.; Rohde, W.; Dörner, G. Increased Number of Galanin-Neurons in the Paraventricular Hypothalamic Nucleus of Neonatally Overfed Weanling Rats. Brain Res. 1999, 818, 160–163. [Google Scholar] [CrossRef]
- Costa, V.M.G.; Andreazzi, A.E.; Bolotari, M.; Lade, C.G.; Guerra, M.O.; Peters, V.M. Effect of Postnatal Overfeeding on the Male and Female Wistar Rat Reproductive Parameters. J. Dev. Orig. Health Dis. 2019, 10, 667–675. [Google Scholar] [CrossRef]
- Behboudi-Gandevani, S.; Ramezani Tehrani, F.; Rostami Dovom, M.; Farahmand, M.; Bahri Khomami, M.; Noroozzadeh, M.; Kabir, A.; Azizi, F. Insulin Resistance in Obesity and Polycystic Ovary Syndrome: Systematic Review and Meta-Analysis of Observational Studies. Gynecol. Endocrinol. 2016, 32, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Hardy, O.T.; Czech, M.P.; Corvera, S. What Causes the Insulin Resistance Underlying Obesity? Curr. Opin. Endocrinol. Diabetes Obes. 2012, 19, 81–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortega, F.J.; Mayas, D.; Moreno-Navarrete, J.M.; Catalán, V.; Gómez-Ambrosi, J.; Esteve, E.; Rodriguez-Hermosa, J.I.; Ruiz, B.; Ricart, W.; Peral, B.; et al. The Gene Expression of the Main Lipogenic Enzymes Is Downregulated in Visceral Adipose Tissue of Obese Subjects. Obesity 2010, 18, 13–20. [Google Scholar] [CrossRef]
- Fernandez, C.; Schuhmann, K.; Herzog, R.; Fielding, B.; Frayn, K.; Shevchenko, A.; James, P.; Holm, C.; Ström, K. Altered Desaturation and Elongation of Fatty Acids in Hormone-Sensitive Lipase Null Mice. PLoS ONE 2011, 6, e21603. [Google Scholar] [CrossRef] [Green Version]
- Mitsuhashi, K.; Senmaru, T.; Fukuda, T.; Yamazaki, M.; Shinomiya, K.; Ueno, M.; Kinoshita, S.; Kitawaki, J.; Katsuyama, M.; Tsujikawa, M.; et al. Testosterone Stimulates Glucose Uptake and GLUT4 Translocation through LKB1/AMPK Signaling in 3T3-L1 Adipocytes. Endocrine 2016, 51, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, S.; Mihaylova, M.M.; Zheng, B.; Hou, X.; Jiang, B.; Park, O.; Luo, Z.; Lefai, E.; Shyy, J.Y.J.; et al. AMPK Phosphorylates and Inhibits SREBP Activity to Attenuate Hepatic Steatosis and Atherosclerosis in Diet-Induced Insulin-Resistant Mice. Cell Metab. 2011, 13, 376–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Després, J.P.; Lemieux, I. Abdominal Obesity and Metabolic Syndrome. Nature 2006, 444, 881–887. [Google Scholar] [CrossRef]
- Varlamov, O.; Bethea, C.L.; Roberts, C.T. Sex-Specific Differences in Lipid and Glucose Metabolism. Front. Endocrinol. 2014, 5, 241. [Google Scholar] [CrossRef] [Green Version]
- Varlamov, O.; Bishop, C.V.; Handu, M.; Takahashi, D.; Srinivasan, S.; White, A.; Roberts, C.T. Combined Androgen Excess and Western-Style Diet Accelerates Adipose Tissue Dysfunction in Young Adult, Female Nonhuman Primates. Hum. Reprod. 2017, 32, 1892–1902. [Google Scholar] [CrossRef]
- True, C.A.; Takahashi, D.L.; Burns, S.E.; Mishler, E.C.; Bond, K.R.; Wilcox, M.C.; Calhoun, A.R.; Bader, L.A.; Dean, T.A.; Ryan, N.D.; et al. Chronic Combined Hyperandrogenemia and Western-Style Diet in Young Female Rhesus Macaques Causes Greater Metabolic Impairments Compared to Either Treatment Alone. Hum. Reprod. 2017, 32, 1880–1891. [Google Scholar] [CrossRef]
- Heilbronn, L.; Smith, S.R.; Ravussin, E. Failure of Fat Cell Proliferation, Mitochondrial Function and Fat Oxidation Results in Ectopic Fat Storage, Insulin Resistance and Type II Diabetes Mellitus. Int. J. Obes. 2004, 28 (Suppl. 4), 12–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gauthier, M.S.; Pérusse, J.R.; Lavoie, M.È.; Sladek, R.; Madiraju, S.R.M.; Ruderman, N.B.; Coulombe, B.; Prentki, M.; Rabasa-Lhoret, R. Increased Subcutaneous Adipose Tissue Expression of Genes Involved in Glycerolipid-Fatty Acid Cycling in Obese Insulin-Resistant versus-Sensitive Individuals. J. Clin. Endocrinol. Metab. 2014, 99, E2518–E2528. [Google Scholar] [CrossRef] [PubMed]
- Silfen, M.E.; Denburg, M.R.; Manibo, A.M.; Lobo, R.A.; Jaffe, R.; Ferin, M.; Levine, L.S.; Oberfield, S.E. Early Endocrine, Metabolic, and Sonographic Characteristics of Polycystic Ovary Syndrome (PCOS): Comparison between Nonobese and Obese Adolescents. J. Clin. Endocrinol. Metab. 2003, 88, 4682–4688. [Google Scholar] [CrossRef] [Green Version]
- Cinti, S. The Adipose Organ at a Glance. DMM Dis. Model. Mech. 2012, 5, 588–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NL-Placebo | NL-DHT | SL-Placebo | SL-DHT | Two-Way ANOVA | |||
|---|---|---|---|---|---|---|---|
| Litter | DHT | Interaction | |||||
| Energy intake (kJ/day/cage) | 475.5 ± 10 | 467.6 ± 15.5 | 512.4 ± 13.6 | 531.1 ± 13.4 | <0.001 | ns | ns |
| Body mass (g) | 230.0 ± 4.6 | 241.0 ± 11.0 | 252.33 ± 4.2 | 283.33 ± 13.0 | <0.001 | <0.05 | ns |
| Mass of VAT (g) | 7.0 ± 0.6 | 8.4 ± 1.4 | 14.0 ± 1.4 | 12.0 ± 1.1 | <0.001 | ns | ns |
| VAT-to-body ratio (×100) | 3.0 ± 0.2 | 3.3 ± 0.4 | 5.3 ± 0.5 | 4.5 ± 0.4 | <0.001 | ns | ns |
| Mass of SAT (g) | 2.1 ± 0.3 | 1.8 ± 0.4 | 2.0 ± 0.17 | 2.2 ± 0.3 | ns | ns | ns |
| SAT-to-body ratio (×100) | 0.9 ± 0.1 | 0.7 ± 0.1 | 0.8 ± 0.1 | 0.8 ± 0.1 | ns | ns | ns |
| Mass of ovary (g) | 0.048 ± 0.003 | 0.027 ± 0.006 | 0.043 ± 0.002 | 0.025 ± 0.004 | ns | <0.001 | ns |
| Ovary-to-body ratio (×1000) | 0.21 ± 0.01 | 0.11 ± 0.03 | 0.17 ± 0.01 | 0.085 ± 0.01 | ns | <0.001 | ns |
| Mass of uterus (g) | 0.49 ± 0.06 | 0.2 ± 0.03 | 0.39 ± 0.05 | 0.24 ± 0.04 | ns | <0.001 | ns |
| Uterus-to-body ratio (×1000) | 2.1 ± 0.22 | 0.85 ± 0.14 | 1.6 ± 0.22 | 0.86 ± 0.16 | ns | <0.001 | ns |
| NL-Placebo | NL-DHT | SL-Placebo | SL-DHT | Two-Way ANOVA | |||
|---|---|---|---|---|---|---|---|
| Litter | DHT | Interaction | |||||
| Triglycerides (mmol/L) | 0.87 ± 0.09 | 0.88 ± 0.11 | 0.73 ± 0.03 | 0.93 ± 0.07 | ns | ns | ns |
| FFA (mmol/L) | 1.53 ± 0.29 | 1.22 ± 0.18 | 1.15 ± 0.21 | 1.20 ± 0.25 | ns | ns | ns |
| Glucose (mmol/L) | 6.00 ± 0.18 | 5.82 ± 0.14 | 5.85 ± 0.14 | 5.83 ± 0.16 | ns | ns | ns |
| Insulin (µU/mL) | 17.0 ± 0.7 | 19.0 ± 2.7 | 23.32 ± 1.2 | 36.0 ± 3.9 ***,###,†† | <0.001 | <0.01 | <0.05 |
| HOMA index | 4.5 ± 0.1 | 4.9 ± 0.6 | 6.1 ± 0.3 | 9.3 ± 1.1 ***,###,†† | <0.001 | <0.05 | <0.05 |
| ipGTT AUC | 1249 ±49.1 | 1266 ± 90.0 | 1442 ± 45.4 | 1578 ± 51.01 | <0.001 | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mićić, B.; Teofilović, A.; Djordjevic, A.; Veličković, N.; Macut, D.; Vojnović Milutinović, D. AMPK Activation Is Important for the Preservation of Insulin Sensitivity in Visceral, but Not in Subcutaneous Adipose Tissue of Postnatally Overfed Rat Model of Polycystic Ovary Syndrome. Int. J. Mol. Sci. 2022, 23, 8942. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23168942
Mićić B, Teofilović A, Djordjevic A, Veličković N, Macut D, Vojnović Milutinović D. AMPK Activation Is Important for the Preservation of Insulin Sensitivity in Visceral, but Not in Subcutaneous Adipose Tissue of Postnatally Overfed Rat Model of Polycystic Ovary Syndrome. International Journal of Molecular Sciences. 2022; 23(16):8942. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23168942
Chicago/Turabian StyleMićić, Bojana, Ana Teofilović, Ana Djordjevic, Nataša Veličković, Djuro Macut, and Danijela Vojnović Milutinović. 2022. "AMPK Activation Is Important for the Preservation of Insulin Sensitivity in Visceral, but Not in Subcutaneous Adipose Tissue of Postnatally Overfed Rat Model of Polycystic Ovary Syndrome" International Journal of Molecular Sciences 23, no. 16: 8942. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23168942