
Identification and Characterization of the Larval Settlement Pheromone Protein Components in Adult Shells of Crassostrea gigas: A Novel Function of Shell Matrix Proteins

 ,
,
Abstract
:1. Introduction
2. Results
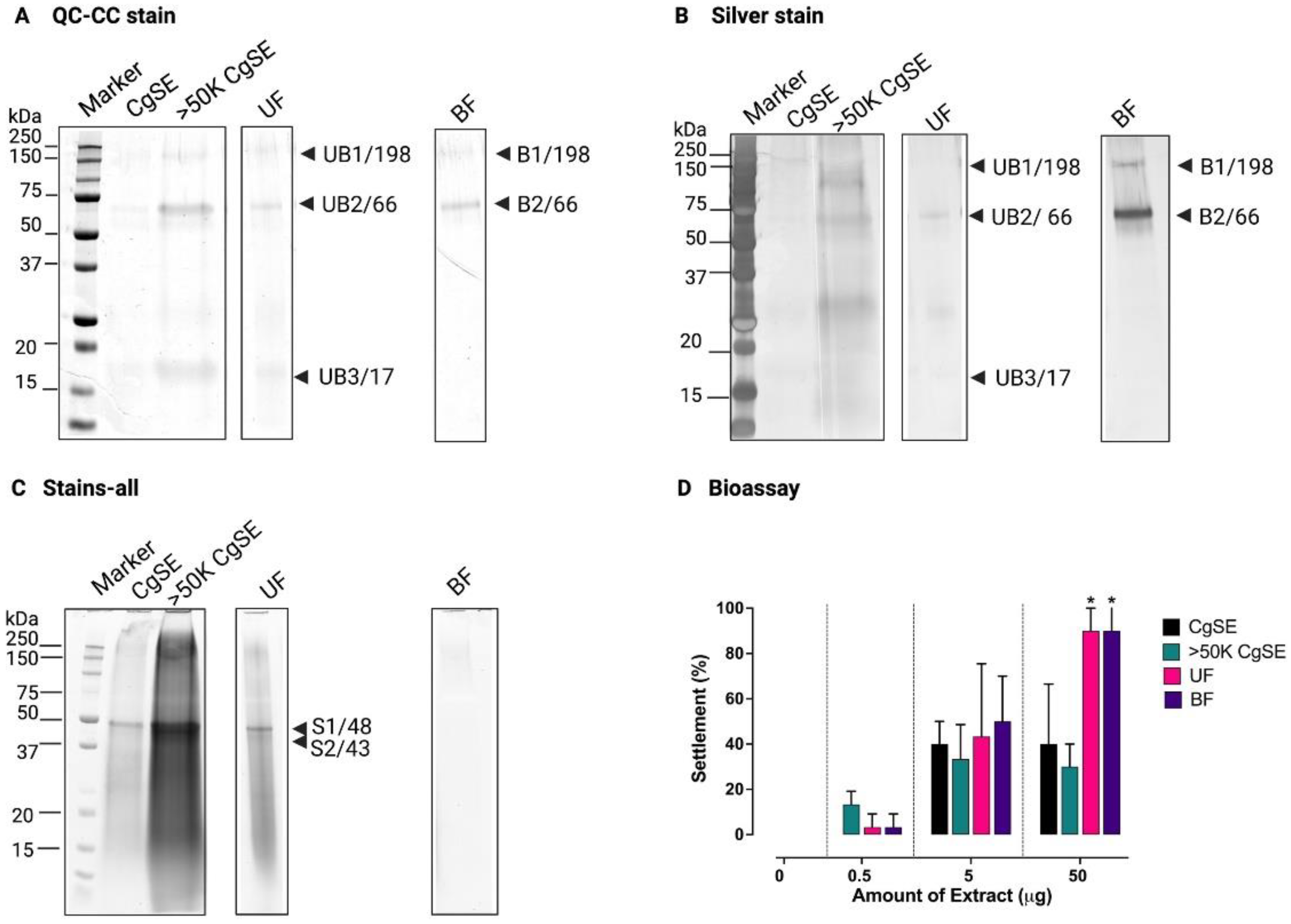
2.1. Protein Separation and Identification of the Settlement Inducing Cues from Crassostrea gigas EDTA-Soluble Shell Matrix Proteins
2.1.1. Protein Separation
2.1.2. Identification by Mass Spectrometry
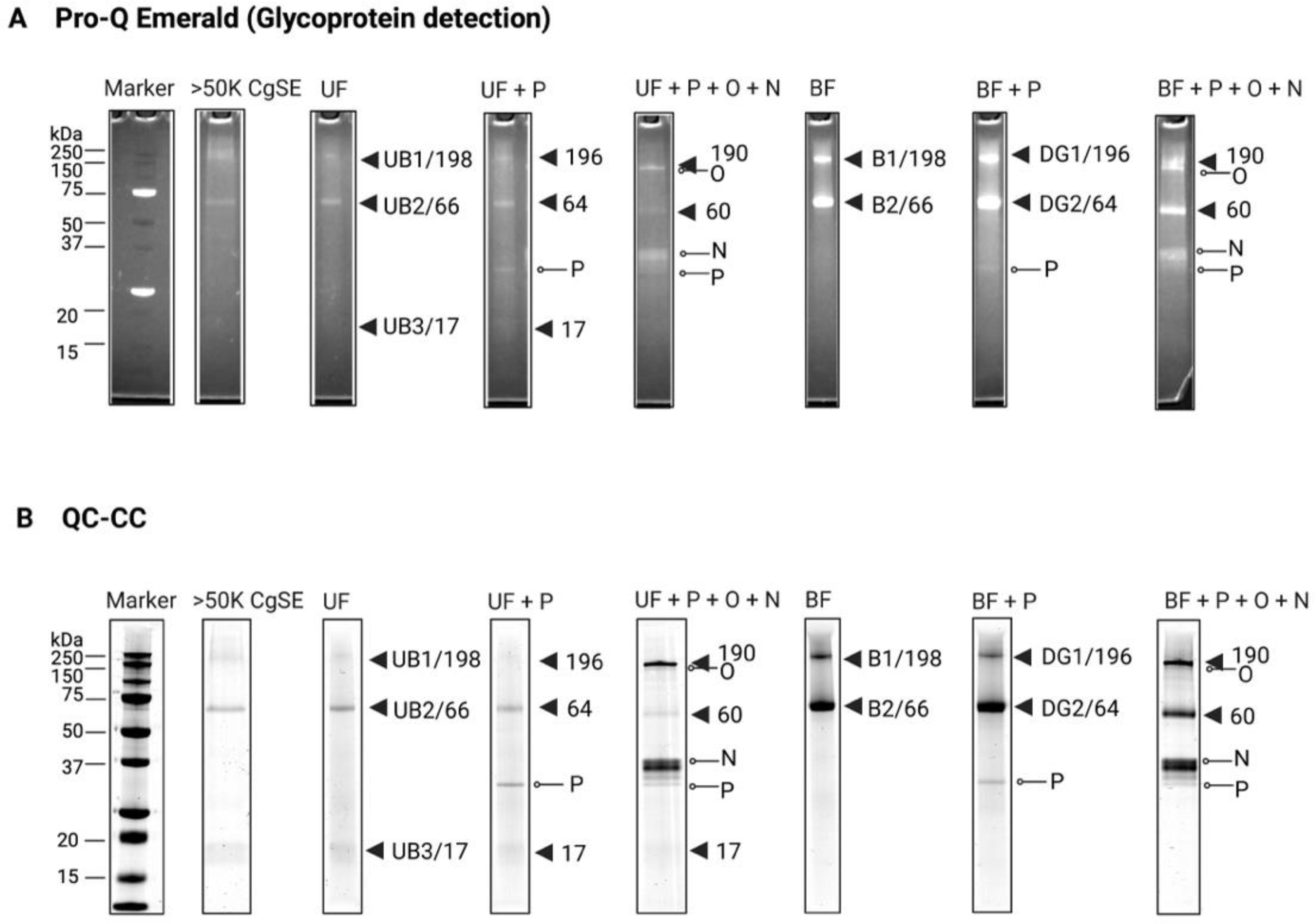
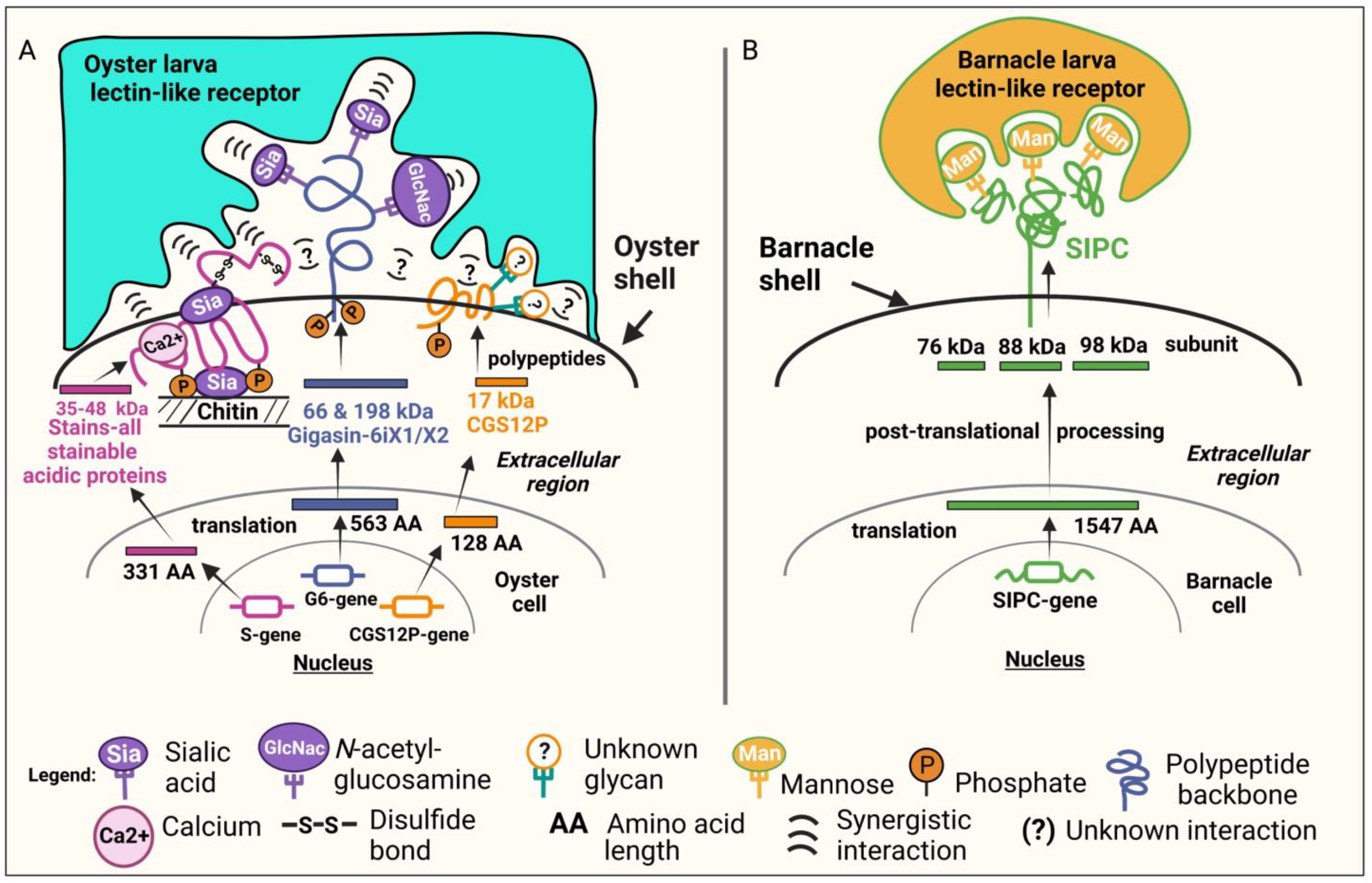
2.2. Biochemical and Molecular Characterization of the Settlement-Inducing Cues from C. gigas EDTA-Soluble Shell Matrix Proteins
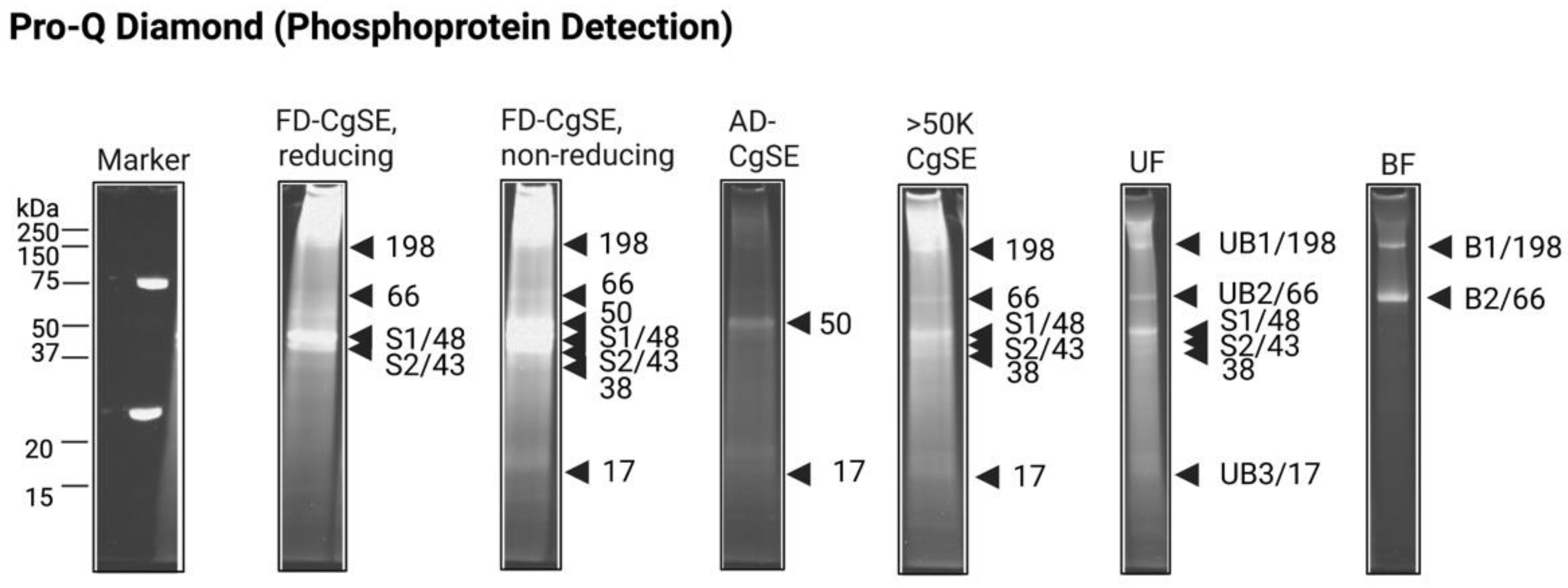
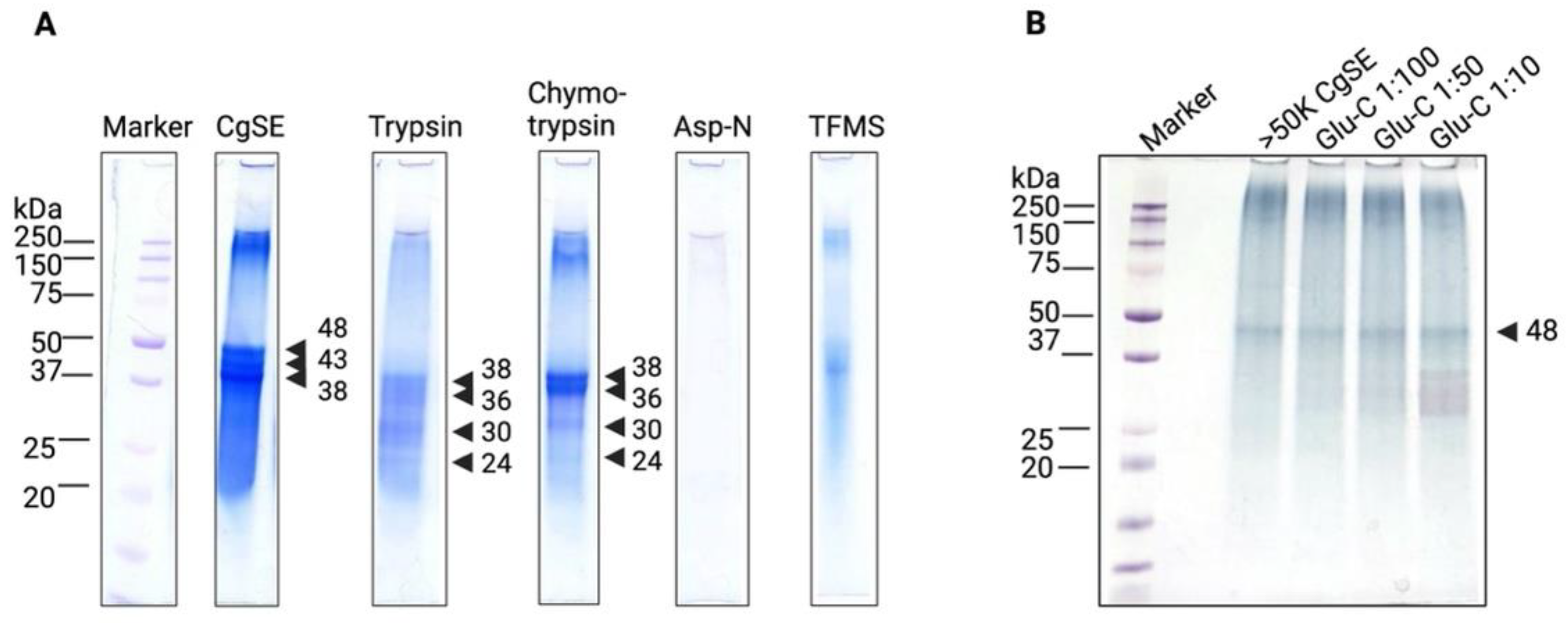
2.2.1. Biochemical Characterization
Post-Translational Modifications (PTMs)
Characterization of Stains-All-Stainable Acidic Proteins by Protease and Chemical Deglycosylation Treatments
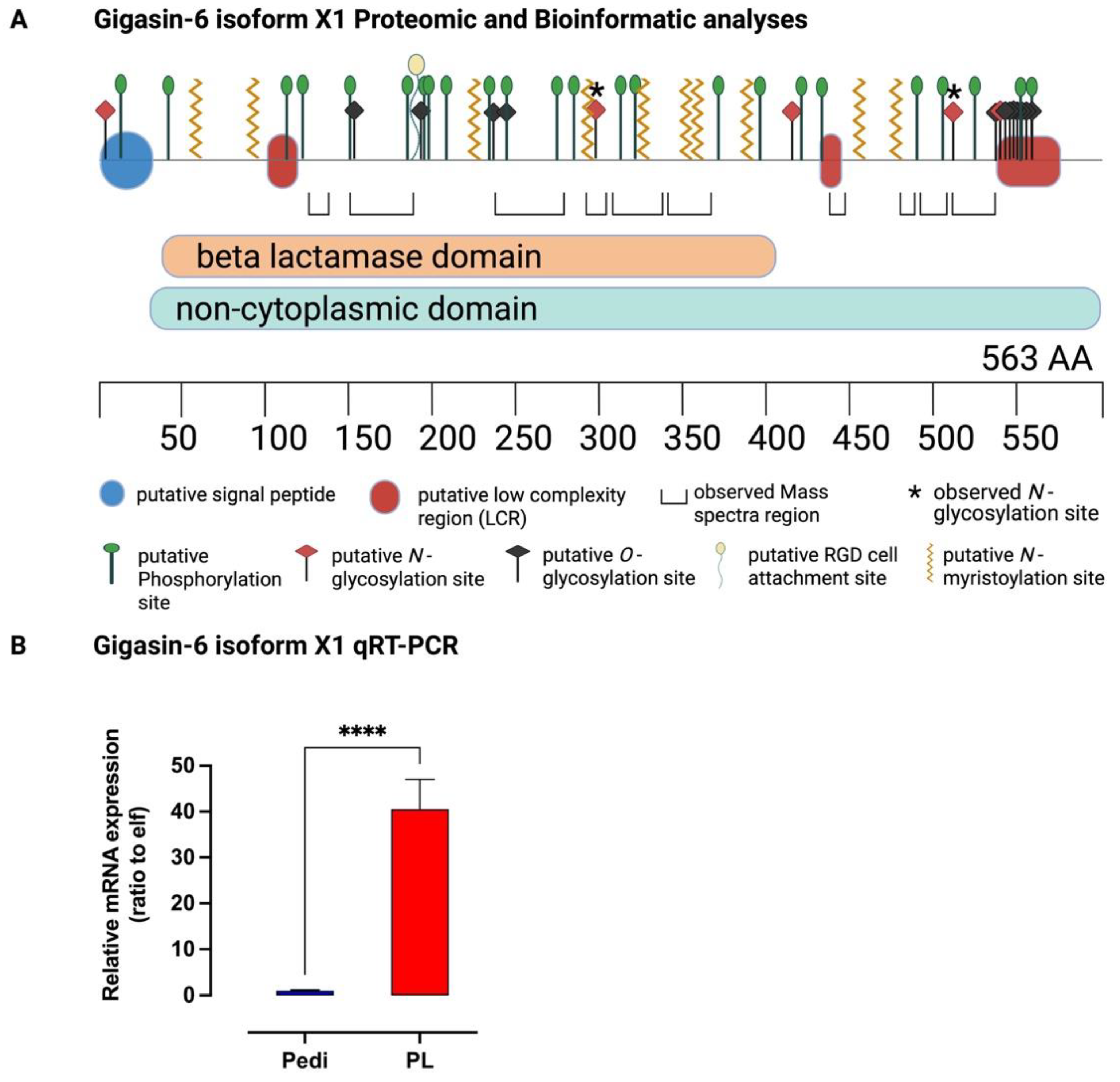
2.2.2. Molecular Characterization
Gigasin-6 isoform X1 and/or X2
Stains-All-Stainable Acidic Proteins
Surface Protein P12p-like
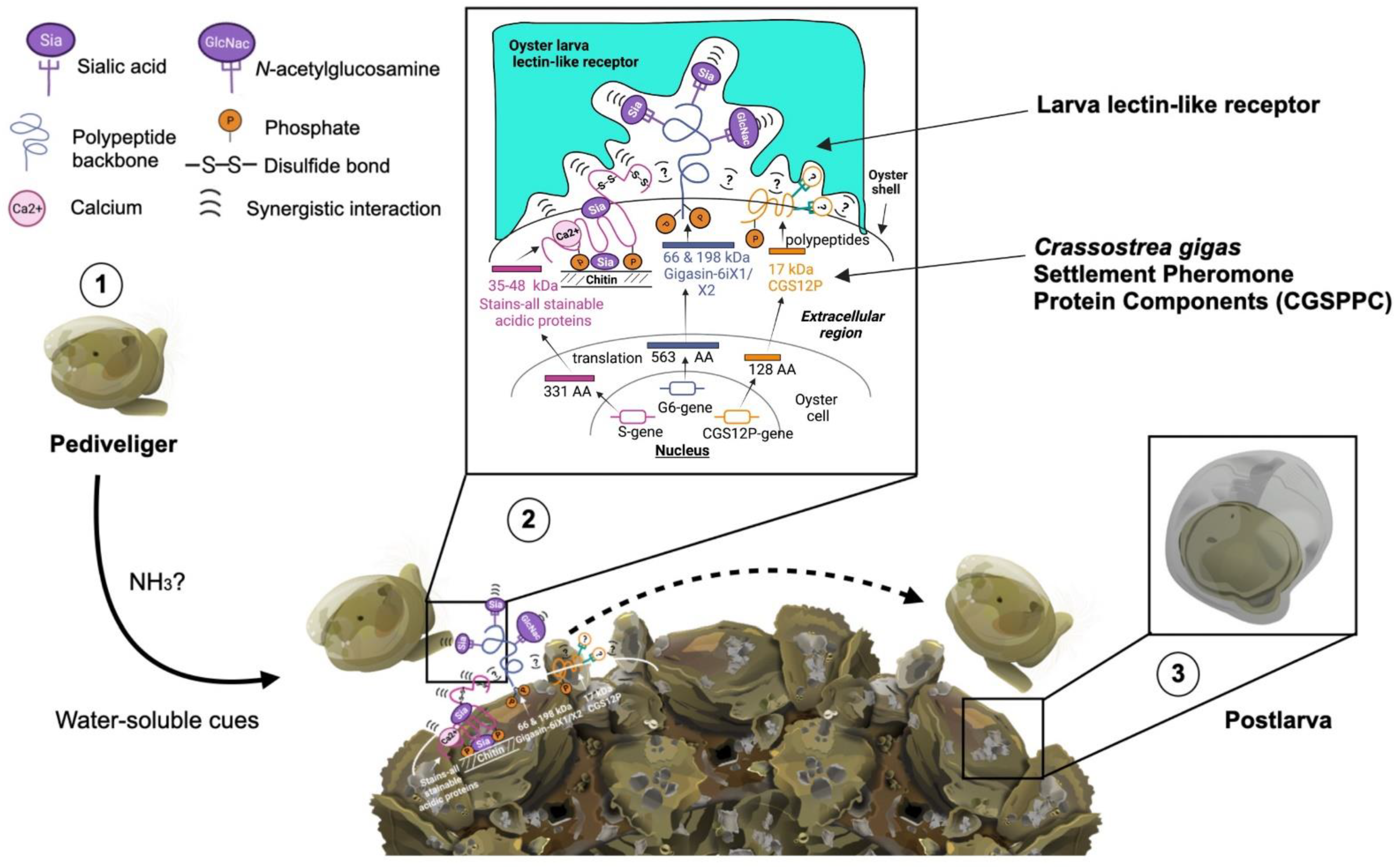
3. Discussion
4. Materials and Methods
4.1. Spawning and Larval Culture
4.2. Extraction of Shell Matrices
4.3. First Protein Separation Approach
4.3.1. Fractionation by Ultrafiltration
4.3.2. Gel Filtration of >50K CgSE
4.3.3. Separation of F2 Proteins
4.4. Protease and TFMS Treatment
4.5. Second Protein Separation Approach
4.5.1. Wheat Germ Agglutinin (WGA) Agarose-Bound Lectin Affinity Chromatography
4.5.2. SDS-PAGE Analysis of WGA Affinity Chromatography-Eluted Fractions
4.6. Glycoprotein Detection
4.7. Phosphoprotein Detection
4.8. Mass Spectrometry Analyses
4.9. Localization of N-glycosylation Sites by Mass Spectrometry Analysis
4.10. Characterization of Protein Sequences
4.11. Molecular Characterization of Gigasin-6 Isoform X1
4.12. Statistical Analyses and Visualization
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hedgecock, D.; Gaffney, P.M.; Goulletquer, P.; Guo, X.; Reece, K.; Warr, G.W. The Case for Sequencing the Pacific Oyster Genome. J. Shellfish Res. 2005, 24, 429–441. [Google Scholar] [CrossRef]
- Zhang, G.; Fang, X.; Guo, X.; Li, L.; Luo, R.; Xu, F.; Yang, P.; Zhang, L.; Wang, X.; Qi, H.; et al. The Oyster Genome Reveals Stress Adaptation and Complexity of Shell Formation. Nature 2012, 490, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Grabowski, J.H.; Brumbaugh, R.D.; Conrad, R.F.; Keeler, A.G.; Opaluch, J.J.; Peterson, C.H.; Piehler, M.F.; Powers, S.P.; Smyth, A.R. Economic Valuation of Ecosystem Services Provided by Oyster Reefs. Bioscience 2012, 62, 900–909. [Google Scholar] [CrossRef]
- Gentry, R.R.; Alleway, H.K.; Bishop, M.J.; Gillies, C.L.; Waters, T.; Jones, R. Exploring the Potential for Marine Aquaculture to Contribute to Ecosystem Services. Rev. Aquac. 2020, 12, 499–512. [Google Scholar] [CrossRef]
- Ridlon, A.D.; Wasson, K.; Waters, T.; Adams, J.; Donatuto, J.; Fleener, G.; Froehlich, H.; Govender, R.; Kornbluth, A.; Lorda, J.; et al. Conservation Aquaculture as a Tool for Imperiled Marine Species: Evaluation of Opportunities and Risks for Olympia Oysters, Ostrea lurida. PLoS ONE 2021, 16, e0252810. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Zhang, G. Transcriptomic and Proteomic Dynamics during Metamorphosis of Pacific Oyster Crassostrea gigas. bioRxiv 2020. [Google Scholar] [CrossRef]
- Fitt, W.K.; Coon, S.L.; Walch, M.; Weiner, R.M.; Colwell, R.R.; Bonar, D.B. Settlement Behavior and Metamorphosis of Oyster Larvae (Crassostrea gigas) in Response to Bacterial Supernatants. Mar. Biol. 1990, 106, 389–394. [Google Scholar] [CrossRef]
- Coon, S.L.; Bonar, D.B.; Weiner, R.M. Induction of Settlement and Metamorphosis of the Pacific Oyster, Crassostrea gigas (Thunberg), by L-DOPA and Catecholamines. J. Exp. Mar. Bio. Ecol. 1985, 94, 211–221. [Google Scholar] [CrossRef]
- Joyce, A.; Vogeler, S. Molluscan Bivalve Settlement and Metamorphosis: Neuroendocrine Inducers and Morphogenetic Responses. Aquaculture 2018, 487, 64–82. [Google Scholar] [CrossRef]
- Yurchenko, O.V.; Skiteva, O.I.; Voronezhskaya, E.E.; Dyachuk, V.A. Nervous System Development in the Pacific Oyster, Crassostrea gigas (Mollusca: Bivalvia). Front. Zool. 2018, 15, 10. [Google Scholar] [CrossRef] [Green Version]
- Marie, B.; Zanella-Cléon, I.; Guichard, N.; Becchi, M.; Marin, F. Novel Proteins from the Calcifying Shell Matrix of the Pacific Oyster Crassostrea gigas. Mar. Biotechnol. 2011, 13, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Song, X.; WANG, T.; Zhu, Q.; Miao, G.; Chen, Y.; Fang, X.; Que, H.; Li, L.; Zhang, G. Evolution and Functional Analysis of the Pif97 Gene of the Pacific Oyster Crassostrea gigas. Curr. Zool. 2013, 59, 109–115. [Google Scholar] [CrossRef]
- De Wit, P.; Durland, E.; Ventura, A.; Langdon, C.J. Gene Expression Correlated with Delay in Shell Formation in Larval Pacific Oysters (Crassostrea gigas) Exposed to Experimental Ocean Acidification Provides Insights into Shell Formation Mechanisms. BMC Genom. 2018, 19, 160. [Google Scholar] [CrossRef] [PubMed]
- Beck, M.W.; Brumbaugh, R.D.; Airoldi, L.; Carranza, A.; Coen, L.D.; Crawford, C.; Defeo, O.; Edgar, G.J.; Hancock, B.; Kay, M.C.; et al. Oyster Reefs at Risk and Recommendations for Conservation, Restoration, and Management. Bioscience 2011, 61, 107–116. [Google Scholar] [CrossRef]
- Hadfield, M.; Paul, V. Natural Chemical Cues for Settlement and Metamorphosis of Marine-Invertebrate Larvae. In Marine Chemical Ecology; McClintok, J.B., Baker, B.J., Eds.; CRC Press: Boca Raton, FL, USA, 2001; pp. 431–461. ISBN 9781420036602. [Google Scholar]
- Prendergast, G.S. Settlement and Behaviour of Marine Fouling Organisms. In Biofouling; Wiley-Blackwell: Oxford, UK, 2010; pp. 30–59. ISBN 9781444315462. [Google Scholar]
- Hadfield, M.G. Biofilms and Marine Invertebrate Larvae: What Bacteria Produce That Larvae Use to Choose Settlement Sites. Ann. Rev. Mar. Sci. 2011, 3, 453–470. [Google Scholar] [CrossRef]
- Highsmith, R.C. Induced Settlement and Metamorphosis of Sand Dollar (Dendraster ecentricus) Larvae in Predator-Free Sites: Adult Sand Dollar Beds. Ecology 1982, 63, 329–337. [Google Scholar] [CrossRef]
- McQuaid, C.D.; Miller, K. Larval Supply and Dispersal. In Biofouling; Wiley-Blackwell: Oxford, UK, 2010; pp. 16–29. ISBN 9781444315462. [Google Scholar]
- Thorson, G. Reproductive and Larval Ecology of Marine Bottom Invertebrates. Biol. Rev. 1950, 25, 1–45. [Google Scholar] [CrossRef]
- Hay, M.E. Marine Chemical Ecology: Chemical Signals and Cues Structure Marine Populations, Communities, and Ecosystems. Ann. Rev. Mar. Sci. 2009, 1, 193–212. [Google Scholar] [CrossRef]
- Bayne, B.L. The Gregarious Behaviour of the Larvae of Ostrea edulis L. at Settlement. J. Mar. Biol. Assoc. U. K. 1969, 49, 327–356. [Google Scholar] [CrossRef]
- Vasquez, H.E.; Hashimoto, K.; Yoshida, A.; Hara, K.; Imai, C.C.; Kitamura, H.; Satuito, C.G. A Glycoprotein in Shells of Conspecifics Induces Larval Settlement of the Pacific Oyster Crassostrea gigas. PLoS ONE 2013, 8, e82358. [Google Scholar] [CrossRef] [Green Version]
- Vasquez, H.E.; Hashimoto, K.; Kitamura, H.; Satuito, C.G. Wheat Germ Agglutinin-Binding Glycoprotein Extract from Shells of Conspecifics Induces Settlement of Larvae of the Pacific Oyster Crassostrea gigas (Thunberg). J. Shellfish Res. 2014, 33, 415–423. [Google Scholar] [CrossRef]
- Diederich, S. Differential Recruitment of Introduced Pacific Oysters and Native Mussels at the North Sea Coast: Coexistence Possible? J. Sea Res. 2005, 53, 269–281. [Google Scholar] [CrossRef]
- Sedanza, M.G.; Kim, H.-J.; Seposo, X.; Yoshida, A.; Yamaguchi, K.; Satuito, C.G. Regulatory Role of Sugars on the Settlement Inducing Activity of a Conspecific Cue in Pacific Oyster Crassostrea gigas. Int. J. Mol. Sci. 2021, 22, 3273. [Google Scholar] [CrossRef] [PubMed]
- Crisp, D.J. Chemical Factors Inducing Settlement in Crassostrea virginica (Gmelin). J. Anim. Ecol. 1967, 36, 329–335. [Google Scholar] [CrossRef]
- Poirier, L.A.; Clements, J.C.; Davidson, J.D.P.; Miron, G.; Davidson, J.; Comeau, L.A. Sink before You Settle: Settlement Behaviour of Eastern Oyster (Crassostrea virginica) Larvae on Artificial Spat Collectors and Natural Substrate. Aquac. Rep. 2019, 13, 100181. [Google Scholar] [CrossRef]
- Tamburri, M.N.; Luckenbach, M.W.; Breitburg, D.L.; Bonniwell, S.M. Settlement of Crassostrea ariakensis Larvae: Effects of Substrate, Biofilms, Sediment and Adult Chemical Cues. J. Shellfish Res. 2008, 27, 601–608. [Google Scholar] [CrossRef]
- Hirata, Y.; Tamura, Y.; Nagasawa, K. Influence of Presoaking Conditions of Spat Collectors in Seawater Containing Adult Pacific Oysters (Crassostrea gigas) on Their Larval Settlement. Nippon. Suisan Gakkaishi 2008, 74, 1017–1023. [Google Scholar] [CrossRef]
- Hirata, Y. Advanced Techniques for Artificial Setting of Oyster Crassostrea gigas. Nippon Suisan Gakkaishi 2005, 71, 725–728. [Google Scholar] [CrossRef]
- Hirata, Y. Effective Pretreatments of Spat-Collectors for Induced Settlement of the Pacific Oyster, Crassostrea gigas. Nippon Suisan Gakkaishi 1998, 64, 610–617. [Google Scholar] [CrossRef]
- Zimmer-Faust, R.K.; Tamburri, M.N. Chemical Identity and Ecological Implications of a Waterborne, Larval Settlement Cue. Limnol. Oceanogr. 1994, 39, 1075–1087. [Google Scholar] [CrossRef]
- Turner, E.J.; Zimmer-Faust, R.K.; Palmer, M.A.; Luckenbach, M.; Pentchef, N.D. Settlement of Oyster (Crassostrea virginica) Larvae: Effects of Water Flow and a Water-soluble Chemical Cue. Limnol. Oceanogr. 1994, 39, 1579–1593. [Google Scholar] [CrossRef]
- Carroll, J.M.; Riddle, K.; Woods, K.E.; Finelli, C.M. Recruitment of the Eastern Oyster, Crassostrea virginica, in Response to Settlement Cues and Predation in North Carolina. J. Exp. Mar. Biol. Ecol. 2015, 463, 1–7. [Google Scholar] [CrossRef]
- Pascual, M.S.; Zampatti, E.A. Evidence of a Chemically Mediated Adult-Larval Interaction Triggering Settlement in Ostrea puelchana: Applications in Hatchery Production. Aquaculture 1995, 133, 33–44. [Google Scholar] [CrossRef]
- Veitch, F.P.; Hidu, H. Gregarious Setting in the American Oyster Crassostrea virginica Gmelin: I. Properties of a Partially Purified “Setting Factor.” Chesap. Sci. 1971, 12, 173–178. [Google Scholar] [CrossRef]
- Coon, S.L.; Walch, M.; Fitt, W.K.; Weiner, R.M.; Bonar, D.B. Ammonia Induces Settlement Behavior in Oyster Larvae. Biol. Bull. 1990, 179, 297–303. [Google Scholar] [CrossRef]
- Myers, J.M.; Veis, A.; Sabsay, B.; Wheeler, A.P. A Method for Enhancing the Sensitivity and Stability of Stains-All for Phosphoproteins Separated in Sodium Dodecyl Sulfate-Polyacrylamide Gels. Anal. Biochem. 1996, 240, 300–302. [Google Scholar] [CrossRef]
- Johnstone, M.B.; Wheeler, A.P.; Falwell, E.P.; Staton, M.E.; Saski, C.A.; Mount, A.S. Folian-Cv1 Is a Member of a Highly Acidic Phosphoprotein Class Derived From the Foliated Layer of the Eastern Oyster (Crassostrea virginica) Shell and Identified in Hemocytes and Mantle. Front. Mar. Sci. 2019, 6, 366. [Google Scholar] [CrossRef]
- Marin, F.; Luquet, G. Molluscan Shell Proteins. Comptes Rendus Palevol 2004, 3, 469–492. [Google Scholar] [CrossRef]
- Marin, F.; Le Roy, N.; Marie, B. The Formation and Mineralization of Mollusk Shell. Front. Biosci.-Sch. 2012, 4, 1099–1125. [Google Scholar] [CrossRef]
- Gotliv, B.-A.; Addadi, L.; Weiner, S. Mollusk Shell Acidic Proteins: In Search of Individual Functions. Chembiochem 2003, 4, 522–529. [Google Scholar] [CrossRef]
- Rivera-Pérez, C.; Hernández-Saavedra, N.Y. Review: Post-Translational Modifications of Marine Shell Matrix Proteins. Comp. Biochem. Physiol. B. Biochem. Mol. Biol. 2021, 256, 110641. [Google Scholar] [CrossRef] [PubMed]
- Dreanno, C.; Matsumura, K.; Dohmae, N.; Takio, K.; Hirota, H.; Kirby, R.R.; Clare, A.S. An α 2-Macroglobulin-like Protein Is the Cue to Gregarious Settlement of the Barnacle Balanus amphitrite. Proc. Natl. Acad. Sci. USA 2006, 103, 14396–14401. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, K.; Nagano, M.; Fusetani, N. Purification of a Larval Settlement-Inducing Protein Complex (SIPC) of the Barnacle, Balanus amphitrite. J. Exp. Zool. 1998, 281, 12–20. [Google Scholar] [CrossRef]
- Cabra, V.; Arreguin, R.; Vazquez-Duhalt, R.; Farres, A. Effect of Temperature and PH on the Secondary Structure and Processes of Oligomerization of 19 KDa Alpha-Zein. Biochim. Biophys. Acta-Proteins Proteom. 2006, 1764, 1110–1118. [Google Scholar] [CrossRef] [PubMed]
- Tatsuno, R.; Yamaguchi, K.; Takatani, T.; Arakawa, O. RT-PCR- and MALDI-TOF Mass Spectrometry-Based Identification and Discrimination of Isoforms Homologous to Pufferfish Saxitoxin- and Tetrodotoxin-Binding Protein in the Plasma of Non-Toxic Cultured Pufferfish (Takifugu rubripes). Biosci. Biotechnol. Biochem. 2013, 77, 208–212. [Google Scholar] [CrossRef]
- Choi, Y.H.; Yamaguchi, K.; Oda, T.; Nam, T.J. Chemical and Mass Spectrometry Characterization of the Red Alga Pyropia yezoensis Chemoprotective Protein (PYP): Protective Activity of the N-Terminal Fragment of PYP1 against Acetaminophen-Induced Cell Death in Chang Liver Cells. Int. J. Mol. Med. 2015, 35, 271–276. [Google Scholar] [CrossRef]
- Magnelli, P.E.; Bielik, A.M.; Guthrie, E.P. Identification and Characterization of Protein Glycosylation Using Specific Endo- and Exoglycosidases. J. Vis. Exp. 2011, e3749. [Google Scholar] [CrossRef]
- Dwek, R.A.; Edge, C.J.; Harvey, D.J.; Wormald, M.R.; Parekh, R.B. Analysis of Glycoprotein-Associated Oligosaccharides. Annu. Rev. Biochem. 1993, 62, 65–100. [Google Scholar] [CrossRef]
- Marin, F. Mollusc Shellomes: Past, Present and Future. J. Struct. Biol. 2020, 212, 107583. [Google Scholar] [CrossRef]
- Marie, B.; Luquet, G.; Pais De Barros, J.-P.; Guichard, N.; Morel, S.; Alcaraz, G.; Bollache, L.; Marin, F. The Shell Matrix of the Freshwater Mussel Unio pictorum (Paleoheterodonta, Unionoida). FEBS J. 2007, 274, 2933–2945. [Google Scholar] [CrossRef]
- Sellinger, O.Z.; Wolfson, M.F. Carboxylmethylation Affects the Proteolysis of Myelin Basic Protein by Staphylococcus aureus V8 Proteinase. Biochim. Biophys. Acta-Protein Struct. Mol. Enzymol. 1991, 1080, 110–118. [Google Scholar] [CrossRef]
- Miyashita, T.; Takagi, R.; Okushima, M.; Nakano, S.; Miyamoto, H.; Nishikawa, E.; Matsushiro, A. Complementary DNA Cloning and Characterization of Pearlin, a New Class of Matrix Protein in the Nacreous Layer of Oyster Pearls. Mar. Biotechnol. 2000, 2, 409–418. [Google Scholar] [CrossRef]
- Campbell, K.P.; MacLennan, D.H.; Jorgensen, A.O. Staining of the Ca2+-Binding Proteins, Calsequestrin, Calmodulin, Troponin C, and S-100, with the Cationic Carbocyanine Dye “Stains-All”. J. Biol. Chem. 1983, 258, 11267–11273. [Google Scholar] [CrossRef]
- Mouchi, V.; Lartaud, F.; Guichard, N.; Immel, F.; de Rafélis, M.; Broussard, C.; Crowley, Q.G.; Marin, F. Chalky versus Foliated: A Discriminant Immunogold Labelling of Shell Microstructures in the Edible Oyster Crassostrea gigas. Mar. Biol. 2016, 163, 256. [Google Scholar] [CrossRef]
- León, I.R.; da Costa Neves-Ferreira, A.G.; da Rocha, S.L.G.; de Oliveira Trugilho, M.R.; Perales, J.; Valente, R.H. Using Mass Spectrometry to Explore the Neglected Glycan Moieties of the Antiophidic Proteins DM43 and DM64. Proteomics 2012, 12, 2753–2765. [Google Scholar] [CrossRef]
- Sedanza, M.G.; Alshaweesh, J.; Gao, Y.; Yoshida, A.; Kim, H.; Yamaguchi, K.; Satuito, C.G. Transcriptome Dynamics of an Oyster Larval Response to a Conspecific Cue-Mediated Settlement Induction in the Pacific Oyster Crassostrea gigas. Diversity 2022, 14, 559. [Google Scholar] [CrossRef]
- Schmitt, P.; Gueguen, Y.; Desmarais, E.; Bachère, E.; de Lorgeril, J. Molecular Diversity of Antimicrobial Effectors in the Oyster Crassostrea gigas. BMC Evol. Biol. 2010, 10, 23. [Google Scholar] [CrossRef]
- Pancsa, R.; Schad, E.; Tantos, A.; Tompa, P. Emergent Functions of Proteins in Non-Stoichiometric Supramolecular Assemblies. Biochim. Biophys. acta. Proteins Proteom. 2019, 1867, 970–979. [Google Scholar] [CrossRef]
- Wojtas, M.; Dobryszycki, P.; Oyhar, A. Intrinsically Disordered Proteins in Biomineralization. In Advanced Topics in Biomineralization; InTech: Rijeka, Croatia, 2012; pp. 1–32. [Google Scholar]
- Alberts, B.; Johnson, A.; Lewis, J.; Morgan, D.; Raff, M.; Roberts, K.; Walter, P. Molecular Biology of the Cell, 6th ed.; Wilson, J., Hunt, T., Eds.; W.W. Norton & Company: New York, NY, USA, 2017; ISBN 9781315735368. [Google Scholar]
- Tompa, P. The Interplay between Structure and Function in Intrinsically Unstructured Proteins. FEBS Lett. 2005, 579, 3346–3354. [Google Scholar] [CrossRef]
- Totten, D.K.; Davidson, F.D.; Wyckoff, R.W. Amino-Acid Composition of Heated Oyster Shells. Proc. Natl. Acad. Sci. USA 1972, 69, 784–785. [Google Scholar] [CrossRef] [Green Version]
- Tommasone, S.; Allabush, F.; Tagger, Y.K.; Norman, J.; Köpf, M.; Tucker, J.H.R.; Mendes, P.M. The Challenges of Glycan Recognition with Natural and Artificial Receptors. Chem. Soc. Rev. 2019, 48, 5488–5505. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.E.; Drickamer, K.; Schnaar, R.L.; Etzler, M.E.; Varki, A. Discovery and Classification of Glycan-Binding Proteins. In Essentials of Glycobiology [Internet]; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2017; ISBN 9780879697709. [Google Scholar]
- Neal, A.L.; Yule, A.B. The Effects of Dissolved Sugars Upon the Temporary Adhesion of Barnacle Cyprids. J. Mar. Biol. Assoc. U. K. 1996, 76, 649–655. [Google Scholar] [CrossRef]
- Thiyagarajan, V. A Review on the Role of Chemical Cues in Habitat Selection by Barnacles: New Insights from Larval Proteomics. J. Exp. Mar. Bio. Ecol. 2010, 392, 22–36. [Google Scholar] [CrossRef]
- Thiyagarajan, V.; Wong, T.; Qian, P.-Y. 2D Gel-Based Proteome and Phosphoproteome Analysis During Larval Metamorphosis in Two Major Marine Biofouling Invertebrates. J. Proteome Res. 2009, 8, 2708–2719. [Google Scholar] [CrossRef]
- Hadfield, M.G.; Carpizo-Ituarte, E.J.; Del Carmen, K.; Nedved, B.T. Metamorphic Competence, a Major Adaptive Convergence in Marine Invertebrate Larvae. Am. Zool. 2001, 41, 1123–1131. [Google Scholar] [CrossRef]
- Calado, R.; Leal, M.C. Trophic Ecology of Benthic Marine Invertebrates with Bi-Phasic Life Cycles: What Are We Still Missing? Adv. Mar. Biol. 2015, 71, 1–70. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, K.; Mori, S.; Nagano, M.; Fusetani, N. Lentil Lectin Inhibits Adult Extract-Induced Settlement of the Barnacle, Balanus amphitrite. J. Exp. Zool. 1998, 280, 213–219. [Google Scholar] [CrossRef]
- Khandeparker, L.; Anil, A.C. Role of Conspecific Cues and Sugars in the Settlement of Cyprids of the Barnacle, Balanus amphitrite. J. Zool. 2011, 284, 206–214. [Google Scholar] [CrossRef]
- Mitchell, R. Colonization by Higher Organisms. In Microbial Adhesion and Aggregation; Marshall, K.C., Ed.; Springer: Berlin/Heidelberg, Germany, 1984; pp. 189–200. [Google Scholar]
- Tamburri, M.N.; Zimmer-Faust, R.K.; Tamplin, M.L. Natural Sources and Properties of Chemical Inducers Mediating Settlement of Oyster Larvae: A Re-Examination. Biol. Bull. 1992, 183, 327–338. [Google Scholar] [CrossRef]
- Bonar, D.B.; Coon, S.L.; Walch, M.; Weiner, R.M.; Fitt, W. Control of Oyster Settlement and Metamorphosis by Endogenous and Exogenous Chemical Cues. Bull. Mar. Sci. 1990, 46, 484–498. [Google Scholar]
- Steinberg, P.D.; De Nys, R.; Kjelleberg, S. Chemical Cues for Surface Colonization. J. Chem. Ecol. 2002, 28, 1935–1951. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Hancock, W.S. Approach to the Comprehensive Analysis of Glycoproteins Isolated from Human Serum Using a Multi-Lectin Affinity Column. J. Chromatogr. A 2004, 1053, 79–88. [Google Scholar] [CrossRef]
- Unno, H.; Nakamura, A.; Mori, S.; Goda, S.; Yamaguchi, K.; Hiemori, K.; Tateno, H.; Hatakeyama, T. Identification, Characterization, and X-Ray Crystallographic Analysis of a Novel Type of Lectin AJLec from the Sea Anemone Anthopleura japonica. Sci. Rep. 2018, 8, 11516. [Google Scholar] [CrossRef]
- Yamaguchi, K. Preparation and Proteomic Analysis of Chloroplast Ribosomes. In Chloroplast Research in Arabidopsis; Methods in Molecular Biology; Springer; Humana Press: Totowa, NJ, USA, 2011; Volume 775. [Google Scholar] [CrossRef]
- Mann, K.; Edsinger-Gonzales, E.; Mann, M. In-Depth Proteomic Analysis of a Mollusc Shell: Acid-Soluble and Acid-Insoluble Matrix of the Limpet Lottia gigantea. Proteome Sci. 2012, 10, 28. [Google Scholar] [CrossRef]
- Teufel, F.; Almagro Armenteros, J.J.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 Predicts All Five Types of Signal Peptides Using Protein Language Models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef]
- Blum, M.; Chang, H.Y.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro Protein Families and Domains Database: 20 Years On. Nucleic Acids Res. 2021, 49, D344–D354. [Google Scholar] [CrossRef]
- De Castro, E.; Sigrist, C.J.A.; Gattiker, A.; Bulliard, V.; Langendijk-Genevaux, P.S.; Gasteiger, E.; Bairoch, A.; Hulo, N. ScanProsite: Detection of PROSITE Signature Matches and ProRule-Associated Functional and Structural Residues in Proteins. Nucleic Acids Res. 2006, 34, W362–W365. [Google Scholar] [CrossRef]
- Schultz, J.; Milpetz, F.; Bork, P.; Ponting, C.P. SMART, a Simple Modular Architecture Research Tool: Identification of Signaling Domains. Proc. Natl. Acad. Sci. USA 1998, 95, 5857–5864. [Google Scholar] [CrossRef]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent Updates, New Developments and Status in 2020. Nucleic Acids Res. 2021, 49, D458–D460. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; Von Heijne, G.; Sonnhammer, E.L.L. Predicting Transmembrane Protein Topology with a Hidden Markov Model: Application to Complete Genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Blom, N.; Gammeltoft, S.; Brunak, S. Sequence and Structure-Based Prediction of Eukaryotic Protein Phosphorylation Sites. J. Mol. Biol. 1999, 294, 1351–1362. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Brunak, S. Prediction of Glycosylation across the Human Proteome and the Correlation to Protein Function. Pac. Symp. Biocomput. 2002, 310–322. [Google Scholar] [CrossRef]
- Steentoft, C.; Vakhrushev, S.Y.; Joshi, H.J.; Kong, Y.; Vester-Christensen, M.B.; Schjoldager, K.T.B.G.; Lavrsen, K.; Dabelsteen, S.; Pedersen, N.B.; Marcos-Silva, L.; et al. Precision Mapping of the Human O-GalNAc Glycoproteome through SimpleCell Technology. EMBO J. 2013, 32, 1478–1488. [Google Scholar] [CrossRef]
- Erdos, G.; Pajkos, M.; Dosztányi, Z. IUPred3: Prediction of Protein Disorder Enhanced with Unambiguous Experimental Annotation and Visualization of Evolutionary Conservation. Nucleic Acids Res. 2021, 49, W297–W303. [Google Scholar] [CrossRef]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and Sequence Analysis Tools Services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef]
- Gao, Y.L.; Yoshida, A.; Liu, J.Y.; Shimizu, T.; Shirota, K.; Shiina, Y.; Osatomi, K. Molecular Cloning and Expression Dynamics of UNC-45B upon Heat Shock in the Muscle of Yellowtail. Aquaculture 2021, 541, 736827. [Google Scholar] [CrossRef]
- Huan, P.; Wang, H.; Liu, B. A Label-Free Proteomic Analysis on Competent Larvae and Juveniles of the Pacific Oyster (Crassostrea gigas). PLoS ONE 2015, 10, e0135008. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; ISBN 3-900051-07-0. [Google Scholar]
- Goldberg, H.A.; Warner, K.J. The Staining of Acidic Proteins on Polyacrylamide Gels: Enhanced Sensitivity and Stability of “Stains-All” Staining in Combination with Silver Nitrate. Anal. Biochem. 1997, 251, 227–233. [Google Scholar] [CrossRef]
- Samata, T.; Ikeda, D.; Kajikawa, A.; Sato, H.; Nogawa, C.; Yamada, D.; Yamazaki, R.; Akiyama, T. A Novel Phosphorylated Glycoprotein in the Shell Matrix of the Oyster Crassostrea nippona. FEBS J. 2008, 275. [Google Scholar] [CrossRef]
- Tarentino, A.L.; Gomez, C.M.; Plummer, T.H. Deglycosylation of Asparagine-Linked Glycans by Peptide:N-Glycosidase F. Biochemistry 1985, 24, 4665–4671. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Band No. | Protein Name a | Accession Number a | Theoretical Mass (kDa) | Observed Mass (kDa) | PS/NMP b | SC c (%) |
|---|---|---|---|---|---|---|
| Unbound fraction components | ||||||
| UB1 | Gigasin-6 isoform X2 | XP_011449648.2 | 63 | 198 | 79/15 | 20 |
| Gigasin-6 isoform X1 | XP_011449647.2 | 64 | 198 | 78/15 | 19 | |
| UB2 | Gigasin-6 isoform X2 | XP_011449648.2 | 63 | 66 | 85/15 | 21 |
| Gigasin-6 isoform X1 | XP_011449647.2 | 64 | 66 | 84/15 | 20 | |
| UB3 | Surface protein P12p-like | XP_034321529.1 | 15 | 17 | 80/8 | 37 |
| Surface protein P12p-like | XP_034319527.1 | 15 | 17 | 80/8 | 37 | |
| S1 | Unidentified Stains-all-stainable acidic protein | N/A | N/A | 48 | N/A | N/A |
| S2 | Unidentified Stains-all-stainable acidic protein | N/A | N/A | 43 | N/A | N/A |
| Bound fraction components | ||||||
| B1 | Gigasin-6 isoform X1 | XP_011449647.2 | 64 | 198 | 97/17 | 21 |
| Gigasin-6 isoform X2 | XP_011449648.2 | 63 | 198 | 96/17 | 22 | |
| B2 | Gigasin-6 isoform X1 | XP_011449647.2 | 64 | 66 | 63/13 | 18 |
| Gigasin-6 isoform X2 | XP_011449648.2 | 63 | 66 | 63/13 | 18 | |
| Band No. | Protein Name a | Accession Number a | Theoretical Mass (kDa) | Observed Mass (kDa) | PS/NMP b | SC c (%) |
|---|---|---|---|---|---|---|
| Bound fraction components | ||||||
| Deglycosylated | ||||||
| DG1 | Gigasin-6 isoform X1 | XP_011449647.2 | 64 | 196 | 73/14 | 21 |
| Gigasin-6 isoform X2 | XP_011449648.2 | 63 | 196 | 72/14 | 22 | |
| DG2 | Gigasin-6 isoform X1 | XP_011449647.2 | 64 | 64 | 91/16 | 21 |
| Gigasin-6 isoform X2 | XP_011449648.2 | 63 | 64 | 90/16 | 22 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sedanza, M.G.; Yoshida, A.; Kim, H.-J.; Yamaguchi, K.; Osatomi, K.; Satuito, C.G. Identification and Characterization of the Larval Settlement Pheromone Protein Components in Adult Shells of Crassostrea gigas: A Novel Function of Shell Matrix Proteins. Int. J. Mol. Sci. 2022, 23, 9816. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23179816
Sedanza MG, Yoshida A, Kim H-J, Yamaguchi K, Osatomi K, Satuito CG. Identification and Characterization of the Larval Settlement Pheromone Protein Components in Adult Shells of Crassostrea gigas: A Novel Function of Shell Matrix Proteins. International Journal of Molecular Sciences. 2022; 23(17):9816. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23179816
Chicago/Turabian StyleSedanza, Mary Grace, Asami Yoshida, Hee-Jin Kim, Kenichi Yamaguchi, Kiyoshi Osatomi, and Cyril Glenn Satuito. 2022. "Identification and Characterization of the Larval Settlement Pheromone Protein Components in Adult Shells of Crassostrea gigas: A Novel Function of Shell Matrix Proteins" International Journal of Molecular Sciences 23, no. 17: 9816. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23179816