
Lipid Droplet Formation Is Regulated by Ser/Thr Phosphatase PPM1D via Dephosphorylation of Perilipin 1

 , and
, and
Abstract
:1. Introduction
2. Results
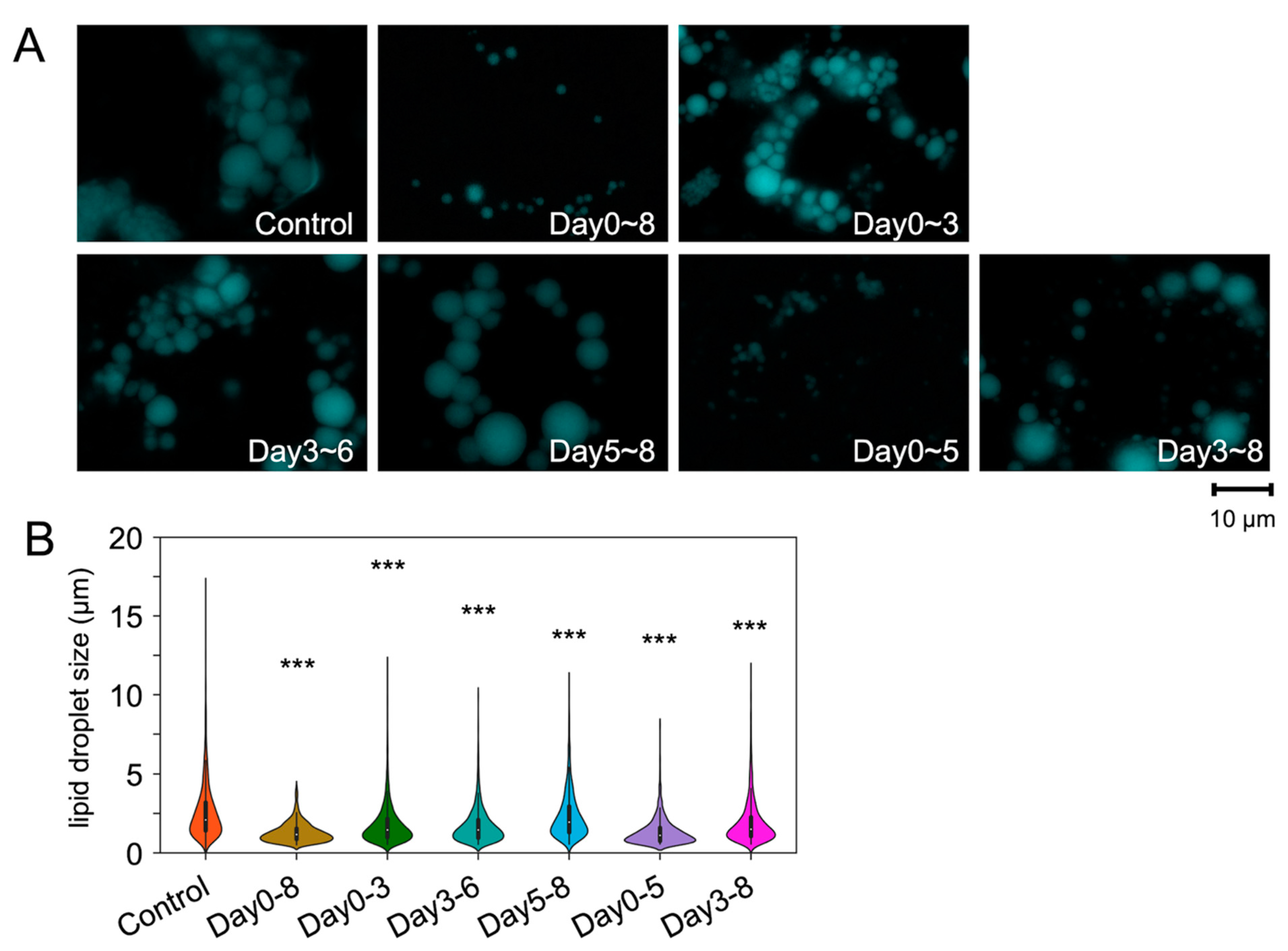
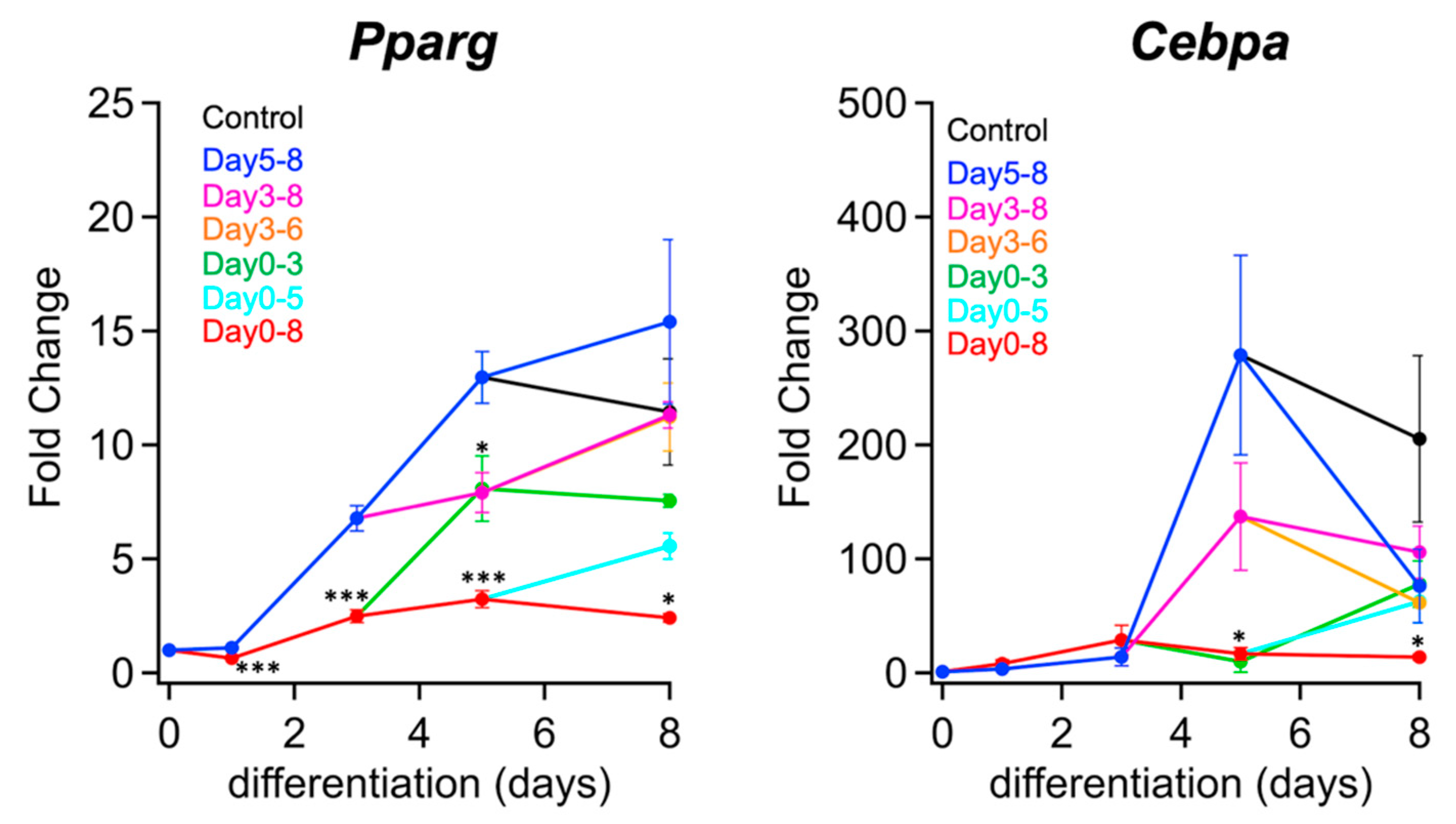
2.1. PPM1D Inhibition in Early Stage Suppressed Adipocyte Differentiation
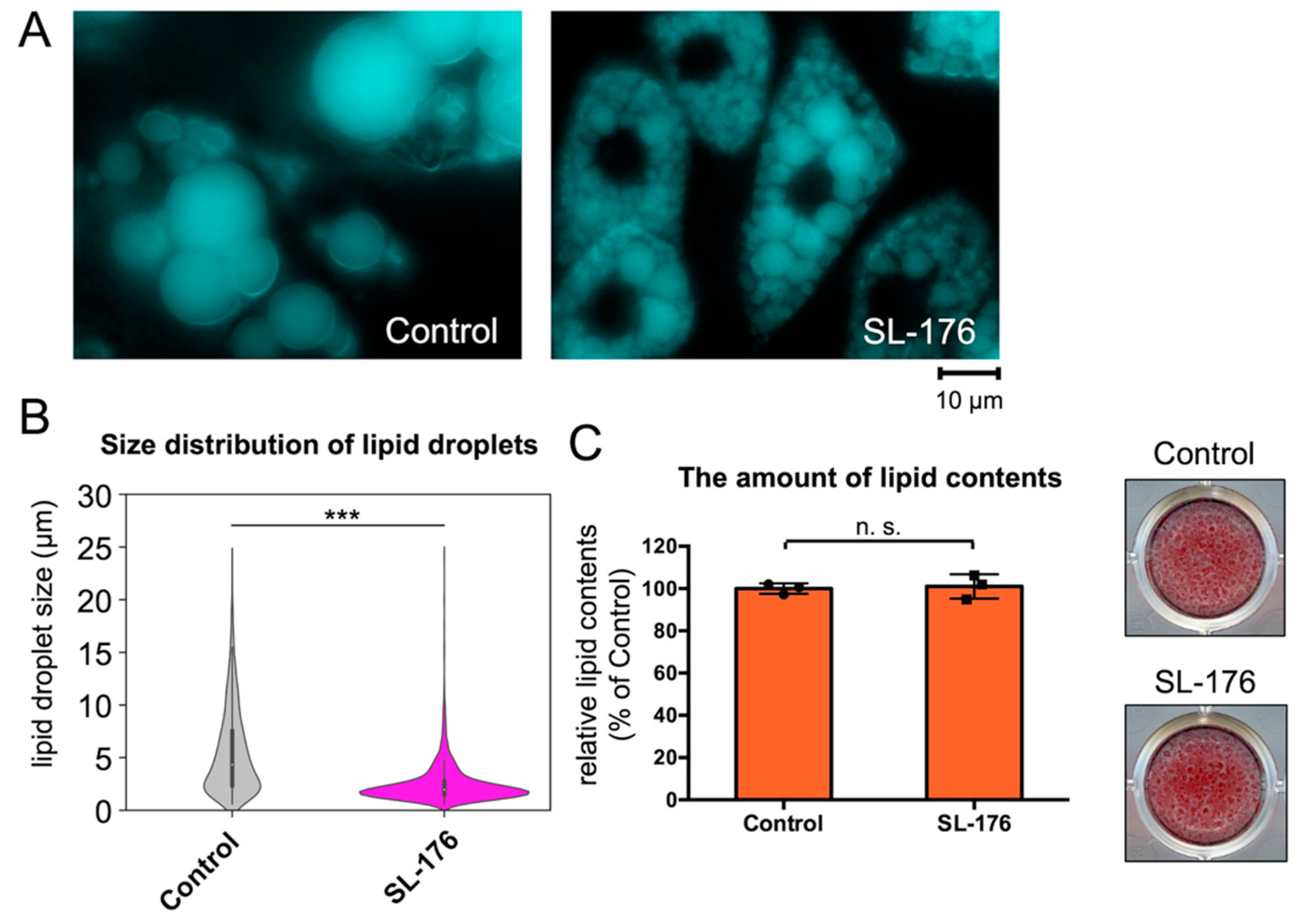
2.2. PPM1D Inhibition in Mature Adipocytes Significantly Reduced the Size of LDs
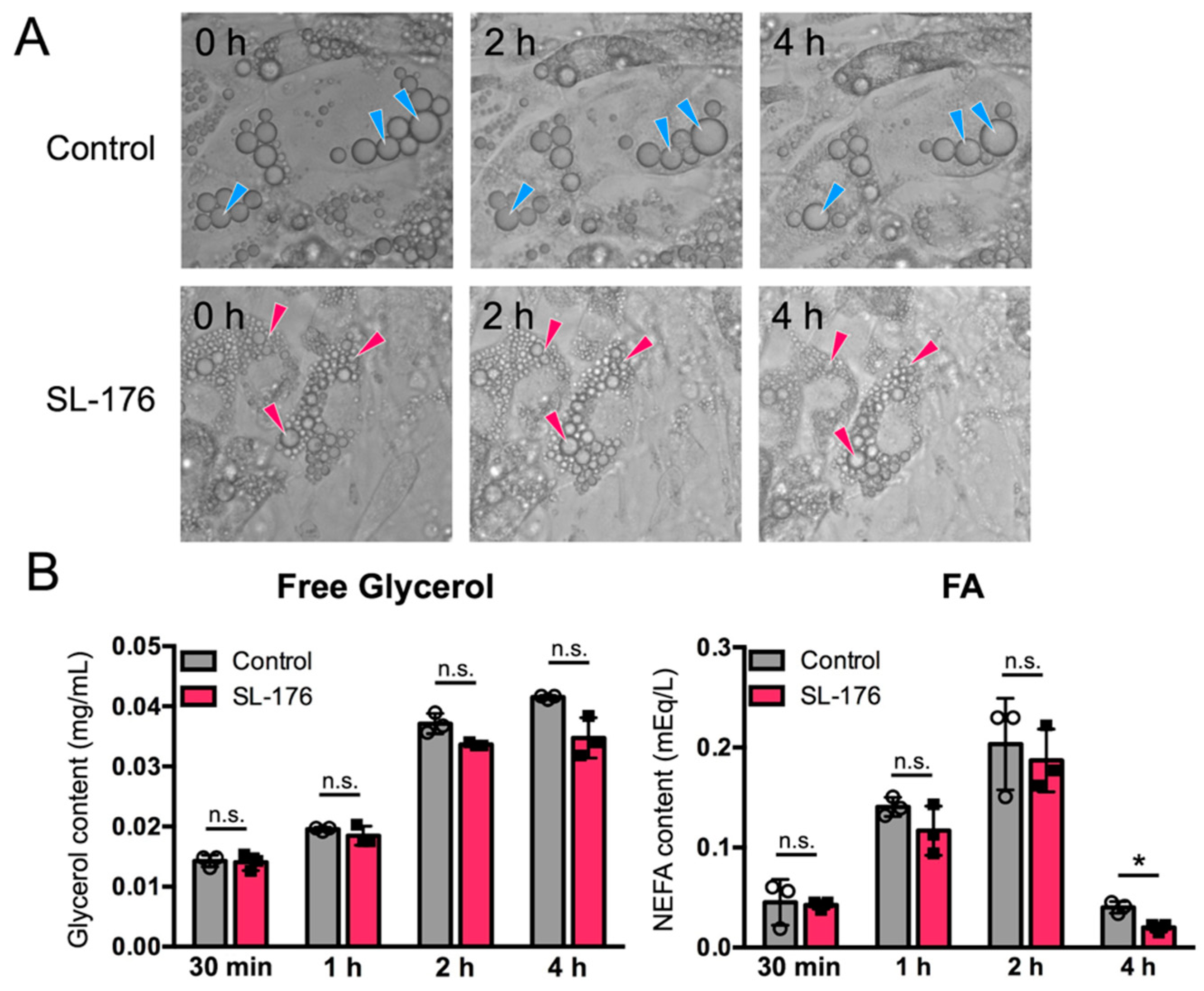
2.3. Small LDs Produced upon PPM1D Inhibition Showed Resistance to Lipolysis
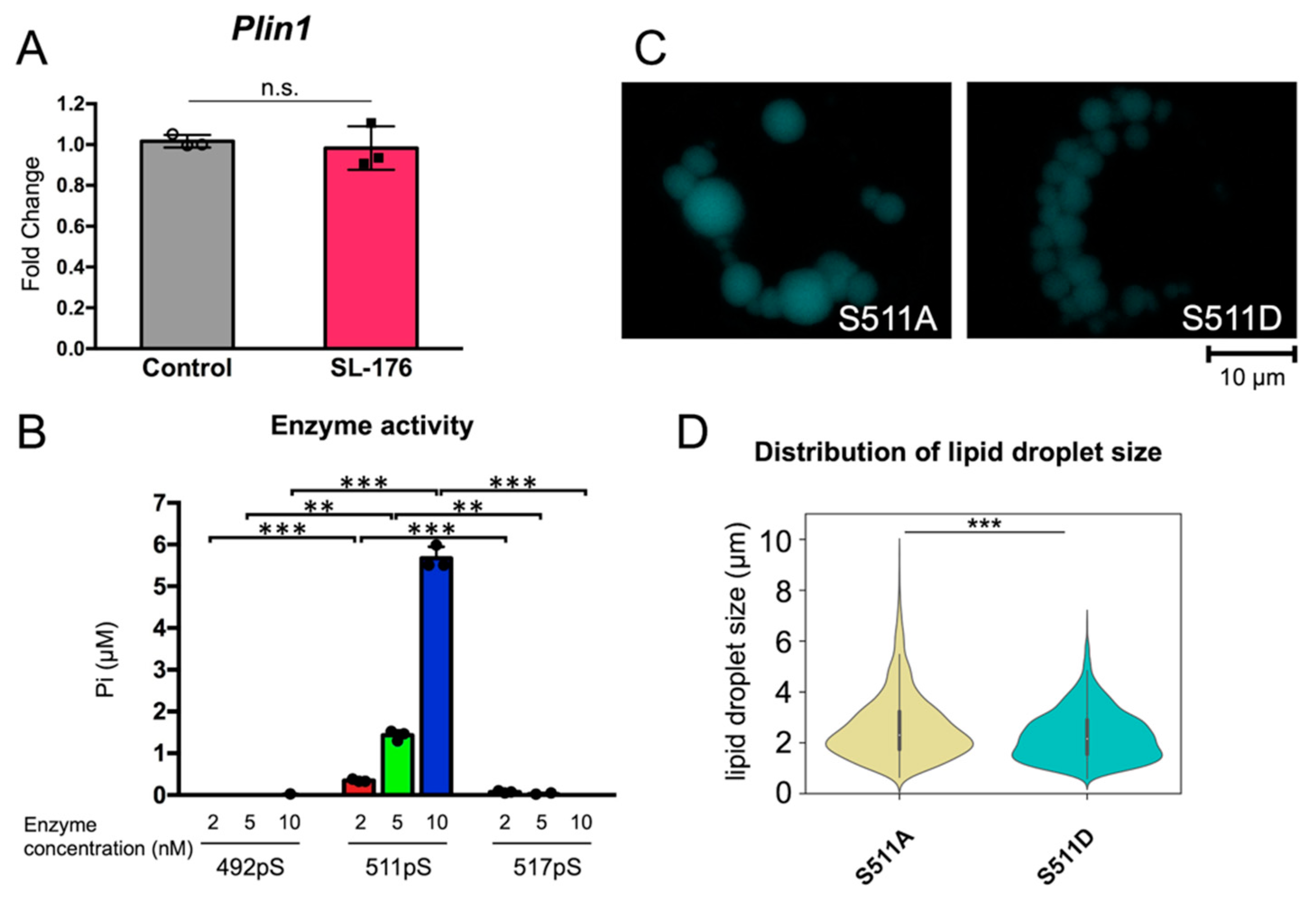
2.4. LD Size was Regulated by Dephosphorylation of Ser511 of Perilipin 1
3. Discussion
4. Materials and Methods
4.1. Cell lines and Materials
4.2. Cell Manipulation
4.3. Oil Red O Staining
4.4. LD imaging and Quantification
4.5. RT-qPCR
4.6. Protein Purification
4.7. In Vitro Phosphatase Assay
4.8. Lipolysis Assay
4.9. Western Blotting
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980-2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef] [Green Version]
- Jeffery, E.; Wing, A.; Holtrup, B.; Sebo, Z.; Kaplan, J.L.; Saavedra-Pena, R.; Church, C.D.; Colman, L.; Berry, R.; Rodeheffer, M.S. The Adipose Tissue Microenvironment Regulates Depot-Specific Adipogenesis in Obesity. Cell Metab. 2016, 24, 142–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, A.L.S.; Barreto, E.A.; Fazolini, N.P.B.; Viola, J.P.B.; Bozza, P.T. Lipid droplets: Platforms with multiple functions in cancer hallmarks. Cell Death Dis. 2020, 11, 105. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, A.S.; Coleman, R.A.; Kraemer, F.B.; McManaman, J.L.; Obin, M.S.; Puri, V.; Yan, Q.W.; Miyoshi, H.; Mashek, D.G. The role of lipid droplets in metabolic disease in rodents and humans. J. Clin. Investig. 2011, 121, 2102–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schott, M.B.; Weller, S.G.; Schulze, R.J.; Krueger, E.W.; Drizyte-Miller, K.; Casey, C.A.; McNiven, M.A. Lipid droplet size directs lipolysis and lipophagy catabolism in hepatocytes. J. Cell Biol. 2019, 218, 3320–3335. [Google Scholar] [CrossRef]
- Paar, M.; Jungst, C.; Steiner, N.A.; Magnes, C.; Sinner, F.; Kolb, D.; Lass, A.; Zimmermann, R.; Zumbusch, A.; Kohlwein, S.D.; et al. Remodeling of lipid droplets during lipolysis and growth in adipocytes. J. Biol. Chem. 2012, 287, 11164–11173. [Google Scholar] [CrossRef] [Green Version]
- Darlington, G.J.; Ross, S.E.; MacDougald, O.A. The role of C/EBP genes in adipocyte differentiation. J. Biol. Chem. 1998, 273, 30057–30060. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Bucher, N.L.; Farmer, S.R. Induction of peroxisome proliferator-activated receptor gamma during the conversion of 3T3 fibroblasts into adipocytes is mediated by C/EBPbeta, C/EBPdelta, and glucocorticoids. Mol. Cell. Biol. 1996, 16, 4128–4136. [Google Scholar] [CrossRef] [Green Version]
- Yeh, W.C.; Cao, Z.; Classon, M.; McKnight, S.L. Cascade regulation of terminal adipocyte differentiation by three members of the C/EBP family of leucine zipper proteins. Genes Dev. 1995, 9, 168–181. [Google Scholar] [CrossRef] [Green Version]
- Rosen, E.D.; Hsu, C.H.; Wang, X.; Sakai, S.; Freeman, M.W.; Gonzalez, F.J.; Spiegelman, B.M. C/EBPalpha induces adipogenesis through PPARgamma: A unified pathway. Genes Dev. 2002, 16, 22–26. [Google Scholar] [CrossRef]
- Arimura, N.; Horiba, T.; Imagawa, M.; Shimizu, M.; Sato, R. The peroxisome proliferator-activated receptor gamma regulates expression of the perilipin gene in adipocytes. J. Biol. Chem. 2004, 279, 10070–10076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenberg, A.S.; Egan, J.J.; Wek, S.A.; Garty, N.B.; Blanchette-Mackie, E.J.; Londos, C. Perilipin, a major hormonally regulated adipocyte-specific phosphoprotein associated with the periphery of lipid storage droplets. J. Biol. Chem. 1991, 266, 11341–11346. [Google Scholar] [CrossRef]
- Martinez-Botas, J.; Anderson, J.B.; Tessier, D.; Lapillonne, A.; Chang, B.H.; Quast, M.J.; Gorenstein, D.; Chen, K.H.; Chan, L. Absence of perilipin results in leanness and reverses obesity in Lepr(db/db) mice. Nat. Genet. 2000, 26, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Gong, J.; Wu, H.; Xu, W.; Wu, L.; Xu, D.; Gao, J.; Wu, J.W.; Yang, H.; Yang, M.; et al. Perilipin1 promotes unilocular lipid droplet formation through the activation of Fsp27 in adipocytes. Nat. Commun. 2013, 4, 1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granneman, J.G.; Moore, H.P.; Krishnamoorthy, R.; Rathod, M. Perilipin controls lipolysis by regulating the interactions of AB-hydrolase containing 5 (Abhd5) and adipose triglyceride lipase (Atgl). J. Biol. Chem. 2009, 284, 34538–34544. [Google Scholar] [CrossRef] [Green Version]
- Girousse, A.; Langin, D. Adipocyte lipases and lipid droplet-associated proteins: Insight from transgenic mouse models. Int. J. Obes. 2012, 36, 581–594. [Google Scholar] [CrossRef] [Green Version]
- Clifford, G.M.; McCormick, D.K.; Londos, C.; Vernon, R.G.; Yeaman, S.J. Dephosphorylation of perilipin by protein phosphatases present in rat adipocytes. FEBS Lett. 1998, 435, 125–129. [Google Scholar] [CrossRef] [Green Version]
- Fiscella, M.; Zhang, H.; Fan, S.; Sakaguchi, K.; Shen, S.; Mercer, W.E.; Vande Woude, G.F.; O’Connor, P.M.; Appella, E. Wip1, a novel human protein phosphatase that is induced in response to ionizing radiation in a p53-dependent manner. Proc. Natl. Acad. Sci. USA 1997, 94, 6048–6053. [Google Scholar] [CrossRef] [Green Version]
- Le Guezennec, X.; Brichkina, A.; Huang, Y.F.; Kostromina, E.; Han, W.; Bulavin, D.V. Wip1-dependent regulation of autophagy, obesity, and atherosclerosis. Cell Metab. 2012, 16, 68–80. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Zhang, L.; Xu, L.; Liu, L.; He, Y.; Zhang, Y.; Huang, X.; Zhao, T.; Wu, L.; Zhao, Y.; et al. WIP1 phosphatase is a critical regulator of adipogenesis through dephosphorylating PPARgamma serine 112. Cell. Mol. Life Sci. 2017, 74, 2067–2079. [Google Scholar] [CrossRef]
- Ogasawara, S.; Kiyota, Y.; Chuman, Y.; Kowata, A.; Yoshimura, F.; Tanino, K.; Kamada, R.; Sakaguchi, K. Novel inhibitors targeting PPM1D phosphatase potently suppress cancer cell proliferation. Bioorg. Med. Chem. 2015, 23, 6246–6249. [Google Scholar] [CrossRef] [PubMed]
- Kamada, R.; Kimura, N.; Yoshimura, F.; Tanino, K.; Sakaguchi, K. Inhibition of lipid droplet formation by Ser/Thr protein phosphatase PPM1D inhibitor, SL-176. PLoS ONE 2019, 14, e0212682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphrey, S.J.; Yang, G.; Yang, P.Y.; Fazakerley, D.J.; Stockli, J.; Yang, J.Y.; James, D.E. Dynamic Adipocyte Phosphoproteome Reveals that Akt Directly Regulates mTORC2. Cell Metab. 2013, 17, 1009–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyoshi, H.; Souza, S.C.; Zhang, H.H.; Strissel, K.J.; Christoffolete, M.A.; Kovsan, J.; Rudich, A.; Kraemer, F.B.; Bianco, A.C.; Obin, M.S.; et al. Perilipin promotes hormone-sensitive lipase-mediated adipocyte lipolysis via phosphorylation-dependent and -independent mechanisms. J. Biol. Chem. 2006, 281, 15837–15844. [Google Scholar] [CrossRef] [Green Version]
- Marcinkiewicz, A.; Gauthier, D.; Garcia, A.; Brasaemle, D.L. The phosphorylation of serine 492 of perilipin a directs lipid droplet fragmentation and dispersion. J. Biol. Chem. 2006, 281, 11901–11909. [Google Scholar] [CrossRef] [Green Version]
- Souza, S.C.; Muliro, K.V.; Liscum, L.; Lien, P.; Yamamoto, M.T.; Schaffer, J.E.; Dallal, G.E.; Wang, X.; Kraemer, F.B.; Obin, M.; et al. Modulation of hormone-sensitive lipase and protein kinase A-mediated lipolysis by perilipin A in an adenoviral reconstituted system. J. Biol. Chem. 2002, 277, 8267–8272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, V.; Rothenberg, A.; Gomez, C.; Cohen, A.W.; Garcia, A.; Bhattacharyya, S.; Shapiro, L.; Dolios, G.; Wang, R.; Lisanti, M.P.; et al. Perilipin A mediates the reversible binding of CGI-58 to lipid droplets in 3T3-L1 adipocytes. J. Biol. Chem. 2004, 279, 42062–42071. [Google Scholar] [CrossRef] [Green Version]
- Puri, V.; Konda, S.; Ranjit, S.; Aouadi, M.; Chawla, A.; Chouinard, M.; Chakladar, A.; Czech, M.P. Fat-specific protein 27, a novel lipid droplet protein that enhances triglyceride storage. J. Biol. Chem. 2007, 282, 34213–34218. [Google Scholar] [CrossRef] [Green Version]
- Namba, K.; Osawa, A.; Nakayama, A.; Mera, A.; Tano, F.; Chuman, Y.; Sakuda, E.; Taketsugu, T.; Sakaguchi, K.; Kitamura, N.; et al. Synthesis of yellow and red fluorescent 1,3a, 6a-triazapentalenes and the theoretical investigation of their optical properties. Chem. Sci. 2015, 6, 1083–1093. [Google Scholar] [CrossRef] [Green Version]
- Kozakai, Y.; Kamada, R.; Furuta, J.; Kiyota, Y.; Chuman, Y.; Sakaguchi, K. PPM1D controls nucleolar formation by up-regulating phosphorylation of nucleophosmin. Sci. Rep. 2016, 6, 33272. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Average Size 1 | Size Distribution of LDs (%) 2 | |||||
|---|---|---|---|---|---|---|
| [µm] | 0–2 µm | 2–4 µm | 4–6 µm | 6–8 µm | >8 µm | |
| Control | 2.60 ± 0.03 | 46.6 | 37.7 | 10.7 | 3.1 | 1.9 |
| Days 0–8 | 1.30 ± 0.02 | 88.2 | 11.4 | 0.3 | 0.0 | 0.0 |
| Days 0–3 | 1.76 ± 0.02 | 71.9 | 23.2 | 3.7 | 0.3 | 0.3 |
| Days 3–6 | 1.73 ± 0.02 | 72.7 | 22.9 | 3.4 | 0.2 | 0.2 |
| Days 5–8 | 2.33 ± 0.03 | 52.8 | 34.7 | 9.5 | 0.6 | 0.6 |
| Days 0–5 | 1.33 ± 0.02 | 85.0 | 13.4 | 1.5 | 0.1 | 0.1 |
| Days 3–8 | 1.91 ± 0.02 | 68.3 | 24.4 | 5.4 | 0.5 | 0.5 |
| Average size 1 | Size Distribution of LDs (%) 2 | |||||
|---|---|---|---|---|---|---|
| [µm] | 0–2 µm | 2–4 µm | 4–6 µm | 6–8 µm | >8 µm | |
| Control | 5.47 ± 0.10 | 19.4 | 27.2 | 17.8 | 13.1 | 22.5 |
| SL-176 | 2.50 ± 0.05 | 53.7 | 33.1 | 8.4 | 2.7 | 2.1 |
| Average Size 1 | Size Distribution of LDs (%) 2 | ||||
|---|---|---|---|---|---|
| [µm] | 0–2 µm | 2–4 µm | 4–6 µm | >6 µm | |
| S511A | 2.61 ± 0.03 | 38.1 | 49.0 | 10.2 | 2.6 |
| S511D | 2.27 ± 0.02 | 44.5 | 49.7 | 5.7 | 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamada, R.; Uno, S.; Kimura, N.; Yoshimura, F.; Tanino, K.; Sakaguchi, K. Lipid Droplet Formation Is Regulated by Ser/Thr Phosphatase PPM1D via Dephosphorylation of Perilipin 1. Int. J. Mol. Sci. 2022, 23, 12046. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231912046
Kamada R, Uno S, Kimura N, Yoshimura F, Tanino K, Sakaguchi K. Lipid Droplet Formation Is Regulated by Ser/Thr Phosphatase PPM1D via Dephosphorylation of Perilipin 1. International Journal of Molecular Sciences. 2022; 23(19):12046. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231912046
Chicago/Turabian StyleKamada, Rui, Sae Uno, Nozomi Kimura, Fumihiko Yoshimura, Keiji Tanino, and Kazuyasu Sakaguchi. 2022. "Lipid Droplet Formation Is Regulated by Ser/Thr Phosphatase PPM1D via Dephosphorylation of Perilipin 1" International Journal of Molecular Sciences 23, no. 19: 12046. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231912046