
Per1/Per2 knockout Affects Spleen Immune Function in Elderly Mice via Inducing Spleen Lymphocyte Ferroptosis
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
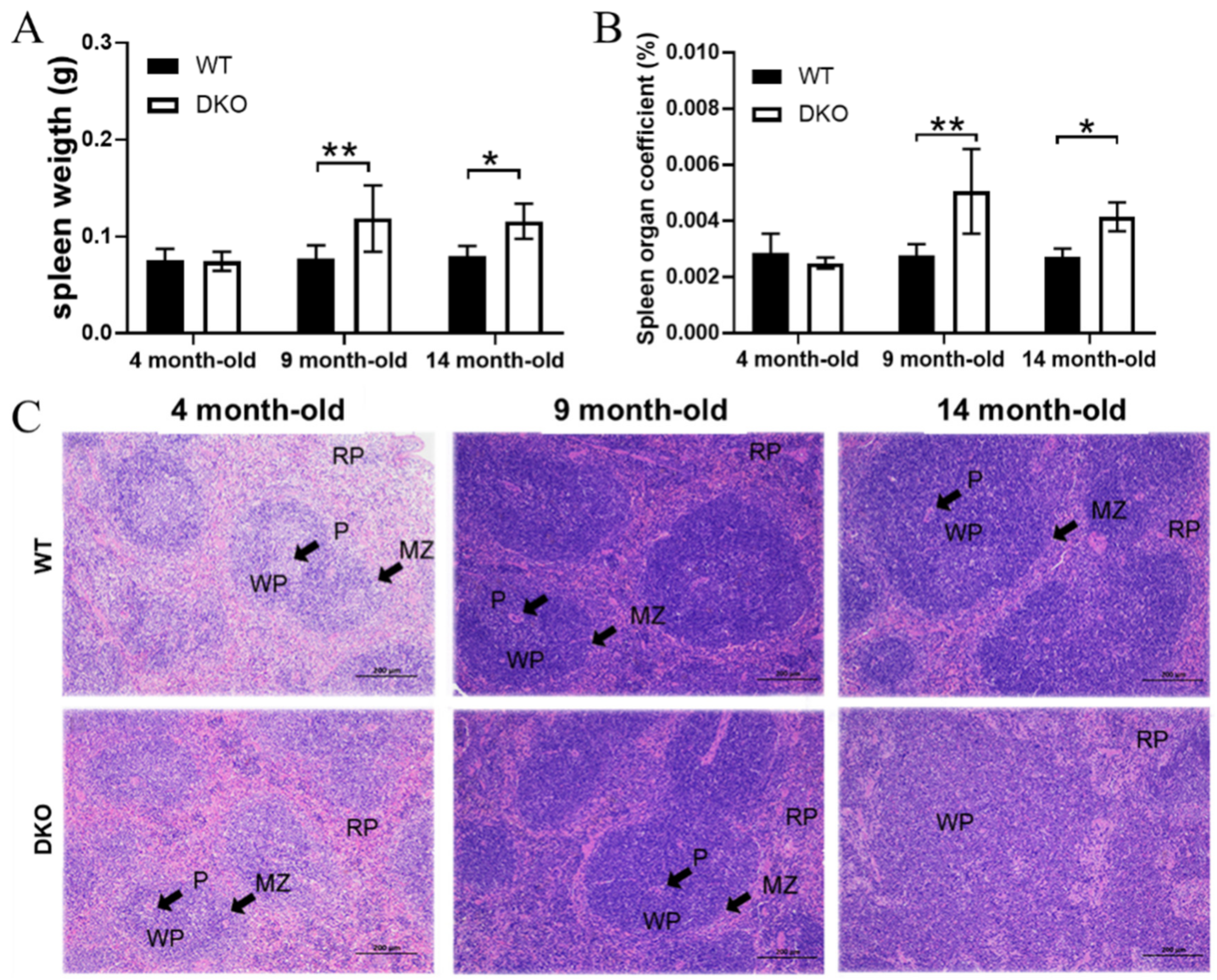
2.1. Effects of Per1/Per2 Knockout on Spleen Weight and Pathological Structure
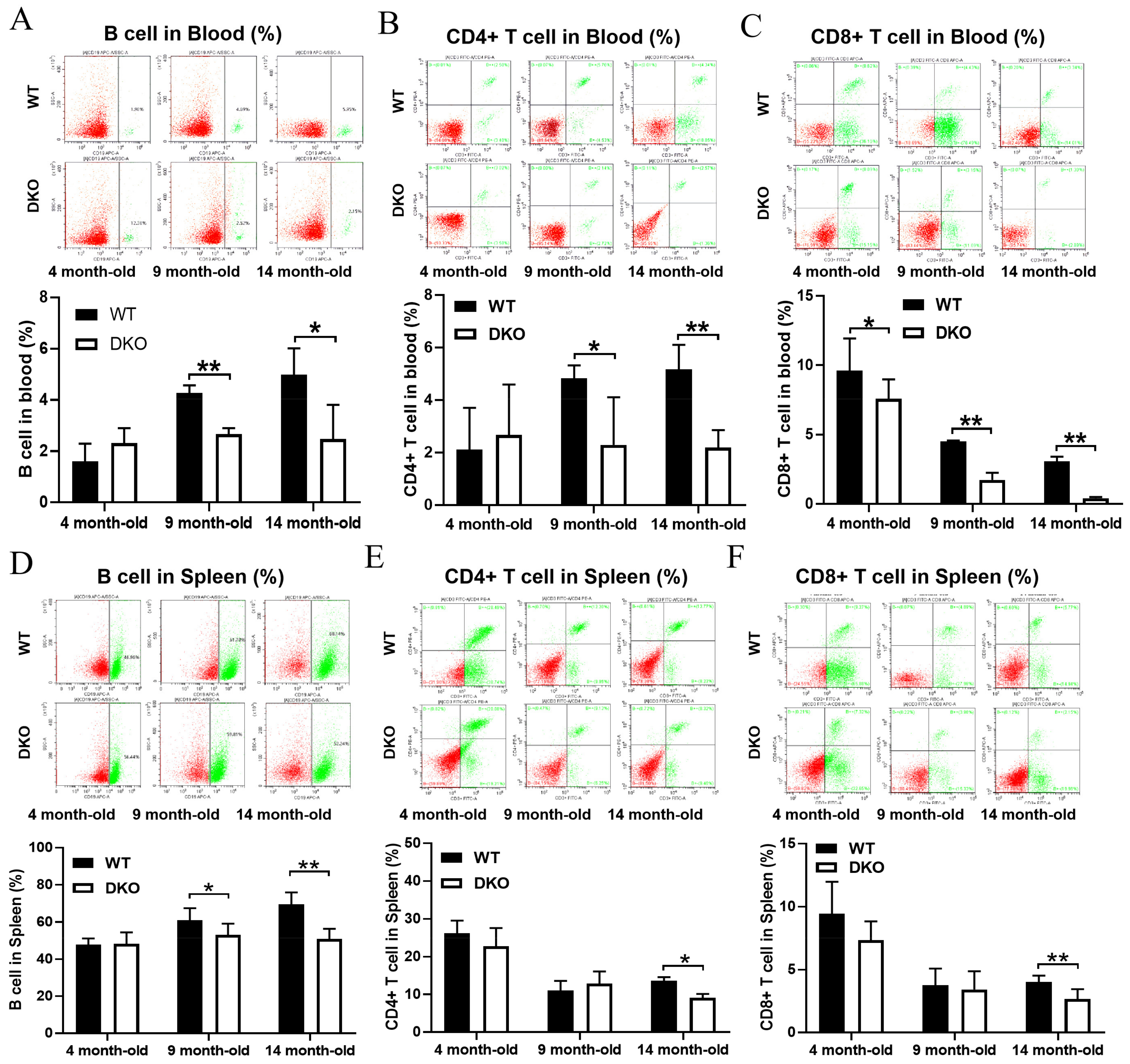
2.2. Levels of B Cells, CD3+-CD4+T Cells, and CD3+-CD8+T Cells in Blood and Spleen of DKO and WT Mice at Different Ages
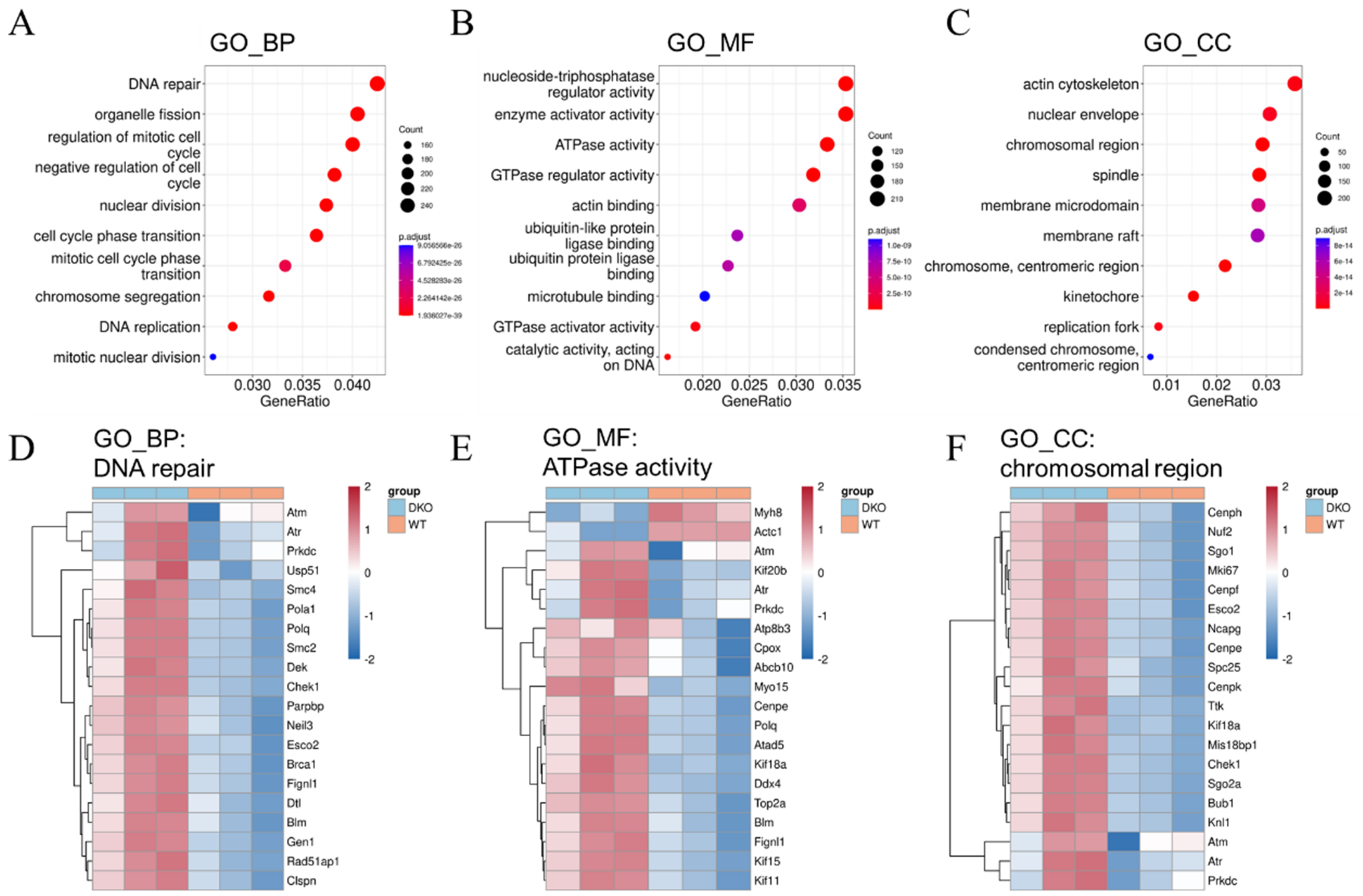
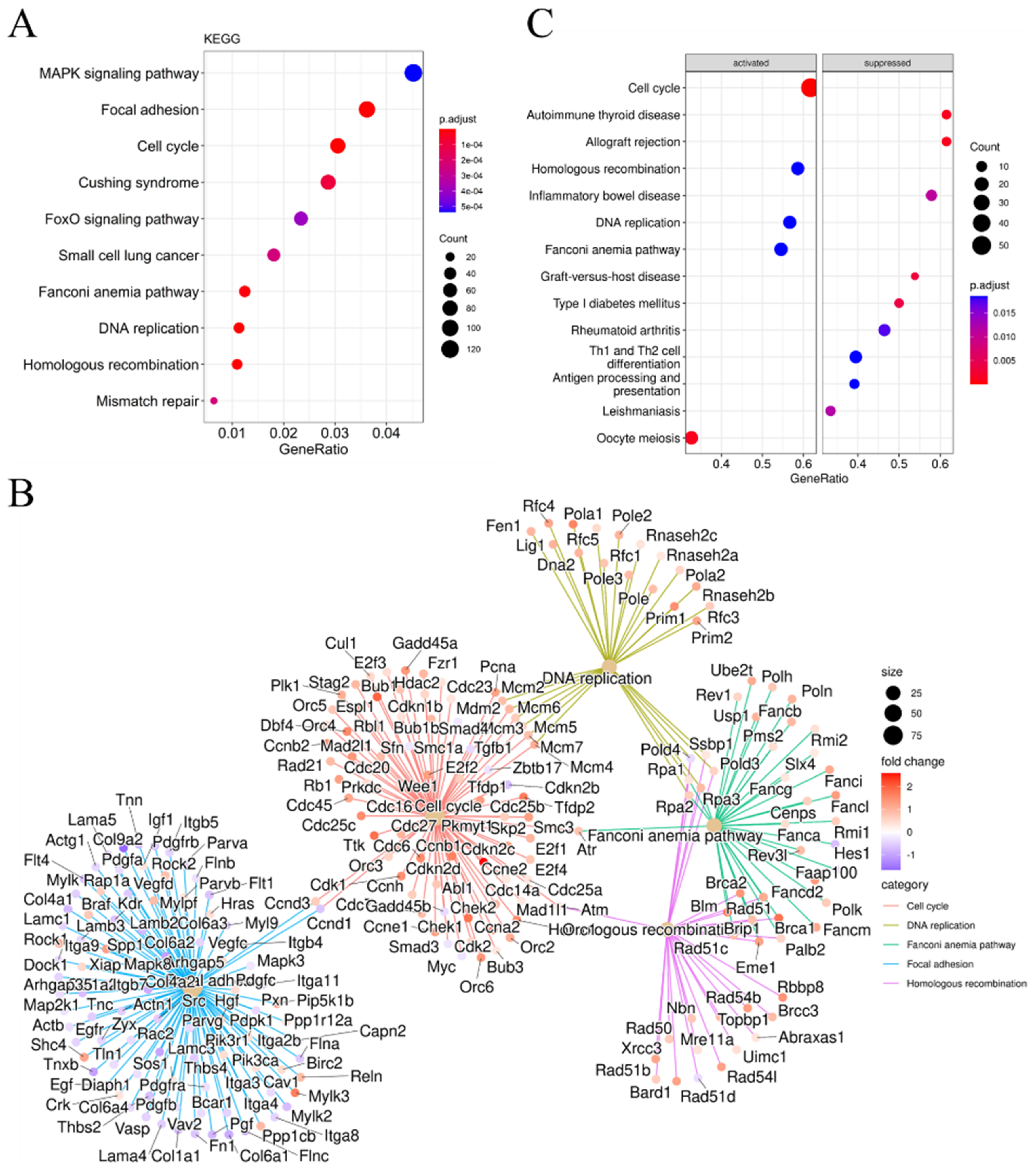
2.3. DNA Damage Response and Oxidative Stress Resistance in DKO Mice Spleen Tissue Detected through Transcriptome Sequencing
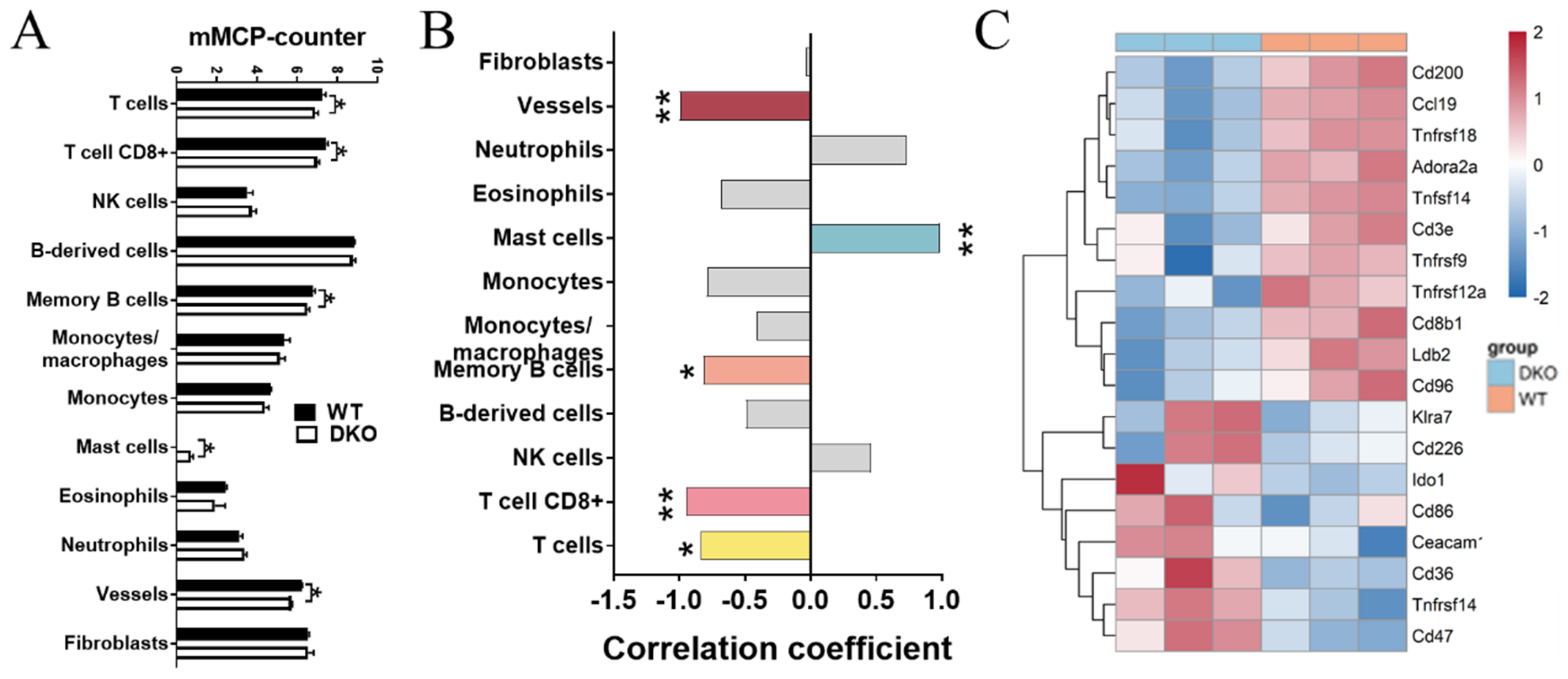
2.4. Differences in the Immune Microenvironment and Immune-Related Signatures between the DKO and WT Mice Spleen Based on Transcriptome Analysis
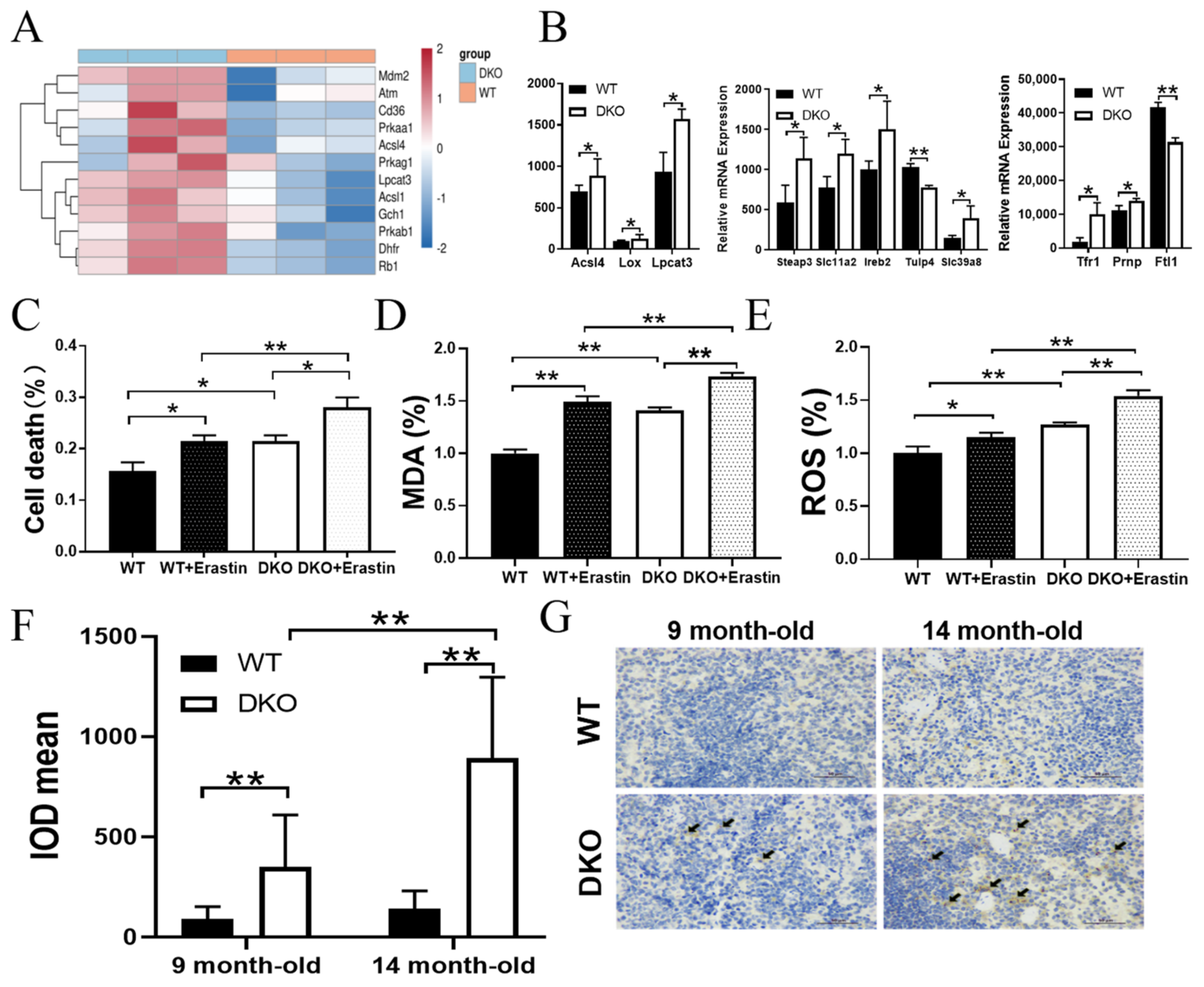
2.5. Per1/Per2 Knockout Activated Ferroptosis in the Spleen of 14-Month-Old Mice
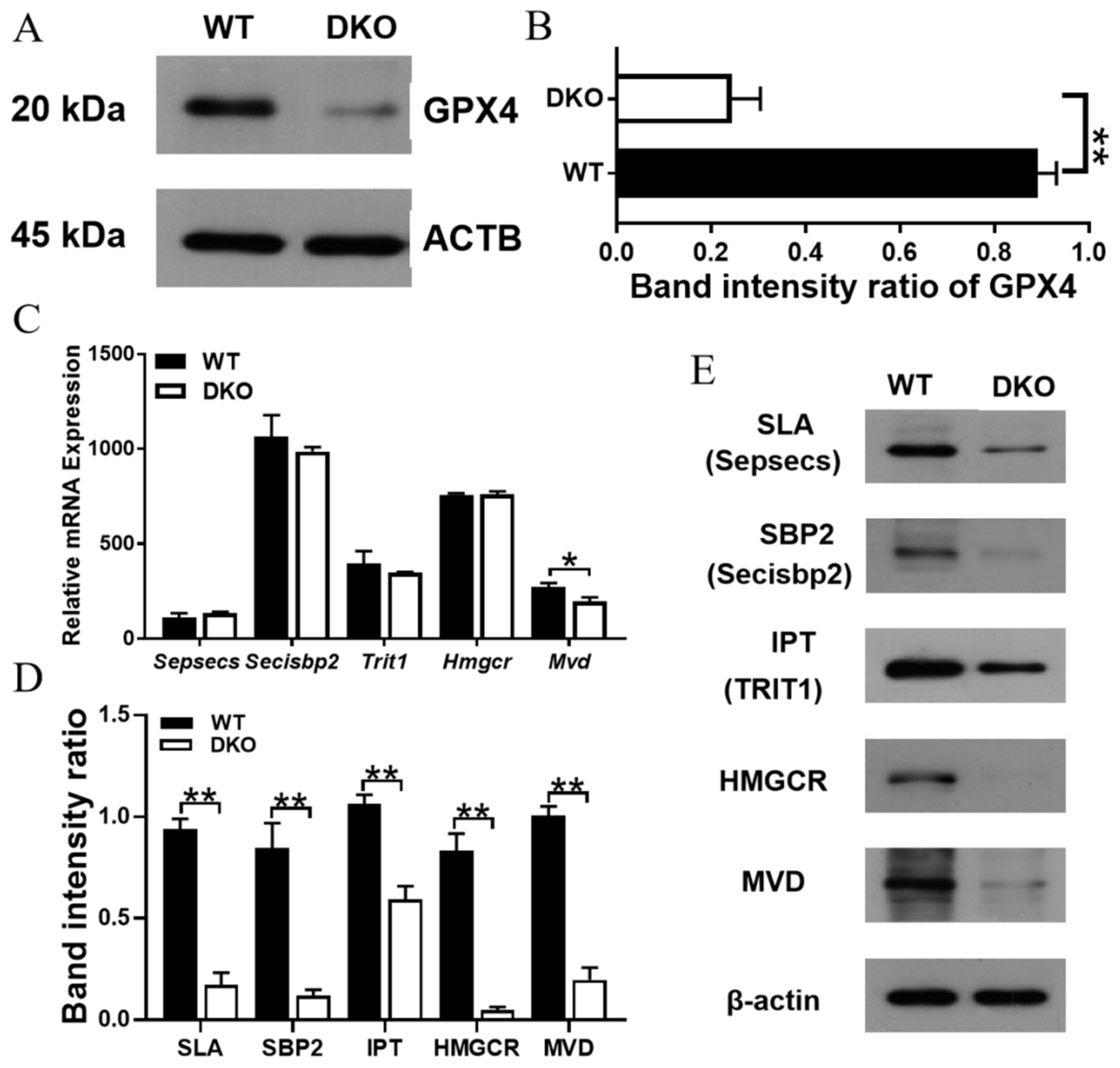
2.6. Per1/Per2 DKO Promotes Ferroptosis in the Spleen of 14-Month-Old Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Histological Study and Immunohistochemical Staining
4.2.1. Histology
4.2.2. Immunohistochemistry
4.3. Flow Cytometry
4.4. BGISEQ-500 RNA-Seq
4.5. Cell Culture
4.6. MDA and ROS Assays
4.6.1. MDA
4.6.2. ROS
4.7. Western Blot Analysis
4.8. qPCR Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bass, J.; Takahashi, J.S. Circadian integration of metabolism and energetics. Science 2010, 330, 1349–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, Y.; Kang, H.S.; Angers, M.; Jetten, A.M. Retinoic acid-related orphan receptor gamma directly regulates neuronal PAS domain protein 2 transcription in vivo. Nucleic Acids Res. 2011, 39, 4769–4782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.; Gammie, S.C. The circadian gene Nr1d1 in the mouse nucleus accumbens modulates sociability and anxiety-related behaviour. Eur. J. Neurosci. 2018, 48, 1924–1943. [Google Scholar] [CrossRef] [PubMed]
- Chellappa, S.L.; Vujovic, N.; Williams, J.S.; Scheer, F. Impact of Circadian Disruption on Cardiovascular Function and Disease. Trends Endocrinol. Metab. 2019, 30, 767–779. [Google Scholar] [CrossRef] [PubMed]
- Konturek, P.C.; Brzozowski, T.; Konturek, S.J. Gut clock: Implication of circadian rhythms in the gastrointestinal tract. J. Physiol. Pharmacol. 2011, 62, 139–150. [Google Scholar]
- Yang, Y.; Zhang, J. Bile acid metabolism and circadian rhythms. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 319, G549–G563. [Google Scholar] [CrossRef]
- Mills, J.; Kuohung, W. Impact of circadian rhythms on female reproduction and infertility treatment success. Curr. Opin. Endocrinol. Diabetes Obes. 2019, 26, 317–321. [Google Scholar] [CrossRef]
- Logan, R.W.; McClung, C.A. Rhythms of life: Circadian disruption and brain disorders across the lifespan. Nat. Rev. Neurosci. 2019, 20, 49–65. [Google Scholar] [CrossRef]
- Cermakian, N.; Stegeman, S.K.; Tekade, K.; Labrecque, N. Circadian rhythms in adaptive immunity and vaccination. Semin. Immunopathol. 2022, 44, 193–207. [Google Scholar] [CrossRef]
- Gray, K.J.; Gibbs, J.E. Adaptive immunity, chronic inflammation and the clock. Semin. Immunopathol. 2022, 44, 209–224. [Google Scholar] [CrossRef]
- Xie, L.; Kang, H.; Xu, Q.; Chen, M.J.; Liao, Y.; Thiyagarajan, M.; O’Donnell, J.; Christensen, D.J.; Nicholson, C.; Iliff, J.J.; et al. Sleep drives metabolite clearance from the adult brain. Science 2013, 342, 373–377. [Google Scholar] [CrossRef]
- Aiello, I.; Fedele, M.L.M.; Roman, F.; Marpegan, L.; Caldart, C.; Chiesa, J.J.; Golombek, D.A.; Finkielstein, C.V.; Paladino, N. Circadian disruption promotes tumor-immune microenvironment remodeling favoring tumor cell proliferation. Sci. Adv. 2020, 6, eaaz4530. [Google Scholar] [CrossRef]
- Lewis, S.M.; Williams, A.; Eisenbarth, S.C. Structure and function of the immune system in the spleen. Sci. Immunol. 2019, 4, eaau6085. [Google Scholar] [CrossRef]
- Lei, G.; Mao, C.; Yan, Y.; Zhuang, L.; Gan, B. Ferroptosis, radiotherapy, and combination therapeutic strategies. Protein Cell 2021, 12, 836–857. [Google Scholar] [CrossRef]
- Becht, E.; Giraldo, N.A.; Lacroix, L.; Buttard, B.; Elarouci, N.; Petitprez, F.; Selves, J.; Laurent-Puig, P.; Sautes-Fridman, C.; Fridman, W.H.; et al. Erratum to: Estimating the population abundance of tissue-infiltrating immune and stromal cell populations using gene expression. Genome Biol. 2016, 17, 249. [Google Scholar] [CrossRef] [Green Version]
- Rutsche, D.; Michalak-Micka, K.; Zielinska, D.; Moll, H.; Moehrlen, U.; Biedermann, T.; Klar, A.S. The Role of CD200-CD200 Receptor in Human Blood and Lymphatic Endothelial Cells in the Regulation of Skin Tissue Inflammation. Cells 2022, 11, 1055. [Google Scholar] [CrossRef]
- Ma, X.; Xiao, L.; Liu, L.; Ye, L.; Su, P.; Bi, E.; Wang, Q.; Yang, M.; Qian, J.; Yi, Q. CD36-mediated ferroptosis dampens intratumoral CD8(+) T cell effector function and impairs their antitumor ability. Cell Metab. 2021, 33, 1001–1012.e1005. [Google Scholar] [CrossRef]
- Zhang, H.L.; Hu, B.X.; Li, Z.L.; Du, T.; Shan, J.L.; Ye, Z.P.; Peng, X.D.; Li, X.; Huang, Y.; Zhu, X.Y.; et al. PKCbetaII phosphorylates ACSL4 to amplify lipid peroxidation to induce ferroptosis. Nat. Cell Biol. 2022, 24, 88–98. [Google Scholar] [CrossRef]
- Zhou, B.R.; Liu, J.; Kang, R.; Klionsky, D.J.; Kroemer, G.; Tang, D.L. Ferroptosis is a type of autophagy-dependent cell death. Semin. Cancer Biol. 2020, 66, 89–100. [Google Scholar] [CrossRef]
- Ganz, F.D. Sleep and immune function. Crit. Care Nurse 2012, 32, e19–e25. [Google Scholar] [CrossRef] [Green Version]
- Pierzyna, M.; Twardoch, M.; Mazur, B. The aging of the immune system. Wiad. Lek. 2015, 68, 388–392. [Google Scholar] [PubMed]
- Vitlic, A.; Lord, J.M.; Phillips, A.C. Stress, ageing and their influence on functional, cellular and molecular aspects of the immune system. Age 2014, 36, 1169–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manoogian, E.N.C.; Panda, S. Circadian rhythms, time-restricted feeding, and healthy aging. Ageing Res. Rev. 2017, 39, 59–67. [Google Scholar] [CrossRef] [PubMed]
- LeBien, T.W.; Tedder, T.F. B lymphocytes: How they develop and function. Blood 2008, 112, 1570–1580. [Google Scholar] [CrossRef] [PubMed]
- Hallgren, J.; Gurish, M.F. Mast Cell Progenitor Trafficking and Maturation. Mast Cell Biol. Contemp. Emerg. Top. 2011, 716, 14–28. [Google Scholar] [CrossRef]
- Christ, P.; Sowa, A.S.; Froy, O.; Lorentz, A. The Circadian Clock Drives Mast Cell Functions in Allergic Reactions. Front. Immunol. 2018, 9, 1526. [Google Scholar] [CrossRef] [Green Version]
- Jackson, C.W.; Pratt, C.M.; Rupprecht, C.P.; Pattanaik, D.; Krishnaswamy, G. Mastocytosis and Mast Cell Activation Disorders: Clearing the Air. Int. J. Mol. Sci. 2021, 22, 11270. [Google Scholar] [CrossRef]
- Akin, C. Mast cell activation syndromes. J. Allergy Clin. Immunol. 2017, 140, 349–355. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.X.; Zhou, P.K. DNA damage response signaling pathways and targets for radiotherapy sensitization in cancer. Signal Transduct. Target. Ther. 2020, 5, 60. [Google Scholar] [CrossRef]
- Bae, Y.S.; Oh, H.; Rhee, S.G.; Yoo, Y.D. Regulation of reactive oxygen species generation in cell signaling. Mol. Cells 2011, 32, 491–509. [Google Scholar] [CrossRef] [Green Version]
- Russo, M.W.; Hoofnagle, J.H.; Gu, J.; Fontana, R.J.; Barnhart, H.; Kleiner, D.E.; Chalasani, N.; Bonkovsky, H.L. Spectrum of statin hepatotoxicity: Experience of the drug-induced liver injury network. Hepatology 2014, 60, 679–686. [Google Scholar] [CrossRef]
- Swierczynski, J.; Goyke, E.; Korczynska, J.; Jankowski, Z. Acetyl-CoA carboxylase and fatty acid synthase activities in human hypothalamus. Neurosci. Lett. 2008, 444, 209–211. [Google Scholar] [CrossRef]
- Chen, X.; Kang, R.; Kroemer, G.; Tang, D. Organelle-specific regulation of ferroptosis. Cell Death Differ. 2021, 28, 2843–2856. [Google Scholar] [CrossRef]
- Hu, Z.; Zhang, H.; Yi, B.; Yang, S.; Liu, J.; Hu, J.; Wang, J.; Cao, K.; Zhang, W. VDR activation attenuate cisplatin induced AKI by inhibiting ferroptosis. Cell Death Dis. 2020, 11, 73. [Google Scholar] [CrossRef] [Green Version]
- Park, E.; Chung, S.W. ROS-mediated autophagy increases intracellular iron levels and ferroptosis by ferritin and transferrin receptor regulation. Cell Death Dis. 2019, 10, 822. [Google Scholar] [CrossRef] [Green Version]
- Moosmann, B.; Behl, C. Selenoproteins, cholesterol-lowering drugs, and the consequences: Revisiting of the mevalonate pathway. Trends Cardiovasc. Med. 2004, 14, 273–281. [Google Scholar] [CrossRef]
- Finkel, T. Signal transduction by mitochondrial oxidants. J. Biol. Chem. 2012, 287, 4434–4440. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Kang, R.; Tang, D. Signaling pathways and defense mechanisms of ferroptosis. FEBS J. 2021. ahead of print. [Google Scholar] [CrossRef]
- Liu, J.; Yang, M.; Kang, R.; Klionsky, D.J.; Tang, D. Autophagic degradation of the circadian clock regulator promotes ferroptosis. Autophagy 2019, 15, 2033–2035. [Google Scholar] [CrossRef]
- Chen, S.P.; Yang, J.; Zhang, Y.; Duan, C.Y.; Liu, Q.; Huang, Z.Y.; Xu, Y.; Zhou, L.; Xu, G.Q. Ubiquitin-conjugating enzyme UBE2O regulates cellular clock function by promoting the degradation of the transcription factor BMAL1. J. Biol. Chem. 2018, 293, 11296–11309. [Google Scholar] [CrossRef]
- Gossan, N.C.; Zhang, F.; Guo, B.Q.; Jin, D.; Yoshitane, H.; Yao, A.Y.; Glossop, N.; Zhang, Y.Q.; Fukada, Y.; Meng, Q.J. The E3 ubiquitin ligase UBE3A is an integral component of the molecular circadian clock through regulating the BMAL1 transcription factor. Nucleic Acids Res. 2014, 42, 5765–5775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Wang, X.L.; Li, Q.; Dong, X.X.; Zhang, J.S.; Yan, Q.C. HIF-1alpha SUMOylation affects the stability and transcriptional activity of HIF-1alpha in human lens epithelial cells. Graefes Arch. Clin. Exp. Ophthalmol. 2015, 253, 1279–1290. [Google Scholar] [CrossRef] [PubMed]
- Stockwell, B.R.; Friedmann Angeli, J.P.; Bayir, H.; Bush, A.I.; Conrad, M.; Dixon, S.J.; Fulda, S.; Gascon, S.; Hatzios, S.K.; Kagan, V.E.; et al. Ferroptosis: A Regulated Cell Death Nexus Linking Metabolism, Redox Biology, and Disease. Cell 2017, 171, 273–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Conrad, M. The Metabolic Underpinnings of Ferroptosis. Cell Metab. 2020, 32, 920–937. [Google Scholar] [CrossRef]
- Wang, P.; Lu, Y.Q. Ferroptosis: A Critical Moderator in the Life Cycle of Immune Cells. Front. Immunol. 2022, 13, 877634. [Google Scholar] [CrossRef]
- Zhang, Z.J.; Silveyra, E.; Jin, N.E.; Ribelayga, C.P. A congenic line of the C57BL/6J mouse strain that is proficient in melatonin synthesis. J. Pineal Res. 2018, 65, e12509. [Google Scholar] [CrossRef]
- Zheng, B.H.; Albrecht, U.; Kaasik, K.; Sage, M.; Lu, W.Q.; Vaishnav, S.; Li, Q.; Sun, Z.S.; Eichele, G.; Bradley, A.; et al. Nonredundant roles of the mPer1 and mPer2 genes in the mammalian circadian clock. Cell 2001, 105, 683–694. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, R.; Zhang, S.; Yu, J.; Yu, X.; Wang, J.; Qiu, Y.; Zhou, W.; Wang, F.; Ren, F.; Liu, Z. Per1/Per2 knockout Affects Spleen Immune Function in Elderly Mice via Inducing Spleen Lymphocyte Ferroptosis. Int. J. Mol. Sci. 2022, 23, 12962. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232112962
He R, Zhang S, Yu J, Yu X, Wang J, Qiu Y, Zhou W, Wang F, Ren F, Liu Z. Per1/Per2 knockout Affects Spleen Immune Function in Elderly Mice via Inducing Spleen Lymphocyte Ferroptosis. International Journal of Molecular Sciences. 2022; 23(21):12962. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232112962
Chicago/Turabian StyleHe, Ruyi, Shijie Zhang, Jiale Yu, Xiaojie Yu, Jian Wang, Yi Qiu, Wenting Zhou, Fangyi Wang, Feng Ren, and Zhiguo Liu. 2022. "Per1/Per2 knockout Affects Spleen Immune Function in Elderly Mice via Inducing Spleen Lymphocyte Ferroptosis" International Journal of Molecular Sciences 23, no. 21: 12962. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232112962