
PRG3 and PRG5 C-Termini: Important Players in Early Neuronal Differentiation
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
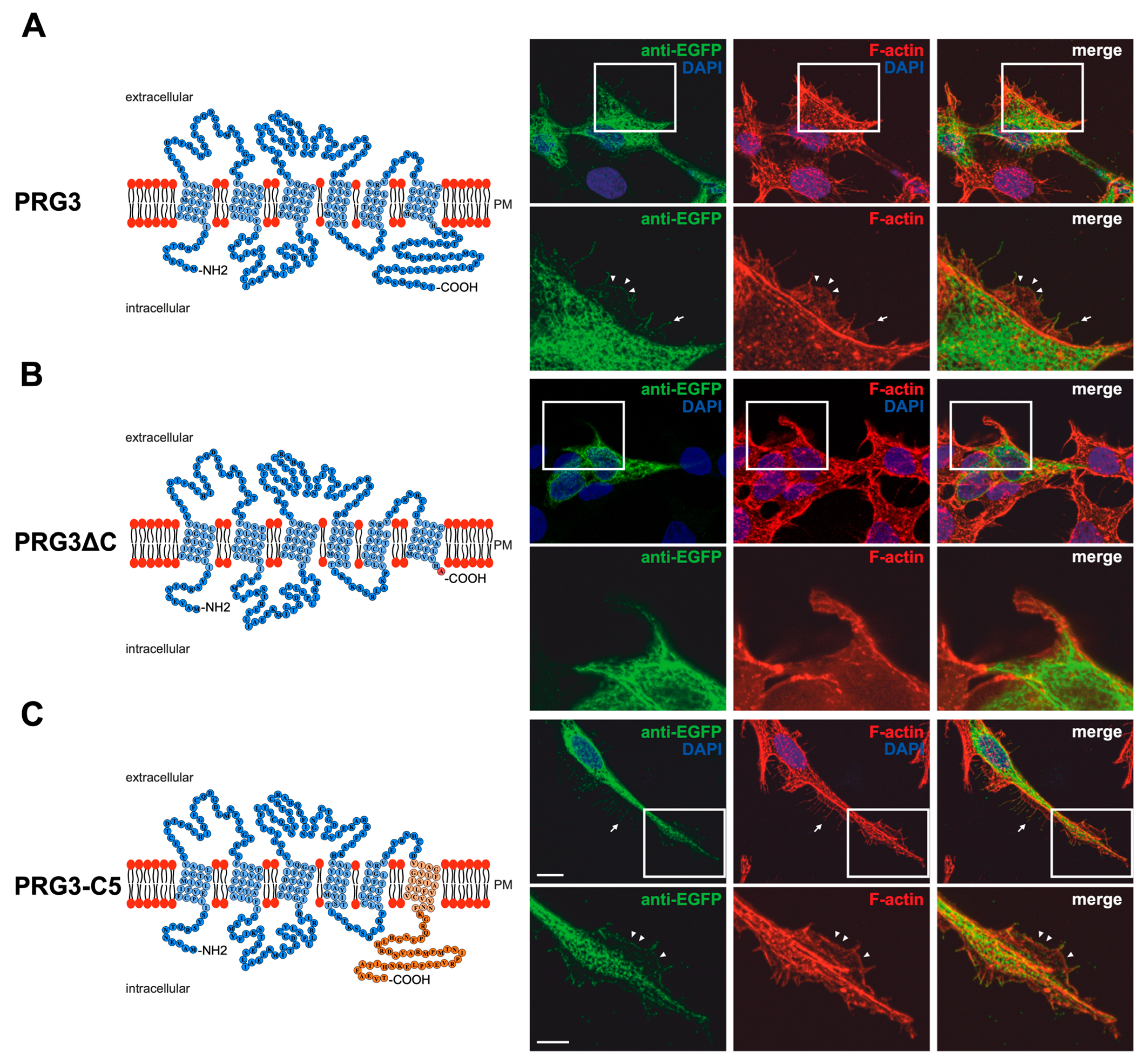
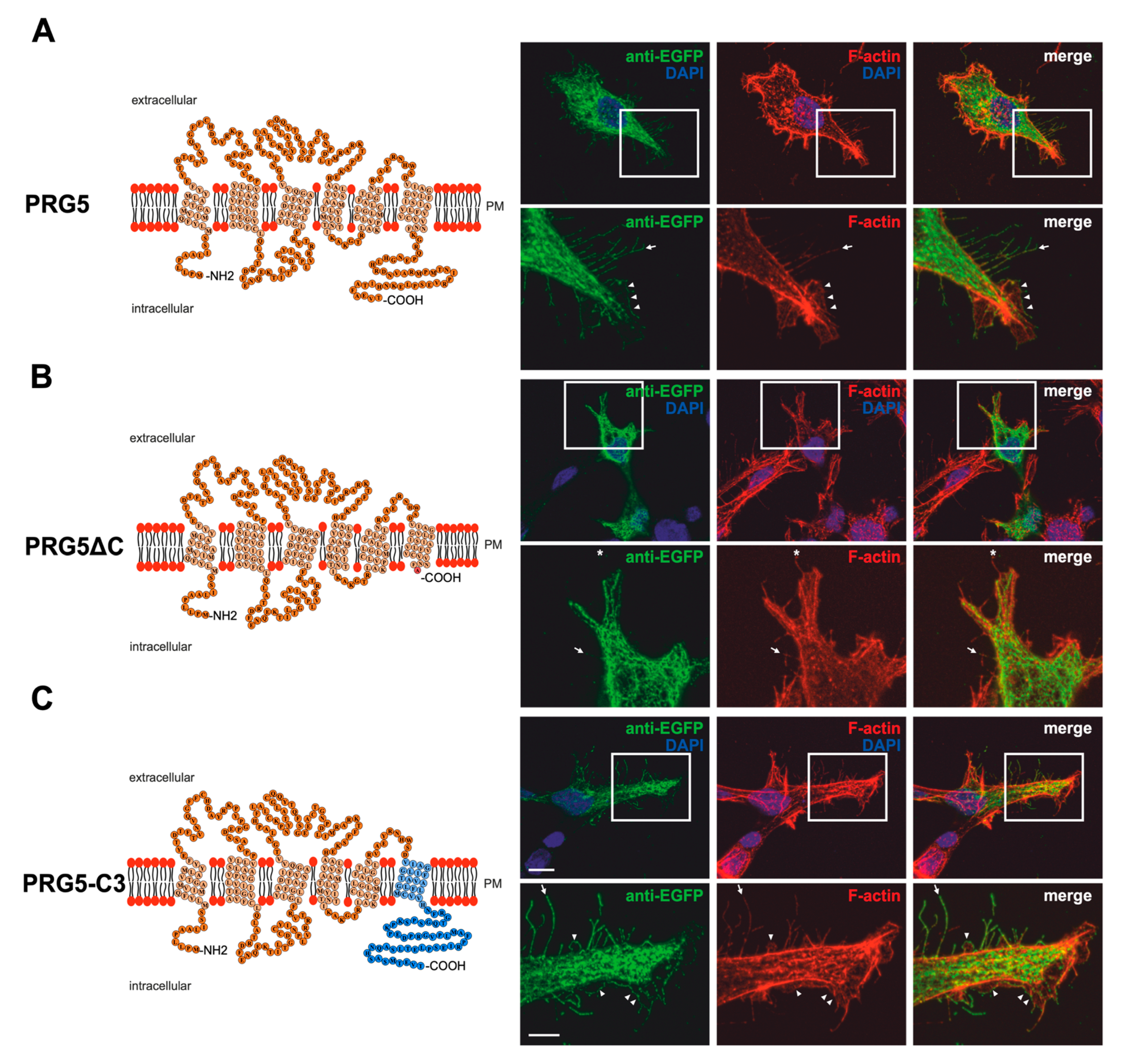
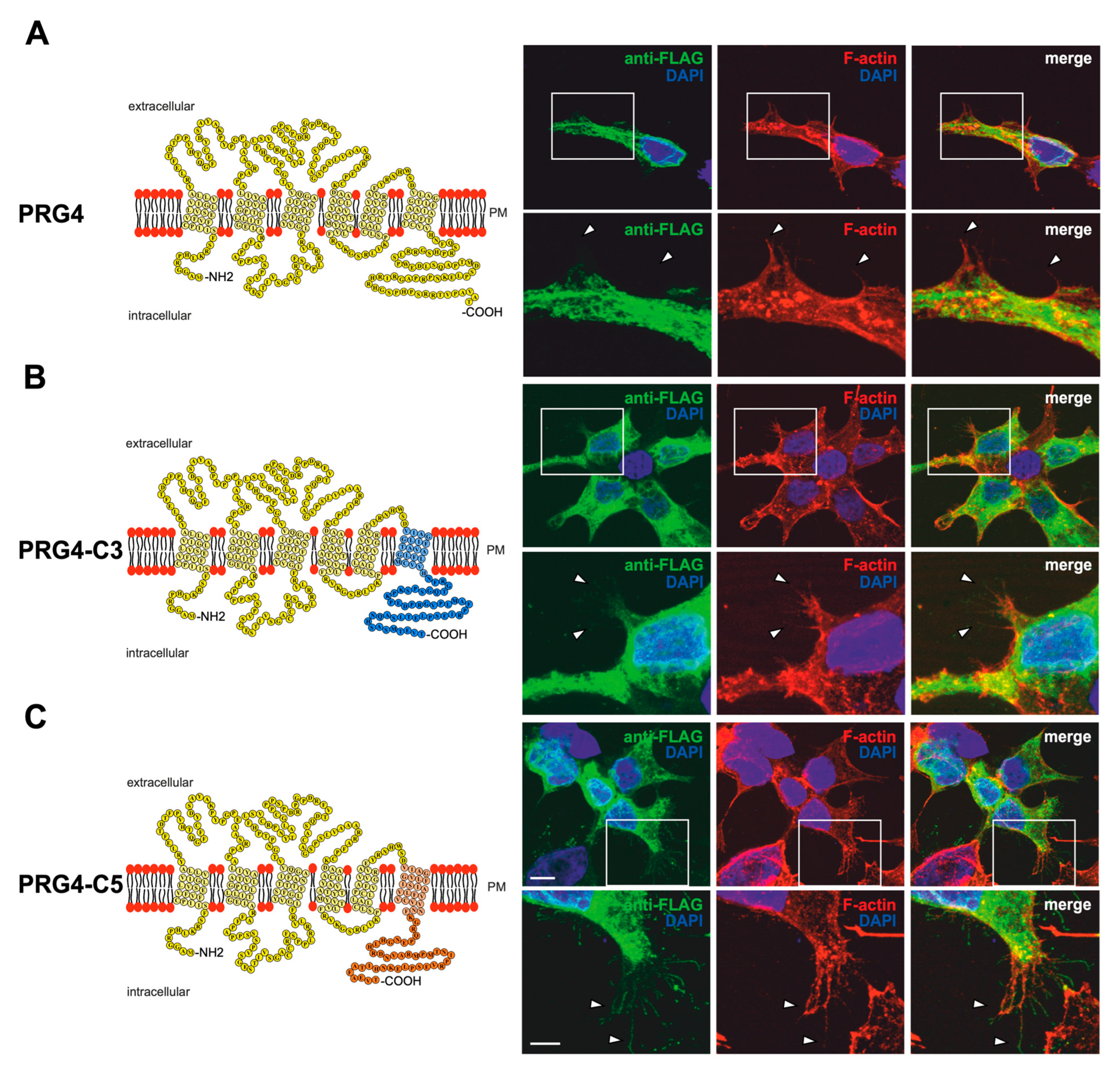
2.1. Membrane Expression of PRG3 and PRG5, also with Swapped C-Termini, but Not with PRG3∆C and PRG5∆C
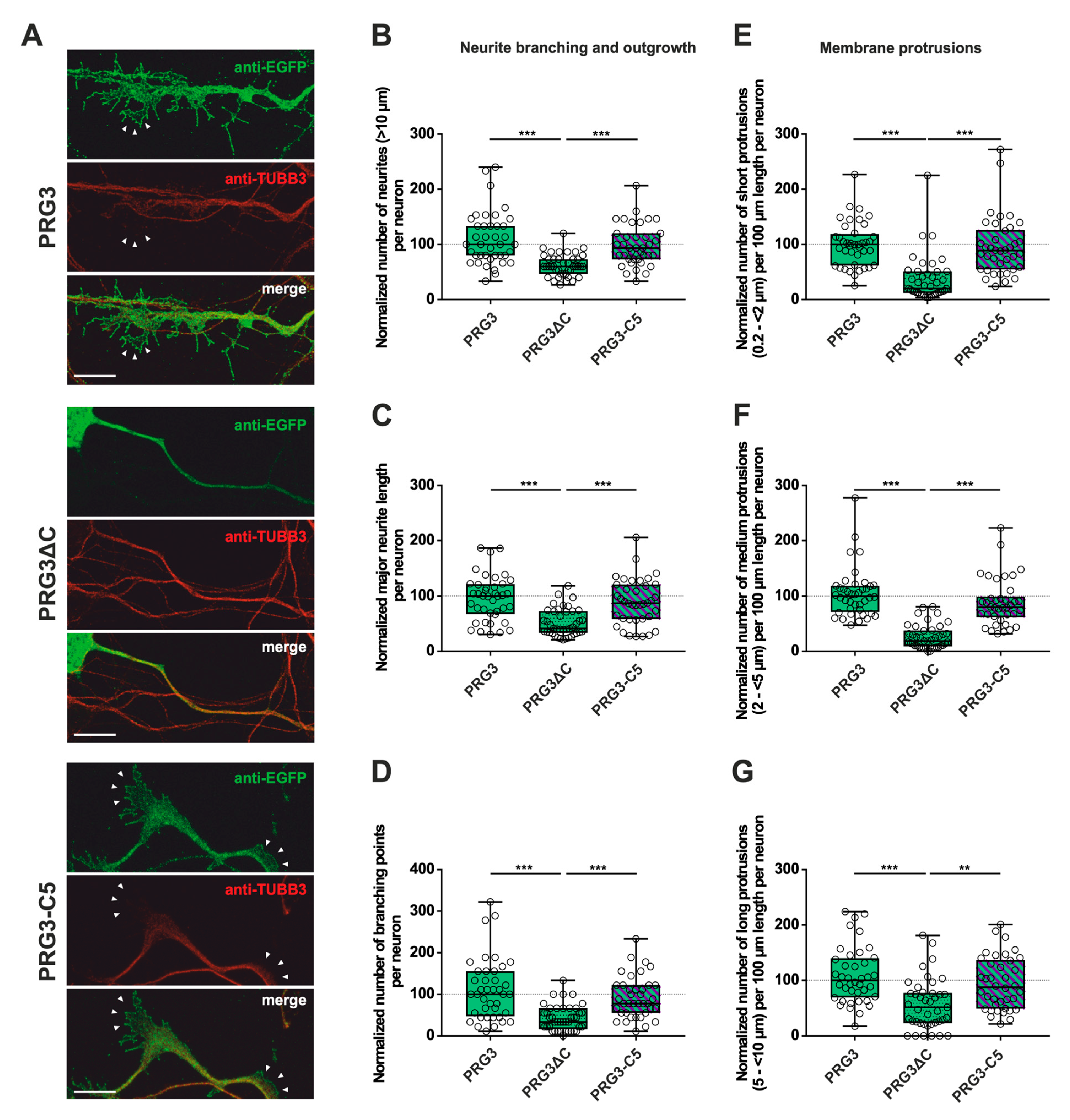
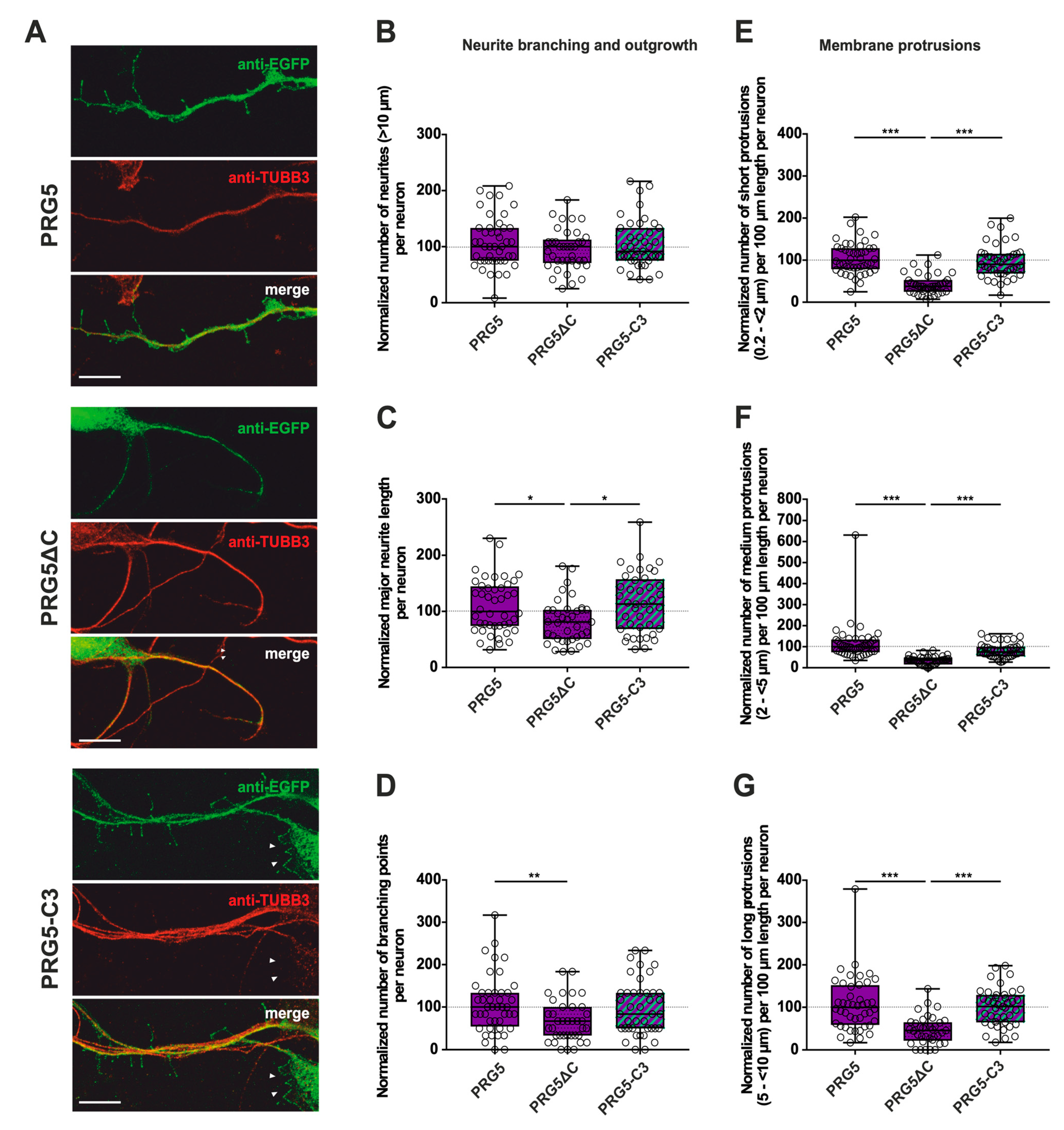
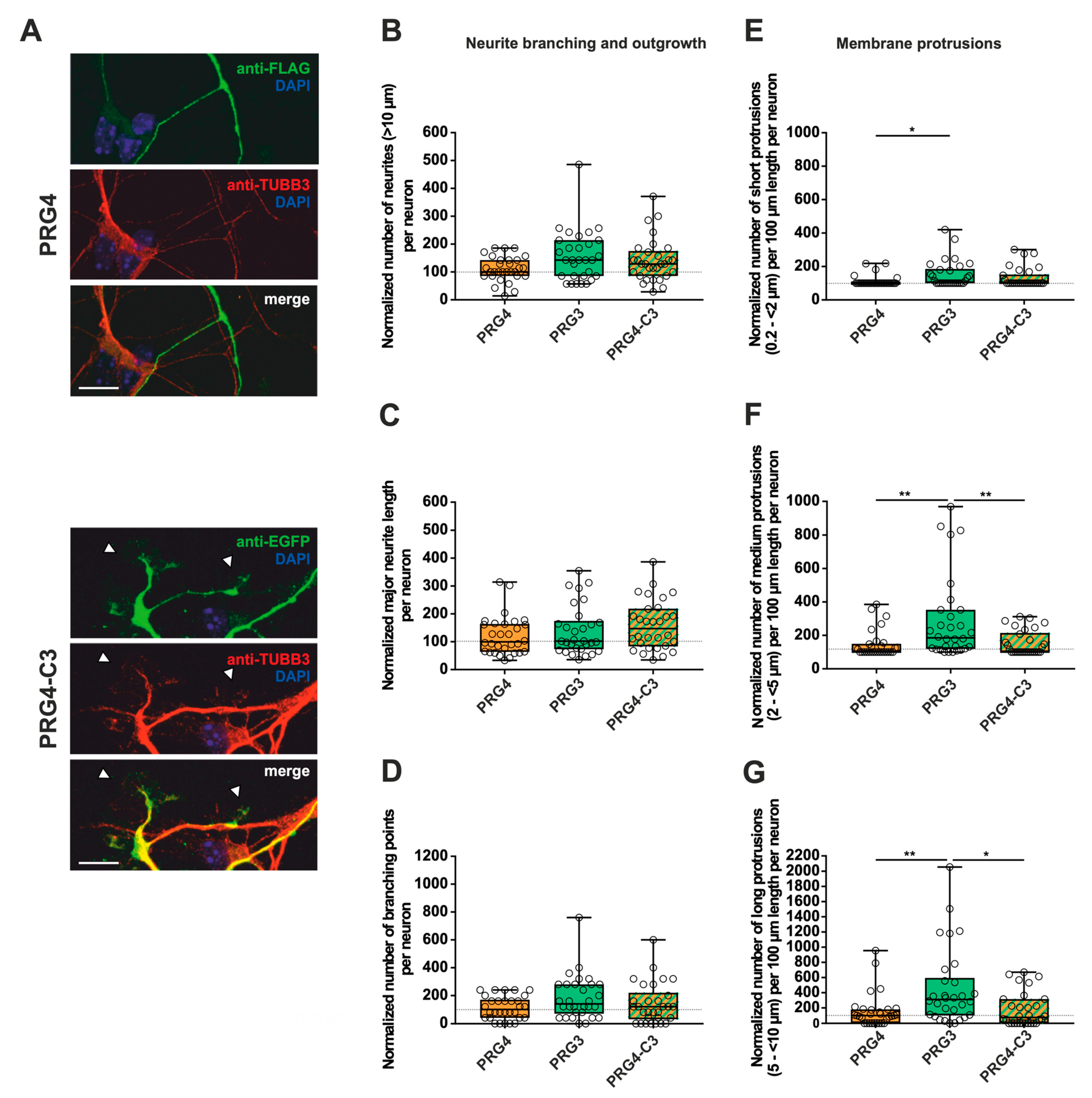
2.2. PRG3 and PRG5 C-Termini Are Important Players in Early Neuronal Differentiation, No Matter Whether the C-Term Came from One or the Other Protein
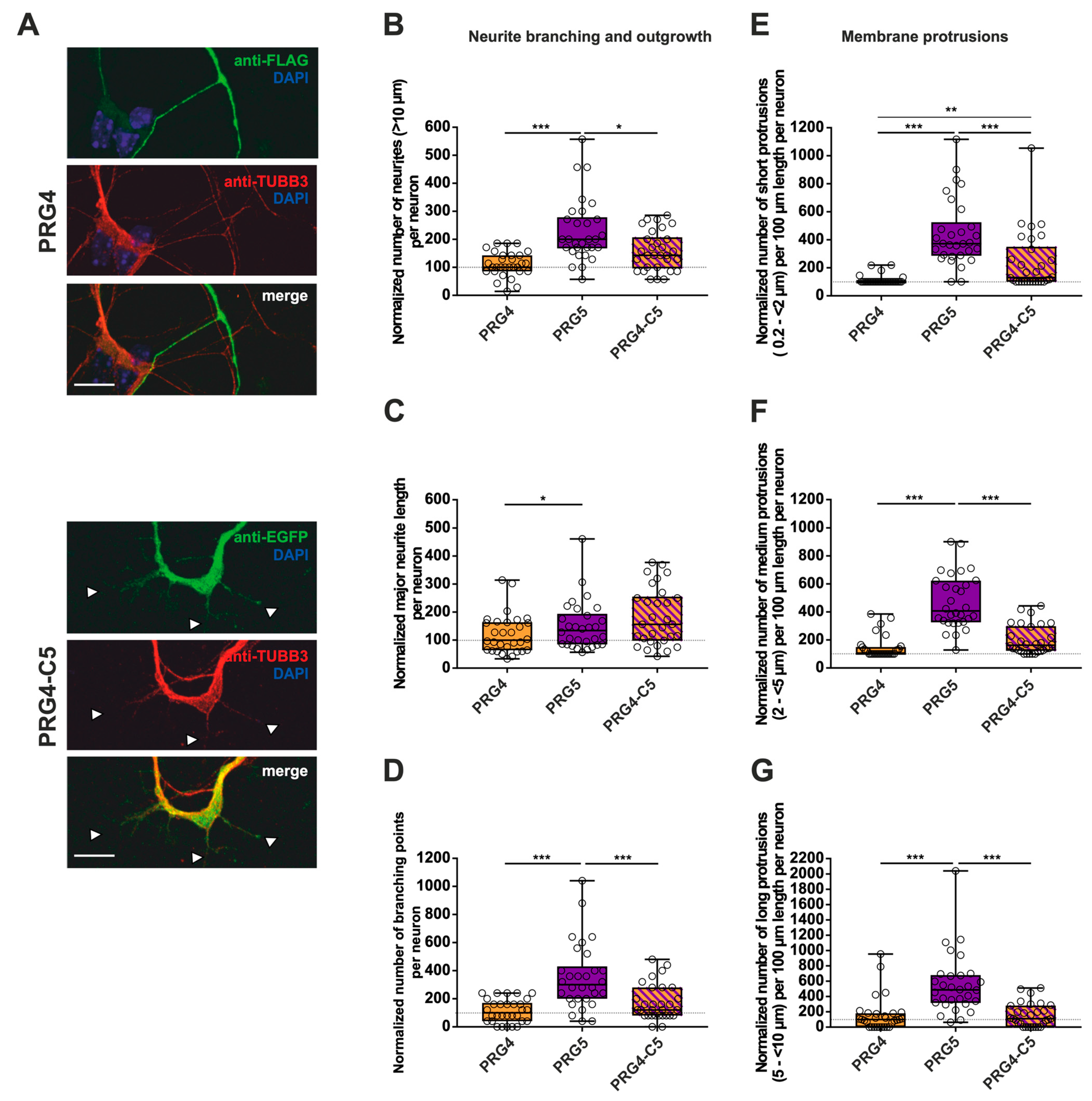
2.3. PRG3 and PRG5 C-Terminal Domains Induce Morphological Changes in PRG4 Overexpressing Non-Neuronal Cells and Neurons—But to a Lesser Extent
3. Discussion
3.1. Plasma Membrane Localization Is Essential for PRG3 and PRG5 Function
3.2. The C-Terminus of PRG3 and PRG5 Plays a Significant Role in Major Neurite Growth
3.3. Only the Full-Length PRG3 or PRG5 Protein Enhances Neurite Branching
3.4. The C-Terminus of PRG3 or PRG5 Is Insufficient to Induce Membrane Protrusions
4. Material and Methods
4.1. Animals
4.2. Plasmids
4.3. Primary Mouse Hippocampal Neuron Cultures
4.4. Cell Culture and Transfection
4.5. Western Blot Analysis
4.6. Immunocytochemistry
4.7. Image Acquisition and Morphological Analysis
4.8. Statistics and Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Götz, M.; Huttner, W.B. The cell biology of neurogenesis. Nat. Rev. Mol. Cell Biol. 2005, 6, 777–788. [Google Scholar] [CrossRef]
- Stiles, J.; Jernigan, T.L. The basics of brain development. Neuropsychol. Rev. 2010, 20, 327–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arimura, N.; Kaibuchi, K. Key regulators in neuronal polarity. Neuron 2005, 48, 881–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yogev, S.; Shen, K. Establishing Neuronal Polarity with Environmental and Intrinsic Mechanisms. Neuron 2017, 96, 638–650. [Google Scholar] [CrossRef] [Green Version]
- Govek, E.E.; Newey, S.E.; Van Aelst, L. The role of the Rho GTPases in neuronal development. Genes Dev. 2005, 19, 1–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, J.S.; Dotti, C.G. Breaking the neuronal sphere: Regulation of the actin cytoskeleton in neuritogenesis. Nat. Rev. Neurosci. 2002, 3, 694–704. [Google Scholar] [CrossRef] [PubMed]
- Broggini, T.; Nitsch, R.; Savaskan, N.E. Plasticity-related gene 5 (PRG5) induces filopodia and neurite growth and impedes lysophosphatidic acid- and nogo-A-mediated axonal retraction. Mol. Biol. Cell 2010, 21, 521–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coiro, P.; Stoenica, L.; Strauss, U.; Brauer, A.U. Plasticity-related gene 5 promotes spine formation in murine hippocampal neurons. J. Biol. Chem. 2014, 289, 24956–24970. [Google Scholar] [CrossRef] [Green Version]
- Velmans, T.; Battefeld, A.; Geist, B.; Farres, A.S.; Strauss, U.; Brauer, A.U. Plasticity-related gene 3 promotes neurite shaft protrusion. BMC Neurosci. 2013, 14, 36. [Google Scholar] [CrossRef] [Green Version]
- Broggini, T.; Schnell, L.; Ghoochani, A.; Mateos, J.M.; Buchfelder, M.; Wiendieck, K.; Schafer, M.K.; Eyupoglu, I.Y.; Savaskan, N.E. Plasticity Related Gene 3 (PRG3) overcomes myelin-associated growth inhibition and promotes functional recovery after spinal cord injury. Aging 2016, 8, 2463–2487. [Google Scholar] [CrossRef]
- Brauer, A.U.; Nitsch, R. Plasticity-related genes (PRGs/LRPs): A brain-specific class of lysophospholipid-modifying proteins. Biochim. Biophys. Acta 2008, 1781, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Peeva, G.P.; Angelova, S.K.; Guntinas-Lichius, O.; Streppel, M.; Irintchev, A.; Schutz, U.; Popratiloff, A.; Savaskan, N.E.; Brauer, A.U.; Alvanou, A.; et al. Improved outcome of facial nerve repair in rats is associated with enhanced regenerative response of motoneurons and augmented neocortical plasticity. Eur. J. Neurosci. 2006, 24, 2152–2162. [Google Scholar] [CrossRef] [PubMed]
- Brauer, A.U.; Savaskan, N.E.; Kuhn, H.; Prehn, S.; Ninnemann, O.; Nitsch, R. A new phospholipid phosphatase, PRG-1, is involved in axon growth and regenerative sprouting. Nat. Neurosci. 2003, 6, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Savaskan, N.E.; Brauer, A.U.; Nitsch, R. Molecular cloning and expression regulation of PRG-3, a new member of the plasticity-related gene family. Eur. J. Neurosci. 2004, 19, 212–220. [Google Scholar] [CrossRef]
- Sigal, Y.J.; Quintero, O.A.; Cheney, R.E.; Morris, A.J. Cdc42 and ARP2/3-independent regulation of filopodia by an integral membrane lipid-phosphatase-related protein. J. Cell Sci. 2007, 120 Pt 2, 340–352. [Google Scholar] [CrossRef] [Green Version]
- Strauss, U.; Brauer, A.U. Current views on regulation and function of plasticity-related genes (PRGs/LPPRs) in the brain. Biochim. Et Biophys. Acta 2013, 1831, 133–138. [Google Scholar] [CrossRef]
- Fuchs, J.; Bareesel, S.; Kroon, C.; Polyzou, A.; Eickholt, B.J.; Leondaritis, G. Plasma membrane phospholipid phosphatase-related proteins as pleiotropic regulators of neuron growth and excitability. Front. Mol. Neurosci. 2022, 15, 984655. [Google Scholar] [CrossRef]
- Gross, I.; Tschigor, T.; Salman, A.L.; Yang, F.; Luo, J.; Vonk, D.; Hipp, M.S.; Neidhardt, J.; Bräuer, A.U. Systematic expression analysis of plasticity-related genes in mouse brain development brings PRG4 into play. Dev. Dyn. 2022, 251, 714–728. [Google Scholar] [CrossRef]
- Gross, I.; Brandt, N.; Vonk, D.; Köper, F.; Wöhlbrand, L.; Rabus, R.; Witt, M.; Heep, A.; Plösch, T.; Hipp, M.S.; et al. Plasticity-Related Gene 5 Is Expressed in a Late Phase of Neurodifferentiation after Neuronal Cell-Fate Determination. Front. Cell. Neurosci. 2022, 16, 797588. [Google Scholar] [CrossRef]
- Yu, P.; Agbaegbu, C.; Malide, D.A.; Wu, X.; Katagiri, Y.; Hammer, J.A.; Geller, H.M. Cooperative interactions of LPPR family members in membrane localization and alteration of cellular morphology. J. Cell Sci. 2015, 128, 3210–3222. [Google Scholar] [CrossRef]
- Kaech, S.; Banker, G. Culturing hippocampal neurons. Nat. Protoc. 2006, 1, 2406–2415. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Bittermann-Rummel, P.; Yakubov, E.; Chen, D.; Broggini, T.; Sehm, T.; Hatipoglu Majernik, G.; Hock, S.W.; Schwarz, M.; Engelhorn, T.; et al. PRG3 induces Ras-dependent oncogenic cooperation in gliomas. Oncotarget 2016, 7, 26692–26708. [Google Scholar] [CrossRef]
- Dotti, C.G.; Sullivan, C.A.; Banker, G.A. The establishment of polarity by hippocampal neurons in culture. J. Neurosci. Off. J. Soc. Neurosci. 1988, 8, 1454–1468. [Google Scholar] [CrossRef] [Green Version]
- Fink, K.L.; Lopez-Giraldez, F.; Kim, I.J.; Strittmatter, S.M.; Cafferty, W.B.J. Identification of Intrinsic Axon Growth Modulators for Intact CNS Neurons after Injury. Cell Rep. 2017, 18, 2687–2701. [Google Scholar] [CrossRef] [PubMed]
- Brewer, G.J.; Torricelli, J.R.; Evege, E.K.; Price, P.J. Optimized survival of hippocampal neurons in B27-supplemented Neurobasal, a new serum-free medium combination. J. Neurosci. Res. 1993, 35, 567–576. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Independent Variable | Group 2 | |||
|---|---|---|---|---|
| Group 1 | PRG3 | PRG3∆C | PRG3-C5 | |
| Number of neurites (>10 µm) per neuron | PRG3 | <0.0001 | 1.000 | |
| PRG3∆C | <0.0001 | <0.0001 | ||
| PRG3-C5 | 1.000 | <0.0001 | ||
| Number of major neurite lengths per neuron | PRG3 | <0.0001 | 1.000 | |
| PRG3∆C | <0.0001 | <0.0001 | ||
| PRG3-C5 | 1.000 | <0.0001 | ||
| Number of branching points per neuron | PRG3 | <0.0001 | 1.000 | |
| PRG3∆C | <0.0001 | <0.0001 | ||
| PRG3-C5 | 1.000 | <0.0001 | ||
| Number of short protrusions (0.2–<2 µm) per 100 µm length per neuron | PRG3 | <0.0001 | 1.000 | |
| PRG3∆C | <0.0001 | <0.0001 | ||
| PRG3-C5 | 1.000 | <0.0001 | ||
| Number of medium protrusions (2–<5 µm) per 100 µm length per neuron | PRG3 | <0.0001 | 0.373 | |
| PRG3∆C | <0.0001 | <0.0001 | ||
| PRG3-C5 | 0.373 | <0.0001 | ||
| Number of long protrusions (5–<10 µm) per 100 µm length per neuron | PRG3 | <0.0001 | 0.699 | |
| PRG3∆C | <0.0001 | 0.003 | ||
| PRG3-C5 | 0.699 | 0.003 | ||
| Independent Variable | Group 2 | |||
|---|---|---|---|---|
| Group 1 | PRG5 | PRG5∆C | PRG5-C3 | |
| Number of neurites (>10 µm) per neuron | PRG5 | n.s. | n.s. | n.s. |
| PRG5∆C | ||||
| PRG5-C3 | ||||
| Number of major neurite lengths per neuron | PRG5 | 0.042 | 1.000 | |
| PRG5∆C | 0.042 | 0.014 | ||
| PRG5-C3 | 1.000 | 0.014 | ||
| Number of branching points per neuron | PRG5 | 0.013 | 1.000 | |
| PRG5∆C | 0.013 | 0.061 | ||
| PRG5-C3 | 1.000 | 0.061 | ||
| Number of short protrusions (0.2–<2 µm) per 100 µm length per neuron | PRG5 | <0.0001 | 1.000 | |
| PRG5∆C | <0.0001 | <0.0001 | ||
| PRG5-C3 | 1.000 | <0.0001 | ||
| Number of medium protrusions (2–<5 µm) per 100 µm length per neuron | PRG5 | <0.0001 | 0.076 | |
| PRG5∆C | <0.0001 | <0.0001 | ||
| PRG5-C3 | 0.076 | <0.0001 | ||
| Number of long protrusions (5–<10 µm) per 100 µm length per neuron | PRG5 | <0.0001 | 1.000 | |
| PRG5∆C | <0.0001 | <0.0001 | ||
| PRG5-C3 | 1.000 | <0.0001 | ||
| Independent Variable | Group 2 | |||
|---|---|---|---|---|
| Group 1 | PRG4 | PRG3 | PRG4-C3 | |
| Number of neurites (>10 µm) per neuron | PRG4 | n.s. | n.s. | n.s. |
| PRG3 | ||||
| PRG4-C3 | ||||
| Number of major neurite lengths per neuron | PRG4 | n.s. | n.s | n.s. |
| PRG3 | ||||
| PRG4-C3 | ||||
| Number of branching points per neuron | PRG4 | n.s. | n.s. | n.s. |
| PRG3 | ||||
| PRG4-C3 | ||||
| Number of short protrusions (0.2–<2 µm) per 100 µm length per neuron | PRG4 | 0.023 | 0.502 | |
| PRG3 | 0.023 | 0.597 | ||
| PRG4-C3 | 0.502 | 0.597 | ||
| Number of medium protrusions (2–<5 µm) per 100 µm length per neuron | PRG4 | 0.002 | 1.000 | |
| PRG3 | 0.002 | 0.005 | ||
| PRG4-C3 | 1.000 | 0.005 | ||
| Number of long protrusions (5–<10 µm) per 100 µm length per neuron | PRG4 | 0.007 | 1.000 | |
| PRG3 | 0.007 | 0.010 | ||
| PRG4-C3 | 1.000 | 0.010 | ||
| Independent Variable | Group 2 | |||
|---|---|---|---|---|
| Group 1 | PRG4 | PRG5 | PRG4-C5 | |
| Number of neurites (>10 µm) per neuron | PRG4 | <0.0001 | 0.052 | |
| PRG5 | <0.0001 | 0.020 | ||
| PRG4-C5 | 0.052 | 0.020 | ||
| Number of major neurite lengths per neuron | PRG4 | 0.516 | 0.040 | |
| PRG5 | 0.516 | 0.805 | ||
| PRG4-C5 | 0.040 | 0.805 | ||
| Number of branching points per neuron | PRG4 | <0.0001 | 0.314 | |
| PRG5 | <0.0001 | 0.002 | ||
| PRG4-C5 | 0.314 | 0.002 | ||
| Number of short protrusions (0.2–<2 µm) per 100 µm length per neuron | PRG4 | <0.0001 | 0.005 | |
| PRG5 | <0.0001 | 0.001 | ||
| PRG4-C5 | 0.005 | 0.001 | ||
| Number of medium protrusions (2–<5 µm) per 100 µm length per neuron | PRG4 | <0.0001 | 0.098 | |
| PRG5 | <0.0001 | <0.0001 | ||
| PRG4-C5 | 0.098 | <0.0001 | ||
| Number of long protrusions (5–<10 µm) per 100 µm length per neuron | PRG4 | <0.0001 | 1.000 | |
| PRG5 | <0.0001 | <0.0001 | ||
| PRG4-C5 | 1.000 | <0.0001 | ||
| Plasmid | Encoded Protein | Abbreviation | Source |
|---|---|---|---|
| pEGFP-N1 | EGFP | EGFP | Clontech/Takara Holdings Shimogyo-ku, Kyoto, Japan |
| pEGFP-N1-mPRG3 | mPRG3-EGFP | PRG3 | [9,14] |
| pEGFP-N1-mPRG3ΔC | mPRG3ΔC-EGFP | PRG3ΔC | This paper |
| pEGFP-N1-mPRG3Δ-C5 | mPRG3ΔC-C5-EGFP | PRG3-C5 | This paper |
| pEGFP-N1-rPRG5 | rPRG5-EGFP | PRG5 | [7,8] |
| pEGFP-N1-rPRG5ΔC | rPRG5ΔC-EGFP | PRG5ΔC | This paper |
| pEGFP-N1-rPRG5Δ-C3 | rPRG5Δ-C3-EGFP | PRG5-C3 | This paper |
| pFLAG-CMV2-mPRG4 | mPRG4-FLAG | PRG4 | [8,14] |
| pEGFP-N1-mPRG4ΔC-C3 | mPRG4ΔC-C3-EGFP | PRG4-C3 | This paper |
| pEGFP-N1-mPRG4ΔC-C5 | mPRG4ΔC-C5-EGFP | PRG4-C5 | This paper |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brandt, N.; Willmer, J.P.; Ayon-Olivas, M.S.; Banicka, V.; Witt, M.; Wree, A.; Groß, I.; Gläser, A.; Hausmann, J.; Bräuer, A.U. PRG3 and PRG5 C-Termini: Important Players in Early Neuronal Differentiation. Int. J. Mol. Sci. 2022, 23, 13007. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113007
Brandt N, Willmer JP, Ayon-Olivas MS, Banicka V, Witt M, Wree A, Groß I, Gläser A, Hausmann J, Bräuer AU. PRG3 and PRG5 C-Termini: Important Players in Early Neuronal Differentiation. International Journal of Molecular Sciences. 2022; 23(21):13007. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113007
Chicago/Turabian StyleBrandt, Nicola, Jan Philipp Willmer, Maurilyn S. Ayon-Olivas, Veronika Banicka, Martin Witt, Andreas Wree, Isabel Groß, Anne Gläser, Jens Hausmann, and Anja U. Bräuer. 2022. "PRG3 and PRG5 C-Termini: Important Players in Early Neuronal Differentiation" International Journal of Molecular Sciences 23, no. 21: 13007. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113007