

A Hypothalamic Mechanism Regulates the Duration of a Migraine Attack: Insights from Microstructural and Temporal Complexity of Cortical Functional Networks Analysis

 ,
,  , , , , and
, , , , and 
Abstract
:1. Introduction
2. Results
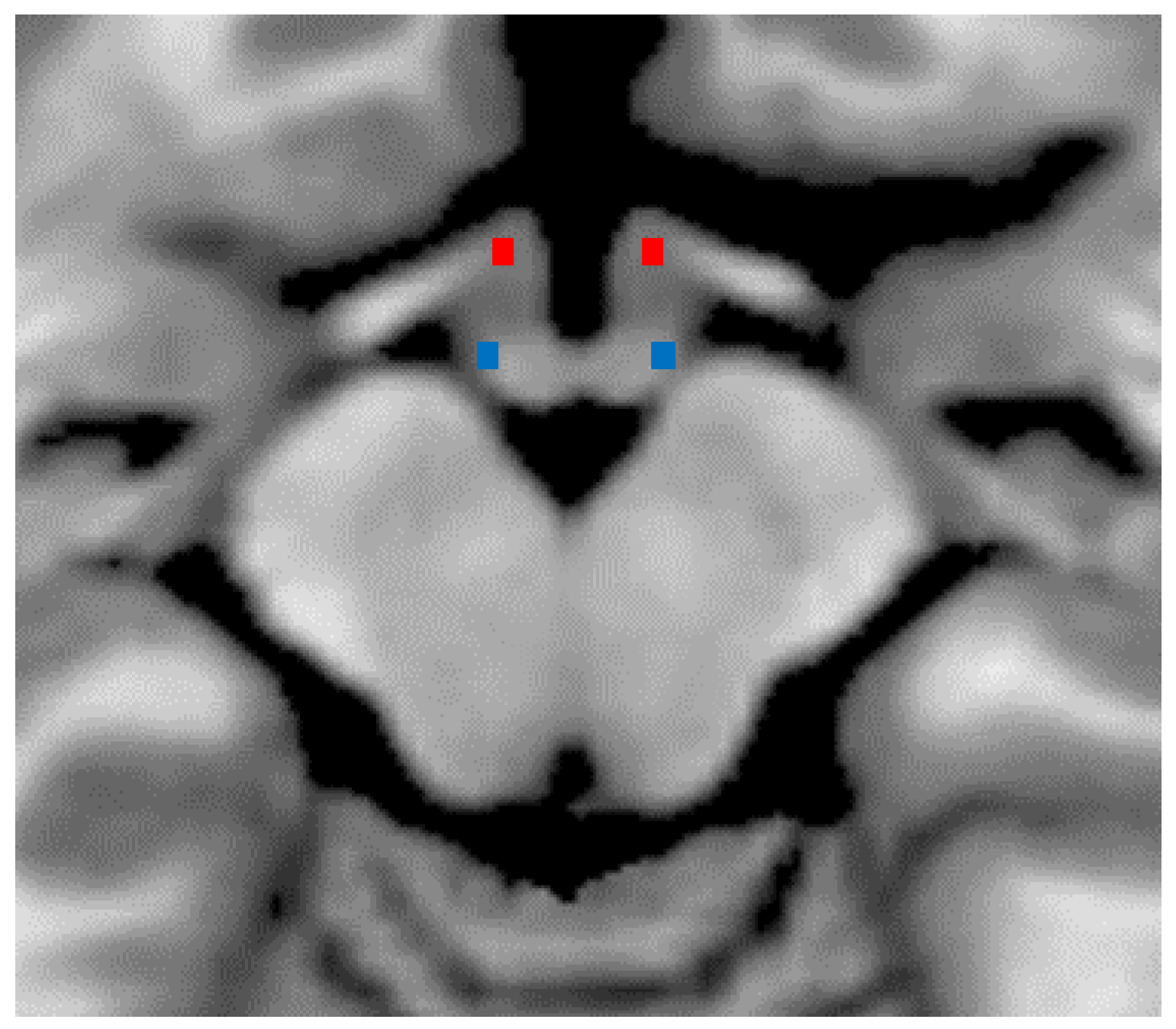
2.1. Characterization of Hypothalamic DTI
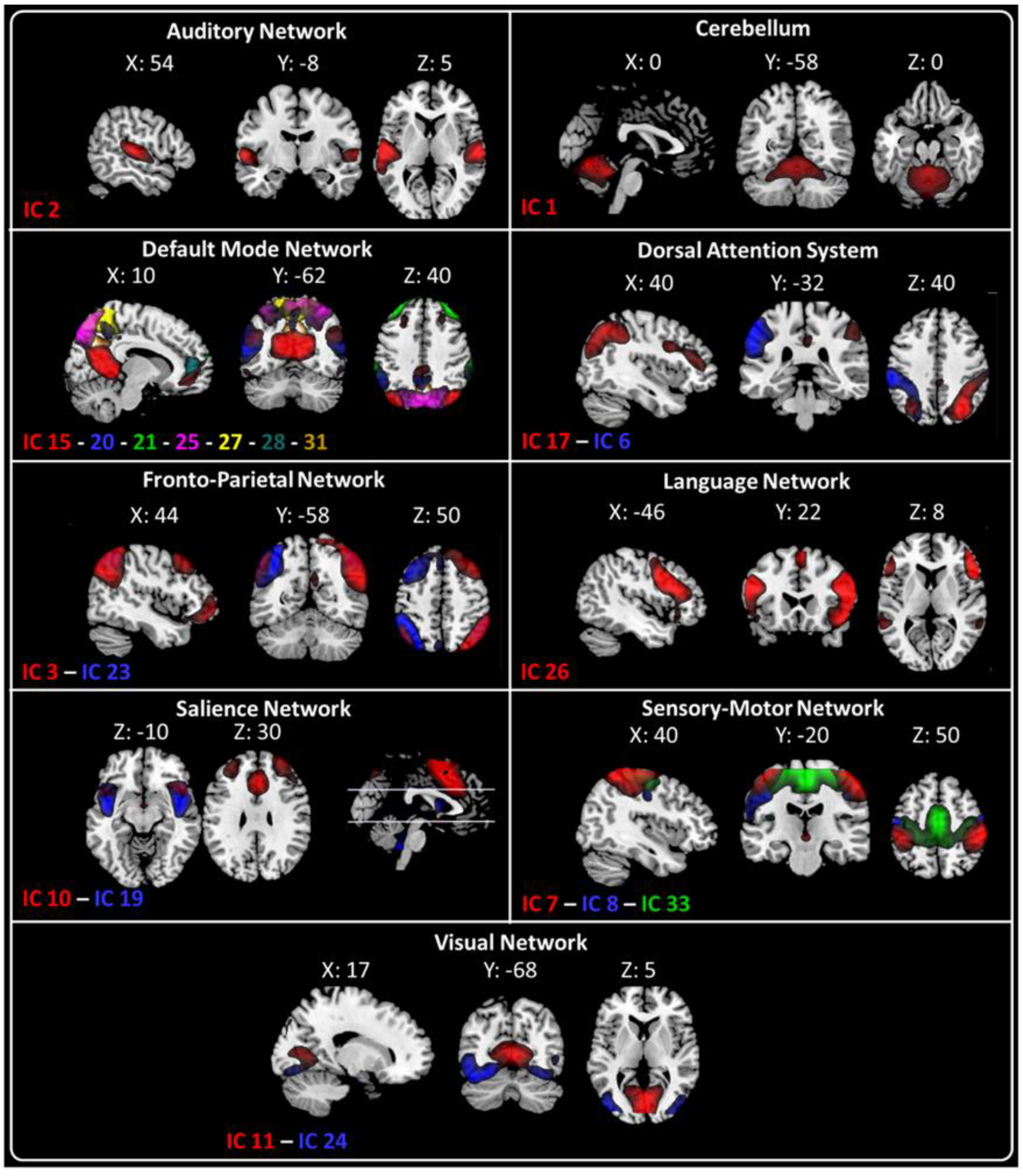
2.2. fMRI Resting-State Networks
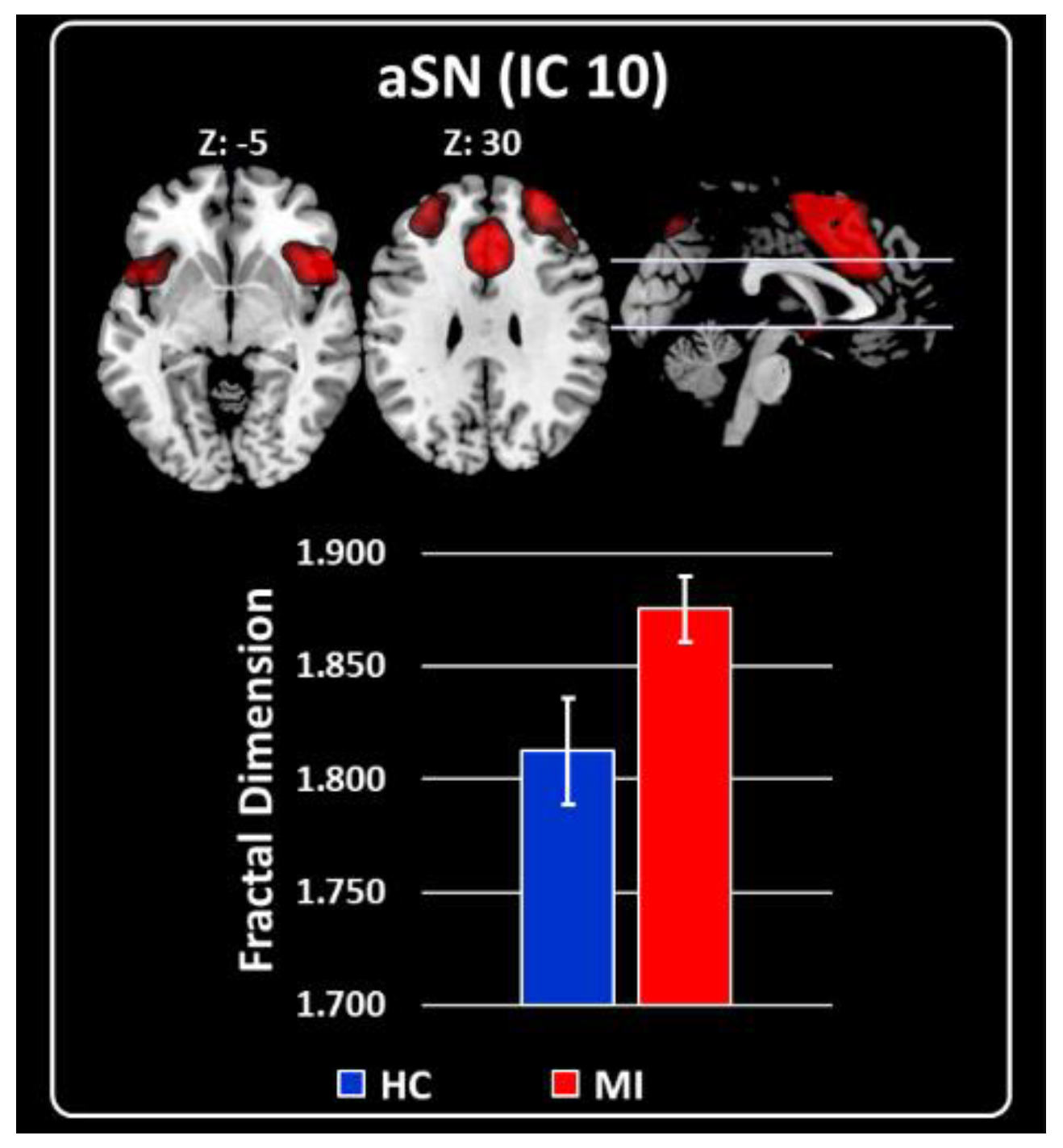
2.3. Characterization of RSNs Using Higuchi’s Fractal Dimension
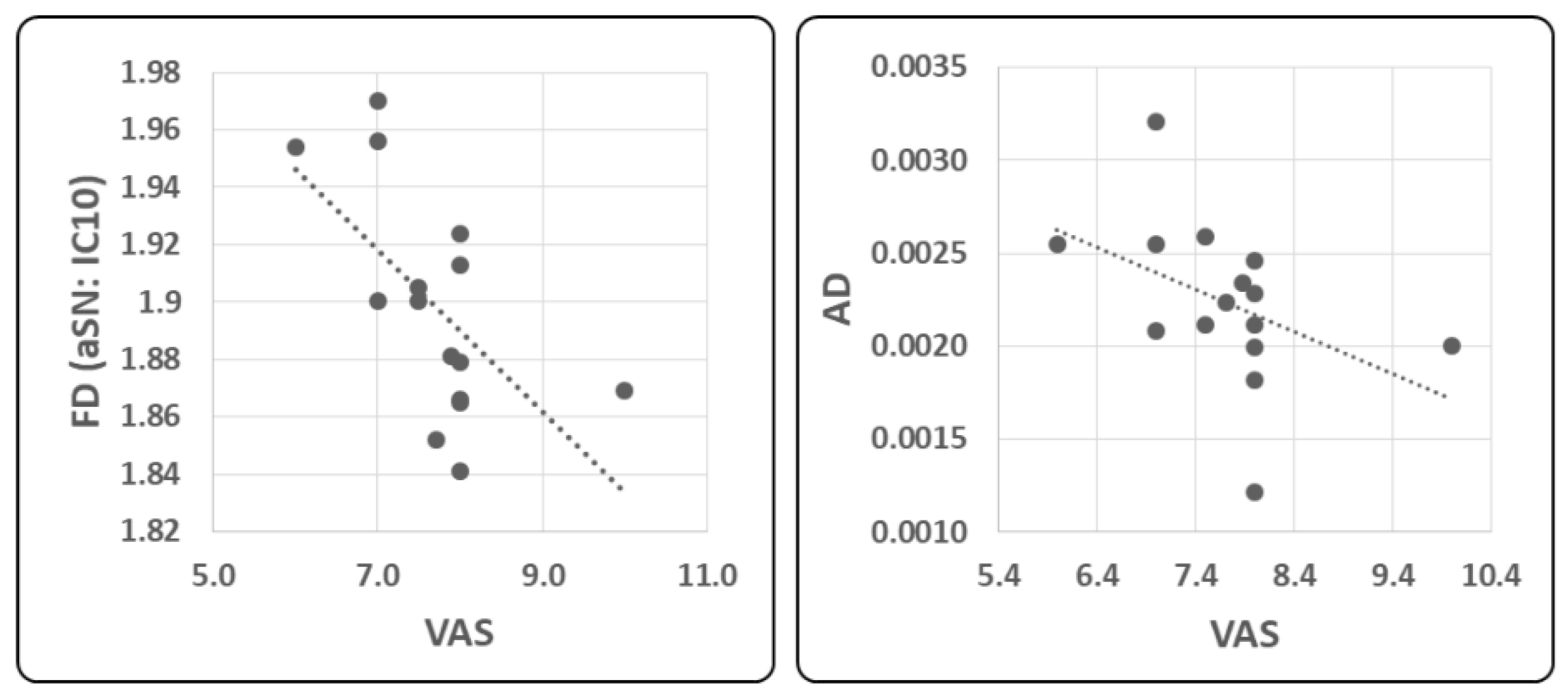
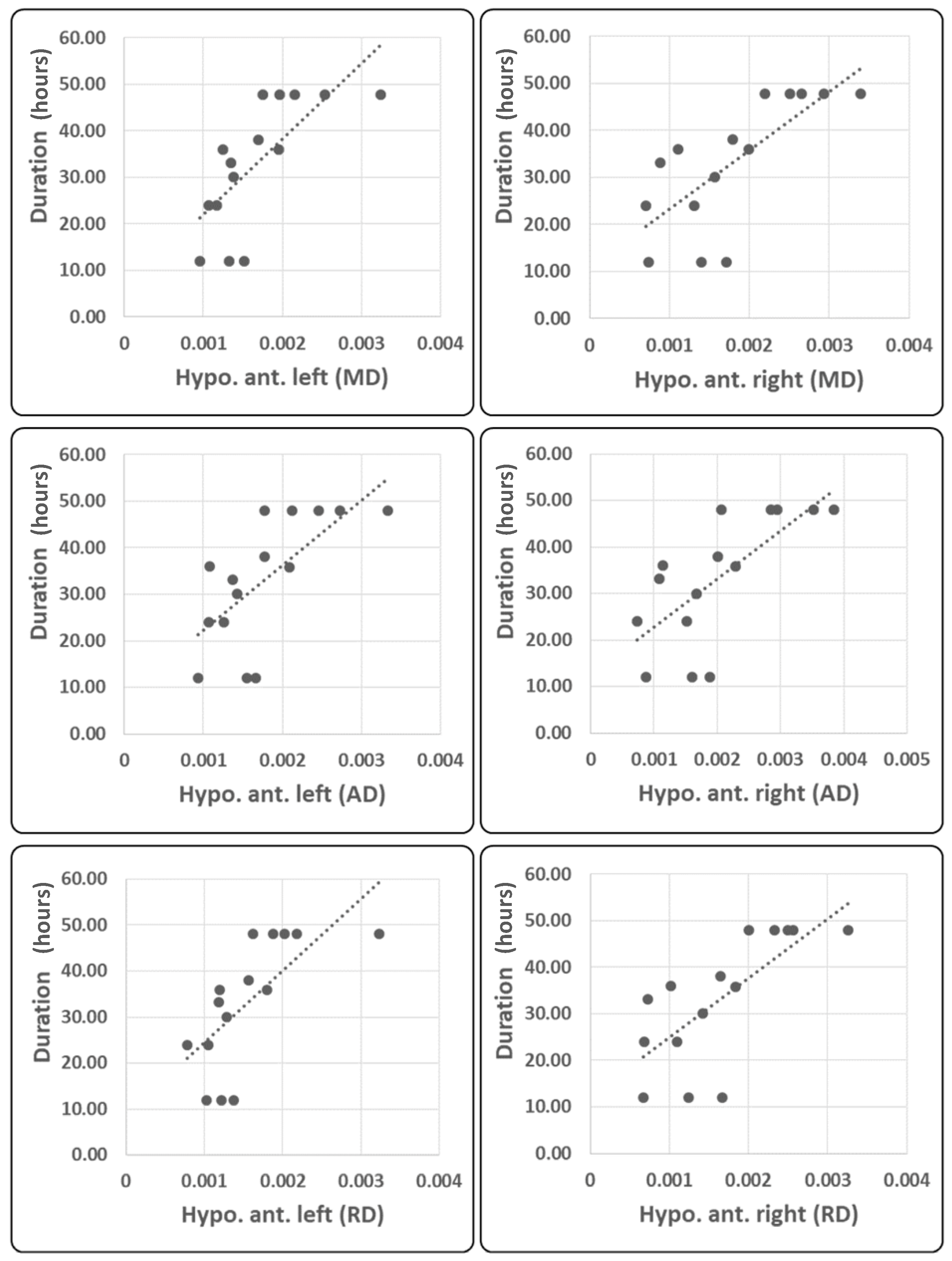
2.4. Correlation Analysis
3. Discussion
3.1. Fractal Dimensionality
3.2. Microstructure of the Hypothalamus
4. Materials and Methods
4.1. Participants
4.2. Diffusion Tensor Imaging (DTI) Data Acquisition and Analysis
4.3. fMRI Data Acquisition and Preprocessing
fMRI Data Analysis
4.4. Characterization of the BOLD RSNs by Higuchi’s Fractal Dimension
Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maniyar, F.; Sprenger, T.; Monteith, T.; Schankin, C.; Goadsby, P.J. Brain activations in the premonitory phase of nitroglycerin-triggered migraine attacks. Brain 2014, 137, 232–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulte, L.H.; May, A. The migraine generator revisited: Continuous scanning of the migraine cycle over 30 days and three spontaneous attacks. Brain 2016, 139, 1987–1993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulte, L.H.; Mehnert, J.; May, A. Longitudinal Neuroimaging over 30 Days: Temporal Characteristics of Migraine. Ann. Neurol. 2020, 87, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Chong, C.D.; Aguilar, M.; Schwedt, T.J. Altered Hypothalamic Region Covariance in Migraine and Cluster Headache: A Structural MRI Study. Headache 2020, 60, 553–563. [Google Scholar] [CrossRef]
- Messina, R.; Rocca, M.A.; Valsasina, P.; Misci, P.; Filippi, M. Clinical correlates of hypothalamic functional changes in migraine patients. Cephalalgia 2021, 42, 279–290. [Google Scholar] [CrossRef]
- Moulton, E.; Becerra, L.; Johnson, A.; Burstein, R.; Borsook, D. Altered hypothalamic functional connectivity with autonomic circuits and the locus coeruleus in migraine. PLoS ONE 2014, 9, e95508. [Google Scholar] [CrossRef] [Green Version]
- Coppola, G.; Parisi, V.; Di Renzo, A.; Pierelli, F. Cortical pain processing in migraine. J. Neural Transm. 2020, 127, 551–566. [Google Scholar] [CrossRef]
- Coppola, G.; Di Renzo, A.; Petolicchio, B.; Tinelli, E.; Di Lorenzo, C.; Serrao, M.; Calistri, V.; Tardioli, S.; Cartocci, G.; Parisi, V.; et al. Increased neural connectivity between the hypothalamus and cortical resting-state functional networks in chronic migraine. J. Neurol. 2020, 267, 185–191. [Google Scholar] [CrossRef]
- Schulte, L.H.; Allers, A.; May, A. Hypothalamus as a mediator of chronic migraine. Neurology 2017, 88, 2011–2016. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, X.; Liu, M.; Ma, L.; Yu, S. Volume of hypothalamus as a diagnostic biomarker of chronic migraine. Front. Neurol. 2019, 10, 606. [Google Scholar] [CrossRef]
- Suárez, L.E.; Markello, R.D.; Betzel, R.F.; Misic, B. Linking Structure and Function in Macroscale Brain Networks. Trends Cogn. Sci. 2020, 24, 302–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tozzi, A. The multidimensional brain. Phys. Life Rev. 2019, 31, 86–103. [Google Scholar] [CrossRef] [PubMed]
- Barbano, L.; Ziccardi, L.; Parisi, V. Correlations between visual morphological, electrophysiological, and acuity changes in chronic non-arteritic ischemic optic neuropathy. Graefe’s Arch. Clin. Exp. Ophthalmol. 2021, 259, 1297–1308. [Google Scholar] [CrossRef] [PubMed]
- Lisicki, M.; D’Ostilio, K.; Coppola, G.; de Noordhout, A.M.; Parisi, V.; Schoenen, J.; Magis, D. Brain correlates of single trial visual evoked potentials in migraine: More than meets the eye. Front. Neurol. 2018, 9, 393. [Google Scholar] [CrossRef] [PubMed]
- Segall, J.M.; Allen, E.A.; Jung, R.E.; Erhardt, E.B.; Arja, S.K.; Kiehl, K.; Calhoun, V.D. Correspondence between structure and function in the human brain at rest. Front. Neuroinform. 2012, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Tavakol, S.; Li, Q.; Royer, J.; Vos de Wael, R.; Larivière, S.; Lowe, A.; Paquola, C.; Jefferies, E.; Hartley, T.; Bernasconi, A.; et al. A Structure–Function Substrate of Memory for Spatial Configurations in Medial and Lateral Temporal Cortices. Cereb. Cortex 2021, 31, 3213–3225. [Google Scholar] [CrossRef]
- Kalinosky, B.T.; Berrios Barillas, R.; Schmit, B.D. Structurofunctional resting-state networks correlate with motor function in chronic stroke. NeuroImage Clin. 2017, 16, 610–623. [Google Scholar] [CrossRef]
- Porcaro, C.; Di Renzo, A.; Tinelli, E.; Di Lorenzo, G.; Seri, S.; Di Lorenzo, C.; Parisi, V.; Caramia, F.; Fiorelli, M.; Di Piero, V.; et al. Hypothalamic structural integrity and temporal complexity of cortical information processing at rest in migraine without aura patients between attacks. Sci. Rep. 2021, 11, 18701. [Google Scholar] [CrossRef]
- Goldberger, A.L.; Peng, C.K.; Lipsitz, L.A. What is physiologic complexity and how does it change with aging and disease? Neurobiol. Aging 2002, 23, 23–26. [Google Scholar] [CrossRef]
- Menon, V.; Uddin, L.Q. Saliency, switching, attention and control: A network model of insula function. Brain Struct. Funct. 2010, 214, 655–667. [Google Scholar] [CrossRef]
- Stankewitz, A.; May, A. Increased limbic and brainstem activity during migraine attacks following olfactory stimulation. Neurology 2011, 77, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Amin, F.M.; Hougaard, A.; Magon, S.; Asghar, M.S.; Ahmad, N.N.; Rostrup, E.; Sprenger, T.; Ashina, M. Change in brain network connectivity during PACAP38-induced migraine attacks: A resting-state functional MRI study. Neurology 2016, 86, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Coppola, G.; Di Renzo, A.; Tinelli, E.; Di Lorenzo, C.; Scapeccia, M.; Parisi, V.; Serrao, M.; Evangelista, M.; Ambrosini, A.; Colonnese, C.; et al. Resting state connectivity between default mode network and insula encodes acute migraine headache. Cephalalgia 2018, 38, 846–854. [Google Scholar] [CrossRef] [PubMed]
- Denuelle, M.; Fabre, N.; Payoux, P.; Chollet, F.; Geraud, G. Hypothalamic activation in spontaneous migraine attacks. Headache 2007, 47, 1418–1426. [Google Scholar] [CrossRef]
- Schulte, L.H.; Menz, M.M.; Haaker, J.; May, A. The migraineur’s brain networks: Continuous resting state fMRI over 30 days. Cephalalgia 2020, 40, 1614–1621. [Google Scholar] [CrossRef]
- Beaulieu, C. The basis of anisotropic water diffusion in the nervous system—A technical review. NMR Biomed. 2002, 15, 435–455. [Google Scholar] [CrossRef]
- Leone, M.; Bussone, G. Pathophysiology of trigeminal autonomic cephalalgias. Lancet Neurol. 2009, 8, 755–764. [Google Scholar] [CrossRef]
- Meylakh, N.; Marciszewski, K.K.; Di Pietro, F.; Macefield, V.G.; Macey, P.M.; Henderson, L.A. Altered regional cerebral blood flow and hypothalamic connectivity immediately prior to a migraine headache. Cephalalgia 2020, 40, 448–460. [Google Scholar] [CrossRef]
- ICHD Headache Classification Committee of the International Headache Society (IHS) The International Classification of Headache Disorders, 3rd edition. Cephalalgia 2018, 38, 1–211. [CrossRef]
- Baroncini, M.; Jissendi, P.; Balland, E.; Besson, P.; Pruvo, J.-P.; Francke, J.-P.; Dewailly, D.; Blond, S.; Prevot, V. MRI atlas of the human hypothalamus. Neuroimage 2012, 59, 168–180. [Google Scholar] [CrossRef]
- Boes, A.D.; Fischer, D.; Geerling, J.C.; Bruss, J.; Saper, C.B.; Fox, M.D. Connectivity of sleep- and wake-promoting regions of the human hypothalamus observed during resting wakefulness. Sleep 2018, 41, zsy108. [Google Scholar] [CrossRef] [PubMed]
- Power, J.D.; Barnes, K.A.; Snyder, A.Z.; Schlaggar, B.L.; Petersen, S.E. Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. Neuroimage 2012, 59, 2142–2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talairach, J.; Tournoux, P. Co-Planar Stereotaxic Atlas of the Human Brain; Georg Thieme Verlag: New York, NY, USA, 1988; ISBN 3137117011. [Google Scholar]
- Allen, E.A.; Damaraju, E.; Plis, S.M.; Erhardt, E.B.; Eichele, T.; Calhoun, V.D. Tracking whole-brain connectivity dynamics in the resting state. Cereb. Cortex 2014, 24, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Calhoun, V.D.; Adali, T.; Pearlson, G.D.; Pekar, J.J. A method for making group inferences from functional MRI data using independent component analysis. Hum. Brain Mapp. 2001, 14, 140–151. [Google Scholar] [CrossRef]
- Porcaro, C.; Zappasodi, F.; Rossini, P.M.; Tecchio, F. Choice of multivariate autoregressive model order affecting real network functional connectivity estimate. Clin. Neurophysiol. 2009, 120, 436–448. [Google Scholar] [CrossRef]
- Calhoun, V.D.; Adali, T.; Pearlson, G.D.; Pekar, J.J. Spatial and temporal independent component analysis of functional MRI data containing a pair of task-related waveforms. Hum. Brain Mapp. 2001, 13, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Griffanti, L.; Douaud, G.; Bijsterbosch, J.; Evangelisti, S.; Alfaro-Almagro, F.; Glasser, M.F.; Duff, E.P.; Fitzgibbon, S.; Westphal, R.; Carone, D.; et al. Hand classification of fMRI ICA noise components. Neuroimage 2017, 154, 188–205. [Google Scholar] [CrossRef]
- Beckmann, C.; DeLuca, M.; Devlin, J.T.; Smith, S.M. Investigations into resting-state connectivity using independent component analysis. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2005, 360, 1001–1013. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, T. Approach to an irregular time series on the basis of the fractal theory. Phys. D Nonlinear Phenom. 1988, 31, 277–283. [Google Scholar] [CrossRef]
- Porcaro, C.; Di Renzo, A.; Tinelli, E.; Di Lorenzo, G.; Parisi, V.; Caramia, F.; Fiorelli, M.; Di Piero, V.; Pierelli, F.; Coppola, G. Haemodynamic activity characterization of resting state networks by fractal analysis and thalamocortical morphofunctional integrity in chronic migraine. J. Headache Pain 2020, 21, 112. [Google Scholar] [CrossRef]
- Porcaro, C.; Mayhew, S.D.; Marino, M.; Mantini, D.; Bagshaw, A.P. Characterisation of Haemodynamic Activity in Resting State Networks by Fractal Analysis. Int. J. Neural Syst. 2020, 30, 2050061. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy Controls (HC: #20) | Ictal Migraine (MI: #15) | |
|---|---|---|
| Women (number) | 11 | 12 |
| Age (years) | 29.7 ± 3.94 | 34.3 ± 10.26 |
| Attacks frequency/month (number) | 4.5 ± 3.88 | |
| Mean attacks duration (h) | 36.3 ± 30.0 | |
| Disease duration (year) | 16.6 ± 9.33 | |
| Mean severity of headache (VAS) (0–10) | 7.7 ± 0.86 | |
| Mean headache-related disability (number) | 2.4 ± 0.43 | |
| Acute medication intake/month (number) | 4.5 ± 3.32 |
| DTI Metric | HC | MI | Age (F; p) | Sex (F; p) | Group (F; p) | R2—adj (%) |
|---|---|---|---|---|---|---|
| FA | 0.271 ± 0.050 | 0.149 ± 0.035 | 0.72; 0.404 | 0.19; 0.668 | 31.78; <0.001 | 62.11 |
| MD | 1.170E − 03 ± 1.630E − 04 | 2.157E − 03 ± 4.425E − 04 | 1.60; 0.217 | 1.08; 0.307 | 41.82; <0.001 | 69.64 |
| AD | 1.450E − 03 ± 1.530E − 04 | 2.238E − 03 ± 4.395E − 04 | 1.08; 0.307 | 1.48; 0.234 | 30.49; <0.001 | 62.02 |
| RD | 1.030E − 03± 1.690E − 04 | 2.021E − 03± 4.282E − 04 | 1.14; 0.295 | 1.37; 0.253 | 45.92; <0.001 | 73.77 |
| Hypothalamic Region | DTI Metric | HC | MI | Age (F; p) | Sex (F; p) | Group (F; p) | R2—adj (%) |
|---|---|---|---|---|---|---|---|
| Anterior Left | FA | 0.174 ± 0.054 | 0.159 ± 0.070 | 0.01; 0.930 | 0.36; 0.551 | 0.16; 0.694 | 0.00 |
| MD | 1.520E − 03 ± 3.788E − 04 | 1.706E − 03 ± 6.391E − 04 | 1.35; 0.255 | 1.71; 0.202 | 0.17; 0.683 | 4.95 | |
| AD | 1.696E − 03 ± 4.514E − 04 | 1.806E − 03 ± 7.044E − 04 | 1.79; 0.192 | 0.71; 0.406 | 0.04; 0.848 | 0.36 | |
| RD | 1.433E − 03 ± 3.282E − 04 | 1.577E − 03 ± 6.369E − 04 | 2.38; 0.135 | 2.98; 0.096 | 0.03; 0.896 | 12.14 | |
| Posterior Left | FA | 0.280 ± 0.161 | 0.116 ± 0.062 | 0.67; 0.421 | 1.30; 0.265 | 7.44; 0.01 | 28.25 |
| MD | 1.576E − 03 ± 5.042E − 04 | 2.939E − 03 ± 4.739E − 04 | 0.21; 0.652 | 0.44; 0.513 | 35.94; <0.001 | 63.07 | |
| AD | 1.825E − 03 ± 4.961E − 04 | 3.190E − 03 ± 5.345E − 04 | 0.05; 0.822 | 0.16; 0.695 | 34.24; <0.001 | 61.03 | |
| RD | 1.468E − 03 ± 5.208E − 04 | 2.815E − 03 ± 4.070E − 04 | 0.02; 0.890 | 0.73; 0.402 | 41.33; <0.001 | 65.04 | |
| Anterior Right | FA | 0.179 ± 0.065 | 0.190 ± 0.117 | 0.00; 0.946 | 0.03; 0.868 | 0.22; 0.643 | 0.00 |
| MD | 1.361E − 03 ± 2.672E − 04 | 1.834E − 03 ± 8.640E − 04 | 5.24; 0.030 | 0.03; 0.870 | 0.44; 0.514 | 18.84 | |
| AD | 1.514E − 03 ± 2.860E − 04 | 2.046E − 03 ± 9.858E − 04 | 6.90; 0.014 | 0.01; 0.942 | 0.24; 0.625 | 22.41 | |
| RD | 1.281E − 03 ± 2.571E − 04 | 1.984E − 03 ± 8.173E − 04 | 4.62; 0.041 | 0.19; 0.668 | 0.33; 0.573 | 16.07 | |
| Posterior Right | FA | 0.319 ± 0.133 | 0.117 ± 0.058 | 0.48; 0.493 | 0.55; 0.465 | 15.46; 0.001 | 43.32 |
| MD | 1.754E − 03 ± 5.962E − 04 | 2.868E − 03 ± 4.083E − 04 | 0.79; 0.381 | 2.14; 0.156 | 21.82; <0.001 | 53.62 | |
| AD | 2.073E − 03 ± 5.808E − 04 | 3.145E − 03 ± 4.572E − 04 | 0.37; 0.550 | 1.57; 0.221 | 20.95; <0.001 | 50.54 | |
| RD | 1.608E − 03 ± 6.013E − 04 | 2.795E − 03 ± 3.166E − 04 | 0.15; 0.703 | 2.46; 0.129 | 30.48; <0.001 | 58.82 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Porcaro, C.; Di Renzo, A.; Tinelli, E.; Parisi, V.; Di Lorenzo, C.; Caramia, F.; Fiorelli, M.; Giuliani, G.; Cioffi, E.; Seri, S.; et al. A Hypothalamic Mechanism Regulates the Duration of a Migraine Attack: Insights from Microstructural and Temporal Complexity of Cortical Functional Networks Analysis. Int. J. Mol. Sci. 2022, 23, 13238. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113238
Porcaro C, Di Renzo A, Tinelli E, Parisi V, Di Lorenzo C, Caramia F, Fiorelli M, Giuliani G, Cioffi E, Seri S, et al. A Hypothalamic Mechanism Regulates the Duration of a Migraine Attack: Insights from Microstructural and Temporal Complexity of Cortical Functional Networks Analysis. International Journal of Molecular Sciences. 2022; 23(21):13238. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113238
Chicago/Turabian StylePorcaro, Camillo, Antonio Di Renzo, Emanuele Tinelli, Vincenzo Parisi, Cherubino Di Lorenzo, Francesca Caramia, Marco Fiorelli, Giada Giuliani, Ettore Cioffi, Stefano Seri, and et al. 2022. "A Hypothalamic Mechanism Regulates the Duration of a Migraine Attack: Insights from Microstructural and Temporal Complexity of Cortical Functional Networks Analysis" International Journal of Molecular Sciences 23, no. 21: 13238. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113238