
Liver Brain Interactions: Focus on FGF21 a Systematic Review
 , , and
, , and
Abstract
:1. Introduction
1.1. Obesity
1.2. Brain Homeostatic Mechanisms and Energy Balance
1.3. Fibroblast Growth Factor21 and Metabolic Control
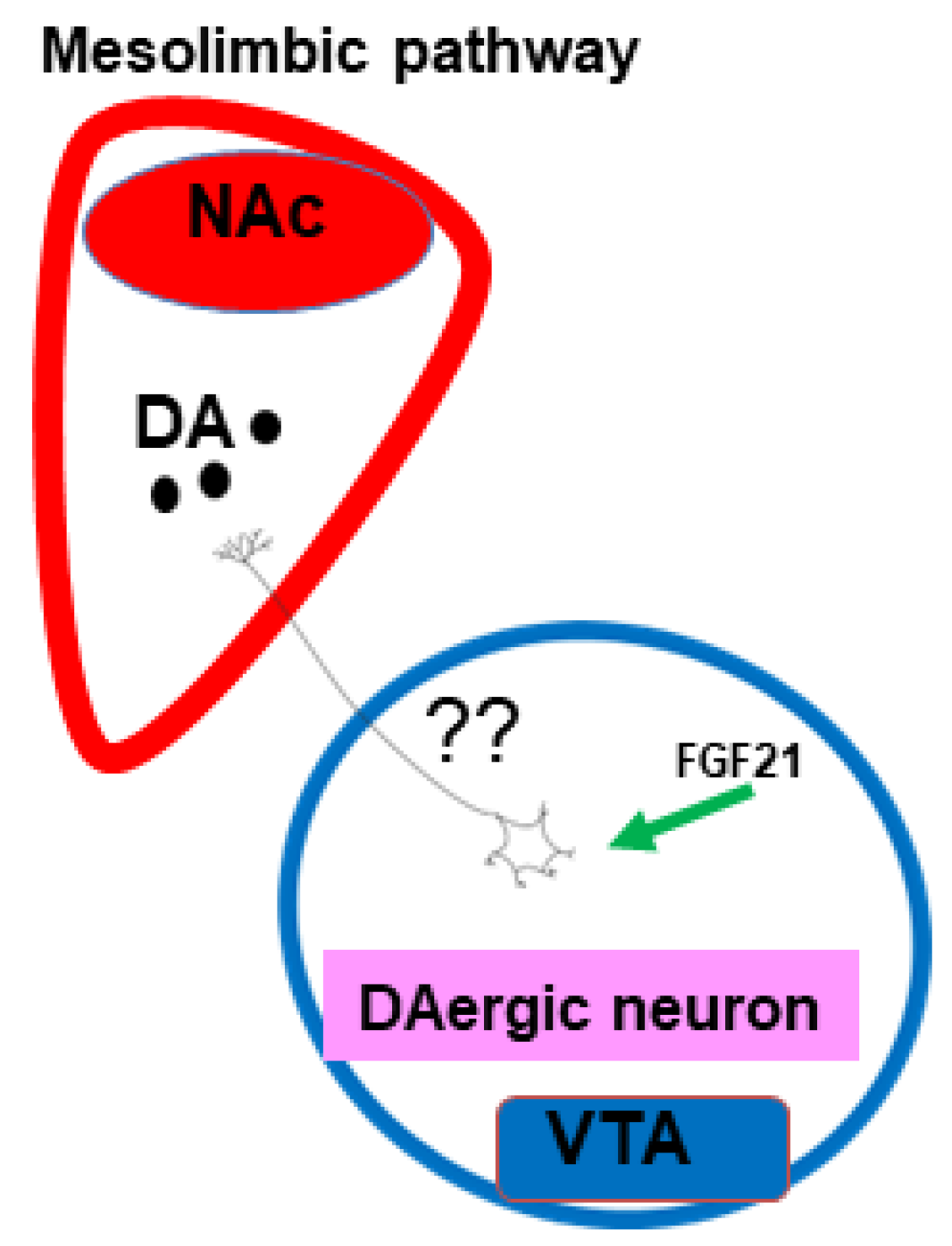
1.4. Central Fibroblast Growth Factor 21 Actions
1.5. Therapeutic Use of FGF21
2. Methods
Study Design, Literature Search and Data Collection
3. Results and Discussion
4. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Farooqi, S.; O’Rahilly, S. Genetics of Obesity in Humans. Endocr. Rev. 2006, 27, 710–718. [Google Scholar] [CrossRef] [PubMed]
- Jéquier, E. Pathways to obesity. Int. J. Obes. 2002, 26, S12–S17. [Google Scholar] [CrossRef] [Green Version]
- NCD Risk Factor Collaboration (NCD-RisC) Rising rural body-mass index is the main driver of the global obesity epidemic in adults. Nature 2019, 569, 260–264. [CrossRef] [PubMed] [Green Version]
- Hill, J.O.; Peters, J.C. Environmental Contributions to the Obesity Epidemic. Science 1998, 280, 1371–1374. [Google Scholar] [CrossRef] [PubMed]
- Berthoud, H.-R.; Morrison, C. The Brain, Appetite, and Obesity. Annu. Rev. Psychol. 2008, 59, 55–92. [Google Scholar] [CrossRef] [PubMed]
- Foster, G.D.; Wadden, T.A.; Kendall, P.C.; Stunkard, A.J.; Al, E. Psychological effects of weight loss and regain: A prospective evaluation. J. Consult. Clin. Psychol. 1996, 64, 752–757. [Google Scholar] [CrossRef]
- Broberger, C. Brain regulation of food intake and appetite: Molecules and networks. J. Intern. Med. 2005, 258, 301–327. [Google Scholar] [CrossRef]
- Jais, A.; Brüning, J.C. Arcuate Nucleus-Dependent Regulation of Metabolism—Pathways to Obesity and Diabetes Mellitus. Endocr. Rev. 2021, 43, 314–328. [Google Scholar] [CrossRef]
- Grundy, S. Metabolic Syndrome: Therapeutic Considerations. Handb. Exp. Pharmacol. 2005, 170, 107–133. [Google Scholar] [CrossRef]
- Huang, P.L. A comprehensive definition for metabolic syndrome. Dis. Model. Mech. 2009, 2, 231–237. [Google Scholar] [CrossRef]
- Angulo, P. Nonalcoholic Fatty Liver Disease. N. Engl. J. Med. 2002, 346, 1221–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quiñones, M.; Fernø, J.; Al-Massadi, O. Ghrelin and liver disease. Rev. Endocr. Metab. Disord. 2019, 21, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Quiñones, M.; Fernø, J.; Diéguez, C.; Nogueiras, R.; Al-Massadi, O. Exciting advances in GPCR-based drugs discovery for treating metabolic disease and future perspectives. Expert Opin. Drug Discov. 2019, 14, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Al Massadi, O.; Nogueiras, R.; Dieguez, C.; Girault, J.-A. Ghrelin and food reward. Neuropharmacology 2019, 148, 131–138. [Google Scholar] [CrossRef]
- Prida, E.; Fernández-González, S.; Pena-León, V.; Pérez-Lois, R.; Fernø, J.; Seoane, L.M.; Quiñones, M.; Al Massadi, O. Crosstalk between Melanin Concentrating Hormone and Endocrine Factors: Implications for Obesity. Int. J. Mol. Sci. 2022, 23, 2436. [Google Scholar] [CrossRef]
- Saper, C.B.; Chou, T.C.; Elmquist, J.K. The Need to Feed: Homeostatic and Hedonic Control of Eating. Neuron 2002, 36, 199–211. [Google Scholar] [CrossRef] [Green Version]
- Schneeberger, M.; Gomis, R.; Claret, M. Hypothalamic and brainstem neuronal circuits controlling homeostatic energy balance. J. Endocrinol. 2014, 220, T25–T46. [Google Scholar] [CrossRef] [Green Version]
- Al-Massadi, O.; Dieguez, C.; Schneeberger, M.; López, M.; Schwaninger, M.; Prevot, V.; Nogueiras, R. Multifaceted actions of melanin-concentrating hormone on mammalian energy homeostasis. Nat. Rev. Endocrinol. 2021, 17, 745–755. [Google Scholar] [CrossRef]
- Al Massadi, O.; López, M.; Tschöp, M.; Diéguez, C.; Nogueiras, R. Current Understanding of the Hypothalamic Ghrelin Pathways Inducing Appetite and Adiposity. Trends Neurosci. 2017, 40, 167–180. [Google Scholar] [CrossRef]
- Timper, K.; Brüning, J.C. Hypothalamic circuits regulating appetite and energy homeostasis: Pathways to obesity. Dis. Model. Mech. 2017, 10, 679–689. [Google Scholar] [CrossRef]
- Friedman, J.M. Modern science versus the stigma of obesity. Nat. Med. 2004, 10, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Myers, M.G.; Olson, D. Central nervous system control of metabolism. Nature 2012, 491, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Valassi, E.; Scacchi, M.; Cavagnini, F. Neuroendocrine control of food intake. Nutr. Metab. Cardiovasc. Dis. 2008, 18, 158–168. [Google Scholar] [CrossRef]
- Quarta, C.; Claret, M.; Zeltser, L.M.; Williams, K.W.; Yeo, G.S.H.; Tschöp, M.H.; Diano, S.; Brüning, J.C.; Cota, D. POMC neuronal heterogeneity in energy balance and beyond: An integrated view. Nat. Metab. 2021, 3, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Yeo, G.S.; Chao, D.H.M.; Siegert, A.-M.; Koerperich, Z.M.; Ericson, M.D.; Simonds, S.E.; Larson, C.M.; Luquet, S.; Clarke, I.; Sharma, S.; et al. The melanocortin pathway and energy homeostasis: From discovery to obesity therapy. Mol. Metab. 2021, 48, 101206. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Nakatake, Y.; Konishi, M.; Itoh, N. Identification of a novel FGF, FGF-21, preferentially expressed in the liver. Biochim. Et Biophys. Acta (BBA)-Gene Struct. Expr. 2000, 1492, 203–206. [Google Scholar] [CrossRef]
- Johnson, C.L.; Weston, J.Y.; Chadi, S.A.; Fazio, E.N.; Huff, M.W.; Kharitonenkov, A.; Köester, A.; Pin, C.L. Fibroblast Growth Factor 21 Reduces the Severity of Cerulein-Induced Pancreatitis in Mice. Gastroenterology 2009, 137, 1795–1804. [Google Scholar] [CrossRef]
- Markan, K.R.; Naber, M.C.; Ameka, M.K.; Anderegg, M.D.; Mangelsdorf, D.J.; Kliewer, S.A.; Mohammadi, M.; Potthoff, M.J. Circulating FGF21 Is Liver Derived and Enhances Glucose Uptake During Refeeding and Overfeeding. Diabetes 2014, 63, 4057–4063. [Google Scholar] [CrossRef] [Green Version]
- Fisher, F.M.; Maratos-Flier, E. Understanding the Physiology of FGF21. Annu. Rev. Physiol. 2016, 78, 223–241. [Google Scholar] [CrossRef] [Green Version]
- Fisher, F.M.; Kleiner, S.; Douris, N.; Fox, E.C.; Mepani, R.J.; Verdeguer, F.; Wu, J.; Kharitonenkov, A.; Flier, J.S.; Maratos-Flier, E.; et al. FGF21 regulates PGC-1α and browning of white adipose tissues in adaptive thermogenesis. Genes Dev. 2012, 26, 271–281. [Google Scholar] [CrossRef]
- Coate, K.C.; Hernandez, G.; Thorne, C.A.; Sun, S.; Le, T.D.; Vale, K.; Kliewer, S.A.; Mangelsdorf, D.J. FGF21 Is an Exocrine Pancreas Secretagogue. Cell Metab. 2017, 25, 472–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inagaki, T.; Dutchak, P.; Zhao, G.; Ding, X.; Gautron, L.; Parameswara, V.; Li, Y.; Goetz, R.; Mohammadi, M.; Esser, V.; et al. Endocrine Regulation of the Fasting Response by PPARα-Mediated Induction of Fibroblast Growth Factor 21. Cell Metab. 2007, 5, 415–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badman, M.K.; Pissios, P.; Kennedy, A.R.; Koukos, G.; Flier, J.S.; Maratos-Flier, E. Hepatic Fibroblast Growth Factor 21 Is Regulated by PPARα and Is a Key Mediator of Hepatic Lipid Metabolism in Ketotic States. Cell Metab. 2007, 5, 426–437. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Liu, Y.B.; Hu, H. Metabolic role of fibroblast growth factor 21 in liver, adipose and nervous system tissues. Biomed. Rep. 2017, 6, 495–502. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.; Boney-Montoya, J.; Owen, B.; Bookout, A.L.; Coate, K.; Mangelsdorf, D.J.; Kliewer, S.A. βKlotho Is Required for Fibroblast Growth Factor 21 Effects on Growth and Metabolism. Cell Metab. 2012, 16, 387–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tacer, K.F.; Bookout, A.L.; Ding, X.; Kurosu, H.; John, G.B.; Wang, L.; Goetz, R.; Mohammadi, M.; Kuro-O, M.; Mangelsdorf, D.J.; et al. Research Resource: Comprehensive Expression Atlas of the Fibroblast Growth Factor System in Adult Mouse. Mol. Endocrinol. 2010, 24, 2050–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bookout, A.L.; De Groot, M.H.M.; Owen, B.; Lee, S.; Gautron, L.; Lawrence, H.L.; Ding, X.; Elmquist, J.K.; Takahashi, J.; Mangelsdorf, D.; et al. FGF21 regulates metabolism and circadian behavior by acting on the nervous system. Nat. Med. 2013, 19, 1147–1152. [Google Scholar] [CrossRef]
- Kurosu, H.; Choi, M.; Ogawa, Y.; Dickson, A.S.; Goetz, R.; Eliseenkova, A.V.; Mohammadi, M.; Rosenblatt, K.P.; Kliewer, S.A.; Kuro-O, M. Tissue-specific Expression of βKlotho and Fibroblast Growth Factor (FGF) Receptor Isoforms Determines Metabolic Activity of FGF19 and FGF21. J. Biol. Chem. 2007, 282, 26687–26695. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, Y.; Kurosu, H.; Yamamoto, M.; Nandi, A.; Rosenblatt, K.P.; Goetz, R.; Eliseenkova, A.V.; Mohammadi, M.; Kuro-O, M. βKlotho is required for metabolic activity of fibroblast growth factor 21. Proc. Natl. Acad. Sci. USA 2007, 104, 7432–7437. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.; Uehara, Y.; Motomura-Matsuzaka, K.; Oki, J.; Koyama, Y.; Kimura, M.; Asada, M.; Komi-Kuramochi, A.; Oka, S.; Toru, T. βKlotho Is Required for Fibroblast Growth Factor (FGF) 21 Signaling through FGF Receptor (FGFR) 1c and FGFR3c. Mol. Endocrinol. 2008, 22, 1006–1014. [Google Scholar] [CrossRef]
- Bono, B.S.; Ly, N.K.K.; Miller, P.A.; Williams-Ikhenoba, J.; Dumiaty, Y.; Chee, M.J. Spatial distribution of beta-klotho mRNA in the mouse hypothalamus, hippocampal region, subiculum, and amygdala. J. Comp. Neurol. 2022, 530, 1634–1657. [Google Scholar] [CrossRef] [PubMed]
- Hultman, K.; Scarlett, J.M.; Baquero, A.F.; Cornea, A.; Zhang, Y.; Salinas, C.B.G.; Brown, J.; Morton, G.J.; Whalen, E.J.; Grove, K.L.; et al. The central fibroblast growth factor receptor/beta klotho system: Comprehensive mapping in Mus musculus and comparisons to nonhuman primate and human samples using an automated in situ hybridization platform. J. Comp. Neurol. 2019, 527, 2069–2085. [Google Scholar] [CrossRef] [PubMed]
- Potthoff, M.J.; Inagaki, T.; Satapati, S.; Ding, X.; He, T.; Goetz, R.; Mohammadi, M.; Finck, B.N.; Mangelsdorf, D.J.; Kliewer, S.A.; et al. FGF21 induces PGC-1α and regulates carbohydrate and fatty acid metabolism during the adaptive starvation response. Proc. Natl. Acad. Sci. USA 2009, 106, 10853–10858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.R.; Kim, S.R.; Cho, W.; Lee, S.-G.; Kim, S.H.; Kim, J.H.; Choi, E.; Kim, J.-H.; Yu, J.-W.; Lee, B.-W.; et al. Short Term Isocaloric Ketogenic Diet Modulates NLRP3 Inflammasome Via B-hydroxybutyrate and Fibroblast Growth Factor 21. Front. Immunol. 2022, 13, 845352. [Google Scholar] [CrossRef] [PubMed]
- Forney, L.A.; Fang, H.; Sims, L.C.; Stone, K.P.; Vincik, L.Y.; Vick, A.M.; Gibson, A.N.; Burk, D.H.; Gettys, T.W. Dietary Methionine Restriction Signals to the Brain Through Fibroblast Growth Factor 21 to Regulate Energy Balance and Remodeling of Adipose Tissue. Obesity 2020, 28, 1912–1921. [Google Scholar] [CrossRef]
- Moro, J.; Chaumontet, C.; Even, P.C.; Blais, A.; Piedcoq, J.; Gaudichon, C.; Tomé, D.; Azzout-Marniche, D. Severe protein deficiency induces hepatic expression and systemic level of FGF21 but inhibits its hypothalamic expression in growing rats. Sci. Rep. 2021, 11, 12436. [Google Scholar] [CrossRef]
- Zhang, X.; Yeung, D.C.Y.; Karpisek, M.; Stejskal, D.; Zhou, Z.-G.; Liu, F.; Wong, R.L.C.; Chow, W.-S.; Tso, A.W.K.; Lam, K.S.L.; et al. Serum FGF21 Levels Are Increased in Obesity and Are Independently Associated with the Metabolic Syndrome in Humans. Diabetes 2008, 57, 1246–1253. [Google Scholar] [CrossRef] [Green Version]
- Mraz, M.; Bartlova, M.; Lacinova, Z.; Michalsky, D.; Kasalicky, M.; Haluzikova, D.; Matoulek, M.; Dostalova, I.; Humenanska, V.; Haluzik, M. Serum concentrations and tissue expression of a novel endocrine regulator fibroblast growth factor-21 in patients with type 2 diabetes and obesity. Clin. Endocrinol. 2009, 71, 369–375. [Google Scholar] [CrossRef]
- Dushay, J.; Chui, P.C.; Gopalakrishnan, G.S.; Varela–Rey, M.; Crawley, M.; Fisher, F.M.; Badman, M.K.; Chantar, M.L.M.; Maratos–Flier, E. Increased Fibroblast Growth Factor 21 in Obesity and Nonalcoholic Fatty Liver Disease. Gastroenterology 2010, 139, 456–463. [Google Scholar] [CrossRef] [Green Version]
- Kiluk, P.; Baran, A.; Kaminski, T.W.; Maciaszek, M.; Flisiak, I. The Level of FGF 21 as a New Risk Factor for the Occurrence of Cardiometabolic Disorders amongst the Psoriatic Patients. J. Clin. Med. 2019, 8, 2206. [Google Scholar] [CrossRef]
- Miehle, K.; Ebert, T.; Kralisch, S.; Hoffmann, A.; Kratzsch, J.; Schlögl, H.; Stumvoll, M.; Fasshauer, M. Serum concentrations of fibroblast growth factor 21 are elevated in patients with congenital or acquired lipodystrophy. Cytokine 2016, 83, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Lindegaard, B.; Hvid, T.; Grøndahl, T.; Frosig, C.; Gerstoft, J.; Hojman, P.; Pedersen, B.K. Expression of Fibroblast Growth Factor-21 in Muscle Is Associated with Lipodystrophy, Insulin Resistance and Lipid Disturbances in Patients with HIV. PLoS ONE 2013, 8, e55632. [Google Scholar] [CrossRef]
- Rodríguez-Nuevo, A.; Díaz-Ramos, A.; Noguera, E.; Díaz-Sáez, F.; Duran, X.; Muñoz, J.P.; Romero, M.; Plana, N.; Sebastián, D.; Tezze, C.; et al. Mitochondrial DNA and TLR9 drive muscle inflammation upon Opa1 deficiency. EMBO J. 2018, 37, e96553. [Google Scholar] [CrossRef]
- Coskun, T.; Bina, H.A.; Schneider, M.A.; Dunbar, J.D.; Hu, C.C.; Chen, Y.; Moller, D.E.; Kharitonenkov, A. Fibroblast Growth Factor 21 Corrects Obesity in Mice. Endocrinology 2008, 149, 6018–6027. [Google Scholar] [CrossRef]
- Kharitonenkov, A.; Shiyanova, T.L.; Koester, A.; Ford, A.M.; Micanovic, R.; Galbreath, E.J.; Sandusky, G.E.; Hammond, L.J.; Moyers, J.S.; Owens, R.A.; et al. FGF-21 as a novel metabolic regulator. J. Clin. Investig. 2005, 115, 1627–1635. [Google Scholar] [CrossRef] [Green Version]
- Tomlinson, E.; Fu, L.; John, L.; Hultgren, B.; Huang, X.; Renz, M.; Stephan, J.P.; Tsai, S.P.; Powell-Braxton, L.; French, D.; et al. Transgenic Mice Expressing Human Fibroblast Growth Factor-19 Display Increased Metabolic Rate and Decreased Adiposity. Endocrinology 2002, 143, 1741–1747. [Google Scholar] [CrossRef]
- Xu, J.; Stanislaus, S.; Chinookoswong, N.; Lau, Y.Y.; Hager, T.; Patel, J.; Ge, H.; Weiszmann, J.; Lu, S.-C.; Graham, M.; et al. Acute glucose-lowering and insulin-sensitizing action of FGF21 in insulin-resistant mouse models—Association with liver and adipose tissue effects. Am. J. Physiol. Metab. 2009, 297, E1105–E1114. [Google Scholar] [CrossRef] [Green Version]
- Kharitonenkov, A.; Wroblewski, V.J.; Koester, A.; Chen, Y.-F.; Clutinger, C.K.; Tigno, X.T.; Hansen, B.C.; Shanafelt, A.B.; Etgen, G.J. The Metabolic State of Diabetic Monkeys Is Regulated by Fibroblast Growth Factor-21. Endocrinology 2007, 148, 774–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, A.C.; Halstead, C.A.; Hansen, B.C.; Irizarry, A.R.; Martin, J.A.; Myers, S.R.; Reynolds, V.L.; Smith, H.W.; Wroblewski, V.J.; Kharitonenkov, A. LY2405319, an Engineered FGF21 Variant, Improves the Metabolic Status of Diabetic Monkeys. PLoS ONE 2013, 8, e65763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, B.; Straarup, E.M.; Heppner, K.M.; Takahashi, D.L.; Raffaele, V.; Dissen, G.A.; Lewandowski, K.; Bödvarsdottir, T.B.; Raun, K.; Grove, K.L.; et al. FGF21 decreases body weight without reducing food intake or bone mineral density in high-fat fed obese rhesus macaque monkeys. Int. J. Obes. 2018, 42, 1151–1160. [Google Scholar] [CrossRef]
- Dutchak, P.A.; Katafuchi, T.; Bookout, A.L.; Choi, J.H.; Yu, R.T.; Mangelsdorf, D.J.; Kliewer, S.A. Fibroblast Growth Factor-21 Regulates PPARγ Activity and the Antidiabetic Actions of Thiazolidinediones. Cell 2012, 148, 556–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Gao, Z.; Zhang, J.; Ye, X.; Xu, A.; Ye, J.; Jia, W. Sodium Butyrate Stimulates Expression of Fibroblast Growth Factor 21 in Liver by Inhibition of Histone Deacetylase 3. Diabetes 2012, 61, 797–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veniant, M.M.; Hale, C.; Helmering, J.; Chen, M.M.; Stanislaus, S.; Busby, J.; Vonderfecht, S.; Xu, J.; Lloyd, D.J. FGF21 Promotes Metabolic Homeostasis via White Adipose and Leptin in Mice. PLoS ONE 2012, 7, e40164. [Google Scholar] [CrossRef]
- Adams, A.C.; Coskun, T.; Cheng, C.C.; O′farrell, L.S.; DuBois, S.L.; Kharitonenkov, A. Fibroblast growth factor 21 is not required for the antidiabetic actions of the thiazoladinediones. Mol. Metab. 2013, 2, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Tian, H.; Lam, K.S.; Lin, S.; Hoo, R.C.; Konishi, M.; Itoh, N.; Wang, Y.; Bornstein, S.R.; Xu, A.; et al. Adiponectin Mediates the Metabolic Effects of FGF21 on Glucose Homeostasis and Insulin Sensitivity in Mice. Cell Metab. 2013, 17, 779–789. [Google Scholar] [CrossRef] [Green Version]
- Holland, W.L.; Adams, A.C.; Brozinick, J.T.; Bui, H.H.; Miyauchi, Y.; Kusminski, C.M.; Bauer, S.M.; Wade, M.; Singhal, E.; Cheng, C.C.; et al. An FGF21-Adiponectin-Ceramide Axis Controls Energy Expenditure and Insulin Action in Mice. Cell Metab. 2013, 17, 790–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hondares, E.; Rosell, M.; Gonzalez, F.J.; Giralt, M.; Iglesias, R.; Villarroya, F. Hepatic FGF21 Expression Is Induced at Birth via PPARα in Response to Milk Intake and Contributes to Thermogenic Activation of Neonatal Brown Fat. Cell Metab. 2010, 11, 206–212. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Hoo, R.L.-C.; Konishi, M.; Itoh, N.; Lee, P.-C.; Ye, H.-Y.; Lam, K.S.-L.; Xu, A. Growth Hormone Induces Hepatic Production of Fibroblast Growth Factor 21 through a Mechanism Dependent on Lipolysis in Adipocytes. J. Biol. Chem. 2011, 286, 34559–34566. [Google Scholar] [CrossRef] [Green Version]
- Hondares, E.; Iglesias, R.; Giralt, A.; Gonzalez, F.J.; Giralt, M.; Mampel, T.; Villarroya, F. Thermogenic Activation Induces FGF21 Expression and Release in Brown Adipose Tissue. J. Biol. Chem. 2011, 286, 12983–12990. [Google Scholar] [CrossRef] [Green Version]
- Potthoff, M.J.; Kliewer, S.A.; Mangelsdorf, D.J. Endocrine fibroblast growth factors 15/19 and 21: From feast to famine. Genes Dev. 2012, 26, 312–324. [Google Scholar] [CrossRef]
- Douris, N.; Stevanovic, D.M.; Fisher, F.M.; Cisu, T.I.; Chee, M.; Nguyen, N.L.; Zarebidaki, E.; Adams, A.C.; Kharitonenkov, A.; Flier, J.S.; et al. Central Fibroblast Growth Factor 21 Browns White Fat via Sympathetic Action in Male Mice. Endocrinology 2015, 156, 2470–2481. [Google Scholar] [CrossRef] [PubMed]
- Owen, B.M.; Ding, X.; Morgan, D.A.; Coate, K.C.; Bookout, A.L.; Rahmouni, K.; Kliewer, S.A.; Mangelsdorf, D.J. FGF21 Acts Centrally to Induce Sympathetic Nerve Activity, Energy Expenditure, and Weight Loss. Cell Metab. 2014, 20, 670–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, T.; Morgan, D.A.; Rahmouni, K.; Sonoda, J.; Fu, X.; Burgess, S.C.; Holland, W.L.; Kliewer, S.A.; Mangelsdorf, D.J. FGF19, FGF21, and an FGFR1/β-Klotho-Activating Antibody Act on the Nervous System to Regulate Body Weight and Glycemia. Cell Metab. 2017, 26, 709–718.e3. [Google Scholar] [CrossRef]
- Marcelin, G.; Jo, Y.-H.; Li, X.; Schwartz, G.J.; Zhang, Y.; Dun, N.J.; Lyu, R.-M.; Blouet, C.; Chang, J.K.; Chua, S., Jr. Central action of FGF19 reduces hypothalamic AGRP/NPY neuron activity and improves glucose metabolism. Mol. Metab. 2014, 3, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Mutsnaini, L.; Kim, C.-S.; Kim, J.; Joe, Y.; Chung, H.T.; Choi, H.-S.; Roh, E.; Kim, M.-S.; Yu, R. Fibroblast growth factor 21 deficiency aggravates obesity-induced hypothalamic inflammation and impairs thermogenic response. Agents Actions 2019, 68, 351–358. [Google Scholar] [CrossRef]
- Liang, Q.; Zhong, L.; Zhang, J.; Wang, Y.; Bornstein, S.R.; Triggle, C.R.; Ding, H.; Lam, K.S.; Xu, A. FGF21 Maintains Glucose Homeostasis by Mediating the Cross Talk Between Liver and Brain During Prolonged Fasting. Diabetes 2014, 63, 4064–4075. [Google Scholar] [CrossRef] [Green Version]
- Inagaki, T.; Lin, V.Y.; Goetz, R.; Mohammadi, M.; Mangelsdorf, D.; Kliewer, S.A. Inhibition of Growth Hormone Signaling by the Fasting-Induced Hormone FGF21. Cell Metab. 2008, 8, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.M.; Laeger, T.; Dehner, M.; Albarado, D.C.; Clarke, B.; Wanders, D.; Burke, S.J.; Collier, J.J.; Qualls-Creekmore, E.; Solon-Biet, S.M.; et al. FGF21 Signals Protein Status to the Brain and Adaptively Regulates Food Choice and Metabolism. Cell Rep. 2019, 27, 2934–2947.e3. [Google Scholar] [CrossRef] [Green Version]
- Blanco, A.M.; Bertucci, J.I.; Unniappan, S. FGF21 Mimics a Fasting-Induced Metabolic State and Increases Appetite in Zebrafish. Sci. Rep. 2020, 10, 6993. [Google Scholar] [CrossRef] [Green Version]
- Song, P.; Zechner, C.; Hernandez, G.; Cánovas, J.; Xie, Y.; Sondhi, V.; Wagner, M.; Stadlbauer, V.; Horvath, A.; Leber, B.; et al. The Hormone FGF21 Stimulates Water Drinking in Response to Ketogenic Diet and Alcohol. Cell Metab. 2018, 27, 1338–1347.e4. [Google Scholar] [CrossRef]
- Hsuchou, H.; Pan, W.; Kastin, A.J. The fasting polypeptide FGF21 can enter brain from blood. Peptides 2007, 28, 2382–2386. [Google Scholar] [CrossRef] [Green Version]
- von Holstein-Rathlou, S.; BonDurant, L.D.; Peltekian, L.; Naber, M.C.; Yin, T.C.; Claflin, K.E.; Urizar, A.I.; Madsen, A.N.; Ratner, C.; Holst, B.; et al. FGF21 Mediates Endocrine Control of Simple Sugar Intake and Sweet Taste Preference by the Liver. Cell Metab. 2015, 23, 335–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larson, K.R.; Chaffin, A.T.-B.; Goodson, M.L.; Fang, Y.; Ryan, K.K. Fibroblast Growth Factor-21 Controls Dietary Protein Intake in Male Mice. Endocrinology 2019, 160, 1069–1080. [Google Scholar] [CrossRef] [PubMed]
- Talukdar, S.; Owen, B.M.; Song, P.; Hernandez, G.; Zhang, Y.; Zhou, Y.; Scott, W.T.; Paratala, B.; Turner, T.; Smith, A.; et al. FGF21 Regulates Sweet and Alcohol Preference. Cell Metab. 2015, 23, 344–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen-Cody, S.O.; Flippo, K.H.; Claflin, K.E.; Yavuz, Y.; Sapouckey, S.A.; Walters, G.C.; Usachev, Y.M.; Atasoy, D.; Gillum, M.P.; Potthoff, M.J. FGF21 Signals to Glutamatergic Neurons in the Ventromedial Hypothalamus to Suppress Carbohydrate Intake. Cell Metab. 2020, 32, 273–286.e6. [Google Scholar] [CrossRef] [PubMed]
- Flippo, K.H.; Jensen-Cody, S.O.; Claflin, K.E.; Potthoff, M.J. FGF21 signaling in glutamatergic neurons is required for weight loss associated with dietary protein dilution. Sci. Rep. 2020, 10, 19521. [Google Scholar] [CrossRef] [PubMed]
- Recinella, L.; Leone, S.; Ferrante, C.; Chiavaroli, A.; Di Nisio, C.; Martinotti, S.; Vacca, M.; Brunetti, L.; Orlando, G. Effects of central fibroblast growth factor 21 (FGF21) in energy balance. J. Biol. Regul. Homeost. Agents 2017, 31, 603–613. [Google Scholar]
- Špolcová, A.; Holubová, M.; Mikulášková, B.; Nagelová, V.; Štofková, A.; Lacinová, Z.; Jurčovičová, J.; Haluzik, M.; Maletínská, L.; Železná, B. Changes in FGF21 Serum Concentrations and Liver mRNA Expression in an Experimental Model of Complete Lipodystrophy and Insulin-Resistant Diabetes. Physiol. Res. 2014, 63, 483–490. [Google Scholar] [CrossRef]
- Geller, S.; Arribat, Y.; Netzahualcoyotzi, C.; Lagarrigue, S.; Carneiro, L.; Zhang, L.; Amati, F.; Lopez-Mejia, I.C.; Pellerin, L. Tanycytes Regulate Lipid Homeostasis by Sensing Free Fatty Acids and Signaling to Key Hypothalamic Neuronal Populations via FGF21 Secretion. Cell Metab. 2019, 30, 833–844.e7. [Google Scholar] [CrossRef]
- Pena-Leon, V.; Folgueira, C.; Barja-Fernández, S.; Pérez-Lois, R.; Lima, N.D.S.; Martin, M.; Heras, V.; Martinez-Martinez, S.; Valero, P.; Iglesias, C.; et al. Prolonged breastfeeding protects from obesity by hypothalamic action of hepatic FGF21. Nat. Metab. 2022, 4, 901–917. [Google Scholar] [CrossRef]
- Zhou, B.; Claflin, K.E.; Flippo, K.H.; Sullivan, A.I.; Asghari, A.; Tadinada, S.M.; Jensen-Cody, S.O.; Abel, T.; Potthoff, M.J. Central FGF21 production regulates memory but not peripheral metabolism. Cell Rep. 2022, 40, 111239. [Google Scholar] [CrossRef] [PubMed]
- Alves, J.M.; Yunker, A.G.; Luo, S.; Jann, K.; Angelo, B.; DeFendis, A.; Pickering, T.A.; Smith, A.; Monterosso, J.R.; Page, K.A. FGF21 response to sucrose is associated with BMI and dorsal striatal signaling in humans. Obesity 2022, 30, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Gimeno, R.E.; Moller, D.E. FGF21-based pharmacotherapy—Potential utility for metabolic disorders. Trends Endocrinol. Metab. 2014, 25, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Gaich, G.; Chien, J.Y.; Fu, H.; Glass, L.C.; Deeg, M.A.; Holland, W.L.; Kharitonenkov, A.; Bumol, T.; Schilske, H.K.; Moller, D.E. The Effects of LY2405319, an FGF21 Analog, in Obese Human Subjects with Type 2 Diabetes. Cell Metab. 2013, 18, 333–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rühlmann, C.; Dannehl, D.; Brodtrück, M.; Adams, A.C.; Stenzel, J.; Lindner, T.; Krause, B.J.; Vollmar, B.; Kuhla, A. Neuroprotective Effects of the FGF21 Analogue LY2405319. J. Alzheimer’s Dis. 2021, 80, 357–369. [Google Scholar] [CrossRef]
- Kim, A.; Somayaji, V.R.; Dong, J.Q.; Rolph, T.P.; Weng, Y.; Chabot, J.R.; Gropp, K.E.; Talukdar, S.; Calle, R.A. Once-weekly administration of a long-acting fibroblast growth factor 21 analogue modulates lipids, bone turnover markers, blood pressure and body weight differently in obese people with hypertriglyceridaemia and in non-human primates. Diabetes Obes. Metab. 2017, 19, 1762–1772. [Google Scholar] [CrossRef]
- Charles, E.D.; Neuschwander-Tetri, B.A.; Frias, J.P.; Kundu, S.; Luo, Y.; Tirucherai, G.S.; Christian, R. Pegbelfermin (BMS-986036), PEGylated FGF21, in Patients with Obesity and Type 2 Diabetes: Results from a Randomized Phase 2 Study. Obesity 2018, 27, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Sanyal, A.; Charles, E.D.; A Neuschwander-Tetri, B.; Loomba, R.; A Harrison, S.; Abdelmalek, M.F.; Lawitz, E.J.; Halegoua-DeMarzio, D.; Kundu, S.; Noviello, S.; et al. Pegbelfermin (BMS-986036), a PEGylated fibroblast growth factor 21 analogue, in patients with non-alcoholic steatohepatitis: A randomised, double-blind, placebo-controlled, phase 2a trial. Lancet 2018, 392, 2705–2717. [Google Scholar] [CrossRef]
- Baruch, A.; Wong, C.; Chinn, L.W.; Vaze, A.; Sonoda, J.; Gelzleichter, T.; Chen, S.; Lewin-Koh, N.; Morrow, L.; Dheerendra, S.; et al. Antibody-mediated activation of the FGFR1/Klothoβ complex corrects metabolic dysfunction and alters food preference in obese humans. Proc. Natl. Acad. Sci. USA 2020, 117, 28992–29000. [Google Scholar] [CrossRef]
- Harrison, S.A.; Rinella, M.E.; Abdelmalek, M.F.; Trotter, J.F.; Paredes, A.H.; Arnold, H.L.; Kugelmas, M.; Bashir, M.R.; Jaros, M.J.; Ling, L.; et al. NGM282 for treatment of non-alcoholic steatohepatitis: A multicentre, randomised, double-blind, placebo-controlled, phase 2 trial. Lancet 2018, 391, 1174–1185. [Google Scholar] [CrossRef]
- Harrison, S.A.; Rossi, S.J.; Paredes, A.H.; Trotter, J.F.; Bashir, M.R.; Guy, C.D.; Banerjee, R.; Jaros, M.J.; Owers, S.; Baxter, B.A.; et al. NGM282 Improves Liver Fibrosis and Histology in 12 Weeks in Patients with Nonalcoholic Steatohepatitis. Hepatology 2019, 71, 1198–1212. [Google Scholar] [CrossRef]
- Tabibian, J.H.; Lindor, K.D. NGM282, an FGF19 analogue, in primary sclerosing cholangitis: A nebulous matter. J. Hepatol. 2019, 70, 348–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, S.A.; Neff, G.; Guy, C.D.; Bashir, M.R.; Paredes, A.H.; Frias, J.P.; Younes, Z.; Trotter, J.F.; Gunn, N.T.; Moussa, S.E.; et al. Efficacy and Safety of Aldafermin, an Engineered FGF19 Analog, in a Randomized, Double-Blind, Placebo-Controlled Trial of Patients with Nonalcoholic Steatohepatitis. Gastroenterology 2021, 160, 219–231.e1. [Google Scholar] [CrossRef]
- Gilroy, C.A.; Capozzi, M.E.; Varanko, A.K.; Tong, J.; D’Alessio, D.A.; Campbell, J.E.; Chilkoti, A. Sustained release of a GLP-1 and FGF21 dual agonist from an injectable depot protects mice from obesity and hyperglycemia. Sci. Adv. 2020, 6, eaaz9890. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Lin, S.; Li, Y.; Liu, L.; Li, X.; Gao, X.; Yan, J.; Gu, B.; Chen, X.; Li, W.; et al. A novel GLP-1 and FGF21 dual agonist has therapeutic potential for diabetes and non-alcoholic steatohepatitis. eBioMedicine 2021, 63, 103202. [Google Scholar] [CrossRef] [PubMed]
- Tschöp, M.H.; Finan, B.; Clemmensen, C.; Gelfanov, V.; Perez-Tilve, D.; Müller, T.D.; DiMarchi, R.D. Unimolecular Polypharmacy for Treatment of Diabetes and Obesity. Cell Metab. 2016, 24, 51–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phase 3 Global Registration Trial. Available online: https://www.prnewswire.com/news-releases/lillys-tirzepatide-delivered-up-to-22-5-weight-loss-in-adults-with-obesity-or-overweight-in-surmount-1--301534871.html (accessed on 29 September 2022).
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. Declaración PRISMA 2020: Una guía actualizada para la publicación de revisiones sistemáticas. Rev. Española Cardiol. 2021, 74, 790–799. [Google Scholar] [CrossRef]
- Staiger, H.; Keuper, M.; Berti, L.; de Angelis, M.H.; Häring, H.-U. Fibroblast Growth Factor 21—Metabolic Role in Mice and Men. Endocr. Rev. 2017, 38, 468–488. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Specie | Administration | Compound | Effect |
|---|---|---|---|---|
| 2020; Gilroy [104] | Wild type mice | Peripheral | GLP-1 and FGF21 dual agonist | Potently reduces BW Decrease fasting glucose Improves NASH |
| 2021; Pan [105] | Ob/ob mice Db/db mice | Peripheral SC | GLP-1 and FGF21 dual agonist | Potently reduces BW Improves glucose Improve NAFLD |
| 2008; Coskun [54] | Ob/ob and DIO mice | Peripheral IV | recombinant human FGF21 | Potently reduces BW Increases EE Improves glucose Improves NAFLD |
| 2005; Kharitonenkov [55] | Ob/ob and db/db mice | Peripheral SC | human FGF21 | Reduces plasma glucose and TG |
| 2009; Xu [57] | Ob/ob and DIO mice | Peripheral IP | recombinant human FGF21 | Improves glucose tolerance and insulin sensitivity |
| 2007; Kharitonenkov [58] | Diabetic non human primates | Peripheral IV SC | recombinant human FGF21 | Mild reduction of BW Reduces plasma glucose and insulin and improves lipid profile |
| 2013; Adams [59] | DIO Mice | Peripheral SC | FGF21 analogue, LY2405319 | Reduction of BW Reduces plasma glucose and insulin and improves lipid profile |
| 2013; Holland [66] | Ob/ob and DIO mice | Peripheral SC | recombinant murine FGF21 | Reduces plasma glucose and improves insulin sensitivity Increases adiponectin and EE |
| 2015; Douris [71] | WT mice | ICV | recombinant murine FGF21 | Energy expenditure, thermogenesis, and “browning” |
| 2014; Owen [72] | WT Mice | ICV | recombinant murine FGF21 | Increases thermogenesis |
| 2017; Lan [73] | DIO mice | Peripheral IP | FGF21 mimetic antibody | Reduces BW, plasma glucose and insulin, and improves lipid profile |
| 2022: Pena-Leon, [90] | Rats | ICV | recombinant human FGF21 | Reduces BW Increases thermogenesis |
| 2013; Gaich [94] | Obese and diabetic Humans | Peripheral SC | FGF21 analog LY2405319 | Reduces BW and plasma insulin and improves lipid profile |
| 2017; Kim [96] | Obese humans | Peripheral IV | PF-05231023 | Not changes in BW Potently reduces TG |
| 2019; Charles [97] | Obese and diabetic humans | Peripheral SC | Pegbelfermin (BMS-986036), PEGylated FGF21 | Non changes in BW Decrease fasting glucose and insulin sensitivity Improves lipid profile Improve fibrosis markers |
| 2019; Sanyal [98] | overweight or obese with NASH | Peripheral SC | Pegbelfermin (BMS-986036), PEGylated FGF21 | Non changes in BW Improves NASH |
| 2020; Baruch [99] | Non human primates | Peripheral IV | bispecific anti-FGFR1/KLB agonist antibody BFKB8488A | Reduces BW Reduction in preference for sweet taste and carbohydrate intake |
| 2020; Baruch [99] | overweight or obese | Peripheral SC | bispecific anti-FGFR1/KLB agonist antibody BFKB8488A | Transient reduction in BW Improve lipid profile Decrease fasting insulin |
| 2018;Harrison [100] | NASH patients | Peripheral S.C. | FGF19 analogue, NGM282 | Reduces BW Improves NASH |
| 2020;Harrison [101] | NASH patients | Peripheral S.C. | FGF19 analogue, NGM282 | Improves NASH |
| 2021;Harrison [103] | NASH patients | Peripheral S.C. | FGF19 analogue NGM282 | No changes in BW Reduced liver fat |
| 2015; Talukdar [84] | DIO mice Monkeys | Peripheral S.C. I.V. | recombinant human FGF21 PF-05231023 | Supresses sweet preferences |
| 2016; Von Holstein-Rathlou [82] | Mice | Peripheral IP and SC | recombinant human FGF21 | Suppresses sugar intake and sweet taste preferences |
| Study | Model | Data | Compound |
|---|---|---|---|
| 2013; Gaich [94] | Mixed effect linear model | BW: LS mean change from baseline = −1.75 (0.65) p < 0.05 | LY2405319 |
| 2017; Kim [96] | Mixed-effects model for repeated measurements | TG: Placebo-adjusted least squares mean 90% CI −62.1, −24.6 | PF-05231023 |
| 2019; Charles [97] | Longitudinal repeated- measures analysis model | Glucose: LS mean Estimates SE, and two-sided 90% confidence intervals (90% CI −3.46 to 0.22) Fibrosis markers: % of change compared with baseline ALT—20%; AST—8% | BMS-986036 |
| 2020; Baruch [99] | Descriptive statistics. | Mean and SD % change for baseline: BW: 2.30 (0.25) Mean % change for baseline: Cardiometabolic parameters: TG: −66% HDL: +34% LDL: −37% Adiponectin: +250% | BFKB8488A |
| 2019; Sanyal [98] | Longitudinal repeated measures analysis and unstructured covariance matrix were used to represent the correlation of the repeated measures within each patient | Hepatic fat: Adjusted mean absolute change compared with placebo Mean and SE, and two-sided 90% confidence intervals (−6.8% vs. −1.3%; p = 0.0004) | BMS-986036 |
| 2018; Harrison [100] | ANCOVA | BW: LS SD, SE (95% CI), −2.0 (0.9; 3.7 to −0.3) p = 0.023 Hepatic fat: LS means With SE, 95% (−11.1 1.4, −13.9 to −8.3; p < 0.0001) | NGM282 (FGF19 analogue) |
| 2020; Harrison [101] | Sensitivity analysis Wilcoxon matched pairs signed rank test | Fibrosis scores: change compared with baseline (0.5; 0.9 to −0) p = 0.035 | NGM282 (FGF19 analogue) |
| 2021; Harrison [103] | ANCOVA | Least squares (LS) Means with standard errors (SE), 95% (difference, −5.0%; 95% CI, −8.0% to −1.9%; p = 0.002) | FGF21 analogue aldafermin |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prida, E.; Álvarez-Delgado, S.; Pérez-Lois, R.; Soto-Tielas, M.; Estany-Gestal, A.; Fernø, J.; Seoane, L.M.; Quiñones, M.; Al-Massadi, O. Liver Brain Interactions: Focus on FGF21 a Systematic Review. Int. J. Mol. Sci. 2022, 23, 13318. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113318
Prida E, Álvarez-Delgado S, Pérez-Lois R, Soto-Tielas M, Estany-Gestal A, Fernø J, Seoane LM, Quiñones M, Al-Massadi O. Liver Brain Interactions: Focus on FGF21 a Systematic Review. International Journal of Molecular Sciences. 2022; 23(21):13318. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113318
Chicago/Turabian StylePrida, Eva, Sara Álvarez-Delgado, Raquel Pérez-Lois, Mateo Soto-Tielas, Ana Estany-Gestal, Johan Fernø, Luisa María Seoane, Mar Quiñones, and Omar Al-Massadi. 2022. "Liver Brain Interactions: Focus on FGF21 a Systematic Review" International Journal of Molecular Sciences 23, no. 21: 13318. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113318