

Effects of Elevated Aluminum Concentration and Distribution on Root Damage, Cell Wall Polysaccharides, and Nutrient Uptake in Different Tolerant Eucalyptus Clones

 , ,
, ,
Abstract
:1. Introduction
2. Results
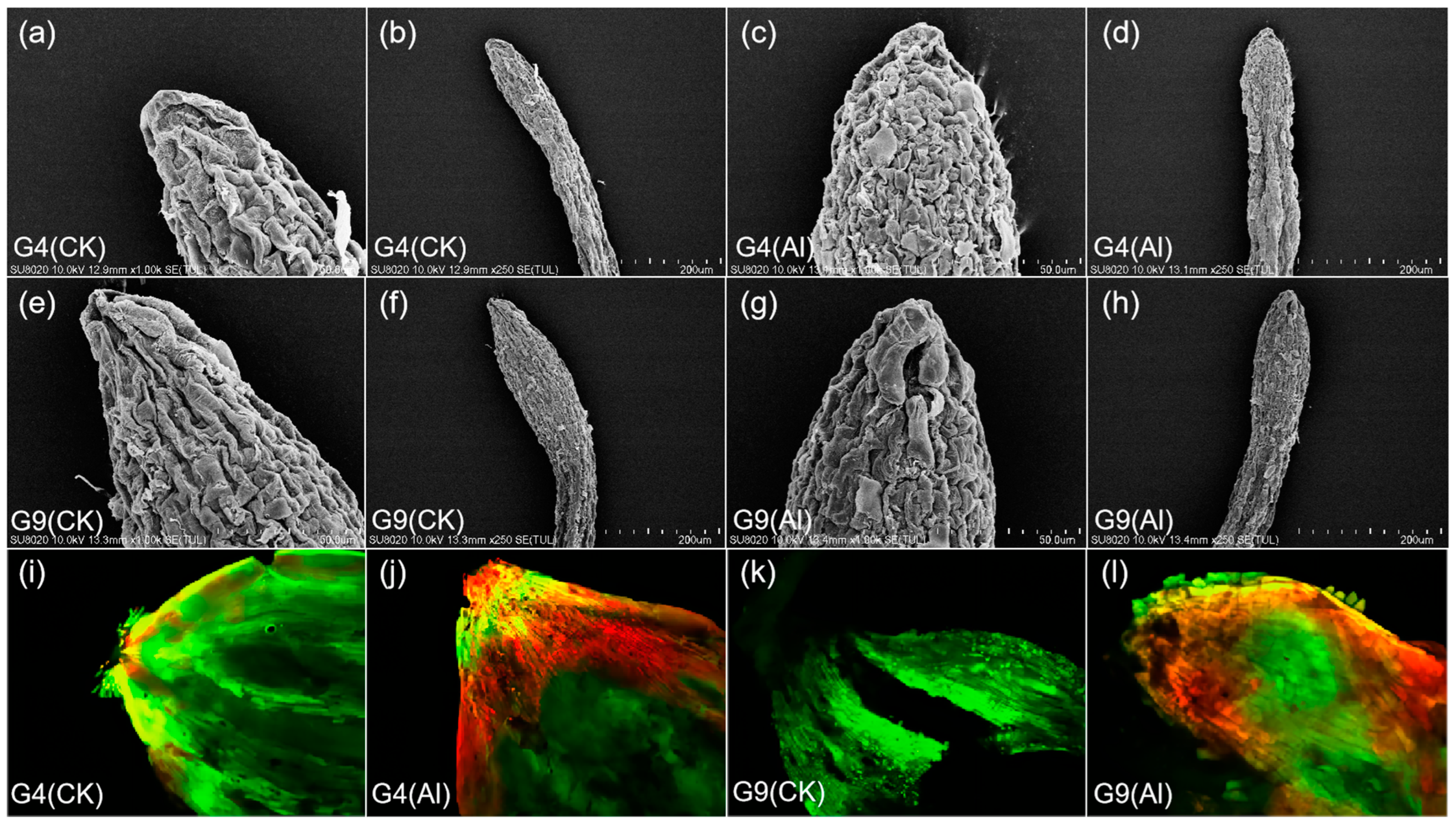
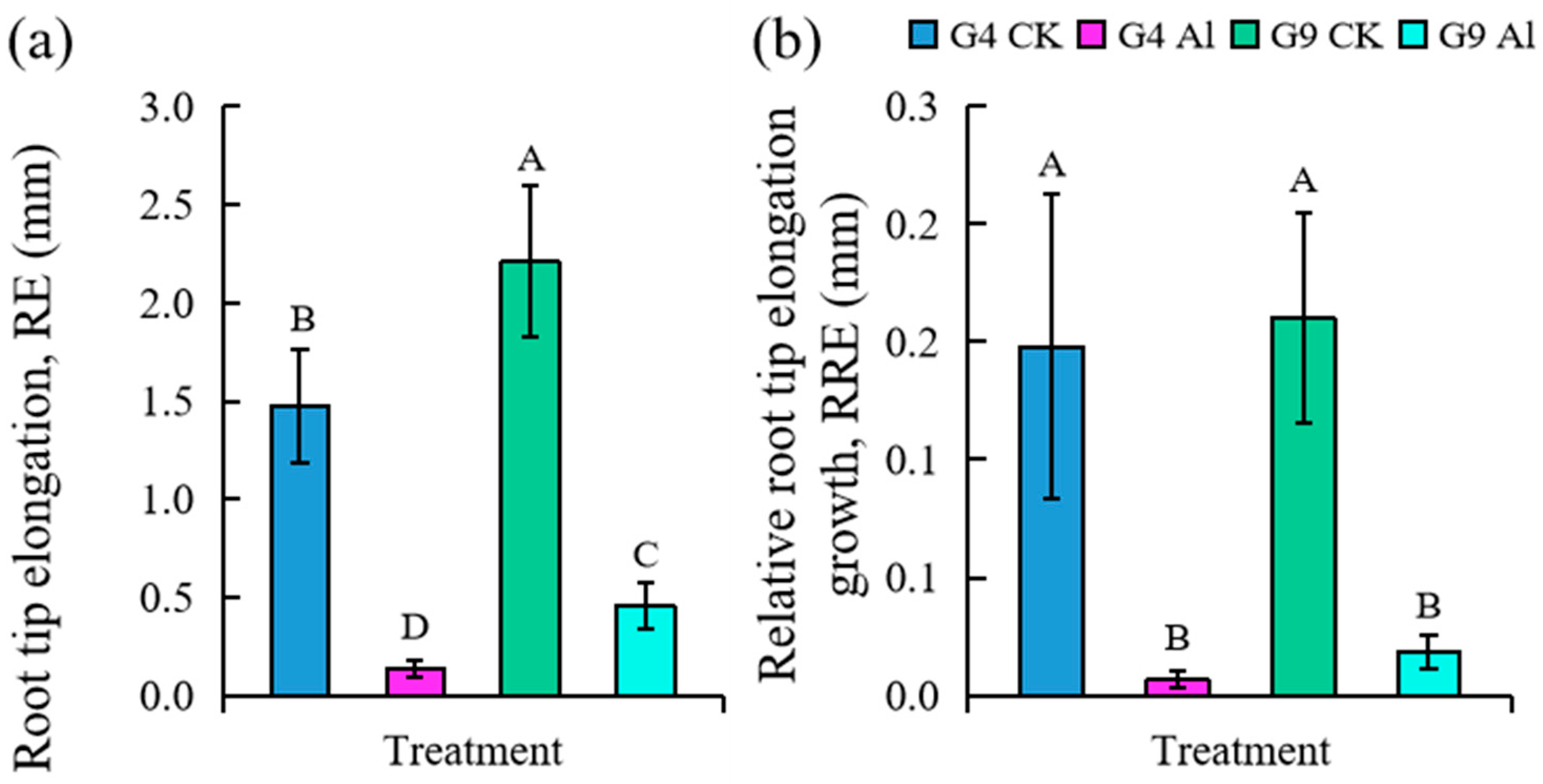
2.1. Effect of Al on Root Tip Surface, PCD Detection, and Change in Root Tip Length
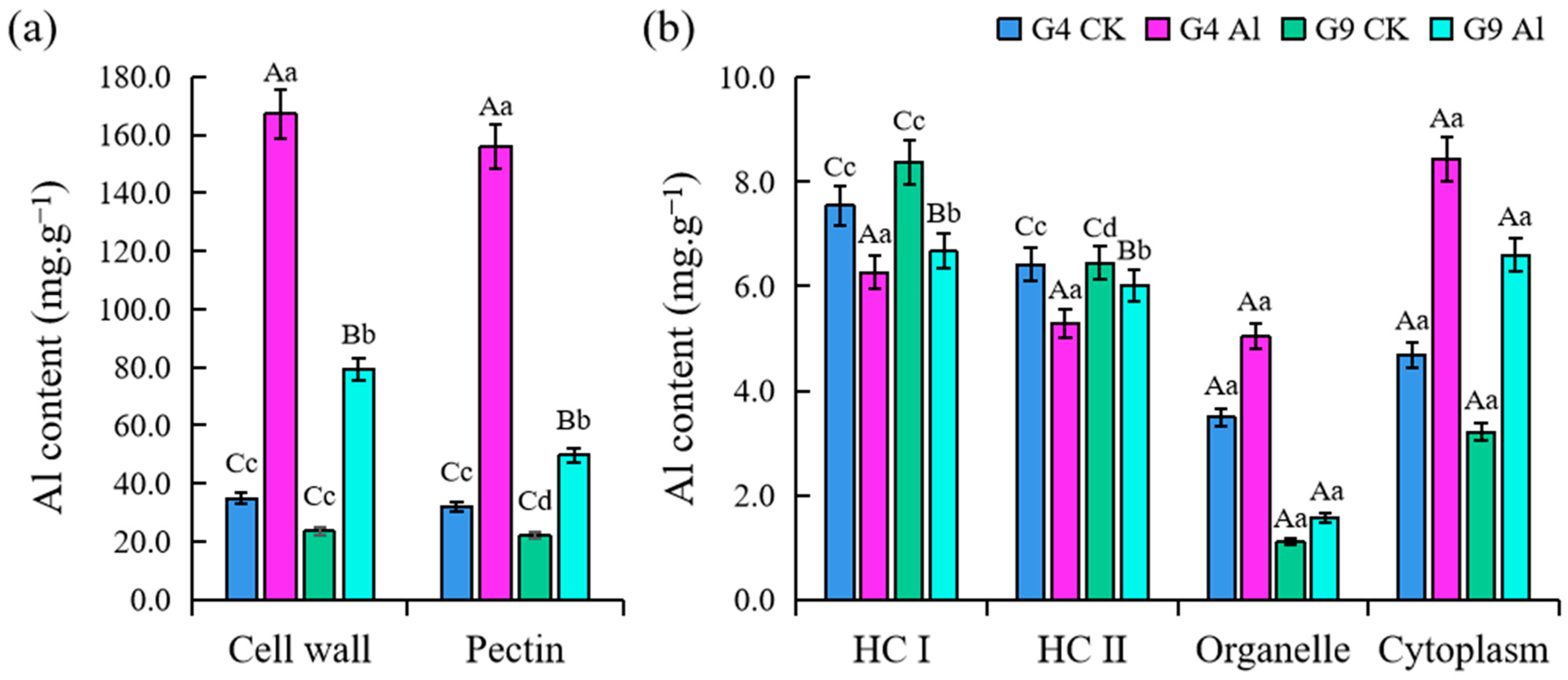
2.2. Distribution of Al3+ in Subcellular Fractions of Root Tip Cells
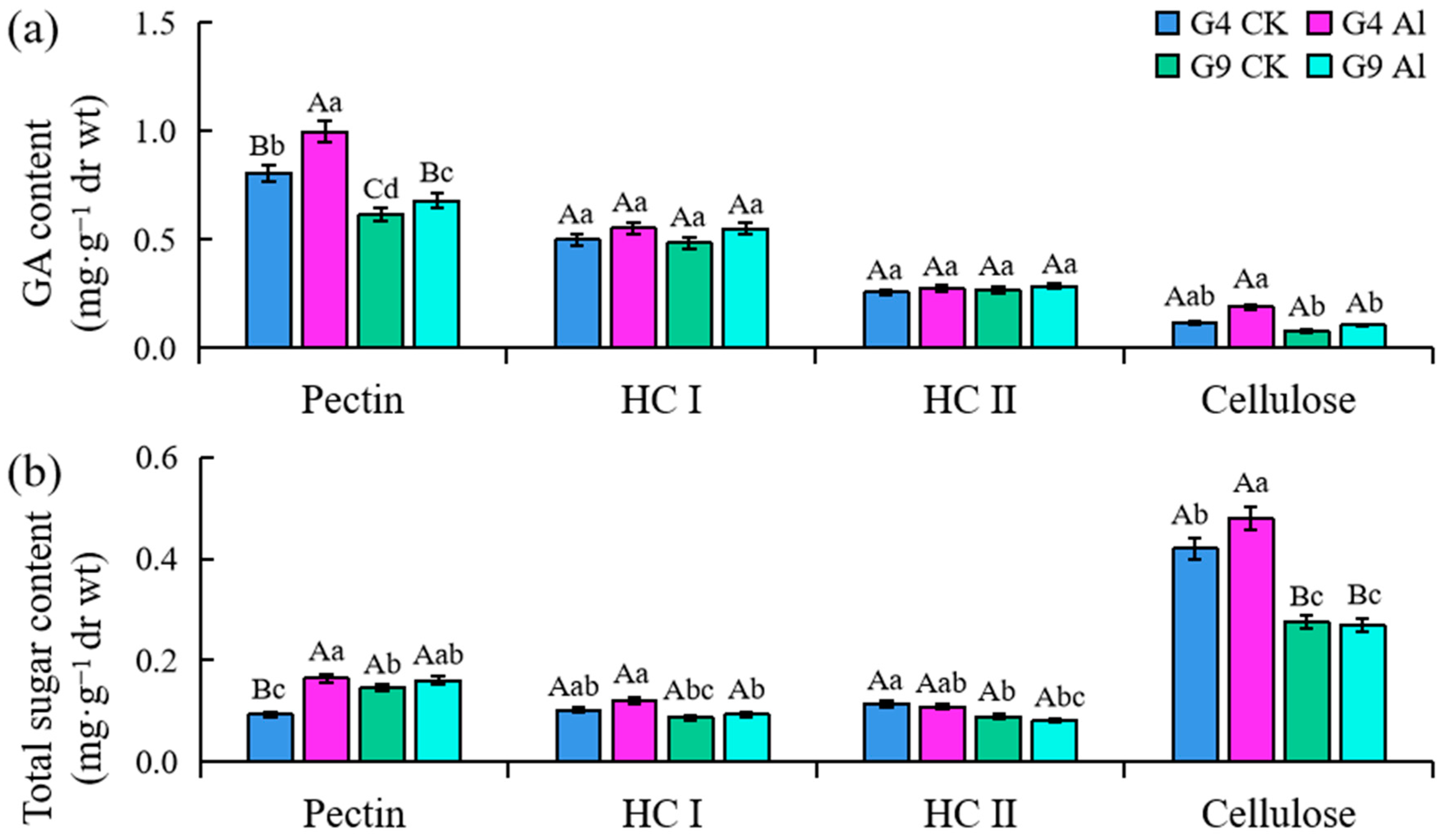
2.3. Contents of Galacturonic Acid (GA) and Total Sugar in Subcellular Components
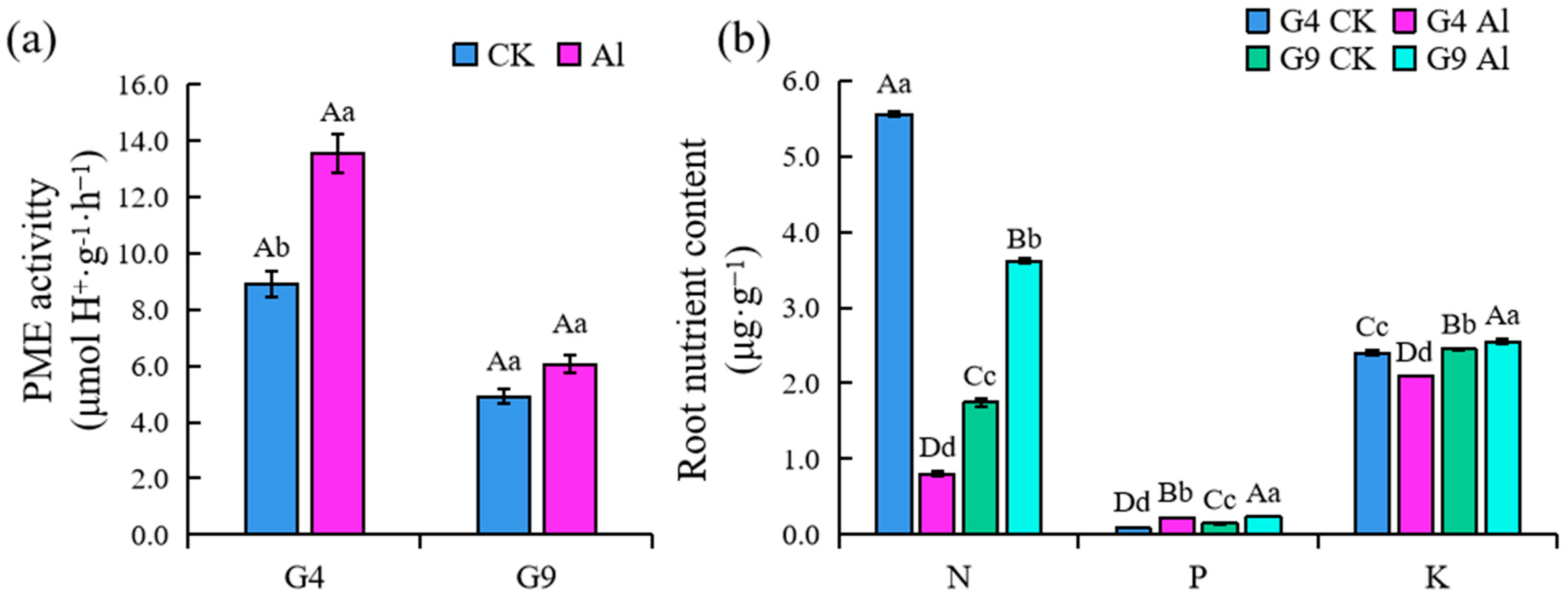
2.4. PME Activity in Root Tip and Root Uptake of N, P, and K
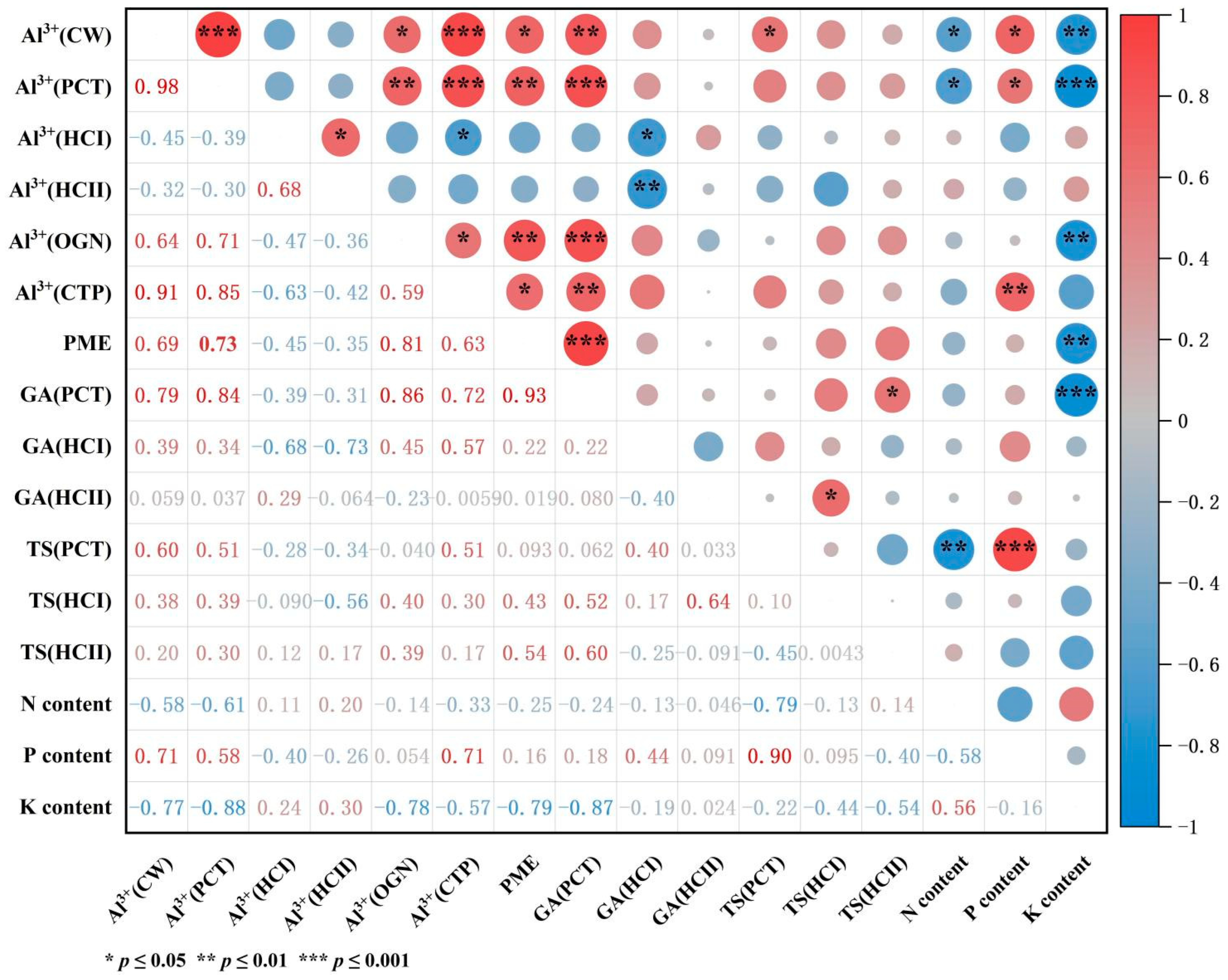
2.5. Correlation Analysis of Al3+ Concentration with N, P, and K Content in Roots, GA and TS Content in Subcellular Fractions, and PME Activity
3. Discussion
3.1. Damage Degree of Root Tip Cells and Characteristics of Programmed Cell Death
3.2. Function of Polysaccharide Components in Root Tip Cell Wall during Al Resistance and Changes in PME Activity
3.3. Root Absorption Characteristics of N, P, and K Nutrients
3.4. Correlation of Aluminum Concentration in Root Tip Subcellular Components with Polysaccharide Content, PME Activity, and Nutrient Content
4. Materials and Methods
4.1. Plant Materials and Experimental Design
4.2. Damage to the Root Tip Surface and PCD Detection
4.3. Cell Wall Extraction in Root Tips
4.4. PME Activity Assay and Determination of Al3+
4.5. Cell Wall Polysaccharide Content and N, P, and K Content in Roots
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, S.H.; Lum, W.C.; Petar, A.; Lubos, K.; Paridah, M.T. Engineering wood products from Eucalyptus spp. Adv. Mater. Sci. Eng. 2022, 2022, 8000780. [Google Scholar]
- Sawaki, Y.; Kihara-Doi, T.; Nishikubo, N.; Kawazu, T.; Kobayashi, Y.; Koyama, H.; Sato, S. Characterization of Al-responsive citrate excretion and citrate-transporting MATEs in Eucalyptus camaldulensis. Planta Int. J. Plant Biol. 2013, 237, 979–989. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.M.; Wen, Y.G.; Yang, M.; Liang, H.W. Correlation analysis on biodiversity and soil physical & chemical properties of Eucalyptus spp. plantations under successive rotation. J. Soil Water Conserv. 2010, 24, 246–250, 256. [Google Scholar]
- Kochian, L.V.; Piñeros, M.A.; Hoekenga, O.A. The physiology, genetics and molecular biology of plant aluminum resistance and toxicity. Plant Soil 2005, 274, 175–195. [Google Scholar] [CrossRef]
- Singh, S.; Tripathi, D.K.; Singh, S.; Sharma, S.; Dubey, N.K.; Chauhan, D.K.; Vaculík, M. Toxicity of aluminium on various levels of plant cells and organism: A review. Environ. Exp. Bot. 2017, 137, 177–193. [Google Scholar] [CrossRef]
- Moriyama, U.; Tomioka, R.; Kojima, M.; Sakakibara, H.; Takenaka, C. Aluminum effect on starch, soluble sugar, and phytohormone in roots of Quercus serrata Thunb. seedlings. Trees 2016, 30, 405–413. [Google Scholar] [CrossRef]
- Liang, Y.H.; Bai, T.D.; Liu, B.; Yu, W.W.; Teng, W.C. Different antioxidant regulation mechanisms in response to aluminum-induced oxidative stress in Eucalyptus species. Ecotox. Environ. Safe. 2022, 241, 113748. [Google Scholar] [CrossRef]
- Tahara, K.; Norisada, M.; Hogetsu, T.; Kojima, K. Aluminum tolerance and aluminum-induced deposition of callose and lignin in the root tips of Melaleuca and Eucalyptus species. J. For. Res. 2005, 10, 325–333. [Google Scholar] [CrossRef]
- Tahara, K.; Hashida, K.; Otsuka, Y.; Ohara, S.; Kojima, K.; Shinohara, K. Identification of a Hydrolyzable Tannin, Oenothein B, as an aluminum-detoxifying ligand in a highly aluminum-resistant tree, Eucalyptus camaldulensis. Plant Physiol. 2014, 164, 683–693. [Google Scholar] [CrossRef] [Green Version]
- Tahara, K.; Kojima, K.; Shinohara, K. Gel-filtration analysis to detect Al-binding ligands in Eucalyptus camaldulensis. J. For. Res. 2015, 20, 411–414. [Google Scholar] [CrossRef]
- Silva, L.F.F.; Lima, M.D.R.; Lima, E.J.A.; Castro, A.R.S.; Junior, U.O.B.; Lobato, A.K.S. Differential behaviours in two species of Eucalyptus ex to aluminium. Indian J. Plant Physiol. 2017, 22, 107–113. [Google Scholar] [CrossRef]
- Takashi, I.; Tsuyoshi, O.; Donghua, L.; Syuntaro, H.; Akio, M. Effect of aluminum on metabolism of organic acids and chemical forms of aluminum in root tips of Eucalyptus camaldulensis Dehnh. Phytochemistry 2013, 94, 142–147. [Google Scholar]
- Yang, M.; Tan, L.; Xu, Y.Y.; Zhao, Y.H.; Cheng, F.; Ye, S.M.; Xin, J.W. Effect of low pH and aluminum toxicity on the photosynthetic characteristics of different fast-growing Eucalyptus vegetatively propagated clones. PLoS ONE 2017, 10, 130963. [Google Scholar] [CrossRef]
- Kinraide, T.B.; Ryan, P.R.; Kochian, L.V. Interactive effects of Al, h, and other cations on root elongation considered in terms of cell-surface electrical potential. Plant Physiol. 1992, 99, 1461–1468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malkanthi, D.R.R.; Yokoyama, K.; Yoshida, T.; Moritsugu, M.; Matsushita, K. Effects of low pH and Al on growth and nutrient uptake of several plants. Soil Sci. Plant Nutr. 2012, 41, 161–165. [Google Scholar] [CrossRef]
- Nguyen, N.T.; Dudzinski, M.J.; Mohapatra, P.K.; Fujita, K. Distribution of accumulated aluminum and changes in cell wall polysaccharides in Eucalyptus camaldulensis and Melaleuca cajuputi under aluminum stress. Soil Sci. Plant Nutr. 2005, 51, 737–740. [Google Scholar] [CrossRef] [Green Version]
- Fabienne, M. Pectin methylesterases: Cell wall enzymes with important roles in plant physiology. Trends Plant Sci. 2001, 6, 414–419. [Google Scholar]
- Yang, J.L.; Zhu, X.F.; Peng, Y.X.; Zheng, C.; Li, G.X.; Liu, Y.; Shi, Y.Z.; Zheng, S.J. Cell wall hemicellulose contributes significantly to aluminum adsorption and root growth in Arabidopsis. Plant Physiol. 2011, 155, 1885–1892. [Google Scholar] [CrossRef] [Green Version]
- Kopittke, P.M.; Menzies, N.W.; Wang, P.; Blamey, F.P.C. Kinetics and nature of aluminium rhizotoxic effects: A review. J. Exp. Bot. 2016, 67, 4451–4467. [Google Scholar] [CrossRef]
- Ma, J.F.; Ryan, P.R.; Delhaize, E. Aluminium tolerance in plants and the complexing role of organic acids. Trends Plant Sci. 2001, 6, 273–278. [Google Scholar] [CrossRef]
- Huang, J.; Han, R.; Ji, F.; Yu, Y.; Wang, R.; Hai, Z.; Liang, W.; Wang, H. Glucose-6-phosphate dehydrogenase and abscisic acid mediate programmed cell death induced by aluminum toxicity in soybean root tips. J. Hazard. Mater. 2022, 425, 127964. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.X.; Liang, Y.C. Silicon prevents aluminum from entering root tip by promoting formation of root border cells in rice. Plant Physiol. Biochem. 2022, 175, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Hawes, M.C.; McLain, J.; Ramirez-Andreotta, M.; Curlango-Rivera, G.; Flores-Lara, Y.; Brigham, L.A. Extracellular trapping of soil contaminants by root border cells: New insights into plant defense. Agronomy 2016, 6, 5. [Google Scholar] [CrossRef]
- Yan, L.; Riaz, M.; Liu, Y.; Zeng, Y.; Jiang, C. Aluminum toxicity could be mitigated with boron by altering the metabolic patterns of amino acids and carbohydrates rather than organic acids in trifoliate orange. Tree Physiol. 2019, 39, 1572–1582. [Google Scholar] [CrossRef]
- Butrinowski, R.T.; Butrinowski, I.T.; Santos, E.L.D.; Picolotto, P.R.; Picolotto, R.A.; Santos, R.F. Water availability in initial development of seedlings in protected environment Eucalyptus grandis. Acta Iguazu 2013, 2, 84–93. [Google Scholar]
- Yang, M.; Cao, X.N.; Wu, Y.M.; Huang, X.L. Effect of acid aluminum on absorption of al and N, P, K in Eucalyptus clones with different aluminum tolerance. Southwest China J. Agric. Sci. 2012, 25, 1061–1064. [Google Scholar]
- Yang, M.; Huang, X.L.; Tan, L.; Wu, Y.M.; Fang, S.Z. Evaluation on aluminum-tolerance of four Eucalyptus clones. J. Cent. South Univ. For. Technol. 2011, 31, 28–33. [Google Scholar]
- Xiao, Z.X.; Yan, G.C.; Ye, M.J.; Liang, Y.C. Silicon relieves aluminum-induced inhibition of cell elongation in rice root apex by reducing the deposition of aluminum in the cell wall. Plant Soil 2021, 462, 189–205. [Google Scholar] [CrossRef]
- Teraoka, T.; Kaneko, M.; Mori, S.; Yoshimura, E. Aluminum rapidly inhibits cellulose synthesis in roots of barley and wheat seedlings. J. Plant Physiol. 2002, 159, 17–23. [Google Scholar] [CrossRef]
- He, H.Y.; He, L.F. Nitric oxide is a suppressor of aluminum-induced mitochondria and caspase-like protease-dependent programmed cell death in plants. Plant Signal. Behav. 2019, 14, 1640566. [Google Scholar] [CrossRef]
- Miyasaka, S.C.; Hawes, M.C. Possible role of root border cells in detection and avoidance of aluminum toxicity. Plant Physiol. 2001, 125, 1978–1987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, J.W.; Zhu, M.Y.; Peng, H.Z.; Wang, L.L. Developmental regulation and biological functions of root border cells in higher plants. Acta Bot. Sin. 2002, 44, 1–8. [Google Scholar]
- Ma, J.F. Syndrome of aluminum toxicity and diversity of aluminum resistance in higher plants. Int. Rev. Cytol. 2007, 264, 225–252. [Google Scholar] [PubMed]
- Brummell, D.A.; Dal Cin, V.; Crisosto, C.H.; Labavitch, J.M. Cell wall metabolism during maturation, ripening and senescence of peach fruit. J. Exp. Bot. 2004, 55, 2029–2039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.F.; Shen, R.; Nagao, S.; Tanimoto, E. Aluminum targets elongating cells by reducing cell wall extensibility in wheat roots. Plant Cell Physiol. 2004, 45, 583–589. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Wang, Q.L.; Zhao, L.F.; Nian, F.Z.; Zhao, Z.Q. Physiological mechanisms of root elongation inhibition induced by changes of root cell wall components under aluminum stress. J. Plant Nutr. Fertil. 2013, 19, 498–509. [Google Scholar]
- Horst, W.J.; Wang, Y.; Eticha, D. The role of the root apoplast in aluminium-induced inhibition of root elongation and in aluminium resistance of plants: A review. Ann. Bot. 2010, 106, 185–197. [Google Scholar] [CrossRef]
- Sun, C.L.; Lu, L.L.; Yu, Y.; Liu, L.J.; Hu, Y.; Ye, Y.Q.; Jin, C.W.; Lin, X.Y. Decreasing methylation of pectin caused by nitric oxide leads to higher aluminium binding in cell walls and greater aluminium sensitivity of wheat roots. J. Exp. Bot. 2016, 67, 979–989. [Google Scholar] [CrossRef] [Green Version]
- Pan, W.H.; Zheng, Z.Z.; Guo, T.R.; Wang, C.; Shuo, J.X.; Pan, J.W. Involvement of PME activity in regulation of Al toxic sensitivity in the root tips of barley. J. Zhejiang Univ. Sci. Ed. 2011, 38, 326–332. [Google Scholar]
- Brunner, I.; Sperisen, C. Aluminum exclusion and aluminum tolerance in woody plants. Front. Plant Sci. 2013, 4, 172. [Google Scholar] [CrossRef]
- Yang, J.L.; Li, Y.Y.; Zhang, Y.J.; Zhang, S.S.; Wu, Y.R.; Wu, P.; Zheng, S.J. Cell wall polysaccharides are specifically involved in the exclusion of aluminum from the rice root apex. Plant Physiol. 2008, 146, 602–611. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Tang, L.; Wang, J.; Zhu, H.; Jin, J.; Yang, J.; Fan, W. Research advances in the mutual mechanisms regulating response of plant roots to phosphate deficiency and aluminum toxicity. Int. J. Mol. Sci. 2022, 23, 1137. [Google Scholar] [CrossRef] [PubMed]
- Li, W.N.; Finnegan, P.M.; Dai, Q.; Guo, D.Q.; Yang, M. Metabolic acclimation supports higher aluminium-induced secretion of citrate and malate in an aluminium-tolerant hybrid clone of Eucalyptus. BMC Plant Biol. 2021, 21, 14. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhao, X.Q.; Chen, R.F.; Dong, X.Y.; Lan, P.; Ma, J.F.; Shen, R.F. Altered cell wall properties are responsible for ammonium-reduced aluminium accumulation in rice roots. Plant Cell Environ. 2015, 38, 1382–1390. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Zeng, F.R.; Qiu, B.Y.; Cai, S.G.; Qiu, L.; Wu, F.B.; Zhang, G.P. Interactive effects of aluminum and chromium stresses on the uptake of nutrients and the metals in barley. Soil Sci. Plant Nutr. 2011, 57, 68–79. [Google Scholar] [CrossRef]
- Dong, Z.X.; Li, Y.B.; Xiao, X.M.; Chen, Y.; Shen, X.F. Silicon effect on growth, nutrient uptake, and yield of peanut (Arachis hypogaea L.) under aluminum stress. J. Plant Nutr. 2018, 41, 2001–2008. [Google Scholar] [CrossRef]
- Li, Y.B.; Xiao, X.M.; Chen, Y.; Shen, X.F. Silicon effect on nutrient acquisition of peanut (Arachis hypogaea L.) under aluminum stress. Commun. Soil Sci. Plant Anal. 2017, 48, 2526–2533. [Google Scholar] [CrossRef]
- Mariano, E.D.; Keltjens, W.G. Long-term effects of aluminum exposure on nutrient uptake by maize genotypes differing in aluminum resistance. J. Plant Nutr. 2005, 28, 323–333. [Google Scholar] [CrossRef]
- Zhao, X.Q.; Chen, R.F.; Shen, R.F. Coadaptation of plants to multiple stresses in acidic soils. Soil Sci. 2014, 179, 503–513. [Google Scholar] [CrossRef]
- Chen, Z.C.; Liao, H. Organic acid anions: An effective defensive weapon for plants against aluminum toxicity and phosphorus deficiency in acidic soils. J. Genet. Genom. 2016, 43, 631–638. [Google Scholar] [CrossRef]
- Magalhaes, J.; Piñeros, M.; Maciel, L.; Kochian, L. Emerging pleiotropic mechanisms underlying aluminum resistance and phosphorus acquisition on acidic soils. Front. Plant Sci. 2018, 9, 1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drouillon, M.; Merckx, R. The role of citric acid as a phosphorus mobilization mechanism in highly P-fixing soils. Gayana. Botánica 2003, 60, 55–62. [Google Scholar] [CrossRef]
- Schmohl, N.; Horst, W.J. Cell wall pectin content modulates aluminium sensitivity of Zea mays (L.) cells grown in suspension culture. Plant Cell Environ. 2000, 23, 735–742. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, Z.Y.; Wu, K.; Xing, C.H.; Chang, H.; Rao, Y.C.; Cai, M.Z. Comparison of aluminum accumulation ability of cell wall polysaccharides in Oryza sativa and Triticum aestivum root tips under aluminum toxicity. Plant Sci. J. 2019, 37, 513–520. [Google Scholar]
- Felipe, F.; Lucelia, B.; Flávio, H.S.R.; Monica, L.R.; Adriana, P.M.; Ricardo, A.A.; José, L. Aluminum-induced stress differently modifies Urochloa genotypes responses on growth and regrowth: Root-to-shoot Al-translocation and oxidative stress. Theor. Exp. Plant Physiol. 2018, 30, 141–152. [Google Scholar]
- Yang, Z.D.; Fang, X.R.; Mu, J.P. Effect of aluminium on the growth and some physiological characters of Eucalyptus seedlings. Guangxi Sci. 1996, 3, 30–33. [Google Scholar]
- Weigel, H.J.; Jäger, H.J. Subcellular distribution and chemical form of cadmium in bean plants. Plant Physiol. 1980, 65, 480–482. [Google Scholar] [CrossRef]
- Bao, S.D. Soil and Agricultural Chemistry Analysis, 3rd ed.; China Agricultural Press: Beijing, China, 2000; pp. 25–114. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| G9 Clone | G4 Clone | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Appraisal Items | Relational Grade (CK) | Ranking (CK) | Relational Grade (Al) | Ranking (Al) | Appraisal Items | Relational Grade (CK) | Ranking (CK) | Relational Grade (Al) | Ranking (Al) |
| TS(CEL) | 0.828 ± 0.084 ** | 5 | 0.997 ± 0.004 ** | 1 | TS(CEL) | 0.980 ± 0.012 | 1 | 0.950 ± 0.031 | 1 |
| TS(PCT) | 0.996 ± 0.004 ** | 1 | 0.900 ± 0.042 ** | 2 | TS(PCT) | 0.888 ± 0.045 | 4 | 0.940 ± 0.052 | 2 |
| GA(HC I) | 0.867 ± 0.099 | 4 | 0.836 ± 0.074 | 3 | GA(PCT) | 0.965 ± 0.024 | 2 | 0.900 ± 0.062 | 3 |
| TS(HC I) | 0.671 ± 0.103 | 7 | 0.822 ± 0.082 | 4 | GA(HC I) | 0.856 ± 0.125 | 5 | 0.882 ± 0.058 | 4 |
| GA(PCT) | 0.949 ± 0.043 ** | 2 | 0.812 ± 0.068 ** | 5 | TS(HC II) | 0.815 ± 0.064 | 6 | 0.842 ± 0.057 | 5 |
| TS(HC II) | 0.911 ± 0.063 | 3 | 0.787 ± 0.161 | 6 | TS(HC I) | 0.742 ± 0.178 | 8 | 0.772 ± 0.115 | 6 |
| GA(HC II) | 0.704 ± 0.237 | 6 | 0.586 ± 0.123 | 7 | GA(HC II) | 0.793 ± 0.119 | 7 | 0.766 ± 0.183 | 7 |
| PME | 0.627 ± 0.126 | 8 | 0.501 ± 0.207 | 8 | PME | 0.895 ± 0.079 | 3 | 0.755 ± 0.160 | 8 |
| GA(CEL) | 0.447 ± 0.097 | 9 | 0.467 ± 0.170 | 9 | GA(CEL) | 0.468 ± 0.178 | 9 | 0.469 ± 0.179 | 9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Ullah, S.; Xu, Y.; Bai, T.; Ye, S.; Jiang, W.; Yang, M. Effects of Elevated Aluminum Concentration and Distribution on Root Damage, Cell Wall Polysaccharides, and Nutrient Uptake in Different Tolerant Eucalyptus Clones. Int. J. Mol. Sci. 2022, 23, 13438. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113438
Li W, Ullah S, Xu Y, Bai T, Ye S, Jiang W, Yang M. Effects of Elevated Aluminum Concentration and Distribution on Root Damage, Cell Wall Polysaccharides, and Nutrient Uptake in Different Tolerant Eucalyptus Clones. International Journal of Molecular Sciences. 2022; 23(21):13438. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113438
Chicago/Turabian StyleLi, Wannian, Saif Ullah, Yuanyuan Xu, Tiandao Bai, Shaoming Ye, Weixin Jiang, and Mei Yang. 2022. "Effects of Elevated Aluminum Concentration and Distribution on Root Damage, Cell Wall Polysaccharides, and Nutrient Uptake in Different Tolerant Eucalyptus Clones" International Journal of Molecular Sciences 23, no. 21: 13438. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113438