
Alteration in the Synaptic and Extrasynaptic Organization of AMPA Receptors in the Hippocampus of P301S Tau Transgenic Mice
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
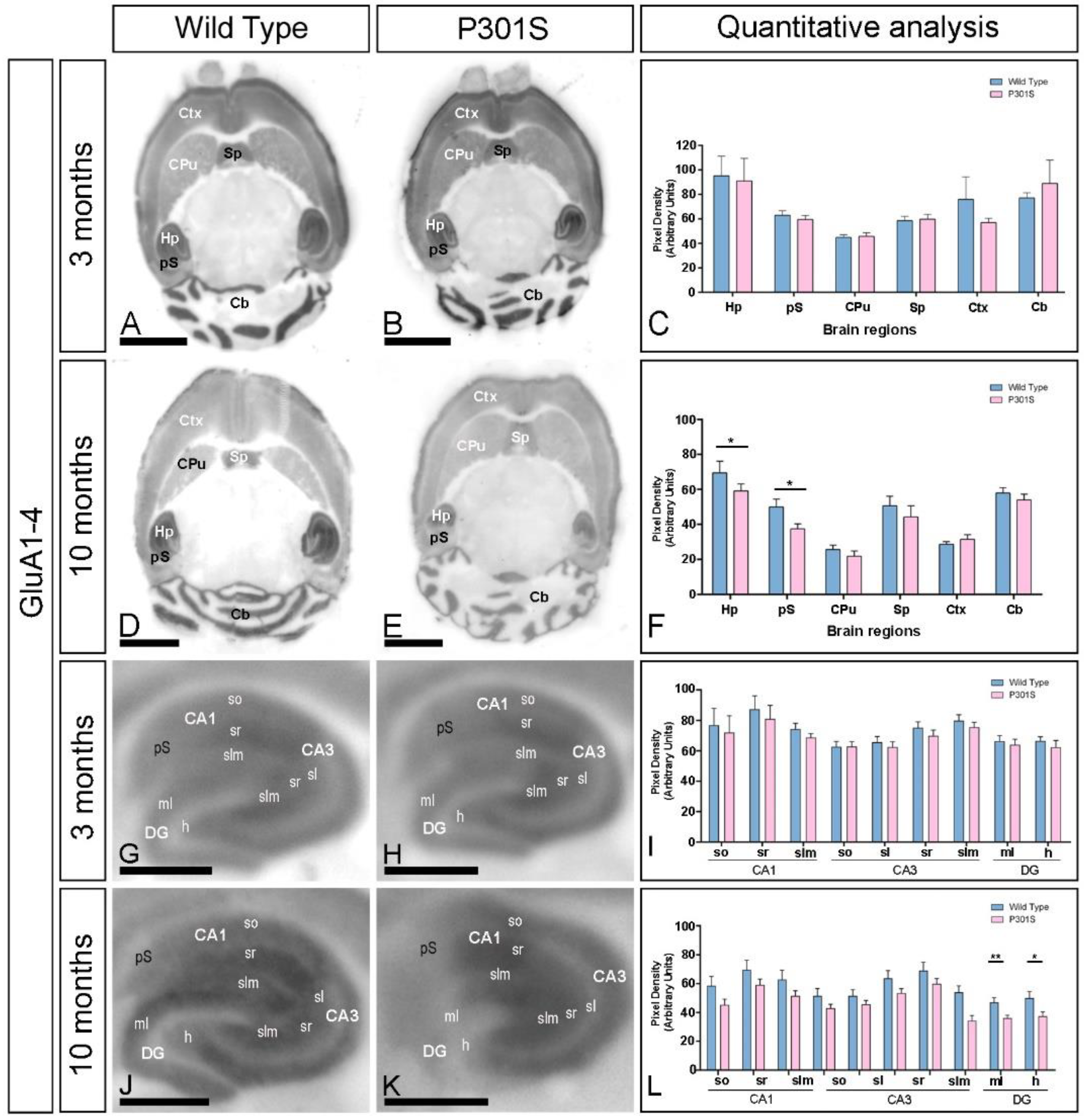
2.1. Age-Dependent Alteration in the GluA1-4 Expression in the Brain of P301S Mice
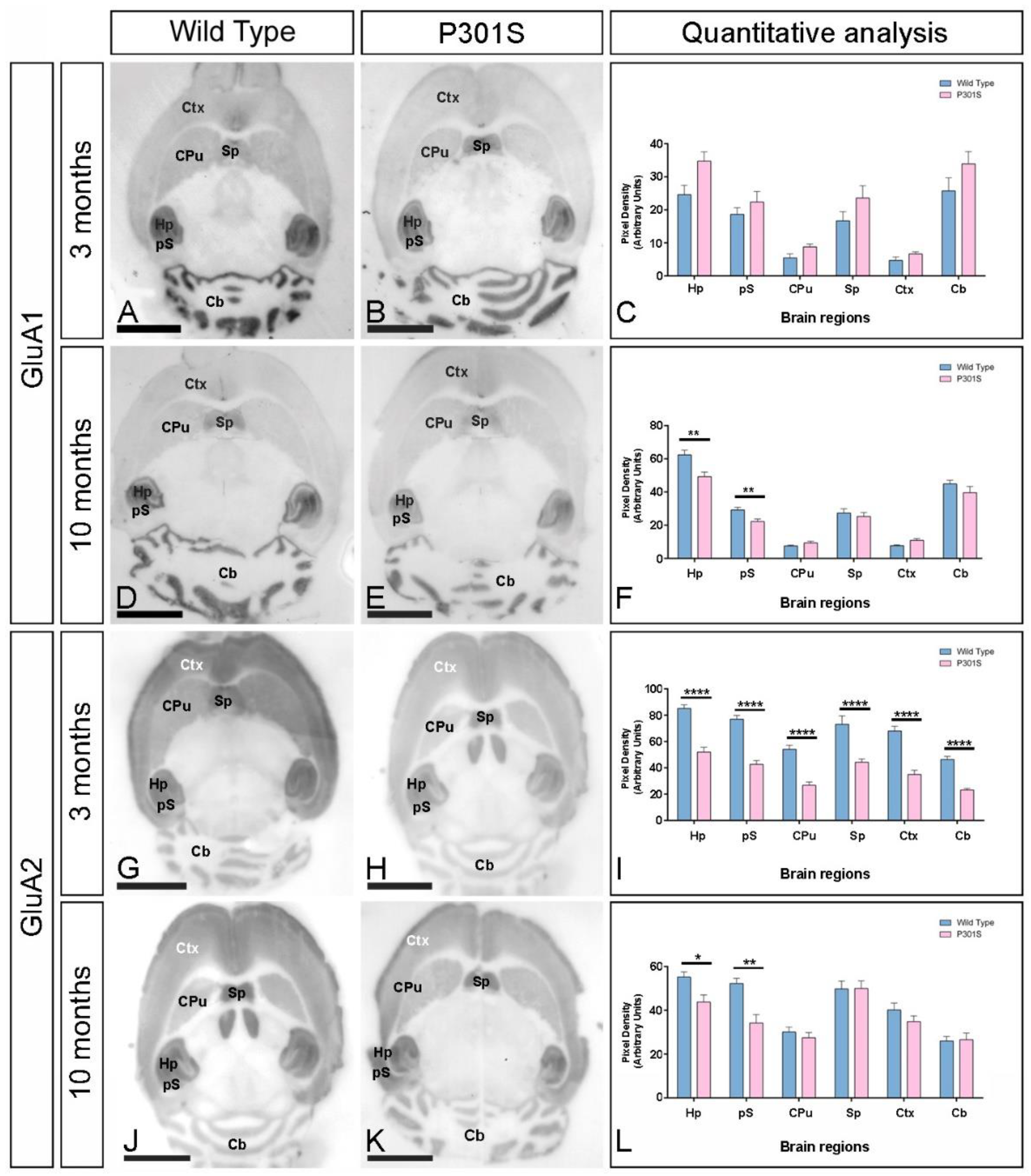
2.2. Early and Differential Alteration of GluA1 and GluA2 Brain Expression in P301S Mice
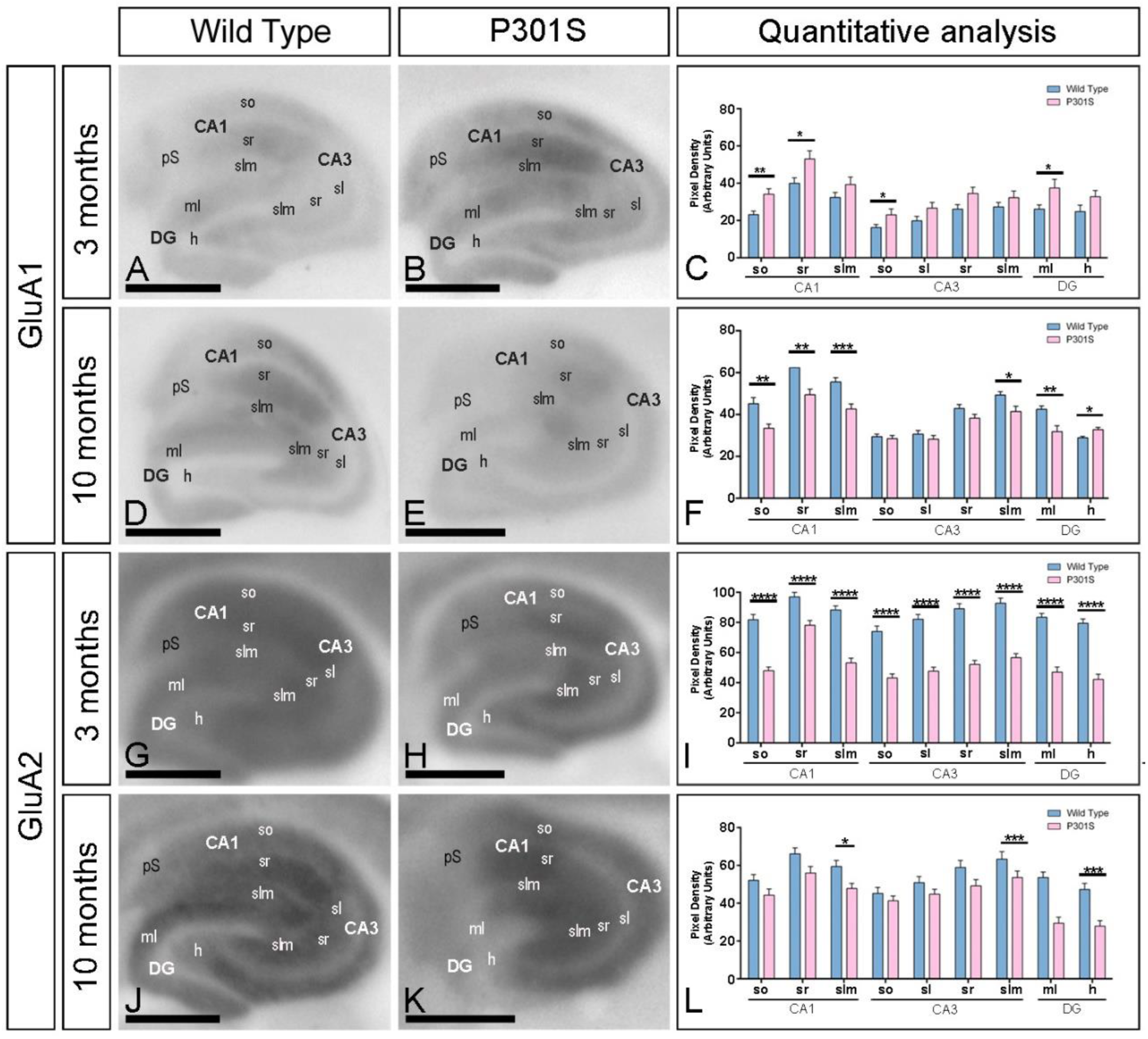
2.3. Reduction in the Expression of AMPA Receptors in the Hippocampus of P301S Mice
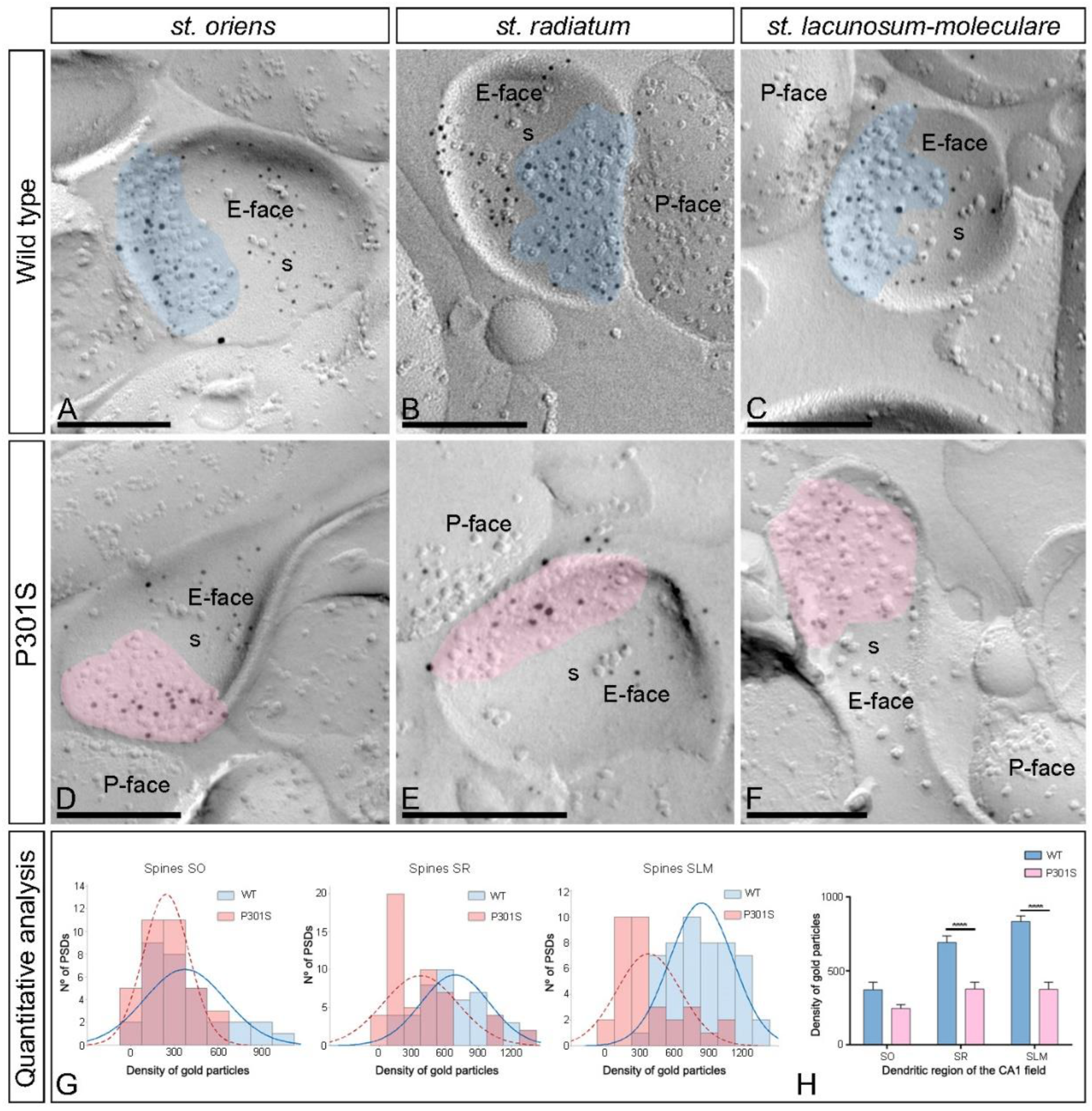
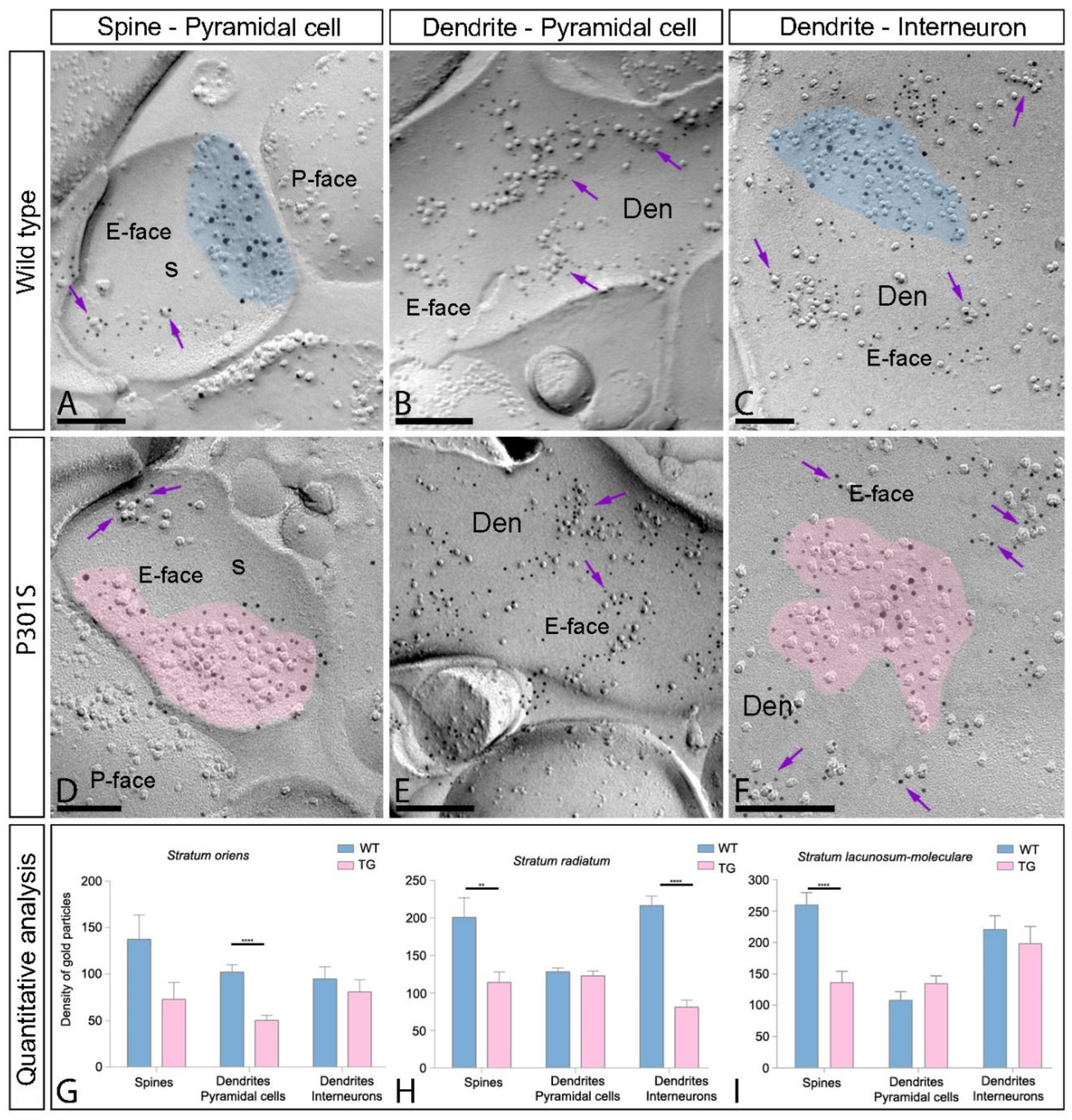
2.4. Differential Reduction of AMPAR s in the Excitatory Synapses in the Spines of P301S Mice
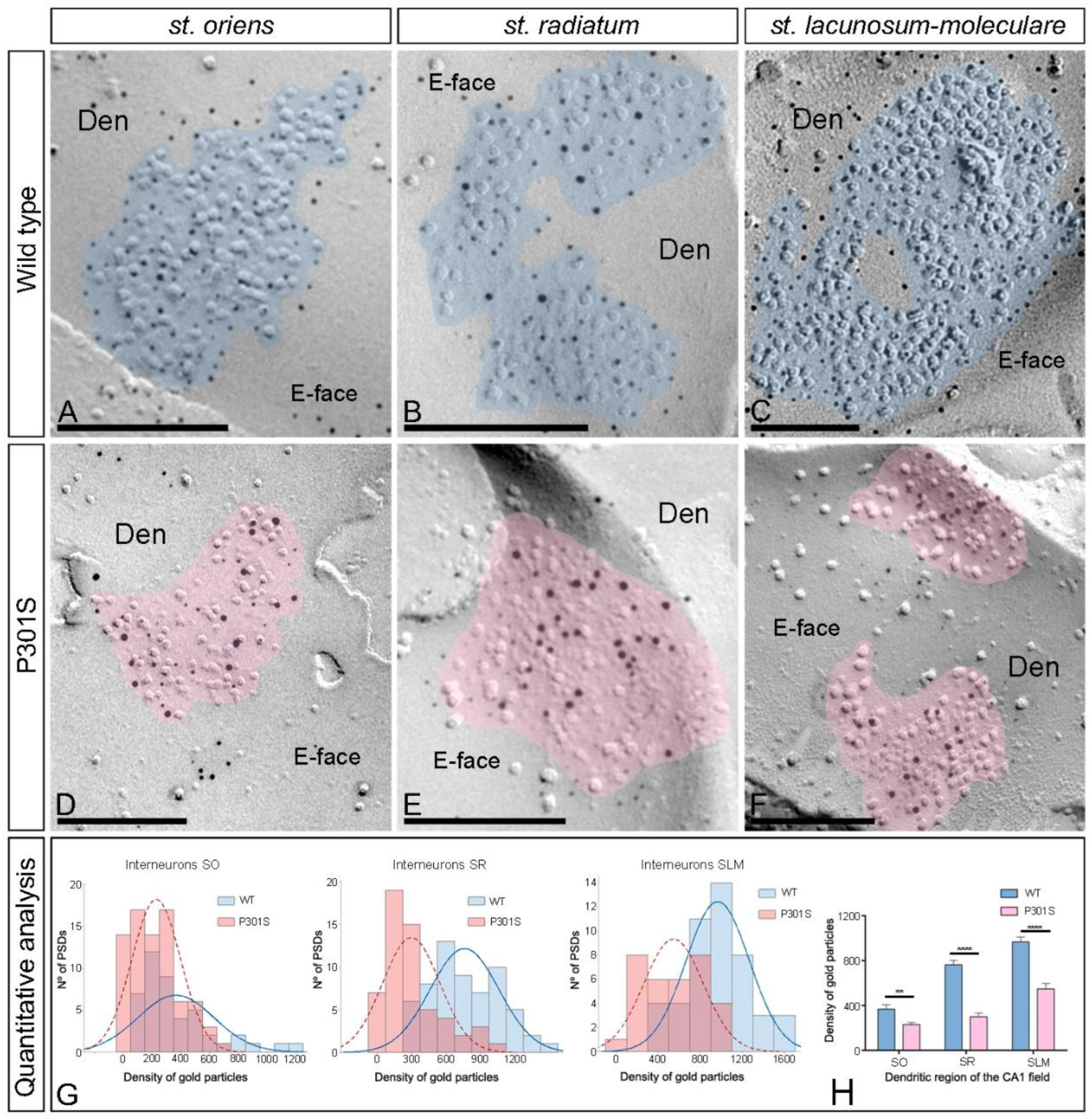
2.5. Reduction of AMPARs in the Excitatory Synapses in the Interneurons of P301S Mice
2.6. Reduction of Extrasynaptic AMPARs in the Hippocampus of P301S Mice
3. Discussion
3.1. Differential Expression of AMPAR Subunits in the Hippocampus of P301S Mice
3.2. Reduction of Synaptic AMPARs in Pyramidal Cells and Interneurons in P301S Mice
3.3. Reduction of Extrasynaptic AMPARs in P301S Mice
4. Material and Methods
4.1. Animals
4.2. Antibodies and Chemicals
4.3. Immunoblots
4.4. Histoblotting
4.5. SDS-Digested Freeze-Fracture Replica Labeling
4.6. Quantification and Analysis of SDS-FRL Data
4.7. Controls
4.8. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hollmann, M.; Heinemann, S. Cloned Glutamate Receptors. Annu. Rev. Neurosci. 1994, 17, 31–108. [Google Scholar] [CrossRef] [PubMed]
- Pin, J.P.; Duvoisin, R. The Metabotropic Glutamate Receptors: Structure and Functions. Neuropharmacology 1995, 34, 1–26. [Google Scholar] [CrossRef]
- Hansen, K.B.; Wollmuth, L.P.; Bowie, D.; Furukawa, H.; Menniti, F.S.; Sobolevsky, A.I.; Swanson, G.T.; Swanger, S.A.; Greger, I.H.; Nakagawa, T.; et al. Structure, Function, and Pharmacology of Glutamate Receptor Ion Channels. Pharm. Rev. 2021, 73, 1469–1658. [Google Scholar] [CrossRef]
- Riedel, G.; Platt, B.; Micheau, J. Glutamate Receptor Function in Learning and Memory. Behav. Brain Res. 2003, 140, 1–47. [Google Scholar] [CrossRef]
- Lee, K.; Goodman, L.; Fourie, C.; Schenk, S.; Leitch, B.; Montgomery, J.M. AMPA Receptors as Therapeutic Targets for Neurological Disorders. Adv. Protein Chem. Struct. Biol. 2016, 103, 203–261. [Google Scholar] [CrossRef]
- Bloom, G.S. Amyloid-β and Tau: The Trigger and Bullet in Alzheimer Disease Pathogenesis. JAMA Neurol. 2014, 71, 505–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Mucke, L. Alzheimer Mechanisms and Therapeutic Strategies. Cell 2012, 148, 1204–1222. [Google Scholar] [CrossRef] [Green Version]
- Boulter, J.; Hollmann, M.; O’Shea-Greenfield, A.; Hartley, M.; Deneris, E.; Maron, C.; Heinemann, S. Molecular Cloning and Functional Expression of Glutamate Receptor Subunit Genes. Science 1990, 249, 1033–1037. [Google Scholar] [CrossRef]
- Hollmann, M.; Hartley, M.; Heinemann, S. Ca2+ Permeability of KA-AMPA—Gated Glutamate Receptor Channels Depends on Subunit Composition. Science 1991, 252, 851–853. [Google Scholar] [CrossRef] [PubMed]
- Keinänen, K.; Wisden, W.; Sommer, B.; Werner, P.; Herb, A.; Verdoorn, T.A.; Sakmann, B.; Seeburg, P.H. A Family of AMPA-Selective Glutamate Receptors. Science 1990, 249, 556–560. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.H.; Hayashi, Y.; Esteban, J.A.; Malinow, R. Subunit-Specific Rules Governing AMPA Receptor Trafficking to Synapses in Hippocampal Pyramidal Neurons. Cell 2001, 105, 331–343. [Google Scholar] [CrossRef]
- Lu, W.; Shi, Y.; Jackson, A.C.; Bjorgan, K.; During, M.J.; Sprengel, R.; Seeburg, P.H.; Nicoll, R.A. Subunit Composition of Synaptic AMPA Receptors Revealed by a Single-Cell Genetic Approach. Neuron 2009, 62, 254–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nusser, Z.; Lujan, R.; Laube, G.; Roberts, J.D.B.; Molnar, E.; Somogyi, P. Cell Type and Pathway Dependence of Synaptic AMPA Receptor Number and Variability in the Hippocampus. Neuron 1998, 21, 545–559. [Google Scholar] [CrossRef] [Green Version]
- Petralia, R.S.; Esteban, J.A.; Wang, Y.X.; Partridge, J.G.; Zhao, H.M.; Wenthold, R.J.; Malinow, R. Selective Acquisition of AMPA Receptors over Postnatal Development Suggests a Molecular Basis for Silent Synapses. Nat. Neurosci. 1999, 2, 31–36. [Google Scholar] [CrossRef]
- Takumi, Y.; Ramírez-León, V.; Laake, P.; Rinvik, E.; Ottersen, O.P. Different Modes of Expression of AMPA and NMDA Receptors in Hippocampal Synapses. Nat. Neurosci. 1999, 2, 618–624. [Google Scholar] [CrossRef] [PubMed]
- Racca, C.; Stephenson, F.A.; Streit, P.; Roberts, J.D.B.; Somogyi, P. NMDA Receptor Content of Synapses in Stratum Radiatum of the Hippocampal CA1 Area. J. Neurosci. 2000, 20, 2512–2522. [Google Scholar] [CrossRef] [Green Version]
- Huganir, R.L.; Nicoll, R.A. AMPARs and Synaptic Plasticity: The Last 25 Years. Neuron 2013, 80, 704–717. [Google Scholar] [CrossRef] [Green Version]
- Nicoll, R.A. A Brief History of Long-Term Potentiation. Neuron 2017, 93, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Jurado, S. AMPA Receptor Trafficking in Natural and Pathological Aging. Front. Mol. Neurosci. 2018, 10, 446. [Google Scholar] [CrossRef] [Green Version]
- Martín-Belmonte, A.; Aguado, C.; Alfaro-Ruíz, R.; Itakura, M.; Moreno-Martínez, A.E.; de la Ossa, L.; Molnár, E.; Fukazawa, Y.; Luján, R. Age-Dependent Shift of AMPA Receptors From Synapses to Intracellular Compartments in Alzheimer’s Disease: Immunocytochemical Analysis of the CA1 Hippocampal Region in APP/PS1 Transgenic Mouse Model. Front. Aging Neurosci. 2020, 12, 577996. [Google Scholar] [CrossRef]
- Yoshiyama, Y.; Higuchi, M.; Zhang, B.; Huang, S.M.; Iwata, N.; Saido, T.C.C.; Maeda, J.; Suhara, T.; Trojanowski, J.Q.; Lee, V.M.Y. Synapse Loss and Microglial Activation Precede Tangles in a P301S Tauopathy Mouse Model. Neuron 2007, 53, 337–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguado, C.; Luján, R. The Histoblot Technique: A Reliable Approach to Analyse Expression Profile of Proteins and to Predict Their Molecular Association. In Neuromethods; Humana Press: New York, NY, USA, 2019; pp. 65–68. [Google Scholar]
- Wenthold, R.J.; Petralia, R.S.; Blahos, J.; Niedzielski, A.S. Evidence for Multiple AMPA Receptor Complexes in Hippocampal CA1/CA2 Neurons. J. Neurosci. 1996, 16, 1982–1989. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, D.J. Alzheimer’s Disease Is a Synaptic Failure. Science 2002, 298, 789–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, D.M.; Selkoe, D.J. Deciphering the Molecular Basis of Memory Failure in Alzheimer’s Disease. Neuron 2004, 44, 181–193. [Google Scholar] [CrossRef] [Green Version]
- Marsh, J.; Alifragis, P. Synaptic Dysfunction in Alzheimer’s Disease: The Effects of Amyloid Beta on Synaptic Vesicle Dynamics as a Novel Target for Therapeutic Intervention. Neural Regen. Res. 2018, 13, 616–623. [Google Scholar] [CrossRef]
- Roberson, E.D.; Scearce-Levie, K.; Palop, J.J.; Yan, F.; Cheng, I.H.; Wu, T.; Gerstein, H.; Yu, G.Q.; Mucke, L. Reducing Endogenous Tau Ameliorates Amyloid Beta-Induced Deficits in an Alzheimer’s Disease Mouse Model. Science 2007, 316, 750–754. [Google Scholar] [CrossRef] [Green Version]
- Ittner, L.M.; Götz, J. Amyloid-β and Tau—A Toxic Pas de Deux in Alzheimer’s Disease. Nat. Rev. Neurosci. 2011, 12, 67–72. [Google Scholar] [CrossRef]
- Choi, S.H.; Kim, Y.H.; Hebisch, M.; Sliwinski, C.; Lee, S.; D’Avanzo, C.; Chen, H.; Hooli, B.; Asselin, C.; Muffat, J.; et al. A Three-Dimensional Human Neural Cell Culture Model of Alzheimer’s Disease. Nature 2014, 515, 274–278. [Google Scholar] [CrossRef] [Green Version]
- Musiek, E.S.; Holtzman, D.M. Three Dimensions of the Amyloid Hypothesis: Time, Space and “Wingmen”. Nat. Neurosci. 2015, 18, 800–806. [Google Scholar] [CrossRef] [Green Version]
- Monaghan, D.T.; Yao, D.; Cotman, C.W. Distribution of [3H]AMPA Binding Sites in Rat Brain as Determined by Quantitative Autoradiography. Brain Res. 1984, 324, 160–164. [Google Scholar] [CrossRef]
- Petralia, R.S.; Wenthold, R.J. Light and Electron Immunocytochemical Localization of AMPA-Selective Glutamate Receptors in the Rat Brain. J. Comp. Neurol. 1992, 318, 329–354. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Kiyama, H.; Tohyama, M. The Differential Expression Patterns of Messenger RNAs Encoding Non-N-Methyl-D-Aspartate Glutamate Receptor Subunits (GluR1-4) in the Rat Brain. Neuroscience 1993, 52, 515–539. [Google Scholar] [CrossRef]
- Cox, D.J.; Racca, C. Differential Dendritic Targeting of AMPA Receptor Subunit MRNAs in Adult Rat Hippocampal Principal Neurons and Interneurons. J. Comp. Neurol. 2013, 521, 1954–2007. [Google Scholar] [CrossRef] [PubMed]
- Helboe, L.; Egebjerg, J.; Barkholt, P.; Volbracht, C. Early Depletion of CA1 Neurons and Late Neurodegeneration in a Mouse Tauopathy Model. Brain Res. 2017, 1665, 22–35. [Google Scholar] [CrossRef]
- Thies, E.; Mandelkow, E.M. Missorting of Tau in Neurons Causes Degeneration of Synapses That Can Be Rescued by the Kinase MARK2/Par-1. J. Neurosci. 2007, 27, 2896–2907. [Google Scholar] [CrossRef] [Green Version]
- Hoover, B.R.; Reed, M.N.; Su, J.; Penrod, R.D.; Kotilinek, L.A.; Grant, M.K.; Pitstick, R.; Carlson, G.A.; Lanier, L.M.; Yuan, L.L.; et al. Tau Mislocalization to Dendritic Spines Mediates Synaptic Dysfunction Independently of Neurodegeneration. Neuron 2010, 68, 1067–1081. [Google Scholar] [CrossRef] [Green Version]
- Greenamyre, J.T.; Penney, J.B.; D’Amato, C.J.; Young, A.B. Dementia of the Alzheimer’s Type: Changes in Hippocampal L-[3H]Glutamate Binding. J. Neurochem. 1987, 48, 543–551. [Google Scholar] [CrossRef] [Green Version]
- Jansen, K.L.R.; Faull, R.L.M.; Dragunow, M.; Synek, B.L. Alzheimer’s Disease: Changes in Hippocampal N-Methyl-D-Aspartate, Quisqualate, Neurotensin, Adenosine, Benzodiazepine, Serotonin and Opioid Receptors--an Autoradiographic Study. Neuroscience 1990, 39, 613–627. [Google Scholar] [CrossRef]
- Penney, J.B.; Maragos, W.F.; Greenamyre, J.T.; Debowey, D.L.; Hollingsworth, Z.; Young, A.B. Excitatory Amino Acid Binding Sites in the Hippocampal Region of Alzheimer’s Disease and Other Dementias. J. Neurol. Neurosurg. Psychiatry 1990, 53, 314–320. [Google Scholar] [CrossRef]
- Shepherd, J.D.; Huganir, R.L. The Cell Biology of Synaptic Plasticity: AMPA Receptor Trafficking. Annu. Rev. Cell Dev. Biol. 2007, 23, 613–643. [Google Scholar] [CrossRef]
- Mansour, M.; Nagarajan, N.; Nehring, R.B.; Clements, J.D.; Rosenmund, C. Heteromeric AMPA Receptors Assemble with a Preferred Subunit Stoichiometry and Spatial Arrangement. Neuron 2001, 32, 841–853. [Google Scholar] [CrossRef] [Green Version]
- Isaac, J.T.R.; Ashby, M.; McBain, C.J. The Role of the GluR2 Subunit in AMPA Receptor Function and Synaptic Plasticity. Neuron 2007, 54, 859–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, H.; O’Brien, R.J.; Fung, E.T.; Lanahan, A.A.; Worley, P.F.; Huganir, R.L. GRIP: A Synaptic PDZ Domain-Containing Protein That Interacts with AMPA Receptors. Nature 1997, 386, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Nishimune, A.; Isaac, J.T.R.; Molnar, E.; Noel, J.; Nash, S.R.; Tagaya, M.; Collingridge, G.L.; Nakanishi, S.; Henley, J.M. NSF Binding to GluR2 Regulates Synaptic Transmission. Neuron 1998, 21, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Pozo, K.; Cingolani, L.A.; Bassani, S.; Laurent, F.; Passafaro, M.; Goda, Y. Β3 Integrin Interacts Directly with GluA2 AMPA Receptor Subunit and Regulates AMPA Receptor Expression in Hippocampal Neurons. Proc. Natl. Acad. Sci. USA 2012, 109, 1323–1328. [Google Scholar] [CrossRef] [Green Version]
- Passafaro, M.; Nakagawa, T.; Sala, C.; Sheng, M. Induction of Dendritic Spines by an Extracellular Domain of AMPA Receptor Subunit GluR2. Nature 2003, 424, 677–681. [Google Scholar] [CrossRef]
- Saglietti, L.; Dequidt, C.; Kamieniarz, K.; Rousset, M.C.; Valnegri, P.; Thoumine, O.; Beretta, F.; Fagni, L.; Choquet, D.; Sala, C.; et al. Extracellular Interactions between GluR2 and N-Cadherin in Spine Regulation. Neuron 2007, 54, 461–477. [Google Scholar] [CrossRef] [Green Version]
- Klausberger, T.; Somogyi, P. Neuronal Diversity and Temporal Dynamics: The Unity of Hippocampal Circuit Operations. Science 2008, 321, 53–57. [Google Scholar] [CrossRef] [Green Version]
- Klausberger, T. GABAergic Interneurons Targeting Dendrites of Pyramidal Cells in the CA1 Area of the Hippocampus. Eur. J. Neurosci. 2009, 30, 947–957. [Google Scholar] [CrossRef]
- Nyíri, G.; Stephenson, F.A.; Freund, T.F.; Somogyi, P. Large Variability in Synaptic N-Methyl-D-Aspartate Receptor Density on Interneurons and a Comparison with Pyramidal-Cell Spines in the Rat Hippocampus. Neuroscience 2003, 119, 347–363. [Google Scholar] [CrossRef]
- Tanaka, J.I.; Matsuzaki, M.; Tarusawa, E.; Momiyama, A.; Molnar, E.; Kasai, H.; Shigemoto, R. Number and Density of AMPA Receptors in Single Synapses in Immature Cerebellum. J. Neurosci. 2005, 25, 799–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, I.; Huganir, R.L. Regulation of AMPA Receptors during Synaptic Plasticity. Trends Neurosci. 2002, 25, 578–588. [Google Scholar] [CrossRef]
- Ittner, L.M.; Ke, Y.D.; Delerue, F.; Bi, M.; Gladbach, A.; van Eersel, J.; Wölfing, H.; Chieng, B.C.; Christie, M.J.; Napier, I.A.; et al. Dendritic Function of Tau Mediates Amyloid-Beta Toxicity in Alzheimer’s Disease Mouse Models. Cell 2010, 142, 387–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Kotilinek, L.A.; Smith, B.; Hlynialuk, C.; Zahs, K.; Ramsden, M.; Cleary, J.; Ashe, K.H. Caspase-2 Cleavage of Tau Reversibly Impairs Memory. Nat. Med. 2016, 22, 1268–1276. [Google Scholar] [CrossRef] [PubMed]
- Verret, L.; Mann, E.O.; Hang, G.B.; Barth, A.M.I.; Cobos, I.; Ho, K.; Devidze, N.; Masliah, E.; Kreitzer, A.C.; Mody, I.; et al. Inhibitory Interneuron Deficit Links Altered Network Activity and Cognitive Dysfunction in Alzheimer Model. Cell 2012, 149, 708–721. [Google Scholar] [CrossRef] [Green Version]
- Palop, J.J.; Mucke, L. Amyloid-Beta-Induced Neuronal Dysfunction in Alzheimer’s Disease: From Synapses toward Neural Networks. Nat. Neurosci. 2010, 13, 812–818. [Google Scholar] [CrossRef] [Green Version]
- Hazra, A.; Gu, F.; Aulakh, A.; Berridge, C.; Eriksen, J.L.; Žiburkus, J. Inhibitory Neuron and Hippocampal Circuit Dysfunction in an Aged Mouse Model of Alzheimer’s Disease. PLoS ONE 2013, 8, e64318. [Google Scholar] [CrossRef] [Green Version]
- Man, H.Y.; Lin, J.W.; Ju, W.H.; Ahmadian, G.; Liu, L.; Becker, L.E.; Sheng, M.; Wang, Y.T. Regulation of AMPA Receptor-Mediated Synaptic Transmission by Clathrin-Dependent Receptor Internalization. Neuron 2000, 25, 649–662. [Google Scholar] [CrossRef] [Green Version]
- Petralia, R.S.; Wang, Y.X.; Wenthold, R.J. Internalization at Glutamatergic Synapses during Development. Eur. J. Neurosci. 2003, 18, 3207–3217. [Google Scholar] [CrossRef]
- Borgdorff, A.J.; Choquet, D. Regulation of AMPA Receptor Lateral Movements. Nature 2002, 417, 649–653. [Google Scholar] [CrossRef]
- Petrini, E.M.; Lu, J.; Cognet, L.; Lounis, B.; Ehlers, M.D.; Choquet, D. Endocytic Trafficking and Recycling Maintain a Pool of Mobile Surface AMPA Receptors Required for Synaptic Potentiation. Neuron 2009, 63, 92–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masugi-Tokita, M.; Shigemoto, R. High-Resolution Quantitative Visualization of Glutamate and GABA Receptors at Central Synapses. Curr. Opin. Neurobiol. 2007, 17, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Pallas-Bazarra, N.; Draffin, J.; Cuadros, R.; Antonio Esteban, J.; Avila, J. Tau Is Required for the Function of Extrasynaptic NMDA Receptors. Sci. Rep. 2019, 9, 9116. [Google Scholar] [CrossRef] [Green Version]
- Alfaro-Ruiz, R.; Aguado, C.; Martín-Belmonte, A.; Moreno-Martínez, A.E.; Merchán-Rubira, J.; Hernández, F.; Ávila, J.; Fukazawa, Y.; Luján, R. Different Modes of Synaptic and Extrasynaptic NMDA Receptor Alteration in the Hippocampus of P301S Tau Transgenic Mice. Brain Pathol. 2022, e13115. [Google Scholar] [CrossRef] [PubMed]
- Opazo, P.; Viana da Silva, S.; Carta, M.; Breillat, C.; Coultrap, S.J.; Grillo-Bosch, D.; Sainlos, M.; Coussen, F.; Bayer, K.U.; Mulle, C.; et al. CaMKII Metaplasticity Drives Aβ Oligomer-Mediated Synaptotoxicity. Cell Rep. 2018, 23, 3137–3145. [Google Scholar] [CrossRef]
- Fukaya, M.; Tsujita, M.; Yamazaki, M.; Kushiya, E.; Abe, M.; Akashi, K.; Natsume, R.; Kano, M.; Kamiya, H.; Watanabe, M.; et al. Abundant distribution of TARP g-8 in synaptic and extrasynaptic surface of hippocampal neurons and its major role inAMPA receptor expression on spines and dendrites. Eur. J. Neurosci. 2006, 24, 2177–2190. [Google Scholar] [CrossRef] [Green Version]
- Siegel, S.J.; Brose, N.; Janssen, W.G.; Gasic, G.P.; Jahn, R.; Heinemann, S.F.; Morrison, J.H. Regional, Cellular, and Ultrastructural Distribution of N-Methyl-D-Aspartate Receptor Subunit 1 in Monkey Hippocampus. Proc. Natl. Acad. Sci. USA 1994, 91, 564–568. [Google Scholar] [CrossRef] [Green Version]
- Iwasato, T.; Datwani, A.; Wolf, A.M.; Nishiyama, H.; Taguchi, Y.; Tonegawa, S.; Knöpfel, T.; Erzurumlu, R.S.; Itohara, S. Cortex-Restricted Disruption of NMDAR1 Impairs Neuronal Patterns in the Barrel Cortex. Nature 2000, 406, 726–731. [Google Scholar] [CrossRef] [Green Version]
- Szabadits, E.; Cserép, C.; Szonyi, A.; Fukazawa, Y.; Shigemoto, R.; Watanabe, M.; Itohara, S.; Freund, T.F.; Nyiri, G. NMDA Receptors in Hippocampal GABAergic Synapses and Their Role in Nitric Oxide Signaling. J. Neurosci. 2011, 31, 5893–5904. [Google Scholar] [CrossRef] [Green Version]
- Luján, R.; Aguado, C.; Ciruela, F.; Cózar, J.; Kleindienst, D.; de la Ossa, L.; Bettler, B.; Wickman, K.; Watanabe, M.; Shigemoto, R.; et al. Differential Association of GABA B Receptors with Their Effector Ion Channels in Purkinje Cells. Brain Struct. Funct. 2018, 223, 1565–1587. [Google Scholar] [CrossRef]
- Harris, K.M.; Landis, D.M.D. Membrane Structure at Synaptic Junctions in Area CA1 of the Rat Hippocampus. Neuroscience 1986, 19, 857–872. [Google Scholar] [CrossRef]
- Antal, M.; Fukazawa, Y.; Eördögh, M.; Muszil, D.; Molnár, E.; Itakura, M.; Takahashi, M.; Shigemoto, R. Numbers, Densities, and Colocalization of AMPA- and NMDA-Type Glutamate Receptors at Individual Synapses in the Superficial Spinal Dorsal Horn of Rats. J. Neurosci. 2008, 28, 9692–9701. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stratum oriens | Stratum radiatum | Stratum lacunosum-moleculare | ||||
|---|---|---|---|---|---|---|
| Spines | Interneurons | Spines | Interneurons | Spines | Interneurons | |
| WT | ||||||
| Excitatory Synapses (n) | 31 | 45 | 48 | 61 | 51 | 47 |
| Area of synapses (µm) | 0.036 ± 0.003 | 0.061 ± 0.004 | 0.032 ± 0.002 | 0.055 ± 0.004 | 0.033 ± 0.002 | 0.063 ± 0.005 |
| Median gold particles | 9 | 19 | 18 | 29 | 22 | 50 |
| Range | 53–1 | 87–2 | 76–4 | 162–9 | 102–8 | 199–11 |
| Density gold particles (µm2) | ||||||
| Mean (±SEM) | 373.25 ± 49.93 | 367.0 ± 39.31 | 692.70 ± 44.43 | 764.07 ± 38.37 | 833.86 ± 38.45 | 967.70 ± 43.95 |
| Median | 333.87 | 301.12 | 627.02 | 723.18 | 818.89 | 1003.70 |
| Range | 1065.06–30.72 | 1172.68–52.22 | 1314.80–185.87 | 1455.36–292.7 | 1276.96–294.75 | 1665.71–378.67 |
| P301S | ||||||
| Excitatory Synapses (n) | 35 | 78 | 52 | 56 | 33 | 33 |
| Area of synapses (µm) | 0.034 ± 0.004 | 0.069 ± 0.004 | 0.029 ± 0.002 | 0.067 ± 0.006 | 0.031 ± 0.002 | 0.068 ± 0.007 |
| Median gold particles | 6 | 11 | 7.5 | 10 | 8 | 28 |
| Range | 47–1 | 62–1 | 47–1 | 151–2 | 43–1 | 135–3 |
| Density gold particles (µm2) | ||||||
| Mean (±SEM) | 245.44 ± 26.64 | 230.02 ± 19.28 | 377.41 ± 46.73 | 300.01 ± 33.29 | 375.59 ± 47.96 | 548.08 ± 49.05 |
| Median | 253.43 | 224.24 | 261.39 | 227.44 | 316.55 | 535.08 |
| Range | 575.28–26.03 | 695.81–14.84 | 1390.82–26.75 | 981.99–34.91 | 1033.89–53.82 | 1001.05–66.32 |
| Stratum oriens | Stratum radiatum | Stratum lacunosum-moleculare | |||||||
|---|---|---|---|---|---|---|---|---|---|
| PC Spines | PC Dendrites | Interneurons | PC Spines | PC Dendrites | Interneurons | PC Spines | PC Dendrites | Interneurons | |
| WT | |||||||||
| Extrasynaptic compartments (n) | 23 | 25 | 38 | 38 | 30 | 58 | 50 | 30 | 48 |
| Density gold particles (µm2) | |||||||||
| Mean (±SEM) | 137.83 ± 26.12 | 102.29 ± 7.75 | 94.94 ± 13.07 | 201 ± 26.02 | 128.3 ± 5.41 | 216.7 ± 12.47 | 260.3 ± 19.13 | 108.7 ± 12.89 | 221.1 ± 21.58 |
| Median | 91 | 102 | 71.56 | 149.3 | 122.5 | 207.5 | 267.6 | 85.46 | 186.2 |
| Range | 401.3–16.94 | 187.8–3.95 | 306.3–5.95 | 711.3–19.26 | 196.1–89.92 | 603.2–74.33 | 629.2–25.67 | 332–14.98 | 765.4–11.17 |
| P301S | |||||||||
| Extrasynaptic compartments (n) | 15 | 31 | 42 | 39 | 23 | 40 | 26 | 28 | 28 |
| Density gold particles (µm2) | |||||||||
| Mean (±SEM) | 72.86 ± 18.23 | 50.29 ± 5.71 | 81.10 ± 13.02 | 114 ± 13.97 | 126.1 ± 6.76 | 81.28 ± 9.40 | 136.8 ± 17.28 | 135.2 ± 11.64 | 198.8 ± 26.52 |
| Median | 40.97 | 42.31 | 58.28 | 92.55 | 117.2 | 60.78 | 140.3 | 150.5 | 197.6 |
| Range | 234.5–18.72 | 126.7–5.55 | 406.4–2.25 | 442.1–11 | 191.3–70.83 | 243.1–6.69 | 382.4–20.68 | 291.3–15.00 | 612.7–19.97 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alfaro-Ruiz, R.; Aguado, C.; Martín-Belmonte, A.; Moreno-Martínez, A.E.; Merchán-Rubira, J.; Hernández, F.; Ávila, J.; Fukazawa, Y.; Luján, R. Alteration in the Synaptic and Extrasynaptic Organization of AMPA Receptors in the Hippocampus of P301S Tau Transgenic Mice. Int. J. Mol. Sci. 2022, 23, 13527. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113527
Alfaro-Ruiz R, Aguado C, Martín-Belmonte A, Moreno-Martínez AE, Merchán-Rubira J, Hernández F, Ávila J, Fukazawa Y, Luján R. Alteration in the Synaptic and Extrasynaptic Organization of AMPA Receptors in the Hippocampus of P301S Tau Transgenic Mice. International Journal of Molecular Sciences. 2022; 23(21):13527. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113527
Chicago/Turabian StyleAlfaro-Ruiz, Rocio, Carolina Aguado, Alejandro Martín-Belmonte, Ana Esther Moreno-Martínez, Jesús Merchán-Rubira, Félix Hernández, Jesús Ávila, Yugo Fukazawa, and Rafael Luján. 2022. "Alteration in the Synaptic and Extrasynaptic Organization of AMPA Receptors in the Hippocampus of P301S Tau Transgenic Mice" International Journal of Molecular Sciences 23, no. 21: 13527. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113527