
Identification of Hypoxia-Related Prognostic Signature and Competing Endogenous RNA Regulatory Axes in Hepatocellular Carcinoma

Abstract
:1. Introduction
2. Results
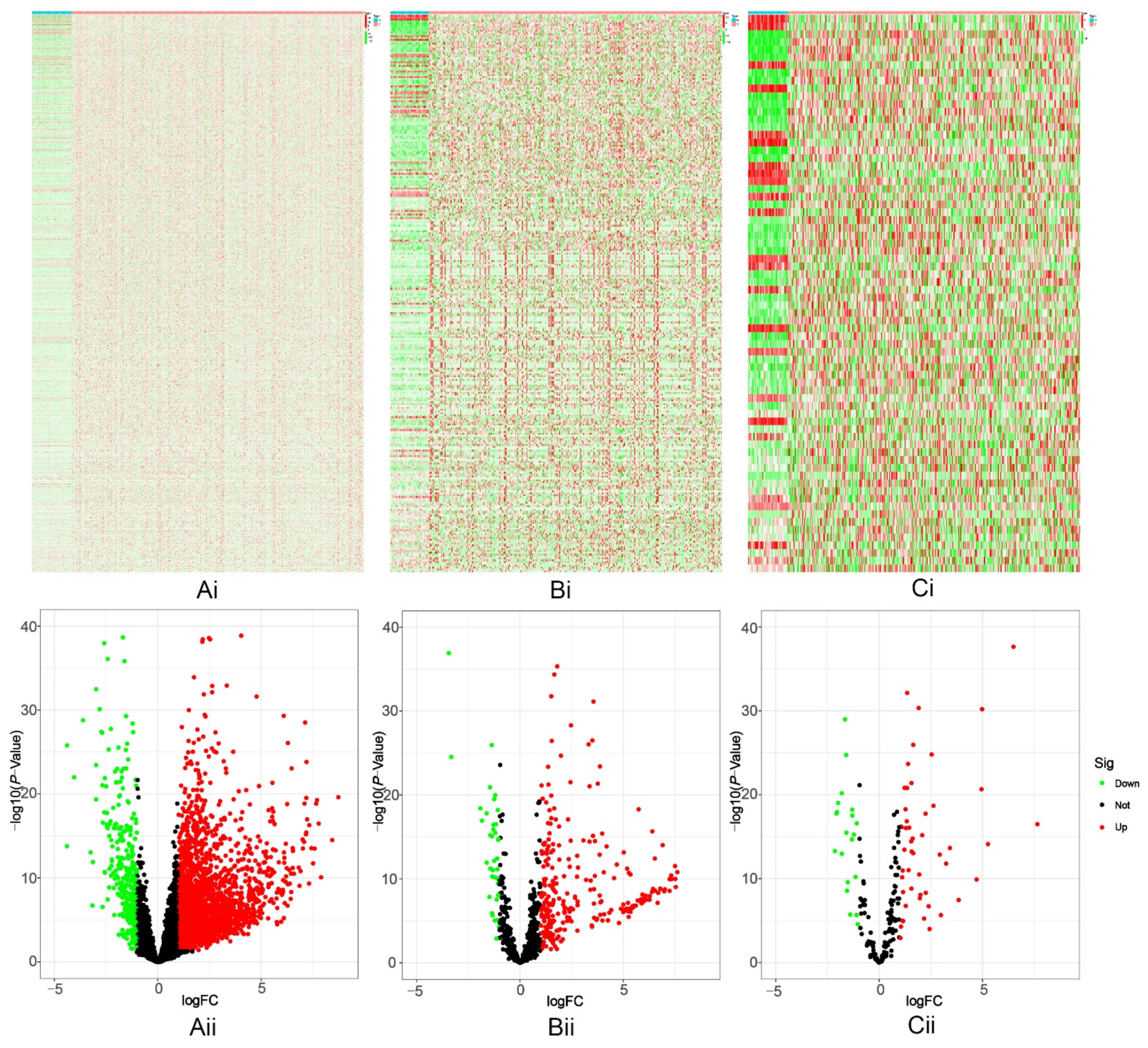
2.1. Screening of Differentially Expressed Genes
2.2. Screening of Prognosis-Related RNAs
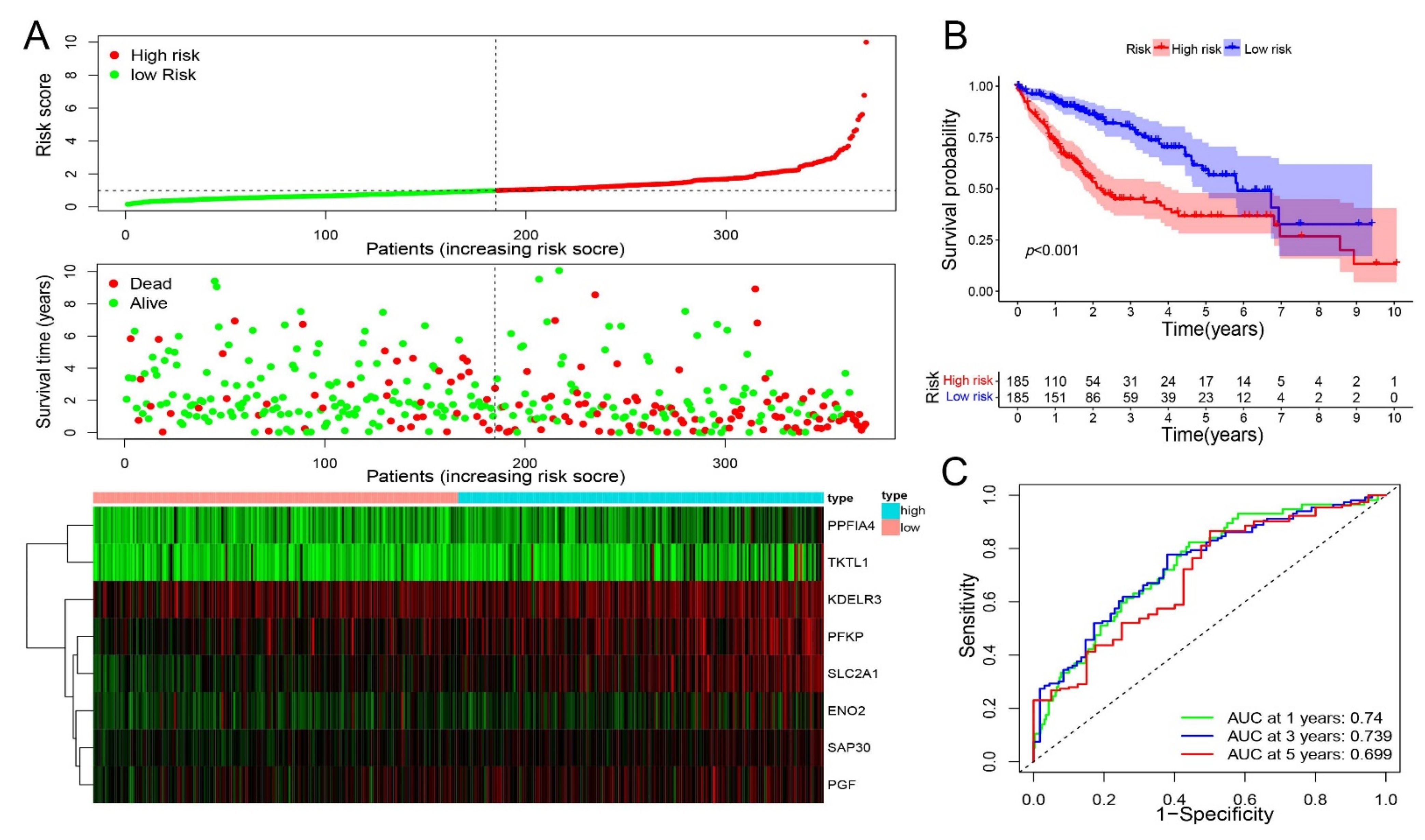
2.3. Construction of a Prognostic Hypoxia-Related Gene Signature
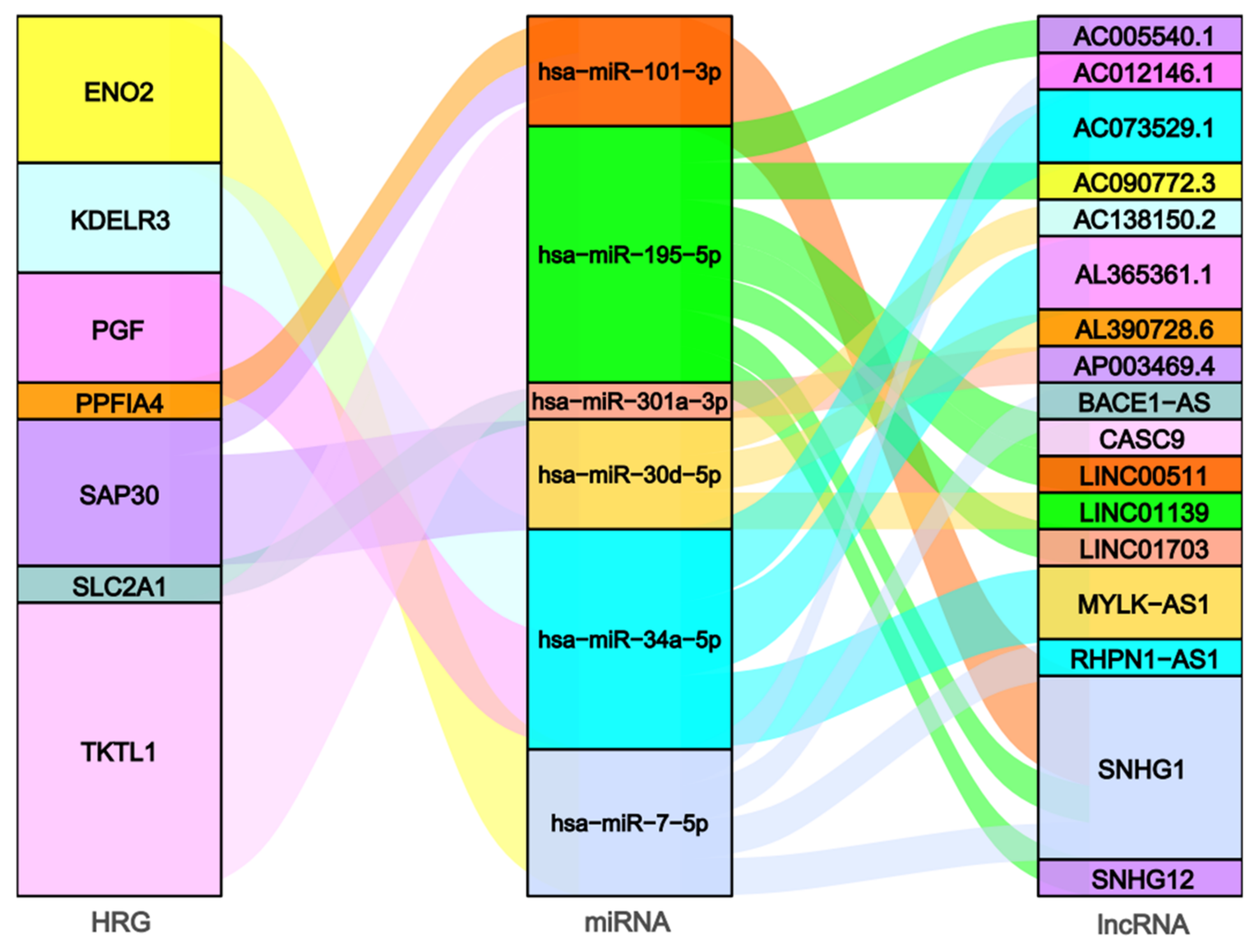
2.4. Construction of the Hypoxia-Related ceRNA Network
2.5. Correlation between ceRNA Network and Tumor Immune Infiltration
2.6. Drug Sensitivity Analysis
2.7. Identification of Potential lncRNA-miRNA-mRNA ceRNA Regulatory Axes
2.8. Gene Set Enrichment Analysis
3. Discussion
4. Materials and Methods
4.1. Data Download and Differential Expression Analysis
4.2. Screening of Prognosis-Related RNA by Univariate Cox Regression Analysis
4.3. Construction of Hypoxia-Related Gene Prognosis Signature by Multivariate Cox Regression Analysis
4.4. Construction of Hypoxia-Related ceRNA Network
4.5. Correlation Analysis between Hypoxic ceRNA Network and Tumor Immune Infiltration
4.6. Drug Sensitivity Analysis
4.7. Construction of lncRNA/miRNA/mRNA Regulatory Axes
4.8. Functional Enrichment Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikaesky, E.; Roayaie, S.; Lencioni, R.; Koike, K.; Rossi, J.; Finn, R.S. Hepatocellular carcinoma. Nat. Rev. Dis. Primers 2021, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, A. Hepatocellular Carcinoma. N. Engl. J. Med. 2019, 380, 1450–1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marrero, J.A.; Kulik, L.M.; Sirlin, C.B.; Zhu, A.X.; Finn, R.S.; Abecassis, M.M.; Roberts, L.R.; Heimbach, J.K. Diagnosis, Staging, and Management of Hepatocellular Carcinoma: 2018 Practice Guidance by the American Association for the Study of Liver Diseases. Hepatology 2018, 68, 723–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nault, J.C.; Villanueva, A. Biomarkers for Hepatobiliary Cancers. Hepatology 2021, 73 (Suppl. 1), 115–127. [Google Scholar] [CrossRef]
- Jing, X.; Yang, F.; Shao, C.; Wei, K.; Xie, M.; Shen, H.; Shu, Y. Role of hypoxia in cancer therapy by regulating the tumor microenvironment. Mol. Cancer 2019, 18, 157. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.Z.; Xie, G.R.; Chen, D. Hypoxia and hepatocellular carcinoma: The therapeutic target for hepatocellular carcinoma. J. Gastroenterol. Hepatol. 2007, 22, 1178–1182. [Google Scholar] [CrossRef]
- Erler, J.T.; Giaccia, A.J. Lysyl oxidase mediates hypoxic control of metastasis. Cancer Res. 2006, 66, 10238–10241. [Google Scholar] [CrossRef] [Green Version]
- Rohr-Udilova, N.; Klinglmuller, F.; Schulte-Hermann, R.; Stift, J.; Herac, M.; Salzmann, M.; Finotello, F.; Timelthaler, G.; Oberhuber, G.; Pinter, M.; et al. Deviations of the immune cell landscape between healthy liver and hepatocellular carcinoma. Sci. Rep. 2018, 8, 6220. [Google Scholar] [CrossRef] [Green Version]
- Palazon, A.; Goldrath, A.W.; Nizet, V.; Johnson, R.S. HIF transcription factors, inflammation, and immunity. Immunity 2014, 41, 518–528. [Google Scholar] [CrossRef]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta Stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Yang, P.; Feng, H.; Zhao, Q.; Liu, H. Using Network Distance Analysis to Predict lncRNA-miRNA Interactions. Interdiscip. Sci. 2021, 13, 535–545. [Google Scholar] [CrossRef]
- Qi, X.; Lin, Y.; Chen, J.; Shen, B. Decoding competing endogenous RNA networks for cancer biomarker discovery. Brief Bioinform. 2020, 21, 441–457. [Google Scholar] [CrossRef]
- Li, S.Y.; Zhu, Y.; Li, R.N.; Huang, J.H.; You, K.; Yuan, Y.F.; Zhuang, S.M. LncRNA Lnc-APUE is Repressed by HNF4alpha and Promotes G1/S Phase Transition and Tumor Growth by Regulating MiR-20b/E2F1 Axis. Adv. Sci. 2021, 8, 2003094. [Google Scholar] [CrossRef]
- Hu, J.J.; Zhou, C.; Luo, X.; Luo, S.Z.; Li, Z.H.; Xu, Z.X.; Xu, M.Y. Linc-SCRG1 accelerates progression of hepatocellular carcinoma as a ceRNA of miR26a to derepress SKP2. J. Exp. Clin. Cancer Res. 2021, 40, 26. [Google Scholar] [CrossRef]
- Wang, D.; Chen, F.; Zeng, T.; Tang, Q.; Chen, B.; Chen, L.; Dong, Y.; Li, X. Comprehensive biological function analysis of lncRNAs in hepatocellular carcinoma. Genes Dis. 2021, 8, 157–167. [Google Scholar] [CrossRef]
- Zhang, X.Z.; Liu, H.; Chen, S.R. Mechanisms of Long Non-Coding RNAs in Cancers and Their Dynamic Regulations. Cancers 2020, 12, 1245. [Google Scholar] [CrossRef]
- Amann, T.; Maegdefrau, U.; Hartmann, A.; Agaimy, A.; Marienhagen, J.; Weiss, T.S.; Stoeltzing, O.; Warnecke, C.; Scholmerich, J.; Oefner, P.J.; et al. GLUT1 expression is increased in hepatocellular carcinoma and promotes tumorigenesis. Am. J. Pathol. 2009, 174, 1544–1552. [Google Scholar] [CrossRef] [Green Version]
- Alpini, G.; Glaser, S.S.; Zhang, J.P.; Francis, H.; Han, Y.; Gong, J.; Stokes, A.; Francis, T.; Hughart, N.; Hubble, L.; et al. Regulation of placenta growth factor by microRNA-125b in hepatocellular cancer. J. Hepatol. 2011, 55, 1339–1345. [Google Scholar] [CrossRef] [Green Version]
- Vandewynckel, Y.P.; Laukens, D.; Devisscher, L.; Bogaerts, E.; Paridaens, A.; van den Bussche, A.; Raevens, S.; Verhelst, X.; van Steenkiste, C.; Jonckx, B.; et al. Placental growth factor inhibition modulates the interplay between hypoxia and unfolded protein response in hepatocellular carcinoma. BMC Cancer 2016, 16, 9. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, J.H.; Guo, C.K.; Cai, P.C. Gene silencing of TKTL1 by RNAi inhibits cell proliferation in human hepatoma cells. Cancer Lett. 2007, 253, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Liu, Y.; Glazer, C.A.; Shao, C.; Bhan, S.; Demokan, S.; Zhao, M.; Rudek, M.A.; Ha, P.K.; Califano, J.A. TKTL1 is activated by promoter hypomethylation and contributes to head and neck squamous cell carcinoma carcinogenesis through increased aerobic glycolysis and HIF1alpha stabilization. Clin. Cancer Res. 2010, 16, 857–866. [Google Scholar] [CrossRef] [Green Version]
- Sha, X.; Wang, K.; Wang, F.; Zhang, C.; Yang, L.; Zhu, X. Silencing PFKP restrains the stemness of hepatocellular carcinoma cells. Exp. Cell Res. 2021, 407, 112789. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.; Yang, D.; Hou, Y.; Liu, S.; Zhao, M.; Qin, Y.; Chen, R.; Teng, Y.; Liu, M. Intracellular citrate accumulation by oxidized ATM-mediated metabolism reprogramming via PFKP and CS enhances hypoxic breast cancer cell invasion and metastasis. Cell Death Dis. 2019, 10, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Y.; Qi, W.; Liu, L.; Zhang, J.; Pang, L.; Gan, T.; Wang, P.; Wang, C.; Chen, H. Identification of Seven-Gene Hypoxia Signature for Predicting Overall Survival of Hepatocellular Carcinoma. Front. Genet. 2021, 12, 637418. [Google Scholar] [CrossRef]
- Snezhkina, A.V.; Nyushko, K.M.; Zaretsky, A.R.; Shagin, D.A.; Sadritdinova, A.F.; Fedorova, M.S.; Guvatova, Z.G.; Abramov, I.S.; Pudova, E.A.; Alekseev, B.Y.; et al. Transcription Factor SAP30 Is Involved in the Activation of NETO2 Gene Expression in Clear Cell Renal Cell Carcinoma. Mol. Biol. 2018, 52, 451–459. [Google Scholar] [CrossRef]
- Chen, Z.; Zou, Y.; Zhang, Y.; Chen, Z.; Wu, F.; Shi, N.; Jin, H. A novel prognostic signature based on four glycolysis-related genes predicts survival and clinical risk of hepatocellular carcinoma. J. Clin. Lab. Anal. 2021, 35, e24005. [Google Scholar] [CrossRef]
- Zheng, Y.; Wu, C.; Yang, J.; Zhao, Y.; Jia, H.; Xue, M.; Xu, D.; Yang, F.; Fu, D.; Wang, C.; et al. Insulin-like growth factor 1-induced enolase 2 deacetylation by HDAC3 promotes metastasis of pancreatic cancer. Signal Transduct. Target Ther. 2020, 5, 53. [Google Scholar] [CrossRef]
- Huang, J.; Yang, M.; Liu, Z.; Li, X.; Wang, J.; Fu, N.; Cao, T.; Yang, X. PPFIA4 Promotes Colon Cancer Cell Proliferation and Migration by Enhancing Tumor Glycolysis. Front. Oncol. 2021, 11, 653200. [Google Scholar] [CrossRef]
- Ahmed, M.Y.; Salah, M.M.; Kassim, S.K.; Abdelaal, A.; Elayat, W.M.; Mohamed, D.A.; Fouly, A.E.; Abu-Zahra, F.A.E. Evaluation of the diagnostic and therapeutic roles of non-coding RNA and cell proliferation related gene association in hepatocellular carcinoma. Gene 2019, 706, 97–105. [Google Scholar] [CrossRef]
- Wan, Y.; Cui, R.; Gu, J.; Zhang, X.; Xiang, X.; Liu, C.; Qu, K.; Lin, T. Identification of Four Oxidative Stress-Responsive MicroRNAs, miR-34a-5p, miR-1915-3p, miR-638, and miR-150-3p, in Hepatocellular Carcinoma. Oxid. Med. Cell Longev. 2017, 2017, 5189138. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Ruan, J.; Liu, X.; Xiao, C.; Xiong, J. MicroRNA-301a-3p suppressed the progression of hepatocellular carcinoma via targeting VGLL4. Pathol. Res. Pract. 2018, 214, 2039–2045. [Google Scholar] [CrossRef]
- Zhuang, H.; Wu, F.; Wei, W.; Dang, Y.; Yang, B.; Ma, X.; Han, F.; Li, Y. Glycine decarboxylase induces autophagy and is downregulated by miRNA-30d-5p in hepatocellular carcinoma. Cell Death Dis. 2019, 10, 192. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Ma, L.; Zhai, L.; Chen, D.; Li, Y.; Shang, Z.; Zhang, Z.; Gao, Y.; Yang, W.; Li, Y.; et al. Construction and validation of a three-microRNA signature as prognostic biomarker in patients with hepatocellular carcinoma. Int. J. Med. Sci. 2021, 18, 984–999. [Google Scholar] [CrossRef]
- Xu, H.; Hu, Y.W.; Zhao, J.Y.; Hu, X.M.; Li, S.F.; Wang, Y.C.; Gao, J.J.; Sha, Y.H.; Kang, C.M.; Lin, L.; et al. MicroRNA-195-5p acts as an anti-oncogene by targeting PHF19 in hepatocellular carcinoma. Oncol. Rep. 2015, 34, 175–182. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.; Shan, X.; Chen, K.; Liu, Y.; Yu, G.; Chen, Q.; Zeng, T.; Zhu, L.; Dang, H.; Chen, F.; et al. LINC00052/miR-101-3p axis inhibits cell proliferation and metastasis by targeting SOX9 in hepatocellular carcinoma. Gene 2018, 679, 138–149. [Google Scholar] [CrossRef]
- Liu, C.; Wang, H.; Tang, L.; Huang, H.; Xu, M.; Lin, Y.; Zhou, L.; Ho, L.; Lu, J.; Ai, X. LncRNA BACE1-AS enhances the invasive and metastatic capacity of hepatocellular carcinoma cells through mediating miR-377-3p/CELF1 axis. Life Sci. 2021, 275, 119288. [Google Scholar] [CrossRef]
- Yao, J.; Fu, J.; Liu, Y.; Qu, W.; Wang, G.; Yan, Z. LncRNA CASC9 promotes proliferation, migration and inhibits apoptosis of hepatocellular carcinoma cells by down-regulating miR-424-5p. Ann. Hepatol. 2021, 23, 100297. [Google Scholar] [CrossRef]
- Hu, P.; Cui, H.; Lei, T.; Li, S.; Mai, E.; Jia, F. Linc00511 Indicates a Poor Prognosis Of Liver Hepatocellular Carcinoma. OncoTargets Ther. 2019, 12, 9367–9376. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.B.; Chu, H.T.; Jia, M.; Li, L. Long noncoding RNA LINC01139 promotes the progression of hepatocellular carcinoma by upregulating MYBL2 via competitively binding to miR-30 family. Biochem. Biophys. Res. Commun. 2020, 525, 581–588. [Google Scholar] [CrossRef]
- Teng, F.; Zhang, J.X.; Chang, Q.M.; Wu, X.B.; Tang, W.G.; Wang, J.F.; Feng, J.F.; Zhang, Z.P.; Hu, Z.Q. LncRNA MYLK-AS1 facilitates tumor progression and angiogenesis by targeting miR-424-5p/E2F7 axis and activating VEGFR-2 signaling pathway in hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2020, 39, 235. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yan, Z.; Wang, L.; Zhang, S.; Gao, M. STAT1-induced upregulation of lncRNA RHPN1-AS1 predicts a poor prognosis of hepatocellular carcinoma and contributes to tumor progression via the miR-485/CDCA5 axis. J. Cell. Biochem. 2020, 121, 4741–4755. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Li, A.; You, Z.; Xu, J.; Zhu, S. Epigenetic silencing of CDKN1A and CDKN2B by SNHG1 promotes the cell cycle, migration and epithelial-mesenchymal transition progression of hepatocellular carcinoma. Cell Death Dis. 2020, 11, 823. [Google Scholar] [CrossRef] [PubMed]
- Lan, T.; Ma, W.; Hong, Z.; Wu, L.; Chen, X.; Yuan, Y. Long non-coding RNA small nucleolar RNA host gene 12 (SNHG12) promotes tumorigenesis and metastasis by targeting miR-199a/b-5p in hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2017, 36, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Zhang, X.; Zhang, X.; Jiang, X.; Li, W. Long Noncoding RNA LINC01703 Exacerbates the Malignant Properties of Non-Small Cell Lung Cancer by Upregulating MACC1 in a MicroRNA-605-3p-Mediated Manner. Oncol. Res. 2022, 28, 913–927. [Google Scholar] [CrossRef]
- Cao, X.; Zhang, Q.; Zhu, Y.; Huo, X.; Bao, J.; Su, M. Derivation, Comprehensive Analysis, and Assay Validation of a Pyroptosis-Related lncRNA Prognostic Signature in Patients with Ovarian Cancer. Front. Oncol. 2022, 12, 780950. [Google Scholar] [CrossRef]
- Lv, Y.; Wei, W.; Huang, Z.; Chen, Z.; Fang, Y.; Pan, L.; Han, X.; Xu, Z. Long non-coding RNA expression profile can predict early recurrence in hepatocellular carcinoma after curative resection. Hepatol. Res. 2018, 48, 1140–1148. [Google Scholar] [CrossRef]
- Fan, T.; Jiang, G.; Shi, R.; Yu, R.; Xiao, X.; Ke, D. Construction of AP003469.4-miRNAs-mRNAs ceRNA network to reveal potential biomarkers for hepatocellular carcinoma. Am. J. Cancer Res. 2022, 12, 1484–1501. [Google Scholar]
- Schito, L.; Semenza, G.L. Hypoxia-Inducible Factors: Master Regulators of Cancer Progression. Trends Cancer 2016, 2, 758–770. [Google Scholar] [CrossRef] [Green Version]
- Abou Khouzam, R.; Goutham, H.V.; Zaarour, R.F.; Chamseddine, A.N.; Francis, A.; Buart, S.; Terry, S.; Chouaib, S. Integrating tumor hypoxic stress in novel and more adaptable strategies for cancer immunotherapy. Semin. Cancer Biol. 2020, 65, 140–154. [Google Scholar] [CrossRef]
- Zhang, Q.; Cheng, Q.; Xia, M.; Huang, X.; He, X.; Liao, J. Hypoxia-Induced lncRNA-NEAT1 Sustains the Growth of Hepatocellular Carcinoma via Regulation of miR-199a-3p/UCK2. Front. Oncol. 2020, 10, 998. [Google Scholar] [CrossRef]
- Hu, G.; Ma, J.; Zhang, J.; Chen, Y.; Liu, H.; Huang, Y.; Zheng, J.; Xu, Y.; Xue, W.; Zhai, W. Hypoxia-induced lncHILAR promotes renal cancer metastasis via ceRNA for the miR-613/206/ 1-1-3p/Jagged-1/Notch/CXCR4 signaling pathway. Mol. Ther. 2021, 29, 2979–2994. [Google Scholar] [CrossRef]
- Prieto, J.; Melero, I.; Sangro, B. Immunological landscape and immunotherapy of hepatocellular carcinoma. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 681–700. [Google Scholar] [CrossRef]
- Lurje, I.; Hammerich, L.; Tacke, F. Dendritic Cell and T Cell Crosstalk in Liver Fibrogenesis and Hepatocarcinogenesis: Implications for Prevention and Therapy of Liver Cancer. Int. J. Mol. Sci. 2020, 21, 7378. [Google Scholar] [CrossRef]
- Qin, M.; Wang, D.; Fang, Y.; Zheng, Z.; Liu, X.; Wu, F.; Wang, L.; Li, X.; Hui, B.; Ma, S.; et al. Current Perspectives on B Lymphocytes in the Immunobiology of Hepatocellular Carcinoma. Front. Oncol. 2021, 11, 647854. [Google Scholar] [CrossRef]
- Dangaj, D.; Bruand, M.; Grimm, A.J.; Ronet, C.; Barras, D.; Duttagupta, P.A.; Lanitis, E.; Duraiswamy, J.; Tanyi, J.L.; Benencia, F.; et al. Cooperation between Constitutive and Inducible Chemokines Enables T Cell Engraftment and Immune Attack in Solid Tumors. Cancer Cell 2019, 35, 885–900.e10. [Google Scholar] [CrossRef]
- Xu, F.; Jin, T.; Zhu, Y.; Dai, C. Immune checkpoint therapy in liver cancer. J. Exp. Clin. Cancer Res. 2018, 37, 110. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Dai, W.; Mao, Y.; Wu, L.; Li, J.; Chen, K.; Yu, Q.; Kong, R.; Li, S.; Zhang, J.; et al. Simvastatin re-sensitizes hepatocellular carcinoma cells to sorafenib by inhibiting HIF-1α/PPAR-γ/PKM2-mediated glycolysis. J. Exp. Clin. Cancer Res. 2020, 39, 24. [Google Scholar] [CrossRef] [Green Version]
- Xiang, Q.; Chen, W.; Ren, M.; Wang, J.; Zhang, H.; Deng, D.Y.; Zhang, L.; Shang, C.; Chen, Y. Cabozantinib suppresses tumor growth and metastasis in hepatocellular carcinoma by a dual blockade of VEGFR2 and MET. Clin. Cancer Res. 2014, 20, 2959–2970. [Google Scholar] [CrossRef] [Green Version]
- Chang, A.Y.; Wang, M. Molecular mechanisms of action and potential biomarkers of growth inhibition of dasatinib (BMS-354825) on hepatocellular carcinoma cells. BMC Cancer 2013, 13, 267. [Google Scholar] [CrossRef] [Green Version]
- Yue, D.; Sun, X. Idelalisib promotes Bim-dependent apoptosis through AKT/FoxO3a in hepatocellular carcinoma. Cell Death Dis. 2018, 9, 935. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Yang, N.; Sun, L.; Zhang, L.; Li, Z.; Li, D.; Qin, T.; Huang, H. LncRNA SNHG1 upregulates ROCK2 to reduce cisplatin sensitivity of NSCLC cells by targeting miR-101-3p. Transl. Cancer Res. 2019, 8, 2141–2150. [Google Scholar] [CrossRef] [PubMed]
- Liberzon, A.; Birger, C.; Thorvaldsdóttir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database (MSigDB) hallmark gene set collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Li, J.H.; Liu, S.; Zhou, H.; Qu, L.H.; Yang, J.H. starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data. Nucleic Acids Res. 2014, 42, D92–D97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S.; Li, B.; Liu, X.S. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017, 77, e108–e110. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, w98–w102. [Google Scholar] [CrossRef] [Green Version]
- Reinhold, W.C.; Sunshine, M.; Liu, H.; Varma, S.; Kohn, K.W.; Morris, J.; Doroshow, J.; Pommier, Y. CellMiner: A web-based suite of genomic and pharmacologic tools to explore transcript and drug patterns in the NCI-60 cell line set. Cancer Res. 2012, 72, 3499–3511. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RNA | Expression Change | Name | p-adj. |
|---|---|---|---|
| lncRNA | Upregulated | MYLK-AS1 | 5.61 × 10−25 |
| SNHG1 | 2.77 × 10−22 | ||
| BACE1-AS | 7.27 × 10−20 | ||
| LINC01703 | 9.28 × 10−18 | ||
| LINC00511 | 4.58 × 10−14 | ||
| SNHG12 | 1.48 × 10−13 | ||
| AP003469.4 | 6.04 × 10−13 | ||
| AC012146.1 | 7.44 × 10−12 | ||
| AL390728.6 | 7.55 × 10−11 | ||
| RHPN1-AS1 | 5.45 × 10−10 | ||
| AC073529.1 | 4.16 × 10−9 | ||
| AC090772.3 | 1.87 × 10−7 | ||
| LINC01139 | 5.27 × 10−7 | ||
| CASC9 | 1.98 × 10−6 | ||
| AC138150.2 | 3.77 × 10−6 | ||
| AC005540.1 | 1.36 × 10−3 | ||
| Downregulated | AL365361.1 | 2.35 × 10−7 | |
| miRNA | Upregulated | hsa-miR-34a-5p | 3.89 × 10−16 |
| hsa-miR-30d-5p | 5.86 × 10−14 | ||
| hsa-miR-301a-3p | 3.08 × 10−13 | ||
| hsa-miR-7-5p | 8.75 × 10−5 | ||
| Downregulated | hsa-miR-101-3p | 1.46 × 10−17 | |
| hsa-miR-195-5p | 5.78 × 10−16 | ||
| mRNA | Upregulated | SAP30 | 4.37 × 10−16 |
| PGF | 1.60 × 10−14 | ||
| PPFIA4 | 5.01 × 10−13 | ||
| KDELR3 | 3.10 × 10−11 | ||
| TKTL1 | 3.57 × 10−10 | ||
| SLC2A1 | 4.02 × 10−9 | ||
| ENO2 | 1.88 × 10−5 |
| mRNA | Feature | Abstract | Citation |
|---|---|---|---|
| SLC2A1 |  | Through cell culture in vitro and immunohistochemical detection, Amann et al. showed that SLC2ZA1 was expressed at high levels and its overexpression promoted the growth and invasion of HCC. The knockdown of SLC2ZA1 inhibited glucose uptake and lactic acid secretion, and deactivated hypoxia transcription factor factor-1 (HIF1) under hypoxic conditions, thereby inhibiting the proliferation and metastasis of hepatocytes. | [18] |
| PGF |  | Alpini et al. showed by immunohistochemistry that PGF (PIGF) protein was notably higher in HCC tissues compared with normal tissues (p < 0.01). PIGF might recruit cyclic hematopoietic progenitor cells and macrophages into a growing tumor to induce tumor angiogenesis, which was associated with the early recurrence of HCC. Through quantitative real-time PCR (qRT-PCR) and immunohistochemistry, Vandewynckel et al. showed that inhibition of PGF deactivated pancreatic endoplasmic reticulum kinase, and further induced tumor angiogenesis to improve oxygen delivery, thus reducing tumor hypoxia in HCC. | [19,20] |
| TKTL1 |  | Zhang et al. showed via cell-based experiments that TKTL1 was overexpressed in human HEPG2 hepatoma cells. Silencing TKTL1 via siRNA, the total transketolase activity of HEPG2 cells was decreased and the proliferative activity was inhibited. Sun et al. showed that overexpressed TKTL1 participated in the occurrence and progression of Head and Neck squamous cell carcinoma by promoting aerobic glycolysis and accumulating HIF1α. | [21,22] |
| PFKP |  | Through western blotting and immunohistochemical experiments, Sha et al. showed that PFKP were upregulated in HCC and its upregulation promoted cancer cell proliferation. Peng et al. showed that hypoxia (1% O2) induced oxidized ATM activation which increased the levels of PFKP and citrate synthase, promoted glucose to be converted into pyruvic acid and citrate, and enhanced the invasion and metastasis of hypoxic breast cancer cells. | [23,24] |
| KDELR3 |  | Bai et al. showed that overexpression of KDELR3 was correlated with poor prognosis by immunohistochemical experiments and real-time fluorescence quantitative PCR (RT-qPCR) analysis and KDELR3 could be a potential therapeutic target in hepatocytes and liver cancer tissues. | [25] |
| SAP30 |   | Snezhkina et al. showed that SAP30 was overexpressed in renal clear cell carcinoma (ccRCC) by qPCR analysis. The overexpression of SAP30 was positively associated with the expression of neuropilin and tolloid-like 2 (NETO2) gene and might activate NETO2 to cause tumor progression and poor prognosis of ccRCC. On the other hand, Chen et al. reported the upregulated expression of SAP30 in HCC via bioinformatics analysis and further constructed a glycolysis-related prognosis model including SAP30 which was connected to the poor prognosis of HCC patients. | [26,27] |
| ENO2 |  | Zheng et al. showed by immunohistochemistry staining and western blotting that the expression of ENO2 was significantly higher in pancreatic ductal adenocarcinoma (PDAC) than in normal tissues. | [28] |
| PPFIA4 |  | Huang et al. reported that the expression level of PPFIA4 was higher in Colon adenocarcinoma cells than in normal cells by qRT-PCR. | [29] |
 the mRNA experimentally reported to be upregulated in HCC in accord with our prediction in HCC.
the mRNA experimentally reported to be upregulated in HCC in accord with our prediction in HCC.  the mRNA computationally reported to be upregulated in HCC in accord with our prediction in HCC.
the mRNA computationally reported to be upregulated in HCC in accord with our prediction in HCC.  the mRNA experimentally reported to be upregulated in other cancers, and its expression in accord with our prediction in hepatocellular carcinoma.
the mRNA experimentally reported to be upregulated in other cancers, and its expression in accord with our prediction in hepatocellular carcinoma.| miRNA | Feature | Abstract | Citation |
|---|---|---|---|
| hsa-miR-301a-3p |  | Hu et al. showed by western blot and Reverse Transcription-PCR (RT-PCR) that the expression of miR-301a-3p in HCC tissues was notably higher than that in normal tissues. The overexpression of miR-301a-3p downregulated the target gene VGLL4, and then enhanced the transcription activity of TEADs, promoting the proliferation, invasion, and chemoresistance of HCC cell. | [32] |
| hsa-miR-30d-5p |  | Zhuang et al. demonstrated that miR-30d-5p was upregulated in hepatocellular carcinoma HCCLM3 and MHCC97L cell lines compared with the normal cell line Huh7 (p < 0.005) by cell culture and transfection, qRT-PCR and Immunohistochemistry (IHC) staining. Silencing miR-30d-5p upregulated the target gene GLDC, decreased p62 expression and further induced cell autophagy, thus suppressing the progression and invasiveness of HCC cells. | [33] |
| hsa-miR-7-5p |  | Through quantity PCR (qPCR) analysis, Zhang et al. showed that the expression of plasma hsa-miR-7-5p in HCC patients was significantly increased compared with normal and benign groups, and further constructed a prognostic model including the overexpression of miR-7-5p which was related to a poor diagnosis of HCC patients. | [34] |
| hsa-miR-195-5p |  | Xu et al. revealed via cell culture, tumor xenograft experiments and RT-qPCR analysis that miR-195-5p was expressed at low levels in HCC tissues, and upregulation of miR-195-5p directly downregulated PHF19 and thus inhibited the proliferation, invasion, and migration of HCC cells in vitro and the growth of xenograft HCC tumors in nude mice. | [35] |
| hsa-miR-101-3p |  | Yan et al. demonstrated by qRT-PCR and in vitro cell culture experiments that miR-101-3p was down-expressed in both HCC tissues and cell lines, and the linker LINC00052 could promote its expression and further down-regulate the miRNA target SOX9, thus inhibiting the proliferation and metastasis of HCC cells. | [36] |
| hsa-miR-34a-5p |   | Ahmed et al. detected by RT-qPCR analysis that miR-34a-5p was significantly down-expressed in HCC than in normal liver tissues, and its downregulation increased the expression level of gene MCM2 to promote HCC development. However, Wan et al. revealed via HCC cell line culture that miR-34a-5p, an oxidative stress-responsive related miRNA, was upregulated in four HCC cell lines with oxidative stress models compared with normal cell lines. | [30,31] |
 the miRNA experimentally reported to be upregulated in HCC in accord with our prediction in HCC.
the miRNA experimentally reported to be upregulated in HCC in accord with our prediction in HCC.  the miRNA experimentally reported to be downregulated in HCC in accord with our prediction in HCC.
the miRNA experimentally reported to be downregulated in HCC in accord with our prediction in HCC.  the miRNA experimentally reported to be down-regulated in HCC which was different from prediction in HCC.
the miRNA experimentally reported to be down-regulated in HCC which was different from prediction in HCC.| lncRNA | Feature | Abstract | Citation |
|---|---|---|---|
| BACE1-AS |  | Liu et al. showed that BACE1-AS was overexpressed in HCC tissues by qRT-PCR analysis. Knocking out BACE1-AS upregulated the expression of MIR-377-3P, and deactivated epithelial–mesenchymal transition (EMT) process, inhibiting the invasion and migration of HCC cells. | [37] |
| CASC9 |  | Yao et al. showed via qRT-PCR and western blot analysis that CASC9 was expressed at high levels in HCC tissues and positively associated with tumor node metastasis (TNM) stage, lymph node metastasis (LNM), tumor size, differentiation degree and α-fetoprotein (AFP). Downregulated CASC9 upregulated the expression level of miR-424-5p, enhanced apoptosis and decreased the proliferation, invasion and migration of HCC cells. | [38] |
| LINC00511 |  | Through qRT-PCR analysis, Hu et al. showed that LINC00511 was up-regulated in HCC tissues and positively correlated with TNM stage, lymphatic metastasis, and poor prognosis. Knockdown of LINC00511 upregulated the level of miR-29c and attenuated the proliferation and migration of Huh7 and Hep3B cells. | [39] |
| LINC01139 |  | Li et al. showed via cell transfection and RT-PCR analysis that the expression of LINC01139 in HCC tissues was significantly increased than that in normal liver tissues. LINC01139 could downregulate miR-30 via acting as a sponge, and further upregulate MYBL2, accordingly aggravating the cancer progression and might serve as a prognostic biomarker for HCC patients. | [40] |
| MYLK-AS1 |  | Through qRT-PCR and dual luciferase reporter analysis, Teng et al. detected that the overexpression of MYLK-AS1 was frequent in both HCC tumor tissues and cell lines. MYLK-AS1 overexpression directly downregulated miR-424-5p, upregulated E2F7 and further activated VEGFR-2 signaling, thus promoting the HCC progression. | [41] |
| RHPN1-AS1 |  | Zhang et al. showed that RHPN1-AS1 expression was elevated in both HCC tissues and cell lines by RT-PCR analysis and cell culture, respectively. Transcription factor STAT1-induced overexpression of RHPN1-AS1 stimulated CDCA5 expressions, further downregulated the expression level of the sponged miR-485 and then promoted the proliferation and metastasis of HCC cells. | [42] |
| SNHG1 |  | Li et al. determined via qRT-PCR and in vitro cell-based experiments that SNHG1 was overexpressed in both HCC tissues and cells. Knockdown of SNHG1 epigenetically increased CDKN2B and CDKN1A in the nucleus and repressed the expression of CDK4 by upregulating the sponged miR-140-5p in the cytoplasm, thus inhibiting cell cycle, growth, metastasis, and EMT process of HCC cells. | [43] |
| SNHG12 |  | Through qRT-PCR analysis and in vitro cell-based experiments, Lan et al. detected that SNHG12 was expressed at higher levels in HCC. Abnormally high expression of SNHG12 downregulated the expression of miR-199a/b-5p, caused the upregulation of gene MLK3, and promoted the progression and metastasis of tumor cells by affecting the NF-κB signaling pathway. | [44] |
| LINC01703 |  | Wang et al. demonstrated via analyses of qRT-PCR and in vitro cell-based experiments that LINC01703 was over-expressed in NSCLC. The upregulation of LINC01703 expression downregulated the sponged miR-605-3p, and promoted the expression of MACC1, subsequently increasing the invasiveness of cancer cells. | [45] |
| AL365361.1 |   | Cao et al. showed that AL365361.1 expression was downregulation in Ovarian cancer patient samples by RT-PCR analysis. Lv et al. showed in a data mining study that down-regulation of AL365361.1 and the other six abnormally expressed lncRNAs formed a lncRNA-based classifier which could efficiently predict the early recurrence after operating resection for HCC. | [46,47] |
| AP003469.4 |  | Fan et al. showed that AP003469.4 was expressed at high levels in HCC by data mining from TCGA and GEO. Up-regulation of AP003469.4 was a biomarker of diagnosis and dismal prognosis in HCC patients (Diagnostic ROC = 0.9048). | [48] |
| AC005540.1 AC012146.1 AC073529.1 AC090772.3 AC138150.2 AL390728.6 |  | In this study, AC005540.1, AC012146.1, AC073529.1, AC090772.3, AC138150.2, and AL390728.6, which are calculated to be upregulated in HCC, were associated with a poor prognosis. The over-expression of these lncRNAs has not been reported in any cancer studies. | \ |
 the lncRNA experimentally reported to be upregulated in HCC in accord with our prediction in HCC.
the lncRNA experimentally reported to be upregulated in HCC in accord with our prediction in HCC.  the lncRNA experimentally reported to be upregulated in other cancers in accord with our prediction in HCC.
the lncRNA experimentally reported to be upregulated in other cancers in accord with our prediction in HCC.  the lncRNAs upregulated in our prediction in HCC which have not been experimentally reported in cancers.
the lncRNAs upregulated in our prediction in HCC which have not been experimentally reported in cancers.  the lncRNA experimentally reported to be downregulated in other cancers in accord with our prediction in HCC.
the lncRNA experimentally reported to be downregulated in other cancers in accord with our prediction in HCC.  the lncRNA downregulated in our prediction in HCC which has not been experimentally verified in cancers.
the lncRNA downregulated in our prediction in HCC which has not been experimentally verified in cancers.Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Y.; Zhang, H.; Chen, L.; Zhang, T.; Xu, N.; Huang, Z. Identification of Hypoxia-Related Prognostic Signature and Competing Endogenous RNA Regulatory Axes in Hepatocellular Carcinoma. Int. J. Mol. Sci. 2022, 23, 13590. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113590
Tang Y, Zhang H, Chen L, Zhang T, Xu N, Huang Z. Identification of Hypoxia-Related Prognostic Signature and Competing Endogenous RNA Regulatory Axes in Hepatocellular Carcinoma. International Journal of Molecular Sciences. 2022; 23(21):13590. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113590
Chicago/Turabian StyleTang, Yulai, Hua Zhang, Lingli Chen, Taomin Zhang, Na Xu, and Zunnan Huang. 2022. "Identification of Hypoxia-Related Prognostic Signature and Competing Endogenous RNA Regulatory Axes in Hepatocellular Carcinoma" International Journal of Molecular Sciences 23, no. 21: 13590. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113590