
Insights into Antiviral Properties and Molecular Mechanisms of Non-Flavonoid Polyphenols against Human Herpesviruses



Abstract
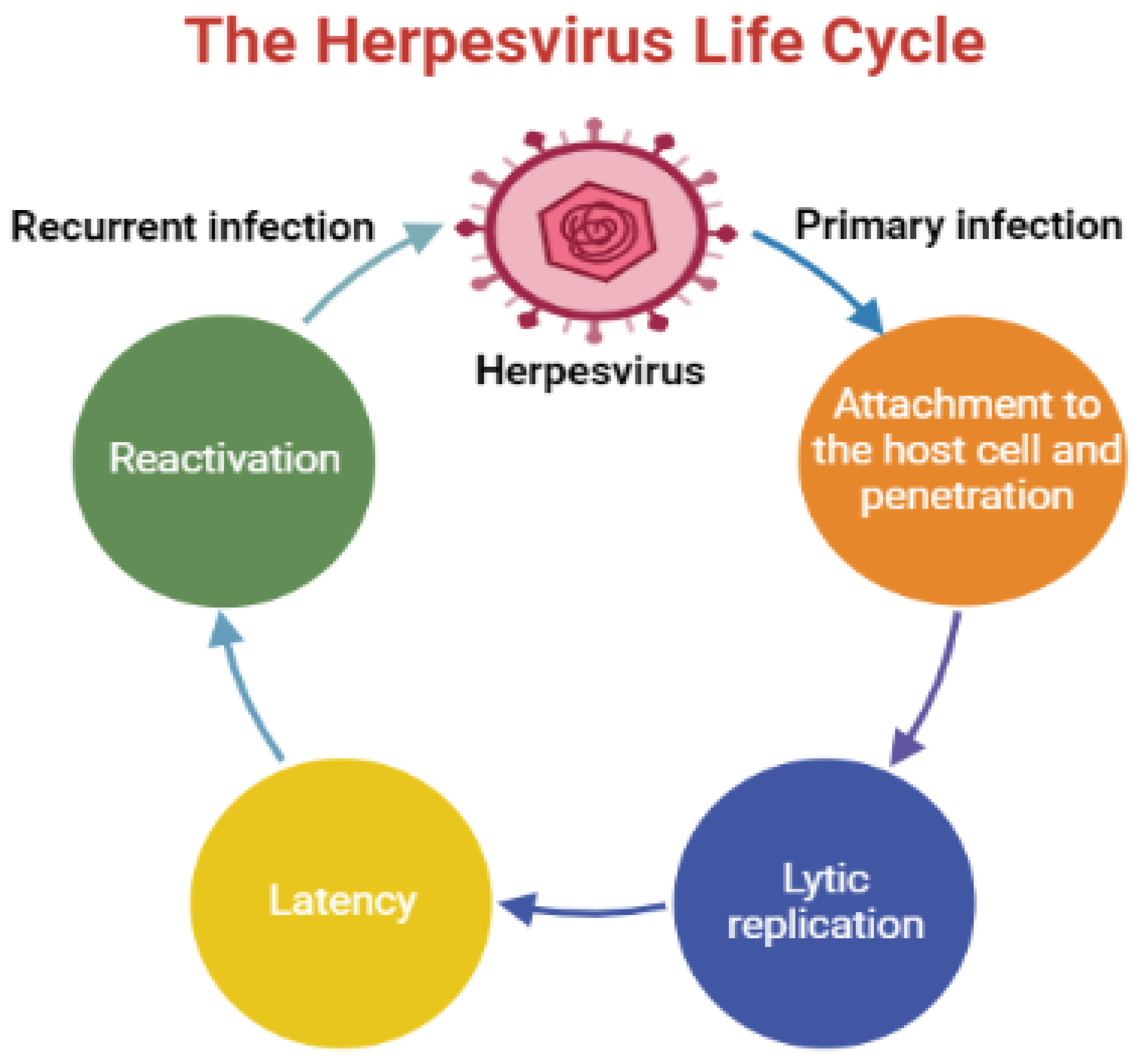
:1. Introduction
2. An Overview of Polyphenols and Their Health Benefits as Antivirals
3. Antiviral Properties of Non-Flavonoid Polyphenols against Alpha-Herpesviruses
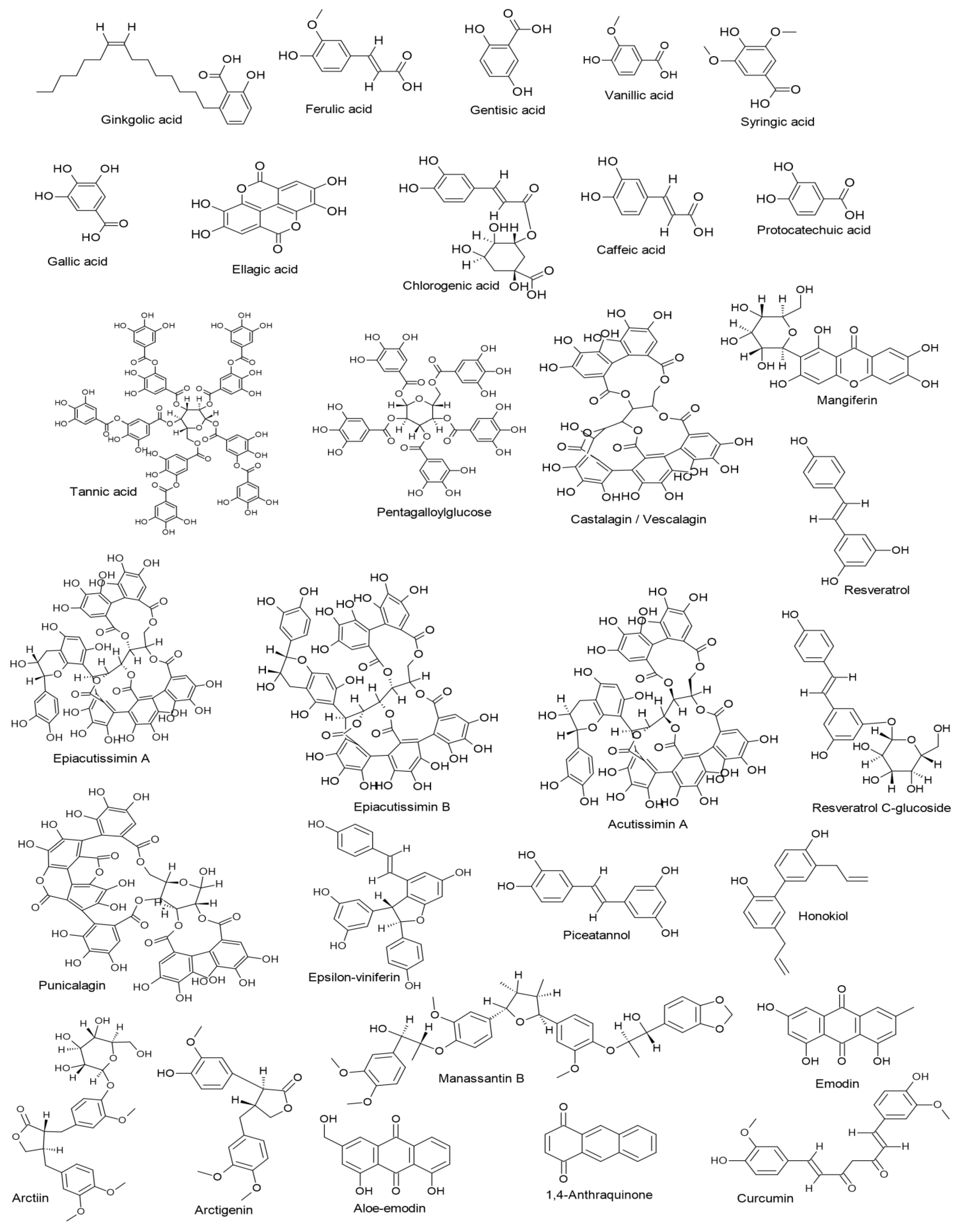
3.1. Phenolic Acids
3.2. Tannins and Their Derivatives
3.3. Xanthones
3.4. Stilbenes and Their Derivatives
3.5. Lignans and Neolignans
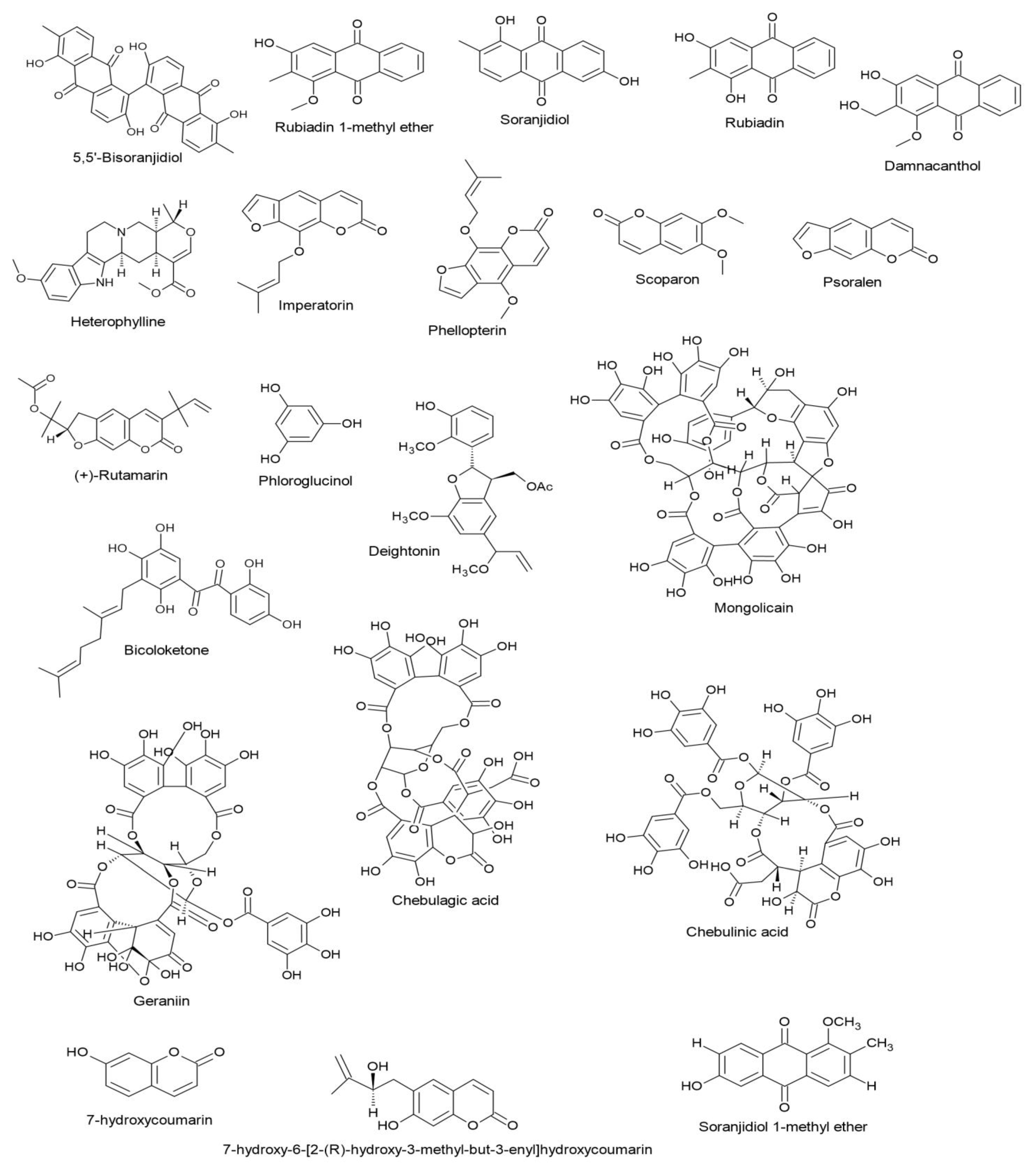
3.6. Anthraquinones and Their Derivatives
3.7. Curcuminoids
3.8. Coumarins
3.9. Other Polyphenols: Phloroglucinol
4. Antiviral Properties of Non-Flavonoid Polyphenols against Beta-Herpesviruses (Human Cytomegalovirus)
4.1. Phenolic Acids
4.2. Stilbenes
4.3. Anthraquinones
4.4. Curcuminoids
5. Antiviral Properties of Non-Flavonoid Polyphenols against Gamma-Herpesviruses
5.1. Phenolic Acids
5.2. Stilbenes
5.3. Lignans
5.4. Anthraquinones
5.5. Curcuminoids
5.6. Coumarins and Furanocoumarins
6. Mechanisms of Action of Non-Flavonoid Polyphenols: A Focus Insight
7. Conclusions and Take-Home Message
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Šudomová, M.; Berchová-Bímová, K.; Mazurakova, A.; Šamec, D.; Kubatka, P.; Hassan, S.T.S. Flavonoids Target Human Herpesviruses That Infect the Nervous System: Mechanisms of Action and Therapeutic Insights. Viruses 2022, 14, 592. [Google Scholar] [CrossRef] [PubMed]
- Šudomová, M.; Berchová-Bímová, K.; Marzocco, S.; Liskova, A.; Kubatka, P.; Hassan, S.T.S. Berberine in Human Oncogenic Herpesvirus Infections and Their Linked Cancers. Viruses 2021, 13, 1014. [Google Scholar] [CrossRef] [PubMed]
- Treml, J.; Gazdová, M.; Šmejkal, K.; Šudomová, M.; Kubatka, P.; Hassan, S.T.S. Natural Products-Derived Chemicals: Breaking Barriers to Novel Anti-HSV Drug Development. Viruses 2020, 12, 154. [Google Scholar] [CrossRef] [Green Version]
- Adler, B.; Sattler, C.; Adler, H. Herpesviruses and Their Host Cells: A Successful Liaison. Trends Microbiol. 2017, 25, 229–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šudomová, M.; Hassan, S.T.S. Nutraceutical Curcumin with Promising Protection against Herpesvirus Infections and Their Associated Inflammation: Mechanisms and Pathways. Microorganisms 2021, 9, 292. [Google Scholar] [CrossRef]
- Wołącewicz, M.; Becht, R.; Grywalska, E.; Niedźwiedzka-Rystwej, P. Herpesviruses in Head and Neck Cancers. Viruses 2020, 12, 172. [Google Scholar] [CrossRef] [Green Version]
- Azab, W.; Osterrieder, K. Initial Contact: The First Steps in Herpesvirus Entry. Adv. Anat. Embryol. Cell Biol. 2017, 223, 1–27. [Google Scholar] [CrossRef]
- Connolly, S.A.; Jardetzky, T.S.; Longnecker, R. The Structural Basis of Herpesvirus Entry. Nat. Rev. Microbiol. 2021, 19, 110–121. [Google Scholar] [CrossRef]
- Cohen, J.I. Herpesvirus Latency. J. Clin. Investig. 2020, 130, 3361–3369. [Google Scholar] [CrossRef]
- Wu, Y.; Yang, Q.; Wang, M.; Chen, S.; Jia, R.; Yang, Q.; Zhu, D.; Liu, M.; Zhao, X.; Zhang, S.; et al. Multifaceted Roles of ICP22/ORF63 Proteins in the Life Cycle of Human Herpesviruses. Front. Microbiol. 2021, 12, 668461. [Google Scholar] [CrossRef]
- Frappier, L. Regulation of Herpesvirus Reactivation by Host MicroRNAs. J. Virol. 2015, 89, 2456–2458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dochnal, S.A.; Francois, A.K.; Cliffe, A.R. De Novo Polycomb Recruitment: Lessons from Latent Herpesviruses. Viruses 2021, 13, 1470. [Google Scholar] [CrossRef] [PubMed]
- Jarosinski, K.W. Interindividual Spread of Herpesviruses. Adv. Anat. Embryol. Cell Biol. 2017, 223, 195–224. [Google Scholar] [CrossRef] [PubMed]
- Poole, C.L.; James, S.H. Antiviral Therapies for Herpesviruses: Current Agents and New Directions. Clin. Ther. 2018, 40, 1282–1298. [Google Scholar] [CrossRef] [Green Version]
- Majewska, A.; Mlynarczyk-Bonikowska, B. 40 Years after the Registration of Acyclovir: Do We Need New Anti-Herpetic Drugs? Int. J. Mol. Sci. 2022, 23, 3431. [Google Scholar] [CrossRef]
- Kłysik, K.; Pietraszek, A.; Karewicz, A.; Nowakowska, M. Acyclovir in the Treatment of Herpes Viruses—A Review. Curr. Med. Chem. 2020, 27, 4118–4137. [Google Scholar] [CrossRef]
- Hassan, S.T.S.; Masarčíková, R.; Berchová, K. Bioactive Natural Products with Anti-Herpes Simplex Virus Properties. J. Pharm. Pharmacol. 2015, 67, 1325–1336. [Google Scholar] [CrossRef]
- Hassan, S.T.S.; Švajdlenka, E.; Berchová-Bímová, K. Hibiscus sabdariffa L. and Its Bioactive Constituents Exhibit Antiviral Activity against HSV-2 and Anti-Enzymatic Properties against Urease by an ESI-MS Based Assay. Molecules 2017, 22, 722. [Google Scholar] [CrossRef]
- Lattanzio, V. Phenolic Compounds: Introduction. In Natural Products; Ramawat, K.G., Mérillon, J.-M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1543–1580. ISBN 978-3-642-22143-9. [Google Scholar]
- Wang, X.; Qi, Y.; Zheng, H. Dietary Polyphenol, Gut Microbiota, and Health Benefits. Antioxidants 2022, 11, 1212. [Google Scholar] [CrossRef]
- Durazzo, A.; Lucarini, M.; Souto, E.B.; Cicala, C.; Caiazzo, E.; Izzo, A.A.; Novellino, E.; Santini, A. Polyphenols: A Concise Overview on the Chemistry, Occurrence, and Human Health. Phytother. Res. 2019, 33, 2221–2243. [Google Scholar] [CrossRef]
- Tuladhar, P.; Sasidharan, S.; Saudagar, P. Role of Phenols and Polyphenols in Plant Defense Response to Biotic and Abiotic Stresses. In Biocontrol Agents and Secondary Metabolites; Elsevier: Amsterdam, The Netherlands, 2021; pp. 419–441. ISBN 978-0-12-822919-4. [Google Scholar]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The Role of Polyphenols in Human Health and Food Systems: A Mini-Review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and Human Health: The Role of Bioavailability. Nutrients 2021, 13, 273. [Google Scholar] [CrossRef]
- Luca, S.V.; Macovei, I.; Bujor, A.; Miron, A.; Skalicka-Woźniak, K.; Aprotosoaie, A.C.; Trifan, A. Bioactivity of Dietary Polyphenols: The Role of Metabolites. Crit. Rev. Food Sci. Nutr. 2020, 60, 626–659. [Google Scholar] [CrossRef]
- Zhang, L.; Han, Z.; Granato, D. Polyphenols in Foods: Classification, Methods of Identification, and Nutritional Aspects in Human Health. Adv. Food Nutr. Res. 2021, 98, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Chojnacka, K.; Skrzypczak, D.; Izydorczyk, G.; Mikula, K.; Szopa, D.; Witek-Krowiak, A. Antiviral Properties of Polyphenols from Plants. Foods 2021, 10, 2277. [Google Scholar] [CrossRef] [PubMed]
- Montenegro-Landívar, M.F.; Tapia-Quirós, P.; Vecino, X.; Reig, M.; Valderrama, C.; Granados, M.; Cortina, J.L.; Saurina, J. Polyphenols and Their Potential Role to Fight Viral Diseases: An Overview. Sci. Total Environ. 2021, 801, 149719. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.T.S.; Šudomová, M.; Berchová-Bímová, K.; Šmejkal, K.; Echeverría, J. Psoromic Acid, a Lichen-Derived Molecule, Inhibits the Replication of HSV-1 and HSV-2, and Inactivates HSV-1 DNA Polymerase: Shedding Light on Antiherpetic Properties. Molecules 2019, 24, 2912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Viejo-Borbolla, A. Pathogenesis and Virulence of Herpes Simplex Virus. Virulence 2021, 12, 2670–2702. [Google Scholar] [CrossRef]
- Gershon, A.A.; Breuer, J.; Cohen, J.I.; Cohrs, R.J.; Gershon, M.D.; Gilden, D.; Grose, C.; Hambleton, S.; Kennedy, P.G.E.; Oxman, M.N.; et al. Varicella Zoster Virus Infection. Nat. Rev. Dis. Prim. 2015, 1, 15016. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, P.G.E.; Gershon, A.A. Clinical Features of Varicella-Zoster Virus Infection. Viruses 2018, 10, E609. [Google Scholar] [CrossRef]
- Azab, W.; Dayaram, A.; Greenwood, A.D.; Osterrieder, N. How Host Specific Are Herpesviruses? Lessons from Herpesviruses Infecting Wild and Endangered Mammals. Annu. Rev. Virol 2018, 5, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Lum, K.K.; Cristea, I.M. Host Innate Immune Response and Viral Immune Evasion During Alphaherpesvirus Infection. Curr. Issues Mol. Biol. 2021, 42, 635–686. [Google Scholar] [CrossRef]
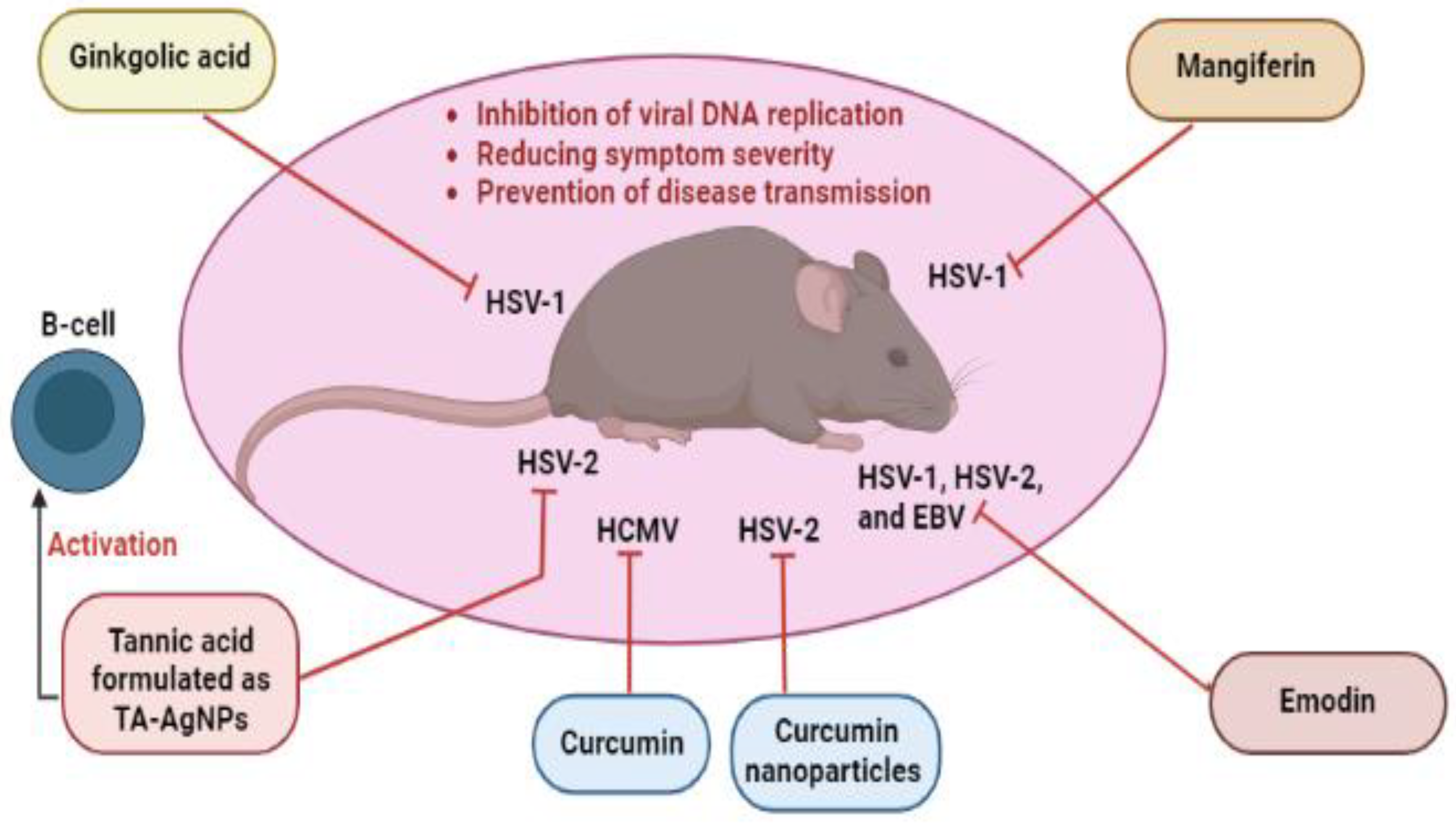
- Borenstein, R.; Hanson, B.A.; Markosyan, R.M.; Gallo, E.S.; Narasipura, S.D.; Bhutta, M.; Shechter, O.; Lurain, N.S.; Cohen, F.S.; Al-Harthi, L.; et al. Ginkgolic Acid Inhibits Fusion of Enveloped Viruses. Sci. Rep. 2020, 10, 4746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sochocka, M.; Sobczyński, M.; Ochnik, M.; Zwolińska, K.; Leszek, J. Hampering Herpesviruses HHV-1 and HHV-2 Infection by Extract of Ginkgo Biloba (EGb) and Its Phytochemical Constituents. Front. Microbiol. 2019, 10, 2367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhutta, M.S.; Shechter, O.; Gallo, E.S.; Martin, S.D.; Jones, E.; Doncel, G.F.; Borenstein, R. Ginkgolic Acid Inhibits Herpes Simplex Virus Type 1 Skin Infection and Prevents Zosteriform Spread in Mice. Viruses 2021, 13, 86. [Google Scholar] [CrossRef] [PubMed]
- Todorova, N.; Rangelov, M.; Dincheva, I.; Badjakov, I.; Enchev, V.; Markova, N. Potential of Hydroxybenzoic Acids from Graptopetalum Paraguayense for Inhibiting of Herpes Simplex Virus DNA Polymerase–Metabolome Profiling, Molecular Docking and Quantum-Chemical Analysis. Pharmacia 2022, 69, 113–123. [Google Scholar] [CrossRef]
- AbouAitah, K.; Allayh, A.K.; Wojnarowicz, J.; Shaker, Y.M.; Swiderska-Sroda, A.; Lojkowski, W. Nanoformulation Composed of Ellagic Acid and Functionalized Zinc Oxide Nanoparticles Inactivates DNA and RNA Viruses. Pharmaceutics 2021, 13, 2174. [Google Scholar] [CrossRef]
- Di Sotto, A.; Di Giacomo, S.; Amatore, D.; Locatelli, M.; Vitalone, A.; Toniolo, C.; Rotino, G.L.; Lo Scalzo, R.; Palamara, A.T.; Marcocci, M.E.; et al. A Polyphenol Rich Extract from Solanum Melongena L. DR2 Peel Exhibits Antioxidant Properties and Anti-Herpes Simplex Virus Type 1 Activity In Vitro. Molecules 2018, 23, E2066. [Google Scholar] [CrossRef] [Green Version]
- Langland, J.; Jacobs, B.; Wagner, C.E.; Ruiz, G.; Cahill, T.M. Antiviral Activity of Metal Chelates of Caffeic Acid and Similar Compounds towards Herpes Simplex, VSV-Ebola Pseudotyped and Vaccinia Viruses. Antivir. Res. 2018, 160, 143–150. [Google Scholar] [CrossRef]
- Kesharwani, A.; Polachira, S.K.; Nair, R.; Agarwal, A.; Mishra, N.N.; Gupta, S.K. Anti-HSV-2 Activity of Terminalia Chebula Retz Extract and Its Constituents, Chebulagic and Chebulinic Acids. BMC Complement. Altern. Med. 2017, 17, 110. [Google Scholar] [CrossRef]
- Siqueira, E.M.D.S.; Lima, T.L.; Boff, L.; Lima, S.G.; Lourenço, E.M.; Ferreira, É.G.; Barbosa, E.G.; Machado, P.R.; Farias, K.J.; Ferreira, L.D.S.; et al. Antiviral Potential of Spondias Mombin L. Leaves Extract Against Herpes Simplex Virus Type-1 Replication Using In Vitro and In Silico Approaches. Planta Med. 2020, 86, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Szymańska, E.; Orłowski, P.; Winnicka, K.; Tomaszewska, E.; Bąska, P.; Celichowski, G.; Grobelny, J.; Basa, A.; Krzyżowska, M. Multifunctional Tannic Acid/Silver Nanoparticle-Based Mucoadhesive Hydrogel for Improved Local Treatment of HSV Infection: In Vitro and In Vivo Studies. IJMS 2018, 19, 387. [Google Scholar] [CrossRef] [Green Version]
- Orłowski, P.; Kowalczyk, A.; Tomaszewska, E.; Ranoszek-Soliwoda, K.; Węgrzyn, A.; Grzesiak, J.; Celichowski, G.; Grobelny, J.; Eriksson, K.; Krzyzowska, M. Antiviral Activity of Tannic Acid Modified Silver Nanoparticles: Potential to Activate Immune Response in Herpes Genitalis. Viruses 2018, 10, 524. [Google Scholar] [CrossRef] [Green Version]
- Vilhelmova-Ilieva, N.; Jacquet, R.; Deffieux, D.; Pouységu, L.; Sylla, T.; Chassaing, S.; Nikolova, I.; Quideau, S.; Galabov, A.S. Anti-Herpes Simplex Virus Type 1 Activity of Specially Selected Groups of Tannins. Drug Res. 2019, 69, 373–374. [Google Scholar] [CrossRef] [PubMed]
- Vilhelmova-Ilieva, N.; Jacquet, R.; Quideau, S.; Galabov, A.S. Ellagitannins as Synergists of ACV on the Replication of ACV-Resistant Strains of HSV 1 and 2. Antivir. Res. 2014, 110, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Arunkumar, J.; Rajarajan, S. Study on Antiviral Activities, Drug-Likeness and Molecular Docking of Bioactive Compounds of Punica Granatum L. to Herpes Simplex Virus-2 (HSV-2). Microb. Pathog. 2018, 118, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Houston, D.M.J.; Bugert, J.J.; Denyer, S.P.; Heard, C.M. Potentiated Virucidal Activity of Pomegranate Rind Extract (PRE) and Punicalagin against Herpes Simplex Virus (HSV) When Co-Administered with Zinc (II) Ions, and Antiviral Activity of PRE against HSV and Aciclovir-Resistant HSV. PLoS ONE 2017, 12, e0179291. [Google Scholar] [CrossRef] [Green Version]
- Bae, S.; Kim, S.Y.; Do, M.H.; Lee, C.H.; Song, Y.-J. 1,2,3,4,6-Penta-O-Galloyl-ß-D-Glucose, a Bioactive Compound in Elaeocarpus Sylvestris Extract, Inhibits Varicella-Zoster Virus Replication. Antivir. Res. 2017, 144, 266–272. [Google Scholar] [CrossRef]
- Rechenchoski, D.Z.; Agostinho, K.F.; Faccin-Galhardi, L.C.; Lonni, A.A.S.G.; da Silva, J.V.H.; de Andrade, F.G.; Cunha, A.P.; Ricardo, N.M.P.S.; Nozawa, C.; Linhares, R.E.C. Mangiferin: A Promising Natural Xanthone from Mangifera Indica for the Control of Acyclovir - Resistant Herpes Simplex Virus 1 Infection. Bioorg. Med. Chem. 2020, 28, 115304. [Google Scholar] [CrossRef]
- Rechenchoski, D.Z.; Samensari, N.L.; Faccin-Galhardi, L.C.; de Almeida, R.R.; Cunha, A.P.; Ricardo, N.M.P.S.; Nozawa, C.; Linhares, R.E.C. The Combination of Dimorphandra Gardneriana Galactomannan and Mangiferin Inhibits Herpes Simplex and Poliovirus. Curr. Pharm. Biotechnol. 2019, 20, 215–221. [Google Scholar] [CrossRef]
- Abba, Y.; Hassim, H.; Hamzah, H.; Noordin, M.M. Antiviral Activity of Resveratrol against Human and Animal Viruses. Adv. Virol. 2015, 2015, 184241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Song, X.; Zhao, X.; Zhang, Y.; Wang, Y.; Jia, R.; Zou, Y.; Li, L.; Yin, Z. Insights into the Anti-Inflammatory and Antiviral Mechanisms of Resveratrol. Mediat. Inflamm. 2022, 2022, 7138756. [Google Scholar] [CrossRef] [PubMed]
- Docherty, J.J.; Fu, M.M.; Stiffler, B.S.; Limperos, R.J.; Pokabla, C.M.; DeLucia, A.L. Resveratrol Inhibition of Herpes Simplex Virus Replication. Antivir. Res. 1999, 43, 145–155. [Google Scholar] [CrossRef]
- Annunziata, G.; Maisto, M.; Schisano, C.; Ciampaglia, R.; Narciso, V.; Tenore, G.C.; Novellino, E. Resveratrol as a Novel Anti-Herpes Simplex Virus Nutraceutical Agent: An Overview. Viruses 2018, 10, 473. [Google Scholar] [CrossRef] [Green Version]
- Ding, L.; Jiang, P.; Xu, X.; Lu, W.; Yang, C.; Zhou, P.; Liu, S. Resveratrol Promotes HSV-2 Replication by Increasing Histone Acetylation and Activating NF-ΚB. Biochem. Pharmacol. 2020, 171, 113691. [Google Scholar] [CrossRef]
- Xiao, J.; Wang, X.; Wu, Y.; Zhao, Q.; Liu, X.; Zhang, G.; Zhao, Z.; Ning, Y.; Wang, K.; Tan, Y.; et al. Synergistic Effect of Resveratrol and HSV-TK/GCV Therapy on Murine Hepatoma Cells. Cancer Biol. Ther. 2019, 20, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Yang, X.-W.; Schols, D.; Mori, M.; Botta, B.; Chevigné, A.; Mulinge, M.; Steinmetz, A.; Schmit, J.-C.; Seguin-Devaux, C. Active Components from Cassia Abbreviata Prevent HIV-1 Entry by Distinct Mechanisms of Action. Int. J. Mol. Sci. 2021, 22, 5052. [Google Scholar] [CrossRef]
- Tarbeeva, D.V.; Krylova, N.V.; Iunikhina, O.V.; Likhatskaya, G.N.; Kalinovskiy, A.I.; Grigorchuk, V.P.; Shchelkanov, M.Y.; Fedoreyev, S.A. Biologically Active Polyphenolic Compounds from Lespedeza Bicolor. Fitoterapia 2022, 157, 105121. [Google Scholar] [CrossRef]
- Squillaci, G.; Zannella, C.; Carbone, V.; Minasi, P.; Folliero, V.; Stelitano, D.; Cara, F.L.; Galdiero, M.; Franci, G.; Morana, A. Grape Canes from Typical Cultivars of Campania (Southern Italy) as a Source of High-Value Bioactive Compounds: Phenolic Profile, Antioxidant and Antimicrobial Activities. Molecules 2021, 26, 2746. [Google Scholar] [CrossRef]
- Liu, S.; Li, L.; Tan, L.; Liang, X. Inhibition of Herpes Simplex Virus-1 Replication by Natural Compound Honokiol. Virol. Sin. 2019, 34, 315–323. [Google Scholar] [CrossRef]
- Dias, M.M.; Zuza, O.; Riani, L.R.; de Faria Pinto, P.; Pinto, P.L.S.; Silva, M.P.; de Moraes, J.; Ataíde, A.C.Z.; de Oliveira Silva, F.; Cecílio, A.B.; et al. In Vitro Schistosomicidal and Antiviral Activities of Arctium Lappa L. (Asteraceae) against Schistosoma Mansoni and Herpes Simplex Virus-1. Biomed. Pharmacother. 2017, 94, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Saidu, M.B.; Kúsz, N.; Tsai, Y.-C.; Vágvölgyi, M.; Berkecz, R.; Kókai, D.; Burián, K.; Hohmann, J.; Rédei, D. Triterpenes and Phenolic Compounds from Euphorbia Deightonii with Antiviral Activity against Herpes Simplex Virus Type-2. Plants 2022, 11, 764. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.-R.; Luo, J.; Hou, W.; Xiao, H.; Yang, Z.-Q. The Effect of Emodin, an Anthraquinone Derivative Extracted from the Roots of Rheum Tanguticum, against Herpes Simplex Virus in Vitro and in Vivo. J. Ethnopharmacol. 2011, 133, 718–723. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, X.; Pan, C.; Cheng, W.; Wang, X.; Yang, Z.; Zheng, L. The Intervention Mechanism of Emodin on TLR3 Pathway in the Process of Central Nervous System Injury Caused by Herpes Virus Infection. Neurol. Res. 2021, 43, 307–313. [Google Scholar] [CrossRef]
- Mugas, M.L.; Marioni, J.; Martinez, F.; Aguilar, J.J.; Cabrera, J.L.; Contigiani, M.S.; Konigheim, B.S.; Núñez-Montoya, S.C. Inactivation of Herpes Simplex Virus by Photosensitizing Anthraquinones Isolated from Heterophyllaea Pustulata. Planta Med. 2021, 87, 716–723. [Google Scholar] [CrossRef]
- Roa-Linares, V.C.; Miranda-Brand, Y.; Tangarife-Castaño, V.; Ochoa, R.; García, P.A.; Castro, M.Á.; Betancur-Galvis, L.; San Feliciano, A. Anti-Herpetic, Anti-Dengue and Antineoplastic Activities of Simple and Heterocycle-Fused Derivatives of Terpenyl-1,4-Naphthoquinone and 1,4-Anthraquinone. Molecules 2019, 24, 1279. [Google Scholar] [CrossRef] [Green Version]
- Soleimani, V.; Sahebkar, A.; Hosseinzadeh, H. Turmeric (Curcuma Longa) and Its Major Constituent (Curcumin) as Nontoxic and Safe Substances: Review. Phytother. Res. 2018, 32, 985–995. [Google Scholar] [CrossRef]
- Kotha, R.R.; Luthria, D.L. Curcumin: Biological, Pharmaceutical, Nutraceutical, and Analytical Aspects. Molecules 2019, 24, 2930. [Google Scholar] [CrossRef] [Green Version]
- Flores, D.J.; Lee, L.H.; Adams, S.D. Inhibition of Curcumin-Treated Herpes Simplex Virus 1 and 2 in Vero Cells. AiM 2016, 6, 276–287. [Google Scholar] [CrossRef] [Green Version]
- Kutluay, S.B.; Doroghazi, J.; Roemer, M.E.; Triezenberg, S.J. Curcumin Inhibits Herpes Simplex Virus Immediate-Early Gene Expression by a Mechanism Independent of P300/CBP Histone Acetyltransferase Activity. Virology 2008, 373, 239–247. [Google Scholar] [CrossRef]
- Vitali, D.; Bagri, P.; Wessels, J.M.; Arora, M.; Ganugula, R.; Parikh, A.; Mandur, T.; Felker, A.; Garg, S.; Kumar, M.N.V.R.; et al. Curcumin Can Decrease Tissue Inflammation and the Severity of HSV-2 Infection in the Female Reproductive Mucosa. IJMS 2020, 21, 337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Wu, L.; Wang, M.; Cheng, A.; Yang, Q.; Wu, Y.; Jia, R.; Zhu, D.; Zhao, X.; Chen, S.; et al. Alpha-Herpesvirus Thymidine Kinase Genes Mediate Viral Virulence and Are Potential Therapeutic Targets. Front. Microbiol. 2019, 10, 941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Halim, S.M.A.; Mamdouh, M.A.; El-Haddad, A.E.; Soliman, S.M. Fabrication of Anti-HSV-1 Curcumin Stabilized Nanostructured Proniosomal Gel: Molecular Docking Studies on Thymidine Kinase Proteins. Sci. Pharm. 2020, 88, 9. [Google Scholar] [CrossRef] [Green Version]
- Badria, F.A.; Abdelaziz, A.E.; Hassan, A.H.; Elgazar, A.A.; Mazyed, E.A. Development of Provesicular Nanodelivery System of Curcumin as a Safe and Effective Antiviral Agent: Statistical Optimization, In Vitro Characterization, and Antiviral Effectiveness. Molecules 2020, 25, 5668. [Google Scholar] [CrossRef]
- Rajtar, B.; Skalicka-Woźniak, K.; Świątek, Ł.; Stec, A.; Boguszewska, A.; Polz-Dacewicz, M. Antiviral Effect of Compounds Derived from Angelica Archangelica L. on Herpes Simplex Virus-1 and Coxsackievirus B3 Infections. Food Chem. Toxicol. 2017, 109, 1026–1031. [Google Scholar] [CrossRef]
- Okba, M.M.; El Gedaily, R.A.; Ashour, R.M. UPLC-PDA-ESI-QTOF-MS Profiling and Potent Anti-HSV-II Activity of Eucalyptus Sideroxylon Leaves. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2017, 1068–1069, 335–342. [Google Scholar] [CrossRef]
- Fulkerson, H.L.; Nogalski, M.T.; Collins-McMillen, D.; Yurochko, A.D. Overview of Human Cytomegalovirus Pathogenesis. Methods Mol. Biol. 2021, 2244, 1–18. [Google Scholar] [CrossRef]
- O’Connor, C.M. Cytomegalovirus (CMV) Infection and Latency. Pathogens 2021, 10, 342. [Google Scholar] [CrossRef]
- Griffiths, P.; Baraniak, I.; Reeves, M. The Pathogenesis of Human Cytomegalovirus. J. Pathol. 2015, 235, 288–297. [Google Scholar] [CrossRef]
- Michaelis, M.; Doerr, H.W.; Cinatl, J. The Story of Human Cytomegalovirus and Cancer: Increasing Evidence and Open Questions. Neoplasia 2009, 11, 1–9. [Google Scholar] [CrossRef]
- Golais, F.; Mrázová, V. Human Alpha and Beta Herpesviruses and Cancer: Passengers or Foes? Folia Microbiol. 2020, 65, 439–449. [Google Scholar] [CrossRef]
- Griffiths, P. Cytomegalovirus Infection of the Central Nervous System. Herpes 2004, 11 (Suppl. 2), 95A–104A. [Google Scholar] [PubMed]
- Tselis, A.C. Cytomegalovirus Infections of the Adult Human Nervous System. Handb. Clin. Neurol. 2014, 123, 307–318. [Google Scholar] [CrossRef]
- Zhang, X.-Y.; Fang, F. Congenital Human Cytomegalovirus Infection and Neurologic Diseases in Newborns. Chin. Med. J. (Engl) 2019, 132, 2109–2118. [Google Scholar] [CrossRef]
- Wang, S.-Y.; Zhang, J.; Xu, X.-G.; Su, H.-L.; Xing, W.-M.; Zhang, Z.-S.; Jin, W.-H.; Dai, J.-H.; Wang, Y.-Z.; He, X.-Y.; et al. Inhibitory Effects of Piceatannol on Human Cytomegalovirus (HCMV) in Vitro. J. Microbiol. 2020, 58, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Alam, Z.; Al-Mahdi, Z.; Zhu, Y.; McKee, Z.; Parris, D.S.; Parikh, H.I.; Kellogg, G.E.; Kuchta, A.; McVoy, M.A. Anti-Cytomegalovirus Activity of the Anthraquinone Atanyl Blue PRL. Antivir. Res. 2015, 114, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; An, Z.; Chen, H.; Wang, Z.; Liu, L. Mechanism of Curcumin Resistance to Human Cytomegalovirus in HELF Cells. BMC Complement. Altern. Med. 2014, 14, 284. [Google Scholar] [CrossRef] [Green Version]
- Lv, Y.; Gong, L.; Wang, Z.; Han, F.; Liu, H.; Lu, X.; Liu, L. Curcumin Inhibits Human Cytomegalovirus by Downregulating Heat Shock Protein 90. Mol. Med. Rep. 2015, 12, 4789–4793. [Google Scholar] [CrossRef] [Green Version]
- Lv, Y.-L.; Jia, Y.; Wan, Z.; An, Z.-L.; Yang, S.; Han, F.-F.; Gong, L.-L.; Xuan, L.-L.; Ren, L.-L.; Zhang, W.; et al. Curcumin Inhibits the Formation of Atherosclerosis in ApoE-/- Mice by Suppressing Cytomegalovirus Activity in Endothelial Cells. Life Sci. 2020, 257, 117658. [Google Scholar] [CrossRef]
- Lv, Y.; Lei, N.; Wang, D.; An, Z.; Li, G.; Han, F.; Liu, H.; Liu, L. Protective Effect of Curcumin against Cytomegalovirus Infection in Balb/c Mice. Environ. Toxicol. Pharmacol. 2014, 37, 1140–1147. [Google Scholar] [CrossRef]
- Möhl, B.S.; Chen, J.; Longnecker, R. Gammaherpesvirus Entry and Fusion: A Tale How Two Human Pathogenic Viruses Enter Their Host Cells. Adv. Virus Res. 2019, 104, 313–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrell, P.J. Epstein-Barr Virus and Cancer. Annu. Rev. Pathol. 2019, 14, 29–53. [Google Scholar] [CrossRef] [PubMed]
- Yiu, S.P.T.; Dorothea, M.; Hui, K.F.; Chiang, A.K.S. Lytic Induction Therapy against Epstein-Barr Virus-Associated Malignancies: Past, Present, and Future. Cancers 2020, 12, 2142. [Google Scholar] [CrossRef]
- Wen, K.W.; Wang, L.; Menke, J.R.; Damania, B. Cancers Associated with Human Gammaherpesviruses. FEBS J. 2021. [Google Scholar] [CrossRef]
- Goncalves, P.H.; Ziegelbauer, J.; Uldrick, T.S.; Yarchoan, R. Kaposi Sarcoma Herpesvirus-Associated Cancers and Related Diseases. Curr. Opin. HIV AIDS 2017, 12, 47–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ackermann, M. Pathogenesis of Gammaherpesvirus Infections. Vet. Microbiol. 2006, 113, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Soldan, S.S.; Lieberman, P.M. Epstein-Barr Virus Infection in the Development of Neurological Disorders. Drug Discov. Today Dis. Models 2020, 32, 35–52. [Google Scholar] [CrossRef]
- Jha, H.C.; Mehta, D.; Lu, J.; El-Naccache, D.; Shukla, S.K.; Kovacsics, C.; Kolson, D.; Robertson, E.S. Gammaherpesvirus Infection of Human Neuronal Cells. mBio 2015, 6, e01844-15. [Google Scholar] [CrossRef] [Green Version]
- Nowalk, A.; Green, M. Epstein-Barr Virus. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Ciccarese, G.; Trave, I.; Herzum, A.; Parodi, A.; Drago, F. Dermatological Manifestations of Epstein-Barr Virus Systemic Infection: A Case Report and Literature Review. Int. J. Dermatol. 2020, 59, 1202–1209. [Google Scholar] [CrossRef]
- Li, S.; Bai, L.; Dong, J.; Sun, R.; Lan, K. Kaposi’s Sarcoma-Associated Herpesvirus: Epidemiology and Molecular Biology. In Infectious Agents Associated Cancers: Epidemiology and Molecular Biology; Cai, Q., Yuan, Z., Lan, K., Eds.; Advances in Experimental Medicine and Biology; Springer: Singapore, 2017; Volume 1018, pp. 91–127. ISBN 978-981-10-5764-9. [Google Scholar]
- Nomura, E.; Hosoda, A.; Morishita, H.; Murakami, A.; Koshimizu, K.; Ohigashi, H.; Taniguchi, H. Synthesis of Novel Polyphenols Consisted of Ferulic and Gallic Acids, and Their Inhibitory Effects on Phorbol Ester-Induced Epstein-Barr Virus Activation and Superoxide Generation. Bioorg. Med. Chem. 2002, 10, 1069–1075. [Google Scholar] [CrossRef]
- Crowley, A.; Connell, J.; Schaffer, K.; Hall, W.; Hassan, J. Is There Diagnostic Value in Detection of Immunoglobulin g Antibodies to the Epstein-Barr Virus Early Antigen? Biores. Open Access 2012, 1, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Boonsopon, S.; Maghsoudlou, A.; Kombo, N.E.; Foster, C.S. A Therapeutic Trial of Valganciclovir in Patients with Uveitis and Positive Epstein-Barr Virus Early Antigen D IgG Titers. Eur. J. Ophthalmol. 2016, 26, 30–35. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, W.-F.; Xu, J.; Kitdamrongtham, W.; Manosroi, A.; Manosroi, J.; Tokuda, H.; Abe, M.; Akihisa, T.; Feng, F. Potential Cancer Chemopreventive and Anticancer Constituents from the Fruits of Ficus Hispida L.f. (Moraceae). J. Ethnopharmacol. 2018, 214, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.-Y.; Chen, C.-Y.; Shyu, H.-W.; Hong, S.; Chen, H.-M.; Chiou, Y.-H.; Lin, K.-H.; Chou, M.-C.; Wang, L.-Y.; Wang, Y.-F. Resveratrol Induces Cell Death and Inhibits Human Herpesvirus 8 Replication in Primary Effusion Lymphoma Cells. Chem. Biol. Interact. 2015, 242, 372–379. [Google Scholar] [CrossRef]
- Wang, Q.; Zhu, N.; Hu, J.; Wang, Y.; Xu, J.; Gu, Q.; Lieberman, P.M.; Yuan, Y. The MTOR Inhibitor Manassantin B Reveals a Crucial Role of MTORC2 Signaling in Epstein-Barr Virus Reactivation. J. Biol. Chem. 2020, 295, 7431–7441. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.-C.; Chen, M.-S.; Cheng, Y.-J.; Ko, Y.-C.; Lin, S.-F.; Chiu, I.-M.; Chen, J.-Y. Emodin Inhibits EBV Reactivation and Represses NPC Tumorigenesis. Cancers 2019, 11, 1795. [Google Scholar] [CrossRef] [Green Version]
- Dheekollu, J.; Wiedmer, A.; Ayyanathan, K.; Deakyne, J.S.; Messick, T.E.; Lieberman, P.M. Cell-Cycle-Dependent EBNA1-DNA Crosslinking Promotes Replication Termination at OriP and Viral Episome Maintenance. Cell 2021, 184, 643–654.e13. [Google Scholar] [CrossRef]
- Jakhmola, S.; Jonniya, N.A.; Sk, M.F.; Rani, A.; Kar, P.; Jha, H.C. Identification of Potential Inhibitors against Epstein–Barr Virus Nuclear Antigen 1 (EBNA1): An Insight from Docking and Molecular Dynamic Simulations. ACS Chem. Neurosci. 2021, 12, 3060–3072. [Google Scholar] [CrossRef]
- Yiu, C.-Y.; Chiu, Y.-J.; Lin, T.-P. The Ethyl Acetate Subfraction of Polygonum Cuspidatum Root Containing Emodin Affect EBV Gene Expression and Induce EBV-Positive Cells Apoptosis. Biol. Pharm. Bull. 2021, 44, 1837–1842. [Google Scholar] [CrossRef]
- Tsai, Y.-C.; Hohmann, J.; El-Shazly, M.; Chang, L.-K.; Dankó, B.; Kúsz, N.; Hsieh, C.-T.; Hunyadi, A.; Chang, F.-R. Bioactive Constituents of Lindernia Crustacea and Its Anti-EBV Effect via Rta Expression Inhibition in the Viral Lytic Cycle. J. Ethnopharmacol. 2020, 250, 112493. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yang, J.; Ji, W.; Wang, C. Curcumin Inhibits Proliferation of Epstein-Barr Virus-Associated Human Nasopharyngeal Carcinoma Cells by Inhibiting EBV Nuclear Antigen 1 Expression. Biomed. Res. Int. 2019, 2019, 8592921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Castillo, M.; Cruz-Robledo, G.; Hernández-Zavala, A.; Córdova, E.J. Curcumin Sensitizes Epstein-Barr-Immortalized Lymphoblastoid Cell Lines to Inorganic Arsenic Toxicity. Exp. Ther. Med. 2021, 22, 872. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhong, C.; Wang, Q.; Chen, W.; Yuan, Y. Curcumin Is an APE1 Redox Inhibitor and Exhibits an Antiviral Activity against KSHV Replication and Pathogenesis. Antivir. Res. 2019, 167, 98–103. [Google Scholar] [CrossRef]
- Wu, T.; Wang, Y.; Yuan, Y. Antiviral Activity of Topoisomerase II Catalytic Inhibitors against Epstein-Barr Virus. Antivir. Res. 2014, 107, 95–101. [Google Scholar] [CrossRef]
- Lin, Y.; Wang, Q.; Gu, Q.; Zhang, H.; Jiang, C.; Hu, J.; Wang, Y.; Yan, Y.; Xu, J. Semisynthesis of (-)-Rutamarin Derivatives and Their Inhibitory Activity on Epstein-Barr Virus Lytic Replication. J. Nat. Prod. 2017, 80, 53–60. [Google Scholar] [CrossRef]
- Xu, B.; Wang, L.; González-Molleda, L.; Wang, Y.; Xu, J.; Yuan, Y. Antiviral Activity of (+)-Rutamarin against Kaposi’s Sarcoma-Associated Herpesvirus by Inhibition of the Catalytic Activity of Human Topoisomerase II. Antimicrob. Agents Chemother. 2014, 58, 563–573. [Google Scholar] [CrossRef] [Green Version]
- Coen, D.M.; Lawler, J.L.; Abraham, J. Herpesvirus DNA Polymerase: Structures, Functions, and Mechanisms. Enzymes 2021, 50, 133–178. [Google Scholar] [CrossRef]
- Piret, J.; Boivin, G. Antiviral Drugs Against Herpesviruses. Adv. Exp. Med. Biol. 2021, 1322, 1–30. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Chemical Class | Herpesvirus | Mechanisms of Action (Inhibition/Downregulation) |
|---|---|---|---|
| Ginkgolic acid | Phenolic acids | HSV-1, HCMV, and EBV | HSV-1 DNA replication, viral structure, ICP27, ICP8, US11, and viral progeny production. HCMV entry and its DNA replication. EBV membrane fusion and gB. |
| Trans-ferulic acid, gentisic acid, vanillic acid, syringic acid, and gallic acid | Phenolic acids | HSV-1 and EBV | HSV-1 DNA polymerase, HSV-1 gB (by vanillic acid), and EBV-EA (by gallic acid). |
| Polyphenol esters consisting of gallic acid and ferulic acid | Phenolic acids | EBV | EBV reactivation. |
| Ellagic acid | Phenolic acids | HSV-2 | HSV-2 DNA replication. |
| Chlorogenic acid and caffeic acid | Phenolic acids | HSV-1 and EBV | HSV-1 gB and EBV-EA (by chlorogenic acid). |
| Caffeic acid chelates | Phenolic acids | HSV-1 and HSV-2 | Enhancement of anti-HSV activity by inhibiting viral DNA replication and viral attachment. |
| Protocatechuic acid | Phenolic acids | HSV-2 and EBV | HSV-2 DNA replication and virion production. EBV-EA. |
| Chebulagic acid and chebulinic acid | Tannins | HSV-2 | HSV-2 DNA replication. |
| Geraniin | Tannins | HSV-1 | HSV-1 gB. |
| Tannic acid formulated as TA-AgNPs and purified tannic acid | Tannins (gallotannins) | HSV-1 and HSV-2 | HSV-1 replication, gC, and gB (purified and formulated tannic acid), HSV-2 replication and improving the anti-HSV-2 immune response by activating B cells. |
| 1,2,3,4,5-penta-O-digalloyl-β-D-glucopyranose, 1,2,3,4,5-penta-O-digalloyl-α-D-glucopyranose, and α/β-3-O-digalloyl-D-glucopyranose (1:1 mixture). | Tannins (gallotannins) | HSV-1 | HSV-1 replication and viral glycoproteins. |
| Pentagalloylglucose | Tannins (gallotannins) | VZV | VZV replication, VZV-induced JNK, and VZV-IE62. |
| Castalagin and vescalagin | Tannins (ellagitannins) | HSV-1 and HSV-2 | In combination with acyclovir, notable inhibition of HSV-1 and HSV-2 replications was observed. |
| Epiacutissimin B, epiacutissimin A, acutissimin A, and mongolicain | Tannins (ellagitannins) | HSV-1 | HSV-1 DNA replication and viral glycoproteins. |
| Punicalagin | Tannins (ellagitannins) | HSV-2 | HSV-2 DNA replication. HSV-2 protease. |
| Mangiferin | Xanthones | HSV-1 | HSV-1 DNA replication and virus particles. |
| Resveratrol | Stilbenes | HSV-1, HSV-2, and KSHV | HSV-1 and HSV-2 replications, viral IE, and CDK9. KSHV latent infection, Rta, and formation of virus progeny. |
| Greco extract contains resveratrol C-glucoside, resveratrol, and epsilon-viniferin | Stilbenes | HSV-1 | HSV-1 particles and viral DNA replication. |
| Piceatannol | Stilbenes | HSV-1, HSV-2, and HCMV | HSV-1 and HSV-2 replications and viral particles. HCMV replication, IE, E, and p16INK4a. |
| Bicoloketone | Stilbenes | HSV-1 | HSV-1 DNA replication. |
| Honokiol | Lignans | HSV-1 | HSV-1 DNA replication, ICP27, VP16, and gD. |
| Arctium lappa L. extract (rich in arctiin and arctigenin) | Lignans | HSV-1 | Viral load and HSV-1 DNA replication. |
| Manassantin B | Lignans | EBV | EBV lytic DNA replication, virion production, BZLF1, AP-1, and mTORC2-mediated phosphorylation of AKT Ser/Thr at Ser-473 and PKCα at Ser-657. |
| Deightonin | Neolignans | HSV-2 | HSV-2 DNA replication. |
| Emodin | Anthraquinones | HSV-1, HSV-2, HCMV, and EBV | HSV-1 and HSV-2 replications, TLR3 pathway and its downstream molecules (TRIF, TRADD, TRAF6, traf3, Nemo, IRF3, and p38), IL-6, TNF-α, and IFN-β. HCMV DNA replication and synthesis. EBV lytic proteins, virion production, SP1, Zta, Rta, EBNA1, BRLF1, BNLF1, and LMP1. |
| Aloe-emodin | Anthraquinones | EBV | EBV lytic cycle and Rta. |
| 5,5′-Bisoranjidiol, rubiadin 1-methyl ether, soranjidiol 1-methyl ether, damnacanthol, soranjidiol, rubiadin, and heterophylline | Anthraquinones | HSV-1 | HSV-1 DNA replication and HSV-1 particles (photo-inactivation). |
| 1,4-Anthraquinone | Anthraquinones | HSV-1 | HSV-1 DNA replication. |
| Curcumin | Curcuminoids | HSV-1, HSV-2, HCMV, EBV, and KSHV | HSV-1 and HSV-2 replications and their adsorption, HSV-1 TK, HSV-1 IE, p300, CBP and HSV-1 DNA polymerase. HCMV (IEA, UL83A, IL-6, TNF-α, Hsp90, ROS, inflammatory cytokines, HMGB1-TLRS-NF-κB). Protection against HCMV by inducing anti-inflammatory and antioxidant activities. EBV (latent and lytic replication, BZLF1, and EBNA1. KSHV (Rta, K8, and APE1-mediated redox function). |
| Imperatorin and phellopterin | Coumarins | HSV-1 | HSV-1 DNA replication. |
| Scoparon | Coumarins | HSV-2 | HSV-2 DNA replication. |
| 7-hydroxycoumarin and 7-hydroxy-6-[2-(R)-hydroxy-3-methyl-but-3-enyl]hydroxycoumarin | Coumarins | EBV | EBV-EA. |
| Psoralen | Furanocoumarins | EBV | EBV-EA. |
| (+)-Rutamarin | Furanocoumarins | EBV and KSHV | EBV (lytic DNA and viral protein synthesis). KSHV (lytic DNA replication and virion production). |
| Phloroglucinol-rich extract (PGRE) | Other polyphenols (phloroglucinol) | HSV-2 | HSV-2 DNA replication and viral protein synthesis. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassan, S.T.S.; Šudomová, M.; Mazurakova, A.; Kubatka, P. Insights into Antiviral Properties and Molecular Mechanisms of Non-Flavonoid Polyphenols against Human Herpesviruses. Int. J. Mol. Sci. 2022, 23, 13891. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232213891
Hassan STS, Šudomová M, Mazurakova A, Kubatka P. Insights into Antiviral Properties and Molecular Mechanisms of Non-Flavonoid Polyphenols against Human Herpesviruses. International Journal of Molecular Sciences. 2022; 23(22):13891. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232213891
Chicago/Turabian StyleHassan, Sherif T. S., Miroslava Šudomová, Alena Mazurakova, and Peter Kubatka. 2022. "Insights into Antiviral Properties and Molecular Mechanisms of Non-Flavonoid Polyphenols against Human Herpesviruses" International Journal of Molecular Sciences 23, no. 22: 13891. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232213891